
What is Known Regarding the Participation of Factor Nrf-2 in Liver Regeneration?

 , , , ,
, , , , {kind=link}
Abstract
:1. Liver
2. Liver Regeneration
3. Keap1-Nrf2-ARE Pathway
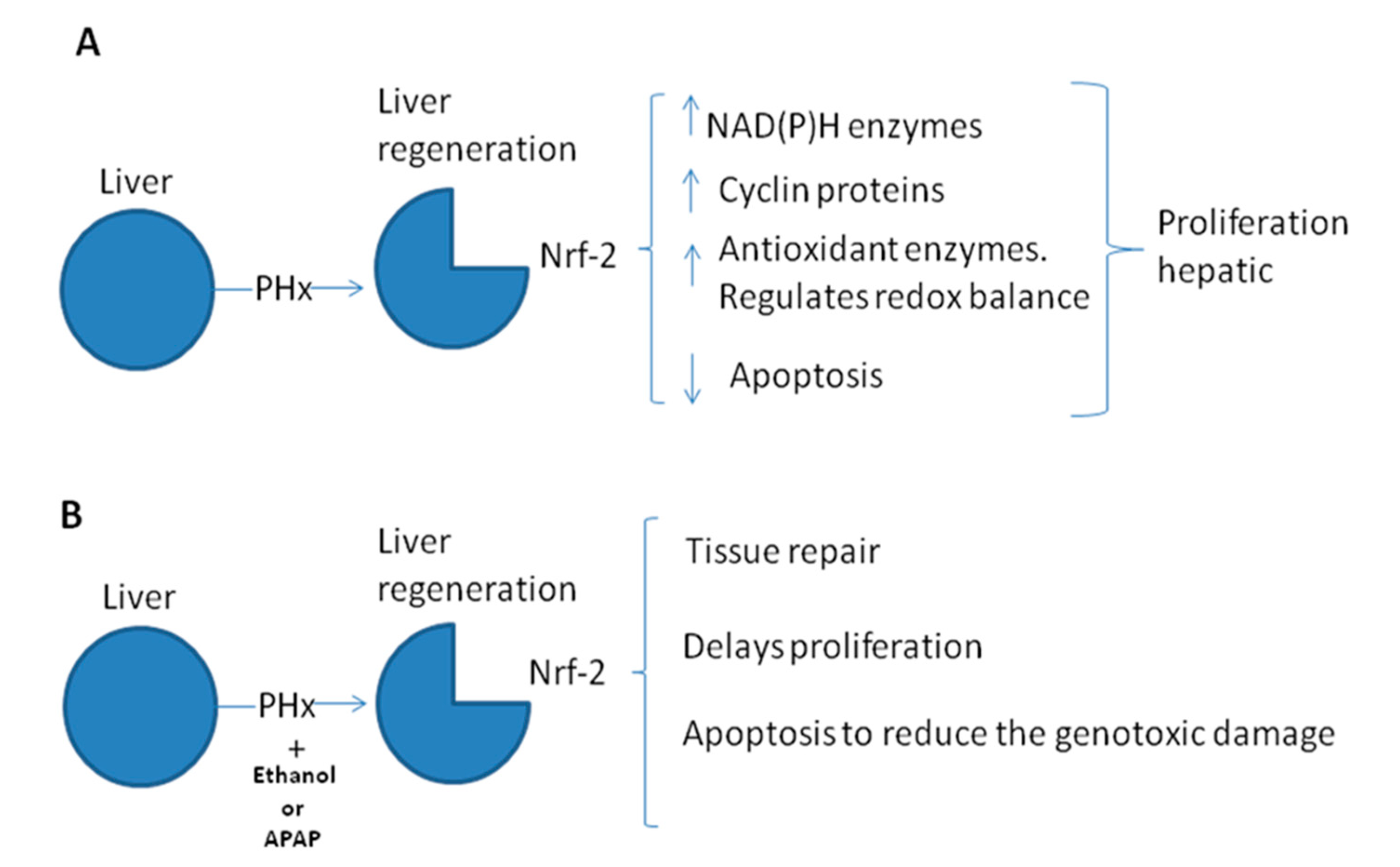
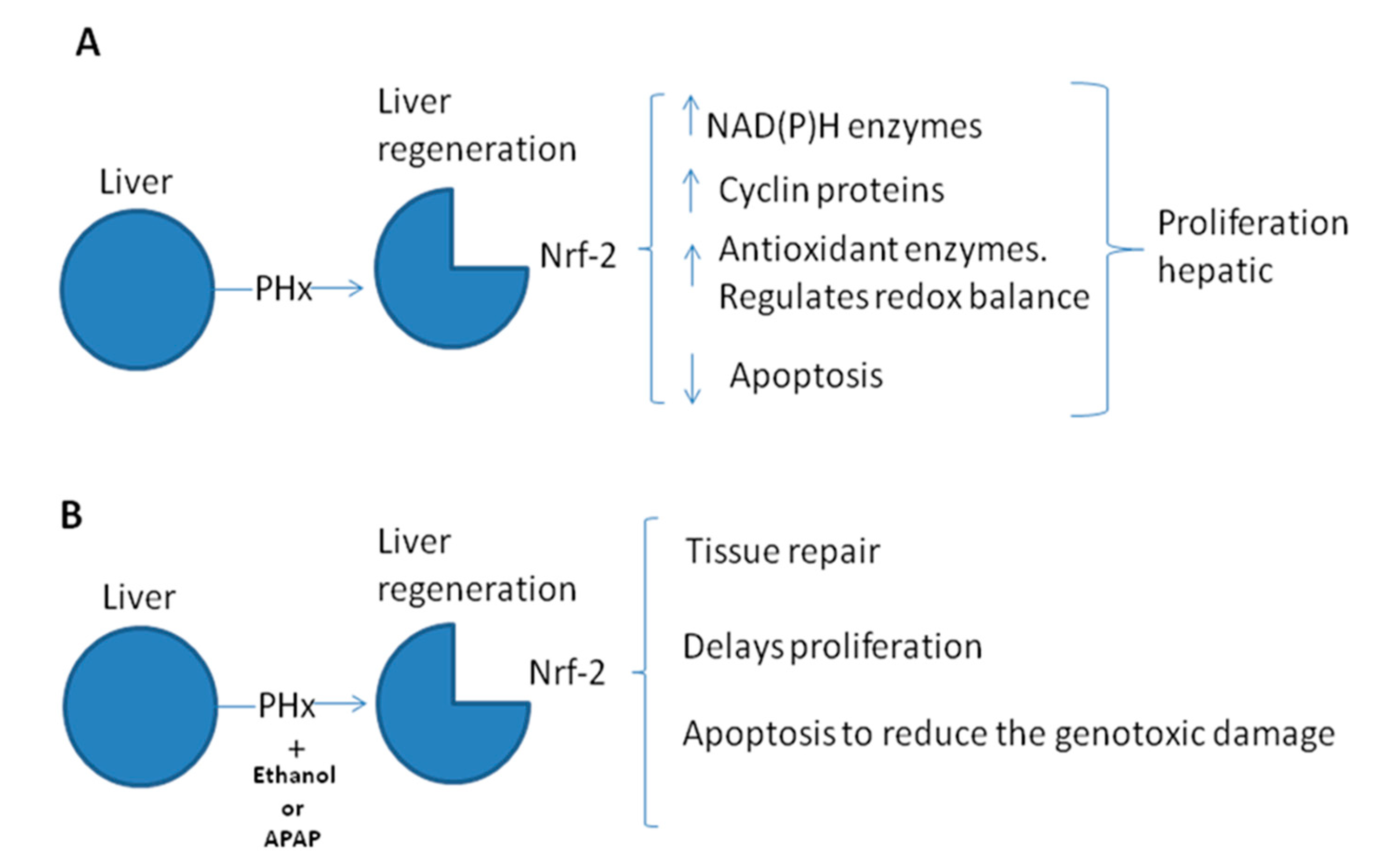
4. Nrf-2 and Liver Regeneration
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Morales González, J.A. Alcohol, Alcoholismo Y Cirrosis. Un Enfoque Multidisciplinario; Universidad Autónoma del Estado de Hidalgo: Pachuca, México, 2007; p. 240. [Google Scholar]
- Ahsan, M.R.; Islam, K.M.; Bulbul, I.J. Hepatoprotective activity of methanol extract of some medicinal plants against carbon tetrachloride-induced hepatotoxicity in rats. Glob. J. Pharmacol. 2009, 3, 116–122. [Google Scholar]
- Madrigal-Santillán, E.; Madrigal-Bujaidar, E.; Álvarez-González, I.; Sumaya-Martínez, M.T.; Gutiérrez-Salinas, J.; Bautista, M.; Morales-González, Á.; García-Luna y González-Rubio, M.; Aguilar-Faisal, J.L.; Morales-González, J.A. Review of natural products with hepatoprotective effects. World J. Gastroenterol. 2014, 20, 14787–14804. [Google Scholar] [CrossRef] [PubMed]
- Higgins, G.M.; Anderson, R.M. Experimental pathology of the liver. I. Restoration of the liver of the white rat following partial surgical removal. ARC Pathol. 1931, 12, 186–202. [Google Scholar]
- Michalopoulos, G.K. Liver regeneration: Molecular mechanism of growth control. FASEB J. 1990, 4, 176–186. [Google Scholar] [PubMed]
- Fausto, N.; Webber, E.M. Control of liver growth. Crit. Rev. Eukaryot. Gene Expr. 1993, 3, 117–135. [Google Scholar] [PubMed]
- Steiner, J.W.; Perz, Z.M.; Taichman, L.B. Cell population dynamics in the liver. A review of quantitative morphological techniques applied to the study of physiological and pathological growth. Exp. Mol. Pathol. 1966, 5, 146–181. [Google Scholar] [CrossRef] [PubMed]
- Fausto, N.; Campbell, J.S.; Riehle, K.J. Liver regeneration. Hepatology 2006, 43, S45–S53. [Google Scholar] [CrossRef] [PubMed]
- Fausto, N.; Campbell, J.S.; Riehle, K.J. Liver regeneration. J. Hepatol. 2012, 57, 692–694. [Google Scholar] [CrossRef] [PubMed]
- Thorgeirsson, S.S. Hepatic stem cells in liver regeneration. FASEB J. 1996, 10, 1249–1256. [Google Scholar] [PubMed]
