
The Evolutionary Young Actin Nucleator Cobl Is Important for Proper Amelogenesis


 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice
2.2. RNA Isolation, cDNA Synthesis and RT-PCR
2.3. In Situ Hybridization
2.4. Immunoblotting Analysis
2.5. Electron Microscopy
2.6. Immunofluorescence Analyses of Tissue Sections and Microscopy
2.7. Examinations of Ameloblast Height and of Ameloblast Actin Cytoskeleton
2.8. Measurement of the Enamel Area at P10
2.9. Experimental Design and Statistical Analyses
3. Results
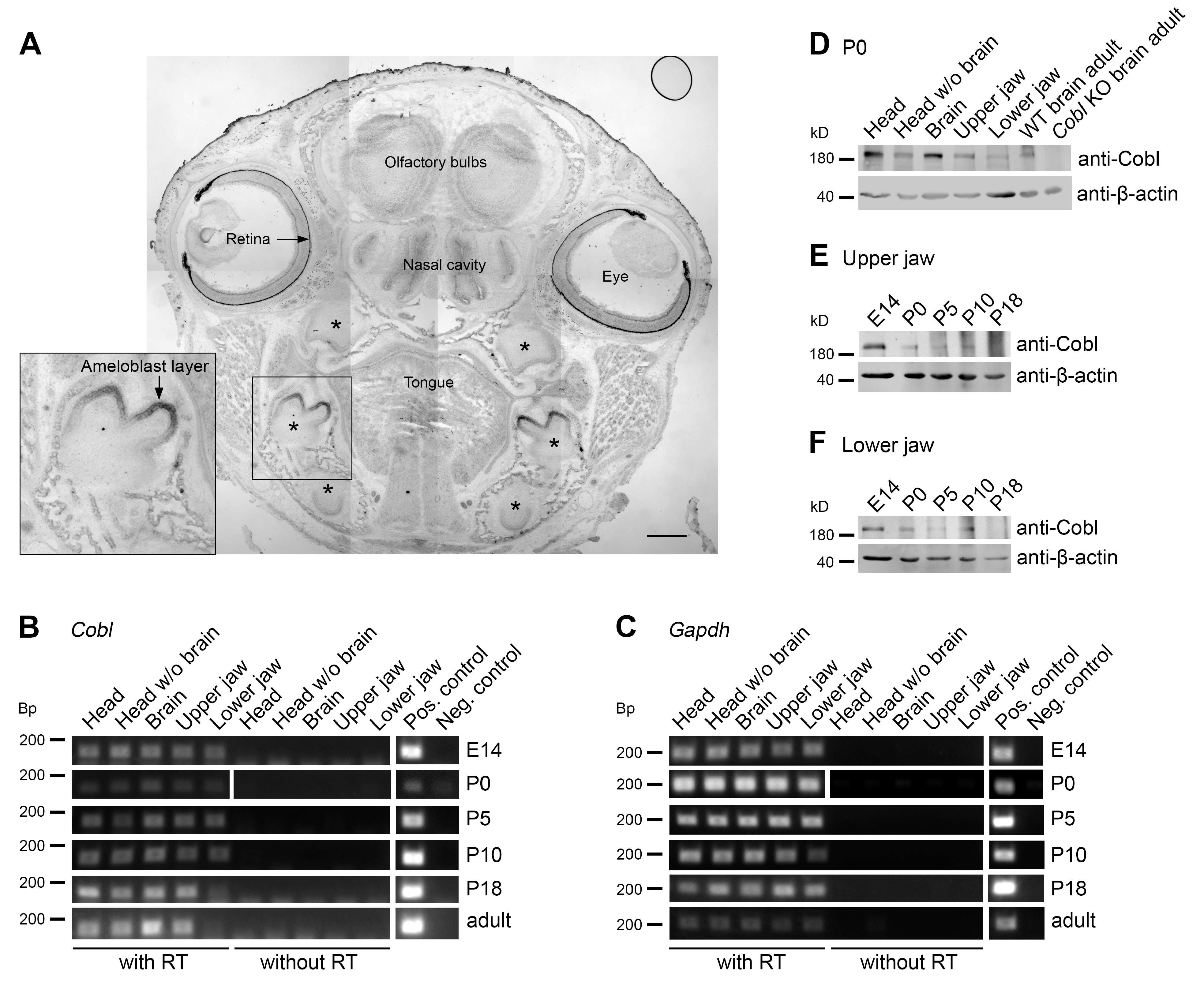
3.1. The Actin Nucleator Cobl Is Expressed in Murine Jaws
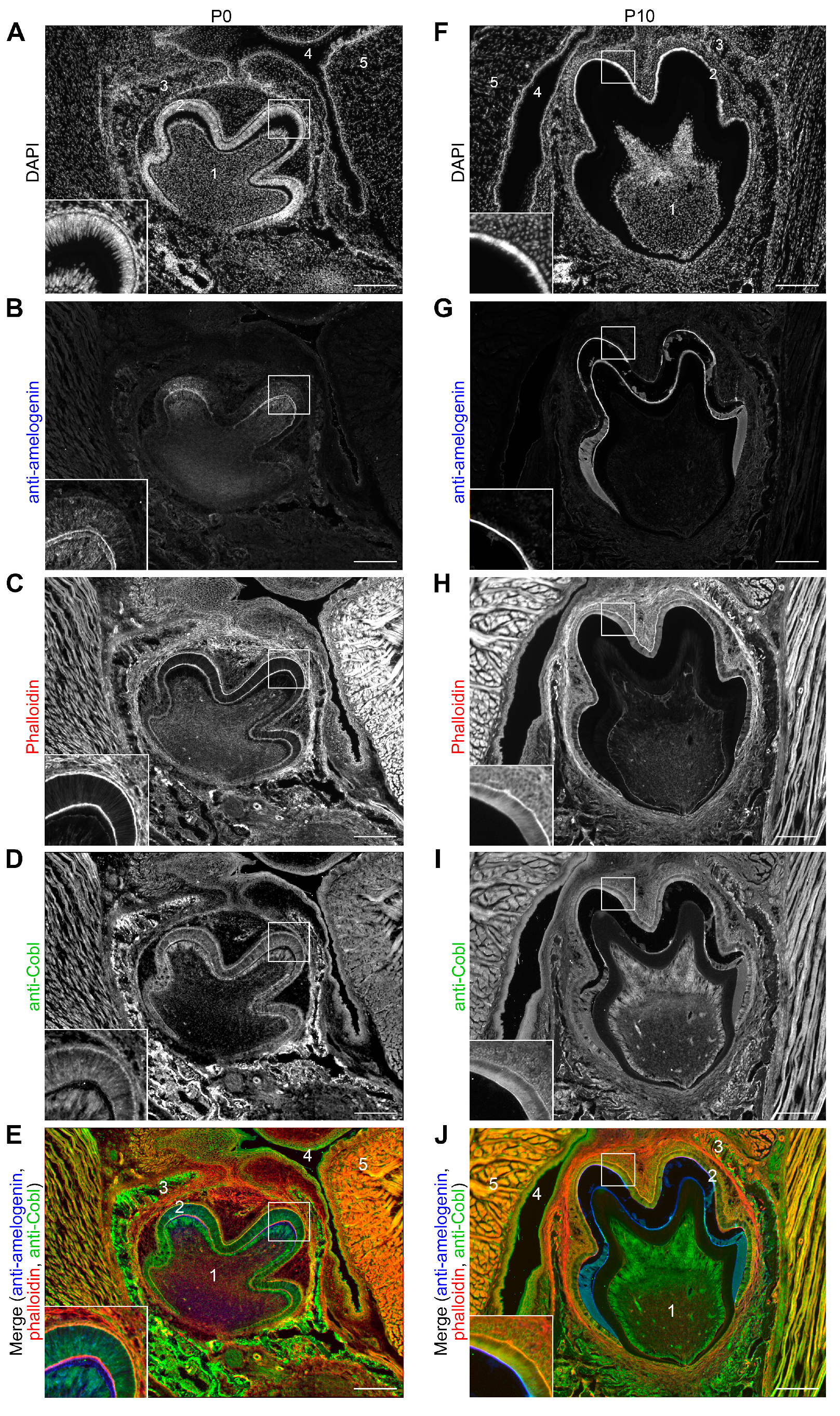
3.2. The Cobl Protein Can Be Detected in the Tooth Germ of a Murine Mandibular Molar at Different Stages of Amelogenesis
3.3. Developing Mandibular Molars of Cobl KO Mice Show an Increased Carbon Content in the Enamel When Compared to WT Molars
3.4. The Absence of Cobl Leads to an Increased Enamel Surface of Developing Mandibular Molars
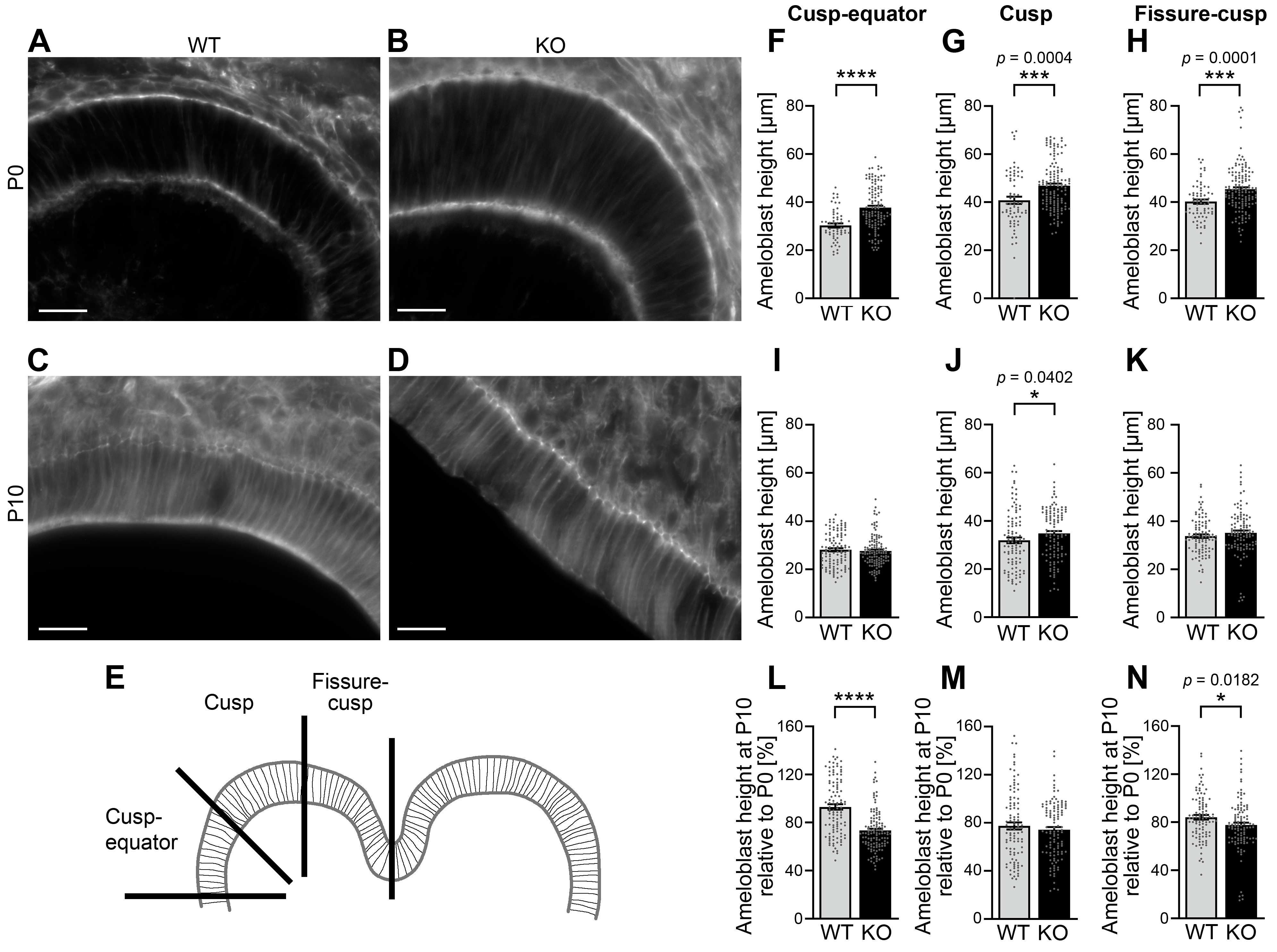
3.5. The Height of Ameloblasts During Amelogenesis Is Increased upon Cobl Deficiency
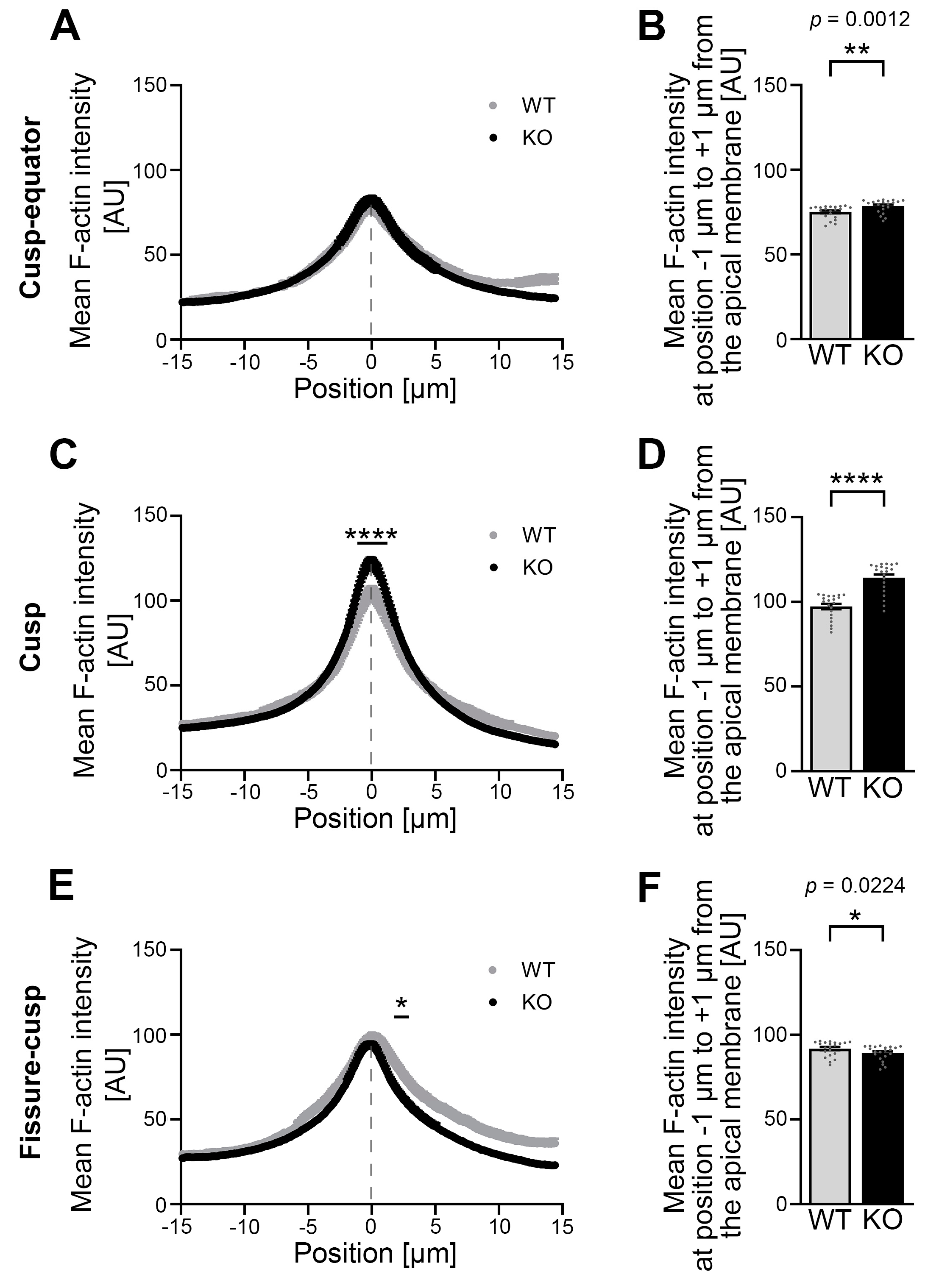
3.6. Developing Ameloblasts of Cobl KO Mice Show an Altered Mean F-Actin Intensity in the Area of the Apical Membrane When Compared to WT Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bartlett, J.D. Dental enamel development: Proteinases and their enamel matrix substrates. ISRN Dent. 2013, 2013, 684607. [Google Scholar] [CrossRef]
