Cardiomyocyte Regeneration


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Stem Cells Used for Cardiomyocyte Regeneration
2.1. BMCs
2.1.1. Adult CSCs
2.1.2. c-Kit Positive Cells
2.2. Cardiosphere-Forming Cells
2.3. Other Stem Cells Relating to the Heart
2.4. Left-Atrium-Derived c-Kit Positive Cells
2.5. Similarity of LA-PCs to Other Stem Cells
3. Signal Transduction of the Differentiation Switch Using LA-PCs
3.1. Microarray Analysis for Identification
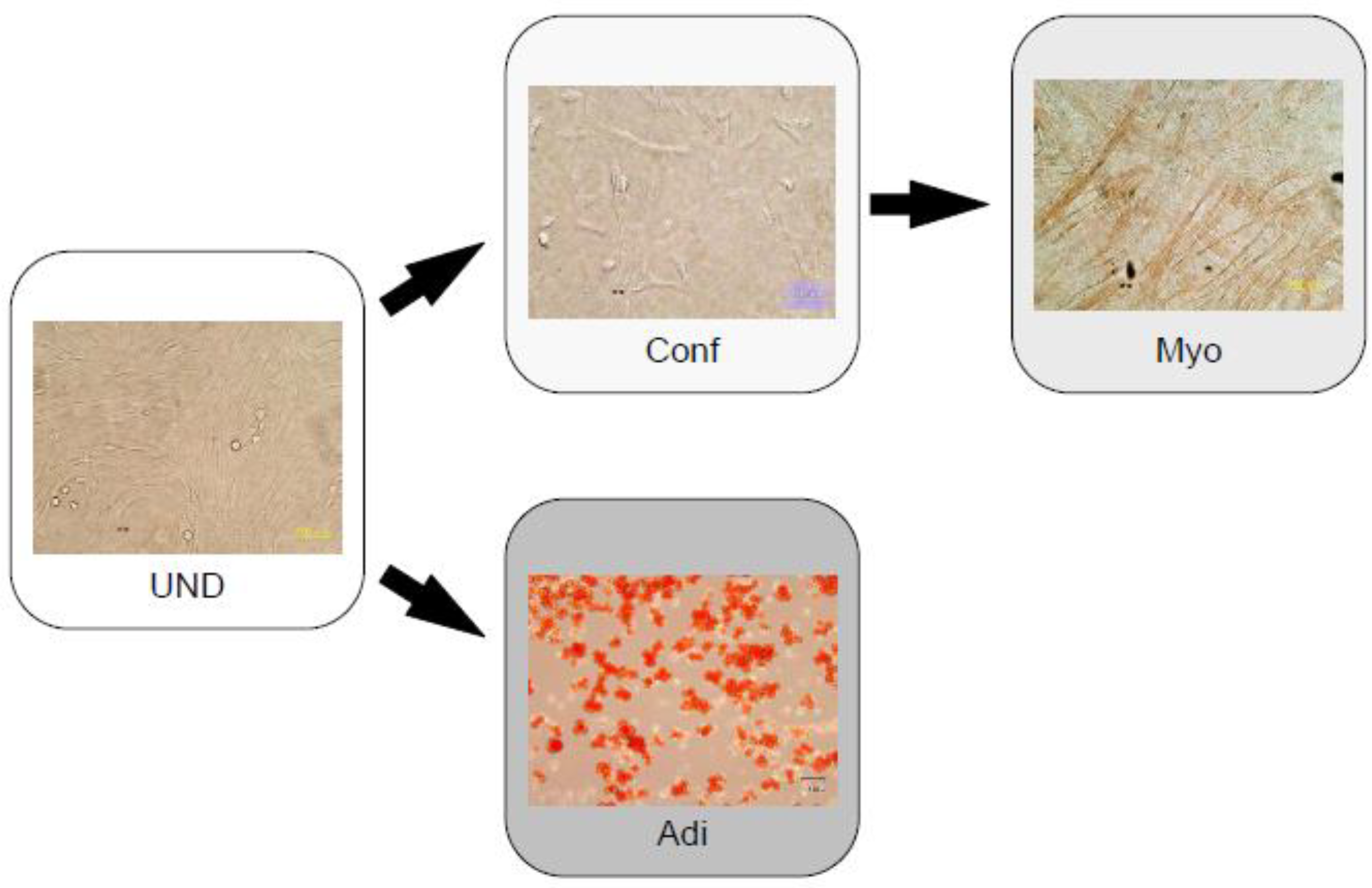
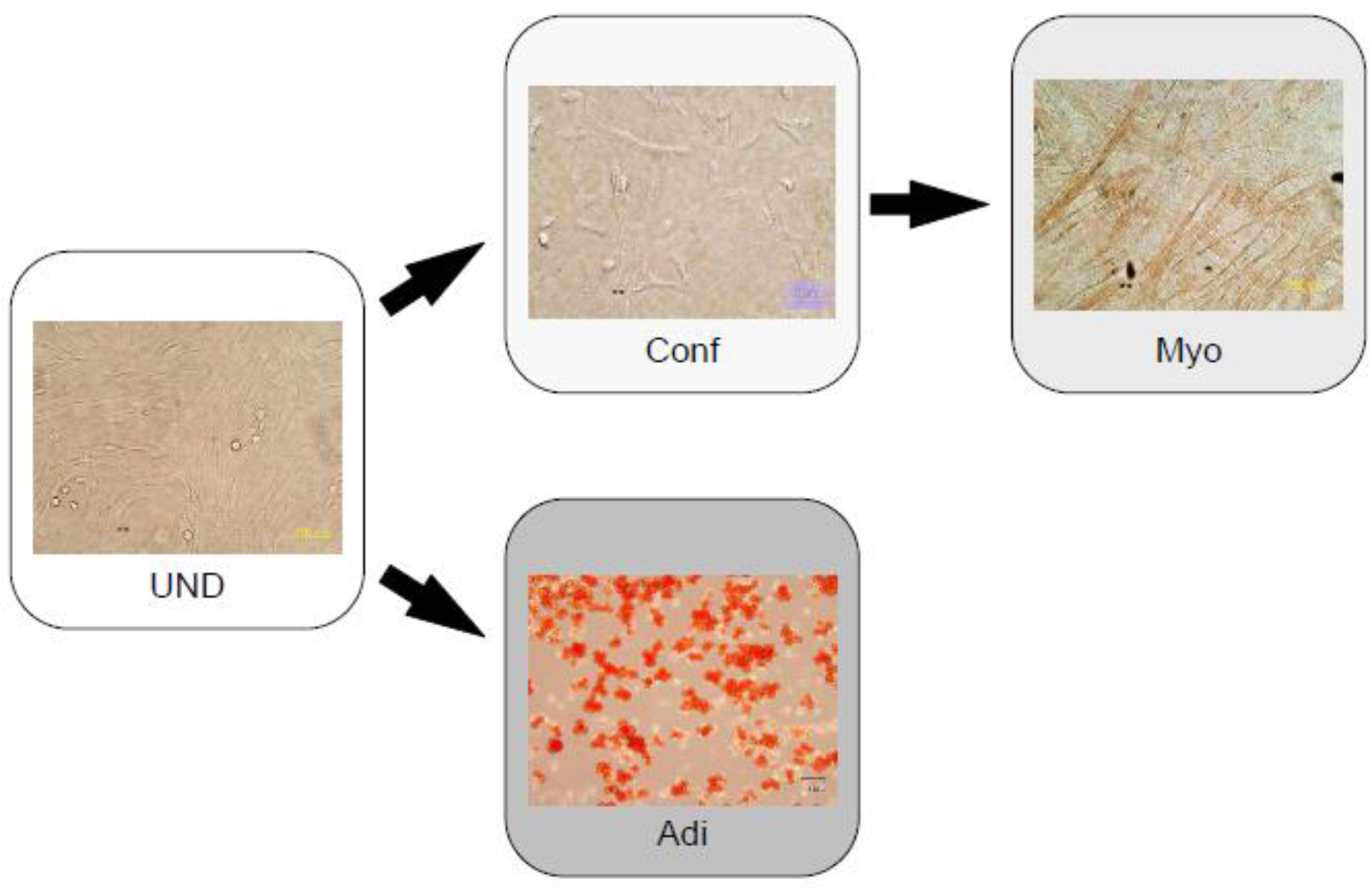
3.2. Noggin and Cardiac Myocytes
4. iPSC-Derived Cardiomyocytes
5. Direct Reprogramming to Induce Cardiomyocyte Differentiation
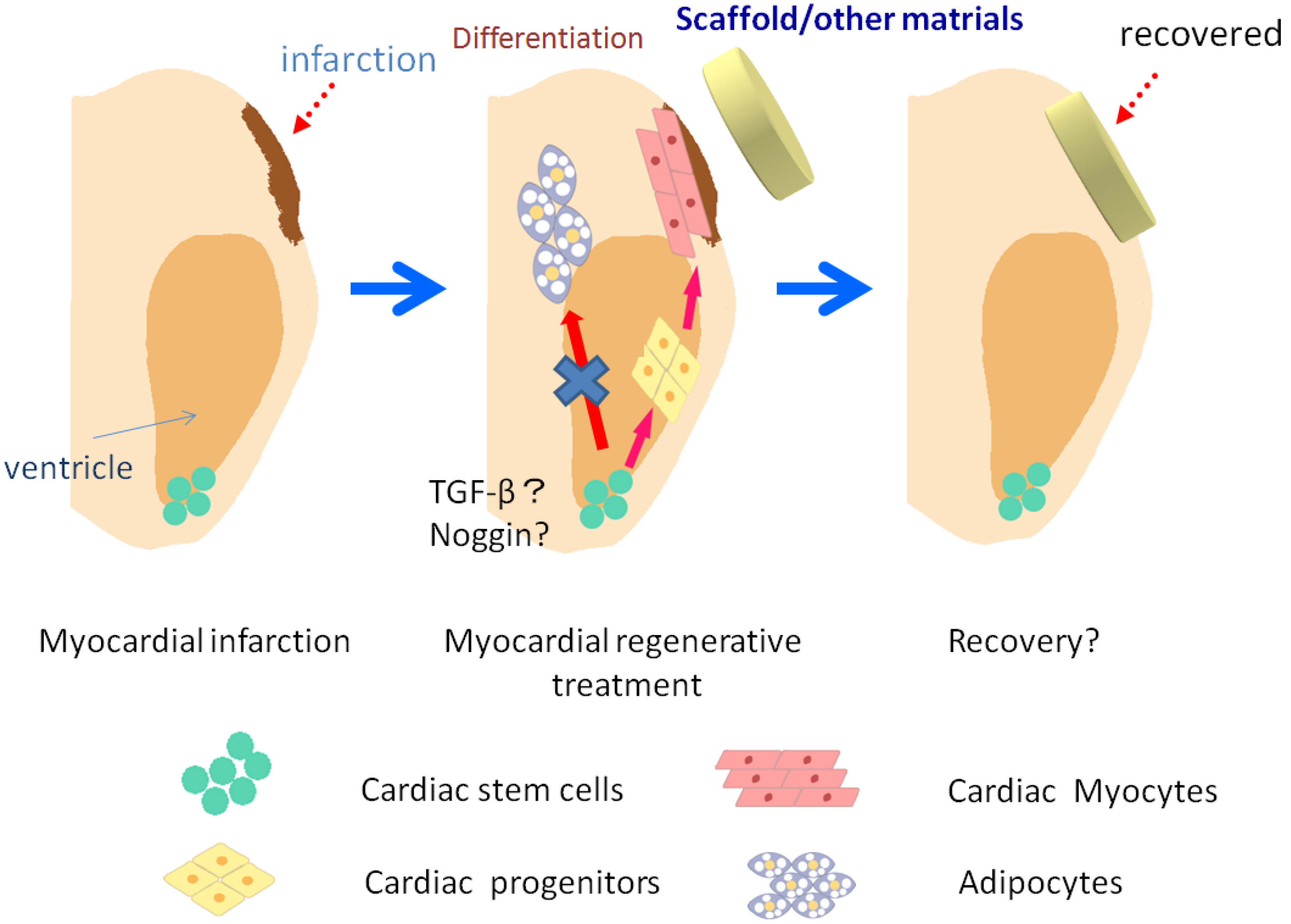
6. Regenerative Medicine Using Stem Cells
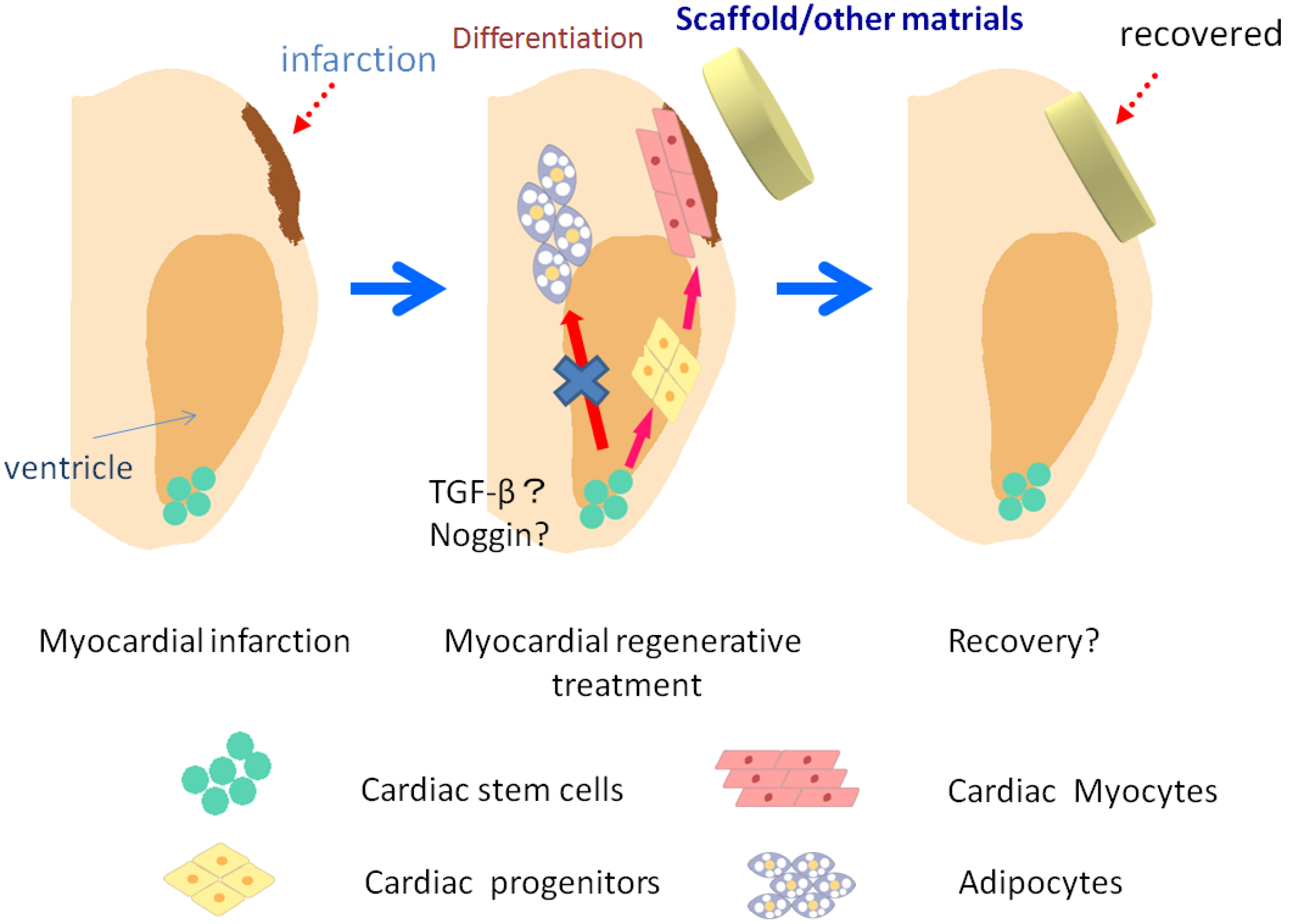
7. Conclusion
References
- Braunwald, E.; Bristow, M. Congestive heart failure: Fifty years of progress. Circulation 2000, 102, IV14–IV23. [Google Scholar]
- Braunwald, E.; Pfeffer, M. Ventricular enlargement and remodeling following acute myocardial infarction: Mechanisms and management. Am. J. Cardiol. 1991, 68, D1–D6. [Google Scholar] [CrossRef]
- Leri, A.; Kajstura, J.; Anversa, P.; Frishman, W.H. Myocardial regeneration and stem cell repair. Curr. Probl. Cardiol. 2008, 33, 91–153. [Google Scholar] [CrossRef]
- Beltrami, A.P.; Urbanek, K.; Kajstura, J.; Yan, S.M.; Finato, N.; Bussani, R.; Nadal-Ginard, B.; Silvestri, F.; Leri, A.; Beltrami, C.A.; et al. Evidence that human cardiac myocytes divide after myocardial infarction. N. Engl. J. Med. 2001, 344, 1750–1757. [Google Scholar] [CrossRef]
- Beltrami, A.P.; Barlucchi, L.; Torella, D.; Baker, M.; Limana, F.; Chimenti, S.; Kasahara, H.; Rota, M.; Musso, E.; Urbanek, K.; et al. Adult cardiac stem cells are multipotent and support myocardial regeneration. Cell 2003, 114, 763–776. [Google Scholar] [CrossRef]
- Matsuura, K.; Nagai, T.; Nishigaki, N.; Oyama, T.; Nishi, J.; Wada, H.; Sano, M.; Toko, H.; Akazawa, H.; Sato, T.; et al. Adult cardiac Sca-1-positive cells differentiate into beating cardiomyocytes. J. Biol. Chem. 2004, 279, 11384–11391. [Google Scholar]
- Messina, E.; De Angelis, L.; Frati, G.; Morrone, S.; Chimenti, S.; Fiordaliso, F.; Salio, M.; Battaglia, M.; Latronico, M.V.; Coletta, M.; et al. Isolation and expansion of adult cardiac stem cells from human and murine heart. Circ. Res. 2004, 95, 911–921. [Google Scholar] [CrossRef]
- Oh, H.; Bradfute, S.B.; Gallardo, T.D.; Nakamura, T.; Gaussin, V.; Mihina, Y.; Pocius, J.; Michael, L.H.; Behringer, R.R.; Garry, D.J.; et al. Cardiac progenitor cells from adult myocardium: homing, differentiation, and fusion after infarction. Proc. Natl. Acad. Sci. USA 2003, 100, 12313–12318. [Google Scholar]
- Linke, A.; Müller, P.; Nurzynska, D.; Casarsa, C.; Torella, D.; Nascimbene, A.; Castaldo, C.; Cascapera, S.; Böhm, M.; Quaini, F.; et al. Stem cells in the dog heart are self-renewing, clonogenic, and multipotent and regenerate infarcted myocardium, improving cardiac function. Proc. Natl. Acad. Sci. USA 2005, 102, 8966–8971. [Google Scholar]
- Bearzi, C.; Rota, M.; Hosoda, T.; Tillmanns, J.; Nascimbene, A.; Angelis, A.D.; Yasuzawa-Amano, S.; Trofimova, I.; Siggins, R.W.; LeCaptitaine, N.; et al. Human cardiac stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 14068–14073. [Google Scholar]
- Urbanek, K.; Torella, D.; Sheikh, F.; Angelis, A.D.; Nurzynska, D.; Silvesti, F.; Beltrami, C.A.; Bussani, R.; Beltrami, A.P.; Quaini, F. Myocardial regeneration by activation of multipotent cardiac stem cells in ischemic heart failure. Proc. Natl. Acad. Sci. USA 2005, 102, 8692–8697. [Google Scholar]