- Gressner, A.M. Cytokines and cellular crosstalk involved in the activation of fat-storing cells. J. Hepatol. 1995, 22, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.I.; Mars, W.M.; Michalopoulos, G.K. Signals and cells involved in regulating liver regeneration. Cell 2012, 13, 1261–1292. [Google Scholar] [CrossRef]
- Francavilla, A.; Ove, P.; Polimeno, L.; Coetzee, M.; Makowka, L.; Barone, M.; van Thiel, D.H.; Starzl, T.E. Regulation of liver size and regeneration: Importance in liver transplantation. Transpl. Proc. 1988, 20, 494–497. [Google Scholar]
- Aguilar-Delfín, I.; López-Barrera, F.; Hernández-Muñoz, R. Selective enhancement of lipid peroxidation in plasma membrane in two experimental models of liver regeneration: Partial hepatectomy and acute administration. Hepatology 1996, 24, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Trejo-Solís, C.; Chagoya de Sánchez, V.; Aranda-Fraustro, A.; Sánchez-Sevilla, L.; Gómez-Ruíz, C.; Hernández-Muñoz, R. Inhibitory effect of vitamin E administration on the progression of liver regeneration induced by partial hepatectomy in rats. Lab. Investig. 2003, 83, 1669–1679. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Farías, C.; Madrigal-Santillán, E.; Gutiérrez-Salinas, J.; Rodríguez-Sánchez, N.; Martínez-Cruz, M.; Valle-Jones, I.; Gramlich-Martínez, I.; Hernández-Ceruelos, A.; Morales-Gonzaléz, J.A. Protective effect of some vitamins against the toxic action of ethanol on liver regeneration induced by partial hepatectomy in rats. World J. Gastroenterol. 2008, 14, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.-H.; Kang, K.-S.; Kwak, M.-K. Effect of redox modulating NRF2 activators on chronic kidney disease. Molecules 2014, 19, 12727–12759. [Google Scholar] [CrossRef] [PubMed]
- Stefanson, A.L.; Bakovic, M. Dietary regulation of Keap1/Nrf2/ARE pathway: Focus on plant-derived compounds and trace minerals. Nutrients 2014, 6, 3777–3801. [Google Scholar] [CrossRef]
- Keum, Y.S.; Choi, B.Y. Molecular and chemical regulation of the Keap1-Nrf2 signaling pathway. Molecules 2014, 19, 100074–100089. [Google Scholar] [CrossRef]