- Lacruz, R.S.; Nanci, A.; Kurtz, I.; Wright, J.T.; Paine, M.L. Regulation of pH During Amelogenesis. Calcif. Tissue Int. 2010, 86, 91–103. [Google Scholar] [CrossRef]
- Hu, J.C.; Hu, Y.; Smith, C.E.; McKee, M.D.; Wright, J.T.; Yamakoshi, Y.; Papagerakis, P.; Hunter, G.K.; Feng, J.Q.; Yamakoshi, F.; et al. Enamel defects and ameloblast-specific expression in Enam knock-out/lacz knock-in mice. J. Biol. Chem. 2008, 283, 10858–10871. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, S. Cytoskeleton, intercellular junctions, planar cell polarity, and cell movement in amelogenesis. J. Oral Biosci. 2017, 59, 197–204. [Google Scholar] [CrossRef]
- Bartlett, J.D.; Smith, C.E. Modulation of cell-cell junctional complexes by matrix metalloproteinases. J. Dent. Res. 2013, 92, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, S.; Fujiwara, K.; Kitamura, H. Formation of the tooth enamel rod pattern and the cytoskeletal organization in secretory ameloblasts of the rat incisor. Eur. J. Cell Biol. 1988, 47, 222–232. [Google Scholar]
- Chesarone, M.A.; Goode, B.L. Actin nucleation and elongation factors: Mechanisms and interplay. Curr. Opin. Cell Biol. 2009, 21, 28–37. [Google Scholar] [CrossRef]
- Dominguez, R. Actin filament nucleation and elongation factors—structure-function relationships. Crit. Rev. Biochem. Mol. Biol. 2009, 44, 351–366. [Google Scholar] [CrossRef]
- Ahuja, R.; Pinyol, R.; Reichenbach, N.; Custer, L.; Klingensmith, J.; Kessels, M.M.; Qualmann, B. Cordon-bleu is an actin nucleation factor and controls neuronal morphology. Cell 2007, 131, 337–350. [Google Scholar] [CrossRef]