- Martin, C.; Meeson, A.; Robertson, S.M.; Hawke, T.J.; Richardson, J.A.; Bates, S.; Goetsch, S.C.; Gallardo, T.D.; Garry, D.J. Persistent expression of the ATP-binding cassette transporter Abcg2 identifies cardiac SP cells in the developing and adult heart. Dev. Biol. 2004, 265, 262–275. [Google Scholar] [CrossRef]
- Pfister, O.; Oikonomopoulos, A.; Sereti, K.I.; Sohn, R.L.; Cullen, D.; Fine, G.C.; Monquet, F.; Westerman, K.; Liao, R. Role of the ATP binding cassette transporter Abcg2 in the phenotype and function of cardiac side population cells. Circ. Res. 2008, 103, 825–835. [Google Scholar] [CrossRef]
- Oyama, T.; Nagai, T.; Wada, H.; Naito, A.T.; Matsuura, K.; Iwanaga, K.; Takahashi, T.; Goto, M.; Mikami, Y.; Yasuda, N.; et al. Cardiac side population cells have a potential to migrate and differentiate into cardiomyocytes in vitro and In vivo. J. Cell. Biol. 2007, 176, 329–341. [Google Scholar] [CrossRef]
- Smith, R.R.; Barile, L.; Cho, H.C.; Leppo, M.K.; Hare, J.M.; Messina, E.; Giacomello, A.M.; Abraham, R.; Marbán, E. Regenerative potential of cardiosphere-derived cells expanded from percutaneous endomyocardial biopsy specimens. Circulation 2007, 115, 896–908. [Google Scholar] [CrossRef]
- Davis, D.R.; Zhang, Y.; Smith, R.R.; Cheng, K.; Terrovitis, J.; Malliaras, K.; Li, T.-S.; White, A.; Makker, R.; Marbán, E. Validation of the cardiosphere method to culture cardiac progenitor cells from myocardial tissue. PLoS One 2009, 4, e7195. [Google Scholar]
- Davis, D.R.; Kizana, E.; Terrovitis, J.; Barth, A.S.; Zhang, Y.; Smith, R.R.; Miake, J.; Marbán, E. Isolation and expansion of functionally-competent cardiac progenitor cells directly from heart biopsies. J. Mol. Cell. Cardiol. 2010, 49, 312–321. [Google Scholar] [CrossRef]
- Johnston, P.V.; Sasano, T.; Mills, K.; Evers, R.; Lee, S.-T.; Smith, R.R.; Lardo, A.C.; Lai, S.; Steenbergen, C.; Gertenblith, G.; et al. Engraftment, differentiation, and functional benefits of autologous cardiosphere-derived cells in porcine ischemic cardiomyopathy. Circulation 2009, 120, 1075–1083. [Google Scholar] [CrossRef]
- Chimenti, I.; Smith, R.R.; Li, T.-S.; Gerstenblith, G.; Messina, E.; Giacomello, A.; Marbán, E. Relative roles of direct regeneration versus paracrine effects of human cardiosphere-derived cells transplanted into infracted mice. Circ. Res. 2010, 106, 971–980. [Google Scholar] [CrossRef]
- Cheng, K.; Li, T.-S.; Malliaras, K.; Davis, D.R.; Zhang, Y.; Marbán, E. Magnetic targeting enhances engraftment and functional benefit of iron-labeled cardiosphere-derived cells in myocardial infarction. Circ. Res. 2010, 106, 1570–1581. [Google Scholar] [CrossRef]
- Malliaras, K.; Li, T.-S.; Luthringer, D.; Terrovitis, J.; Cheng, K.; Chakravarty, T.; Galang, G.; Zhang, Y.; Schoenhoff, F.; Eyk, J.V.; et al. Safety and efficacy of allogenic cell therapy in infarcted rats transplanted with mismatched cardiosphere-derived cells. Circulation 2012, 125, 100–112. [Google Scholar]
- Lee, S.-T.; White, A.J.; Matsushita, S.; Malliaras, K.; Steenbergen, C.; Zhang, Y.; Li, T.-S.; Terrovitis, J.; Yee, K.; Simsir, S.; et al. Intramyocardial injection of autologous cardiospheres or cardiosphere-derived cells preserves function and minimizes adverse ventricular remodeling in pigs with heart failure post-myocardial infarction. J. Am. Coll. Cardiol. 2011, 57, 455–465. [Google Scholar]