- Han, X.; Shen, T.; Lou, H. Dietary poliphenols and their biological significance. Int. J. Mol. Sci. 2007, 8, 950–988. [Google Scholar] [CrossRef]
- Zou, Y.; Hu, M.; Lee, J.; Nambiar, S.M.; Garcia, V.; Bao, Q.; Chan, J.Y.; Dai, G. Nrf2 is essential for timely M-phase entry of replicating hepatocytes during liver regeneration. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 18. [Google Scholar] [CrossRef]
- Zou, Y.; Lee, J.; Nambiar, S.M.; Hu, M.; Rui, W.; Bao, Q.; Chan, J.Y.; Dai, G. Nrf2 is involved in maintaining hepatocyte identity during liver regeneration. PLoS ONE 2014, 9, e107423. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Zou, Y.; Nambiar, S.M.; Lee, J.; Yang, Y.; Dai, G. Keap1 modulates the redox cycle and hepatocyte cell cycle in regenerating liver. Cell Cycle 2014, 13, 2349–2358. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Chen, P.; Tan, H.; Zeng, H.; Jiang, Y.; Wang, Y.; Wang, Y.; Hou, X.; Bi, H.; Huang, M. Dynamic and coordinated regulation of KEAP1-NRF2-ARE and p53/p21 signaling pathways is associated with acetaminophen injury responsive liver regeneration. Drug Metab. Dispos. 2014, 42, 1532–1539. [Google Scholar] [CrossRef] [PubMed]
- Dayoub, R.; Vogel, A.; Schuett, J.; Lupke, M.; Spieker, S.M.; Kettern, N.; Hildt, E.; Melter, M.; Weiss, T.S. Nrf2 activates augmenter of liver regeneration (ALR) via antioxidant response element and links oxidative stress to liver regeneration. Mol. Med. 2013, 19, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Köhler, U.A.; Kurinna, S.; Schwitter, D.; Marti, A.; Schäfer, M.; Hellerbrand, C.; Speicher, T.; Werner, S. Activated Nrf2 impairs liver regeneration in mice by activation of genes involved in cell-cycle control and apoptosis. Hepatology 2014, 60, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.M.; Yang, J.H.; Ki, S.H. Role of the Nrf2-ARE pathway in liver diseases. Oxid. Med. Cell. Longev. 2013. [Google Scholar] [CrossRef]
- Buitrago-Molina, L.E.; Marhenke, S.; Longerich, T.; Sharma, A.D.; Boukouris, A.E.; Geffers, R.; Guigas, B.; Manns, M.P.; Vogel, A. The degree of liver injury determines the role of p21 in liver regeneration and hepatocarcinogenesis in mice. Hepatology 2013, 58, 1143–1152. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, N.; Shin, S.; Slocum, S.L.; Agoston, E.S.; Wakabayashi, J.; Kwak, M.K.; Misra, V.; Biswal, S.; Yamamoto, M.; Kensler, T.W. Regulation of notch1 signaling by Nrf2: Implications for tissue regeneration. Sci. Signal. 2010, 3. [Google Scholar] [CrossRef]