- Pizarro-Cerdá, J.; Chorev, D.S.; Geiger, B.; Cossart, P. The Diverse Family of Arp2/3 Complexes. Trends Cell Biol. 2017, 27, 93–100. [Google Scholar] [CrossRef]
- Schwintzer, L.; Koch, N.; Ahuja, R.; Grimm, J.; Kessels, M.M.; Qualmann, B. The functions of the actin nucleator Cobl in cellular morphogenesis critically depend on syndapin I. EMBO J. 2011, 30, 3147–3159. [Google Scholar] [CrossRef] [PubMed]
- Haag, N.; Schwintzer, L.; Ahuja, R.; Koch, N.; Grimm, J.; Heuer, H.; Qualmann, B.; Kessels, M.M. The actin nucleator Cobl is crucial for Purkinje cell development and works in close conjunction with the F-actin binding protein Abp1. J. Neurosci. 2012, 32, 17842–17856. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Koch, D.; González Delgado, J.; Günther, M.; Witte, O.W.; Kessels, M.M.; Frahm, C.; Qualmann, B. Poststroke dendritic arbor regrowth requires the actin nucleator Cobl. PLoS Biol. 2021, 19, e3001399. [Google Scholar] [CrossRef]
- Hou, W.; Izadi, M.; Nemitz, S.; Haag, N.; Kessels, M.M.; Qualmann, B. The Actin Nucleator Cobl Is Controlled by Calcium and Calmodulin. PLoS Biol. 2015, 13, e1002233. [Google Scholar] [CrossRef]
- Izadi, M.; Seemann, E.; Schlobinski, D.; Schwintzer, L.; Qualmann, B.; Kessels, M.M. Functional interdependence of the actin nucleator Cobl and Cobl-like in dendritic arbor development. eLife 2021, 10, e67718. [Google Scholar] [CrossRef]
- Beer, A.J.; González Delgado, J.; Steiniger, F.; Qualmann, B.; Kessels, M.M. The actin nucleator Cobl organises the terminal web of enterocytes. Sci. Rep. 2020, 10, 11156. [Google Scholar] [CrossRef]