- White, A.J.; Smith, R.R.; Matsushita, S.; Chakravarty, T.; Czer, L.S.C.; Burton, K.; Schwarz, E.R.; Davis, D.R.; Wang, Q.; Reinsmoen, N.L.; et al. Intrinsic cardiac origin of human cardiosphere-derived cells. Eur. Heart J. 2011, 172, 68–75. [Google Scholar]
- Lautamki, R.; Terrovitis, J.; Bonios, M.; Yu, J.; Tsui, B.M.; Abraham, M.R.; Bengel, F.M. Perfusion defect size predicts engraftment but not early retention of intra-myocardially injected cardiosphere-derived cells after acute myocardial infarction. Basic Res. Cardiol. 2011, 106, 1379–1386. [Google Scholar] [CrossRef]
- Carr, C.A.; Stuckey, D.J.; Tan, J.J.; Tan, S.C.; Gomes, R.S.M.; Camelliti, P.; Messina, E.; Giacomello, A.; Ellison, G.M.; Clarke, K. Cardiosphere-derived cells improve function in the infarcted rat heart for at least 16 weeks-an MRI study. PLoS One 2011, 6, e25669. [Google Scholar]
- Makker, R.R.; Smith, R.R.; Cheng, K.; Malliaras, K.; Thomson, L.E.J.; Berman, D.; Czer, L.S.C.; Marban, L.; Mendizabal, A.; Johnston, P.V.; et al. Intracoronary cardiosphere-derived cells for heart regeneration after myocardial infarction (CADUCEUS): A prospective, randomized phase 1 trial. Lancet 2012, in press. [Google Scholar]
- Bolli, R.; Chugh, A.R.; D’Amario, D.; Loughran, J.H.; Stoddard, M.F.; Ikram, S.; Beache, G.M.; Wagner, S.G.; Leri, A.; Hosoda, T.; et al. Cardiac stem cells in patients with ischaemic cardiomyopathy (SCIPIO): Initial results of a randomized phase 1 trial. Lancet 2011, 378, 1847–1857. [Google Scholar] [CrossRef]
- Eisenberg, J.M.; Burns, L.; Eisenberg, C.A. Hematopoietic cells from bone marrow have the potential to differentiate into cardiomyocytes in vitro. Anat. Rec. A Discov. Mol. Cell Evol. Biol. 2003, 274, 870–882. [Google Scholar]
- Orlic, D.; Kajstura, J.; Chimenti, S.; Jakonik, I.; Anderson, S.M.; Li, B.; Pickel, J.; Mckay, R.; Nadal-Ginard, B.; Bodine, D.M. Bone marrow cells regenerate infarcted myocardium. Nature 2001, 410, 701–705. [Google Scholar] [CrossRef]
- Psaltis, P.J.; Zannettino, A.C.W.; Worthley, S.G.; Gronthos, S. Concise review: Mesenchymal stromal cells: Potential for cardiovascular repair. Stem Cells 2008, 26, 2201–2210. [Google Scholar] [CrossRef]
- Abarbanell, A.M.; Coffey, A.C.; Fehrenbacher, J.W.; Beckman, D.J.; Herrmann, J.L.; Weil, B.; Meldrum, D.R. Proinflammatory cytokine effects on mesenchymal stem cell therapy for the ischemic heart. Ann. Thorac. Surg. 2009, 88, 1036–1043. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Martin, B.J. Mesenchymal stem cells and their potential as cardiac therapeutics. Circ. Res. 2004, 95, 9–20. [Google Scholar] [CrossRef]
- Uemura, R.; Xu, M.; Ahmad, N.; Ashraf, M. Bone marrow stem cells prevent left ventricular remodeling of ischemic heart through paracrine signaling. Circ. Res. 2006, 98, 1414–1421. [Google Scholar] [CrossRef]
- Murry, C.; Soonpaa, M.; Reinecke, H.; Nakajima, H.; Nakajima, H.O.; Rubart, M.; Pasumarthi, K.B.; Virag, J.I.; Bartelmez, S.H.; Poppa, V.; et al. Hematopoietic stem cells do not differentiate into cardiac myocytes in myocardial infarcts. Nature 2004, 428, 664–668. [Google Scholar] [CrossRef]
- Fazel, S.; Cimini, M.; Chen, L.; Li, S.; Angoulvant, D.; Fedak, P.; Verma, S.; Weisel, R.D.; Keating, A.; Li, R. Cardioprotective c-kit+ cells are from the bone marrow and regulate the myocardial balance of angiogenic cytokines. J. Clin. Invest. 2006, 116, 1865–1877. [Google Scholar]
- Chein, K.R. Lost and found: cardiac stem cell therapy revisited. J. Clin. Invest. 2006, 116, 1838–1840. [Google Scholar] [CrossRef]