- Beyer, T.A.; Xu, W.; Teupser, D.; auf dem Keller, U.; Bugnon, P.; Hildt, E.; Thiery, J.; Kan, Y.W.; Werner, S. Impaired liver regeneration in Nrf2 knockout mice: Role of ROS-mediated insulin/IGF-1 resistance. EMBO J. 2008, 27, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Madrigal-Santillán, E.; Bautista, M.; Gayosso-de-Lucio, J.A.; Reyes-Rosales, Y.; Posadas-Mondragon, A.; Morales-González, A.; Soriano-Ursúa, M.A.; García-Machorro, J.; Madrigal-Bujaidar, E.; Álvarez-González, I.; et al. Hepatoprotective effect of Geranium schiedeanum against ethanol toxicity during liver regeneration. World J. Gastroenterol. 2015, in press. [Google Scholar]
- Michalopoulos, G.K. Nrf2, not always friendly but perhaps misunderstood. Hepatology 2014, 60, 461–463. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morales-González, J.A.; Madrigal-Santillán, E.; Morales-González, Á.; Bautista, M.; Gayosso-Islas, E.; Sánchez-Moreno, C. What is Known Regarding the Participation of Factor Nrf-2 in Liver Regeneration? Cells 2015, 4, 169-177. https://doi.org/10.3390/cells4020169
Morales-González JA, Madrigal-Santillán E, Morales-González Á, Bautista M, Gayosso-Islas E, Sánchez-Moreno C. What is Known Regarding the Participation of Factor Nrf-2 in Liver Regeneration? Cells. 2015; 4(2):169-177. https://doi.org/10.3390/cells4020169
Chicago/Turabian StyleMorales-González, José A., Eduardo Madrigal-Santillán, Ángel Morales-González, Mirandeli Bautista, Evila Gayosso-Islas, and Cecilia Sánchez-Moreno. 2015. "What is Known Regarding the Participation of Factor Nrf-2 in Liver Regeneration?" Cells 4, no. 2: 169-177. https://doi.org/10.3390/cells4020169
APA StyleMorales-González, J. A., Madrigal-Santillán, E., Morales-González, Á., Bautista, M., Gayosso-Islas, E., & Sánchez-Moreno, C. (2015). What is Known Regarding the Participation of Factor Nrf-2 in Liver Regeneration? Cells, 4(2), 169-177. https://doi.org/10.3390/cells4020169