- Haag, N.; Schüler, S.; Nietzsche, S.; Hübner, C.A.; Strenzke, N.; Qualmann, B.; Kessels, M.M. The Actin Nucleator Cobl Is Critical for Centriolar Positioning, Postnatal Planar Cell Polarity Refinement, and Function of the Cochlea. Cell Rep. 2018, 24, 2418–2431.e6. [Google Scholar] [CrossRef]
- Hou, W.; Nemitz, S.; Schopper, S.; Nielsen, M.L.; Kessels, M.M.; Qualmann, B. Arginine Methylation by PRMT2 Controls the Functions of the Actin Nucleator Cobl. Dev. Cell 2018, 45, 262–275.e8. [Google Scholar] [CrossRef] [PubMed]
- Izadi, M.; Schlobinski, D.; Lahr, M.; Schwintzer, L.; Qualmann, B.; Kessels, M.M. Cobl-like promotes actin filament formation and dendritic branching using only a single WH2 domain. J. Cell Biol. 2018, 217, 211–230. [Google Scholar] [CrossRef]
- Karnovsky, M.J. A Formaldehyde-Glutaraldehyde Fixative of High Osmolality for Use in Electron Microscopy. J. Cell Biol. 1965, 27, 1A–149A. [Google Scholar]
- Schüler, S.; Hauptmann, J.; Perner, B.; Kessels, M.M.; Englert, C.; Qualmann, B. Ciliated sensory hair cell formation and function require the F-BAR protein syndapin I and the WH2 domain-based actin nucleator Cobl. J. Cell Sci. 2013, 126, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Lacruz, R.S.; Smith, C.E.; Bringas, P., Jr.; Chen, Y.B.; Smith, S.M.; Snead, M.L.; Kurtz, I.; Hacia, J.G.; Hubbard, M.J.; Paine, M.L. Identification of novel candidate genes involved in mineralization of dental enamel by genome-wide transcript profiling. J. Cell Physiol. 2012, 227, 2264–2275. [Google Scholar] [CrossRef] [PubMed]