- Sun, J.; Li, S.; Liu, S.; Wu, J.; Weisel, R.D.; Zhuo, Y.; Yau, F.M.; Li, R.; Fazel, S.S. Improvement in cardiac function after bone marrow cell therapy is associated with increase in myocardial inflammation. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H43–H50. [Google Scholar]
- Fazel, S.S.; Chen, L.; Angoulvant, D.; Li, S.; Weisel, R.D.; Keating, A.; Li, R. Activation of c-kit is necessary for mobilization of reparative bone marrow progenitor cells in response to cardiac injury. FASEB J. 2008, 22, 930–940. [Google Scholar]
- Cimini, M.; Fazel, S.; Zhuo, S.; Xaymardan, M.; Fujii, H.; Weisel, R.D.; Li, R. c-Kit dysfunction impairs myocardial healing after infarction. Circulation 2007, 116, I77–I82. [Google Scholar]
- Gaebel, R.; Furlani, D.; Sorg, H.; Polchow, B.; Frank, J.; Bieback, K.; Wang, W.; Klopsch, C.; Ong, L.; Li, W.; et al. Cell origin of human mesenchymal stem cells determines a different healing performance in cardiac regeneration. PLoS One 2011, 6, e15652. [Google Scholar]
- Anversa, P.; Kajstura, J.; Leri, A.; Bolli, R. Life and death of cardiac stem cells. A paradigm shift in cardiac biology. Circulation 2006, 113, 1451–1463. [Google Scholar] [CrossRef]
- Hosoda, T.; Kajstura, J.; Leri, A.; Anversa, P. Mechanism of myocardial regeneration. Circ. J. 2010, 74, 13–17. [Google Scholar] [CrossRef]
- Tallini, Y.; Greene, K.; Craven, M.; Spealman, A.; Breitbach, M.; Smith, J.; Fisher, P.J.; Steffey, M.; Hesse, M.; Doran, R.M.; et al. c-Kit expression identifies cardiovascular precursors in the neonatal heart. Proc. Natl. Acad. Sci. USA 2009, 106, 1808–1813. [Google Scholar]
- Kawaguchi, N. Stem cells for cardiac regeneration and possible roles of the transforming growth factor family. Biomol. Concept. 2012, 3, 99–106. [Google Scholar]
- Miyamoto, S.; Kawaguchi, N.; Ellison, G.M.; Shin’oka, T.; Matsuoka, R.; Kurosawa, H. Characterization of long-term cultured cardiac stem cells (CSCs) derived from adult rat hearts. Stem Cells Dev. 2010, 19, 105–116. [Google Scholar] [CrossRef]
- Torella, D.; Rota, M.; Nurzynska, D.; Musso, E.; Monsen, A.; Shiraishi, I.; Zias, E.; Walsh, K.; Rosenzweig, A.; Sussman, M.A.; et al. Cardiac stem cell and myocyte aging, heart failure, and insulin-like growth factor-1 overexpression. Circ. Res. 2004, 94, 514–524. [Google Scholar] [CrossRef]
- Wang, L.; Ma, W.; Markovich, R.; Chen, J.W.; Wang, P.H. Regulation of cardiomyocyte apoptotic signaling by insulin-like growth factor I. Circ. Res. 1998, 83, 516–252. [Google Scholar] [CrossRef]
- Fujio, Y.; Nguyen, T.; Wencker, D.; Kitsis, R.N.; Walsh, K. Akt promotes survival of cardiomyocytes in vitro and protects against ischemia-reperfusion injury in mouse heart. Circulation 2000, 101, 660–667. [Google Scholar] [CrossRef]
- Kawaguchi, N.; Smith, A.J.; Waring, C.D.; Hasan, M.K.; Miyamoto, S.; Matsuoka, R.; Ellison, G.M. c-Kit pos GATA-4 high rat cardiac stem cells foster adult cardiomyocyte survival through IGF-1 paracrine signalling. PLoS One 2010, 5, e14297. [Google Scholar]
- Kawaguchi, N. Adult cardiac-derived stem cells: Differentiation and survival regulators. In Vitamins and Hormones; Litwack, G., Ed.; Academic Press Elsevier Inc.: Oxford, UK, 2011; Volume 87, pp. 111–125. [Google Scholar]
- Suslov, O.; Kukekov, V.; Ignatova, T.; Steindler, D. Neural stem cell heterogeneity demonstrated by molecular phenotyping of clonal neurospheres. Proc. Natl. Acad. Sci. USA 2002, 99, 14506–11451. [Google Scholar]