- Simmer, J.P.; Richardson, A.S.; Wang, S.K.; Reid, B.M.; Bai, Y.; Hu, Y.; Hu, J.C. Ameloblast transcriptome changes from secretory to maturation stages. Connect. Tissue Res. 2014, 55, 29–32. [Google Scholar] [CrossRef]
- Kranz, S.; Heyder, M.; Mueller, S.; Guellmar, A.; Krafft, C.; Nietzsche, S.; Tschirpke, C.; Herold, V.; Sigusch, B.; Reise, M. Remineralization of Artificially Demineralized Human Enamel and Dentin Samples by Zinc-Carbonate Hydroxyapatite Nanocrystals. Materials 2022, 15, 7173. [Google Scholar] [CrossRef]
- Lacruz, R.S.; Habelitz, S.; Wright, J.T.; Paine, M.L. Dental enamel formation and implications for oral health and disease. Physiol. Rev. 2017, 97, 939–993. [Google Scholar] [CrossRef]
- Hu, Y.; Smith, C.E.; Richardson, A.S.; Bartlett, J.D.; Hu, J.C.; Simmer, J.P. MMP20, KLK4, and MMP20/KLK4 double null mice define roles for matrix proteases during dental enamel formation. Mol. Genet. Genom. Med. 2016, 4, 178–196. [Google Scholar] [CrossRef] [PubMed]
- Núñez, S.M.; Chun, Y.P.; Ganss, B.; Hu, Y.; Richardson, A.S.; Schmitz, J.E.; Fajardo, R.; Yang, J.; Hu, J.C.; Simmer, J.P. Maturation stage enamel malformations in Amtn and Klk4 null mice. Matrix Biol. 2016, 52–54, 219–233. [Google Scholar] [CrossRef]
- Crawford, P.J.; Aldred, M.; Bloch-Zupan, A. Amelogenesis imperfecta. Orphanet J. Rare Dis. 2007, 2, 17. [Google Scholar] [CrossRef]
- Li, Y.; Pugach, M.K.; Kuehl, M.A.; Peng, L.; Bouchard, J.; Hwang, S.Y.; Gibson, C.W. Dental enamel structure is altered by expression of dominant negative RhoA in ameloblasts. Cells Tissues Organs 2011, 194, 227–231. [Google Scholar] [CrossRef]
- Zheng, J.; Nie, X.; He, L.; Yoon, A.J.; Wu, L.; Zhang, X.; Vats, M.; Schiff, M.D.; Xiang, L.; Tian, Z.; et al. Epithelial Cdc42 Deletion Induced Enamel Organ Defects and Cystogenesis. J. Dent. Res. 2018, 97, 1346–1354. [Google Scholar] [CrossRef]
- Husson, C.; Renault, L.; Didry, D.; Pantaloni, D.; Carlier, M.F. Cordon-Bleu uses WH2 domains as multifunctional dynamizers of actin filament assembly. Mol. Cell 2011, 43, 464–477. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Walker, M.; Trinick, J.; Pernier, J.; Montaville, P.; Carlier, M.F. Mutagenetic and electron microscopy analysis of actin filament severing by Cordon-Bleu, a WH2 domain protein. Cytoskeleton 2014, 71, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Tsukita, K.; Kitamata, M.; Kashihara, H.; Yano, T.; Fujiwara, I.; Day, T.F.; Katsuno, T.; Kim, J.; Takenaga, F.; Tanaka, H.; et al. Phase separation of an actin nucleator by junctional microtubules regulates epithelial function. Sci. Adv. 2023, 9, eadf6358. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.; Terhoeven, N. The bilaterian roots of cordon-bleu. BMC Res. Notes 2013, 6, 393. [Google Scholar] [CrossRef]
- Chen, X.; Ni, F.; Tian, X.; Kondrashkina, E.; Wang, Q.; Ma, J. Structural basis of actin filament nucleation by tandem W domains. Cell Rep. 2013, 3, 1910–1920. [Google Scholar] [CrossRef]
- Zuchero, J.B.; Coutts, A.S.; Quinlan, M.E.; Thangue, N.B.; Mullins, R.D. p53-cofactor JMY is a multifunctional actin nucleation factor. Nat. Cell Biol. 2009, 11, 451–459. [Google Scholar] [CrossRef]