- Andersen, D.C.; Andersen, P.; Schneider, M.; Jensen, H.B.; Sheikh, S.P. Murine “cardiospheres” are not a source of stem cells with cardiomyogenic potential. Stem Cells 2009, 27, 1571–1581. [Google Scholar] [CrossRef]
- Machida, M.; Takagaki, Y.; Matsuoka, R.; Kawaguchi, N. Proteomic comparison spherical aggregates and adherent cells of cardiac stem cells. Int. J. Cardiol. 2011, 153, 296–305. [Google Scholar] [CrossRef]
- Fathi, A.; Pakzad, M.; Taei, A.; Brink, T.C.; Pirhaji, L.; Ruiz, G.; Bordbar, M.S.T.; Gourabi, H.; Adjaye, J.; Baharvand, H.; et al. Comparative proteome and transcriptome analyses of embryonic stem cells during embryoid body-based differentiation. Proteomics 2009, 9, 4859–4870. [Google Scholar] [CrossRef]
- Wojakowski, W.; Tendera, M.; Michalowska, A.; Majka, M.; Kucia, M.; Malanskiewicz, K.; Wyderka, R.; Ochala, A.; Ratajczak, M.Z. Mobilization of CD34/CXCR4 +, CD34/CD117 +, c-met + stem cells, and mononuclear cells expressing early cardiac, muscle, and endothelin markers into peripheral blood in patients with acute myocardial infarction. Circulation 2004, 110, 3213–3220. [Google Scholar] [CrossRef]
- Tang, J.; Wang, J.; Zhang, L.; Zheng, F.; Yang, J.; Kong, X.; Guo, L.; Chen, L.; Huang, Y.; Wan, Y.; et al. VEGF/SDF-1 promotes cardiac stem cell mobilization and myocardial repair in infarcted heart. Cadiovasc. Res. 2011, 91, 402–411. [Google Scholar] [CrossRef]
- Morimoto, H.; Takahashi, M.; Shiba, Y.; Izawa, A.; Ise, H.; Hongo, M.; Hatake, K.; Motoyoshi, K.; Ikeda, U. Bone marrow-derived CXCR4+ cells mobilized by macrophages colony-stimulating factor participate in the reduction of infarct area and improvement of cardiac remodeling after myocardial infarction in mice. Am. J. Pathol. 2007, 171, 755–766. [Google Scholar] [CrossRef]
- Laugwitz, K.; Moretti, A.; Lam, J.; Gruber, P.; Chen, Y.; Woodard, S.; Lin, L.Z.; Cai, C.L.; Lu, M.M.; Reth, M.; et al. Postnatal isl1 + cardioblasts enter fully differentiated cardiomyocyte lineages. Nature 2005, 433, 647–653. [Google Scholar]
- Moretti, A.; Caron, L.; Nakano, A.; Lam, J.T.; Bernshausen, A.; Chen, Y.; Qyang, Y.; Bu, L.; Sasaki, M.; Martin-Puig, S.; et al. Mtipotent embryonic isl1 + progenitor cells lead to cardiac smooth muscle and endothelial cell diversification. Cell 2006, 127, 1151–1165. [Google Scholar] [CrossRef]
- Ott, H.C.; Matthiesen, T.S.; Brechtken, J.; Grindle, S.; Goh, S.K.; Nelson, W.; Tayler, D.A. The adult human as a source for stem cells: Repair strategies with embryonic-like progenitor cells. Nat. Clin. Pract. Cardiovasc. Med. 2007, 4, S27–S39. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–172. [Google Scholar] [CrossRef]
- Park, I.H.; Zhao, R.; West, J.A.; Yabuuchi, A.; Huo, H.; Ince, T.A.; Lerou, P.H.; Lensch, M.W.; Daley, G.Q. Reprogramming of human somatic cells to pluripotency with defined factors. Nature 2008, 451, 141–146. [Google Scholar]
- Hinescu, M.E.; Popescu, L.M. Interstitial Cajal-like cells (ICLC) in human atrial myocardium. J. Cell Mol. Med. 2005, 9, 972–975. [Google Scholar] [CrossRef]
- Kostin, S.; Popescu, L.M. A distinct type of cell in myocardium: Interstitial Cajal-like cells (ICLC). J. Cell Mol. Med. 2009, 13, 295–308. [Google Scholar] [CrossRef]
- Gherghiceanu, M.; Manole, C.G.; Popescu, L.M. Telocytes in endocarium: Electron microscope evidence. J. Cell Mol. Med. 2010, 14, 2330–2334. [Google Scholar] [CrossRef]
- Popescu, L.M.; Manole, C.G.; Gherghiceanu, M.; Aedelean, A.; Nicolescu, M.I.; Hinescu, M.E.; Kostin, S. Telocytes in human epicardium. J. Cell Mol. Med. 2010, 14, 2085–2093. [Google Scholar] [CrossRef]
- Hasan, M.K.; Komoike, Y.; Tsunesumi, S.; Nakao, R.; Nagao, H.; Matsuoka, R.; Kawaguchi, N. Myogenic differentiation in atrium derived adult cardiac pluripotent cells and the transcriptional regulation of GATA4 and myogenin on ANP promoter. Genes Cells 2010, 15, 439–453. [Google Scholar]
- Danalache, B.; Paquin, J.; Donghao, W.; Gyrgorczyk, R.; Moore, J.C.; Mummery, C.L.; Gutkowska, J.; Jankowski, M. Nitric oxide signaling in oxytocin-mediated cardiomyogenesis. Stem Cells 2007, 25, 679–688. [Google Scholar]
- Minasi, M.G.; Riminucci, M.; De Angelis, L.; Borello, U.; Berarducci, B.; Innocenzi, A.; Caprioli, A.; Sirabella, D.; Baiocchi, M.; De Maria, R.; et al. The meso-angioblast: A multipotent, self-renewing cell that originates from the dorsal aorta and differentiates into most mesodermal tissues. Development 2002, 129, 2773–2783. [Google Scholar]
- Kawaguchi, N.; Nakao, R.; Yamaguchi, M.; Ogawa, D.; Matsuoka, R. TGF-β superfamily regulates a switch that mediates differentiation either into adipocytes or myocytes in left atrium derived pluripotent cells (LA-PCS). Biochem. Biophys. Res. Commun. 2010, 396, 619–625. [Google Scholar] [CrossRef]
- Gilbert, S.F. Developmental Biology, 7th ed; Sinauer Associates Inc.: Sunderland, MA, USA, 2003. [Google Scholar]
- Abdel-Latif, A.; Zuba-Surma, E.K.; Case, J.; Tiwari, S.; Hunt, G.; Ranjan, S.; Vincent, R.J.; Srour, E.F.; Bolli, R.; Dawn, B. TGF-β1 enhances cardiomyogenic differentiation of skeletal muscle-derived adult primitive cells. Basic Res. Cardiol. 2008, 103, 514–524. [Google Scholar] [CrossRef]
- Singh, R.; Bhasin, S.; Braga, M.; Artaza, J.N.; Pervin, S.; Taylor, W.E.; Krishnan, V.; Sinha, S.K.; Rajavashisth, T.B.; Jasuja, R. Regulation of myogenic differentiation by androgens: Cross talk between androgen receptor/β-catenin and follistatin/transforming growth factor-β signaling pathways. Endocrinology 2009, 150, 1259–1268. [Google Scholar]
- Sordella, R.; Jiang, W.; Chen, G.; Curto, M.; Settleman, J. Modulation of Rho GTPase signalling regulates a switch between adipogenesis and myogenesis. Cell 2003, 113, 147–158. [Google Scholar] [CrossRef]
- Yuasa, S.; Itabashi, Y.; Koshimizu, U.; Tanaka, T.; Sugimura, K.; Kinoshita, M.; Hattori, F.; Fukami, S.; Shimazaki, T.; Ogawa, S.; et al. Transient inhibition of BMP signaling by noggin induces cardiomyocyte differentiation of mouse embryonic stem cells. Nat. Biotechnol. 2005, 23, 607–611. [Google Scholar] [CrossRef]
- Kawaguchi, N.; Hayama, E.; Furutani, Y.; Nakanishi, T. Prospective in vitro models of channelopathies and cardiomyopathies. Stem Cell Int. 2012. [Google Scholar] [CrossRef]
- Yang, L.; Soonpaa, M.H.; Adler, E.D.; Roepke, T.K.; Kattman, S.J.; Kennedy, M.; Henckaerts, E.; Bonham, K.; Abbott, G.W.; Linden, R.M.; et al. Human cardiovascular progenitor cells develop from a KDR+ embryonic-stem-cell-derived population. Nature 2008, 453, 524–529. [Google Scholar]
- Ren, Y.; Lee, M.Y.; Schliffke, S.; Paavola, J.; Amos, P.J.; Ge, X.; Ye, M.; Zhu, S.; Senyei, G.; Lum, L.; et al. Small molecule Wnt inhibitors enhance the efficiency of BMP-4-directed cardiac differentiation of human pluripotent stem cells. J. Mol. Cell. Cardiol. 2011, 51, 280–287. [Google Scholar] [CrossRef]
- Kattman, S.J.; Witty, A.D.; Gagliardi, M.; Dubois, N.C.; Niapour, M.; Hotta, A.; Ellis, J.; Keller, G. Stage-specific optimization of activin/nodal and BMP signaling promotes cardiac differentiation of mouse and human pluripotent stem cell lines. Cell Stem Cell 2011, 8, 228–240. [Google Scholar] [CrossRef]