- Quinlan, M.E.; Heuser, J.E.; Kerkhoff, E.; Mullins, R.D. Drosophila Spire is an actin nucleation factor. Nature 2005, 433, 382–388. [Google Scholar] [CrossRef]
- Mahuzier, A.; Shihavuddin, A.; Fournier, C.; Lansade, P.; Faucourt, M.; Menezes, N.; Meunier, A.; Garfa-Traoré, M.; Carlier, M.F.; Voituriez, R.; et al. Ependymal cilia beating induces an actin network to protect centrioles against shear stress. Nat. Commun 2018, 9, 2279. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janitzek, H.; González Delgado, J.; Haag, N.; Seemann, E.; Nietzsche, S.; Sigusch, B.; Qualmann, B.; Kessels, M.M. The Evolutionary Young Actin Nucleator Cobl Is Important for Proper Amelogenesis. Cells 2025, 14, 359. https://doi.org/10.3390/cells14050359
Janitzek H, González Delgado J, Haag N, Seemann E, Nietzsche S, Sigusch B, Qualmann B, Kessels MM. The Evolutionary Young Actin Nucleator Cobl Is Important for Proper Amelogenesis. Cells. 2025; 14(5):359. https://doi.org/10.3390/cells14050359
Chicago/Turabian StyleJanitzek, Hannes, Jule González Delgado, Natja Haag, Eric Seemann, Sandor Nietzsche, Bernd Sigusch, Britta Qualmann, and Michael Manfred Kessels. 2025. "The Evolutionary Young Actin Nucleator Cobl Is Important for Proper Amelogenesis" Cells 14, no. 5: 359. https://doi.org/10.3390/cells14050359
APA StyleJanitzek, H., González Delgado, J., Haag, N., Seemann, E., Nietzsche, S., Sigusch, B., Qualmann, B., & Kessels, M. M. (2025). The Evolutionary Young Actin Nucleator Cobl Is Important for Proper Amelogenesis. Cells, 14(5), 359. https://doi.org/10.3390/cells14050359