- Freund, C.; Mummery, C.L. Prospects for pluripotent stem cell-derived cardiomyocytes in cardiac cell therapy and as disease models. J. Cell. Biochem. 2008, 107, 592–599. [Google Scholar] [CrossRef]
- Yamashita, J.K. ES and iPS cell research for cardiovascular regeneration. Exp. Cell Res. 2010, 316, 2555–2559. [Google Scholar] [CrossRef]
- Zhang, J.; Wilson, G.F.; Soerens, A.G.; Koonce, C.H.; Yu, J.; Palecek, S.P.; Thomson, J.A.; Kamp, T.J. Functional cardiomyocytes derived from human induced pluripotent stem cells. Circ. Res. 2009, 104, e30–e41. [Google Scholar] [CrossRef]
- Khalil, J.; Xi, M.; Shishechian, N.; Hannes, T.; Pfannkuche, K.; Liang, H.; Fatima, A.; Haustein, M.; Suhr, F.; Bloch, W.; et al. Comparison of contractile behavior of native murine ventricular tissue and cardiomyocytes derived from embryonic or induced pluripotent stem cells. FASEB J. 2010, 24, 2739–2751. [Google Scholar] [CrossRef]
- Burridge, P.W.; Thompson, S.; Millrod, M.A.; Weinberg, S.; Yuan, X.; Peters, A.; Mahairaki, V.; Koliatsos, V.E.; Tung, L.; Zambidis, E.T. A universal system for highly efficient cardiac differentiation of human induced pluripotent stem cells that eliminates interline variability. PLoS One 2011, 6, e18293. [Google Scholar]
- Cao, N.; Liu, Z.; Chen, Z.; Wang, J.; Chen, T.; Zhao, X.; Ma, Y.; Qin, L.; Kang, J.; Wei, B.; et al. Ascorbic acid enhances the cardiac differentiation of induced pluripotent stem cells through promoting the proliferation of cardiac progenitor cells. Cell Res. 2012, 22, 219–236. [Google Scholar] [CrossRef]
- Li, L.; Larabee, S.M.; Chen, S.; Basiri, L.; Yamaguchi, S.; Zakaria, A.; Gallicano, G.I. Novel 5′TOP mRNAs regulated by ribosomal S6 kinase are important for cardiomyocyte development: S6 kinase suppression limits cardiac differentiation and promotes pluripotent cells toward a neural lineage. Stem Cells Dev. 2011, 1, 1538–1548. [Google Scholar]
- Kempf, H.; Lecina, M.; Ting, S.; Zweigerdt, R.; Oh, S. Distinct regulation of mitogen-activated protein kinase activities is coupled with enhanced cardiac differentiation of human embryonic stem cells. Stem Cell Res. 2011, 7, 198–209. [Google Scholar] [CrossRef]
- Sachinidis, A.; Schwengberg, S.; Hippler-Altenburg, R.; Mariappan, D.; Kamisetti, N.; Seelig, B.; Berkessel, A.; Hescheler, J. Identification of small signaling molecules promoting cardiac-specific differentiation of mouse embryonic stem cells. Cell. Physiol. Biochem. 2006, 18, 303–314. [Google Scholar] [CrossRef]
- Hao, J.; Daleo, M.A.; Murphy, C.K.; Yu, P.B.; Ho, J.N.; Hu, J.; Peterson, R.T.; Hatzopoulos, A.K.; Hong, C.C. Dorsomorphin, a selective small molecule inhibitor of BMP signaling, promotes cardiomyogenesis in embryonic stem cell. PLoS One 2008, 3, e2094. [Google Scholar]
- Wang, H.J.; Hao, J.; Hong, C.C. Cardiac induction of embryonic stem cells by a small molecule inhibitor of Wnt/β-catenin signaling. ACS Chem. Biol. 2011, 6, 92–197. [Google Scholar]
- Hattori, F.; Chen, H.; Yamashita, H.; Tohyama, S.; Satoh, Y.S.; Yuasa, S.; Li, W.; Yamakawa, H.; Tanaka, T.; Onitsuka, T.; et al. Nngenetic method for purifying stem cell-derived cardiomyocytes. Nat. Methods 2010, 7, 61–66. [Google Scholar] [CrossRef]
- Egashira, T.; Yuasa, S.; Fukuda, K. Induced pluripotent stem cells in cardiovascular medicine. Stem Cells Int. 2011. [Google Scholar] [CrossRef]
- Dubois, N.C.; Craft, A.M.; Sharma, P.; Elliott, D.A.; Stanley, E.G.; Elefanty, A.G.; Gramolini, A.; Keller, G. SIRPA is a specific cell-surface marker for isolating cardiomyocytes derived from human pluripotent stem cells. Nat. Biotechnol. 2011, 29, 1011–1018. [Google Scholar]
- Ieda, M.; Fu, J.; Delgado-Olguin, P.; Vadantham, V.; Hayashi, Y.; Bruneau, B.G.; Srivastava, D. Direct reprogramming of fibroblasts into functional cardiomyocytes by defined factors. Cell 2010, 142, 375–386. [Google Scholar] [CrossRef]
- Qian, L.; Huang, Y.; Spencer, C.J.; Foley, A.; Vedantham, V.; Liu, L.; Conway, S.J.; Fu, J.-D.; Srivastava, D. In vivo reprogramming of murine cardiac fibroblasts into induced cardiomyocytes. Nature 2012, 485, 593–598. [Google Scholar] [CrossRef]
- Song, K.; Nam, Y.-J.; Luo, X.; Qi, X.; Tan, W.; Huang, G.N.; Acharya, A.; Smith, C.L.; Tailquist, M.D.; Neilson, E.G.; et al. Heart repair by reprogramming non-myocytes with cardiac transcription factors. Nature 2012, 485, 599–604. [Google Scholar] [CrossRef]
- Moskowitz, I.P.G.; Kim, J.B.; Moore, M.L.; Wolf, C.M.; Peterson, M.A.; Shendure, J.; Nobrega, M.A.; Yokota, Y.; Berul, C.; Izumo, S.; et al. A molecular pathway including Id2, Tbx5, and Nkx2-5 required for cardiac conduction system development. Cell 2007, 129, 1365–1376. [Google Scholar] [CrossRef]
- Gimble, J.; Katz, A.; Bunnell, B. Adipose-derived stem cells for regenerative medicine. Circ. Res. 2007, 100, 1249–1260. [Google Scholar] [CrossRef]
- Madonna, R.; Geng, Y.; De Caterina, R. Adipose tissue-derived stem cells: characterization and potential for cardiovascular repair. Arterioscler Thromb. Vasc. Biol. 2009, 29, 1723–1729. [Google Scholar] [CrossRef]
- Hosseinkhani, H.; Hosseinkhani, M.; Hattori, S.; Matsuoka, R.; Kawaguchi, N. Micro and nano-scale in vitro 3D culture system for cardiac stem cells. J. Biomed. Mater. Res. Part A 2010, 94, 1–8. [Google Scholar]
- Sekine, H.; Shimizu, T.; Dobashi, I.; Matsuura, K.; Hagiwara, H.; Takahashi, M.; kobayashi, E.; Yamato, M.; Okano, T. Cardiac cell sheet transplantation improves damaged heart function via superior cell survival in comparison with dissociated cell injection. Tissue Eng. Part A 2011, 17, 2973–2980. [Google Scholar] [CrossRef]
- Matsuura, K.; Masuda, S.; Haraguchi, Y.; Yasuda, N.; Shimizu, T.; Hagiwara, N.; Zandstra, P.W.; Okano, T. Creation of mouse embryonic stem cell-derived cardiac cell sheets. Biomaterials 2011, 32, 7355–7362. [Google Scholar] [CrossRef]
- Jiang, X.; Wang, T.; Li, X.; Wu, D.; Zheng, X.; Chen, J.; Peng, B.; Jiang, H.; Huang, C.; Zhang, X. Injection of a novel synthetic hydrogel preserves left ventricle function after myocardial infarction. J. Biomed. Mater. Res. 2009, 90, A472–A477. [Google Scholar]
- Chen, Q.; Ishii, H.; Thouas, G.A.; Lyon, A.R.; Wright, J.S.; Blaker, J.J.; Chrzanowski, W.; Boccaccini, A.R.; Ali, N.N.; Knowles, J.C.; et al. An elastomeric patch derived from poly(glycerol sebacate) for delivery of embryonic stem cells to the heart. Biomaterials 2010, 31, 3883–3893. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kawaguchi, N.; Nakanishi, T. Cardiomyocyte Regeneration. Cells 2013, 2, 67-82. https://doi.org/10.3390/cells2010067
Kawaguchi N, Nakanishi T. Cardiomyocyte Regeneration. Cells. 2013; 2(1):67-82. https://doi.org/10.3390/cells2010067
Chicago/Turabian StyleKawaguchi, Nanako, and Toshio Nakanishi. 2013. "Cardiomyocyte Regeneration" Cells 2, no. 1: 67-82. https://doi.org/10.3390/cells2010067
APA StyleKawaguchi, N., & Nakanishi, T. (2013). Cardiomyocyte Regeneration. Cells, 2(1), 67-82. https://doi.org/10.3390/cells2010067