
PTPA Governs Stress-Responsive Differentiation and Metabolic Homeostasis in Toxoplasma gondii

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Parasite Strain, Culture Conditions, and Electroporation
2.2. Endogenous Tagging
2.3. Indirect Immunofluorescence Assay
2.4. Proliferation Assay
2.5. Invasion Assay
2.6. Egress Assay
2.7. Plaque Assay
2.8. In Vitro Differentiation Assay
2.9. PTPA Protein 3D Structure Prediction, Phylogenetic Analysis, and Molecular Docking
2.10. Transmission Electron Microscopy (TEM) of Toxoplasma Ultrastructure
2.11. Starch Granule Staining and Analysis
2.12. PTPA Protein Expression and Pull-Down
2.13. Proteomic Profiling and Analysis
3. Results
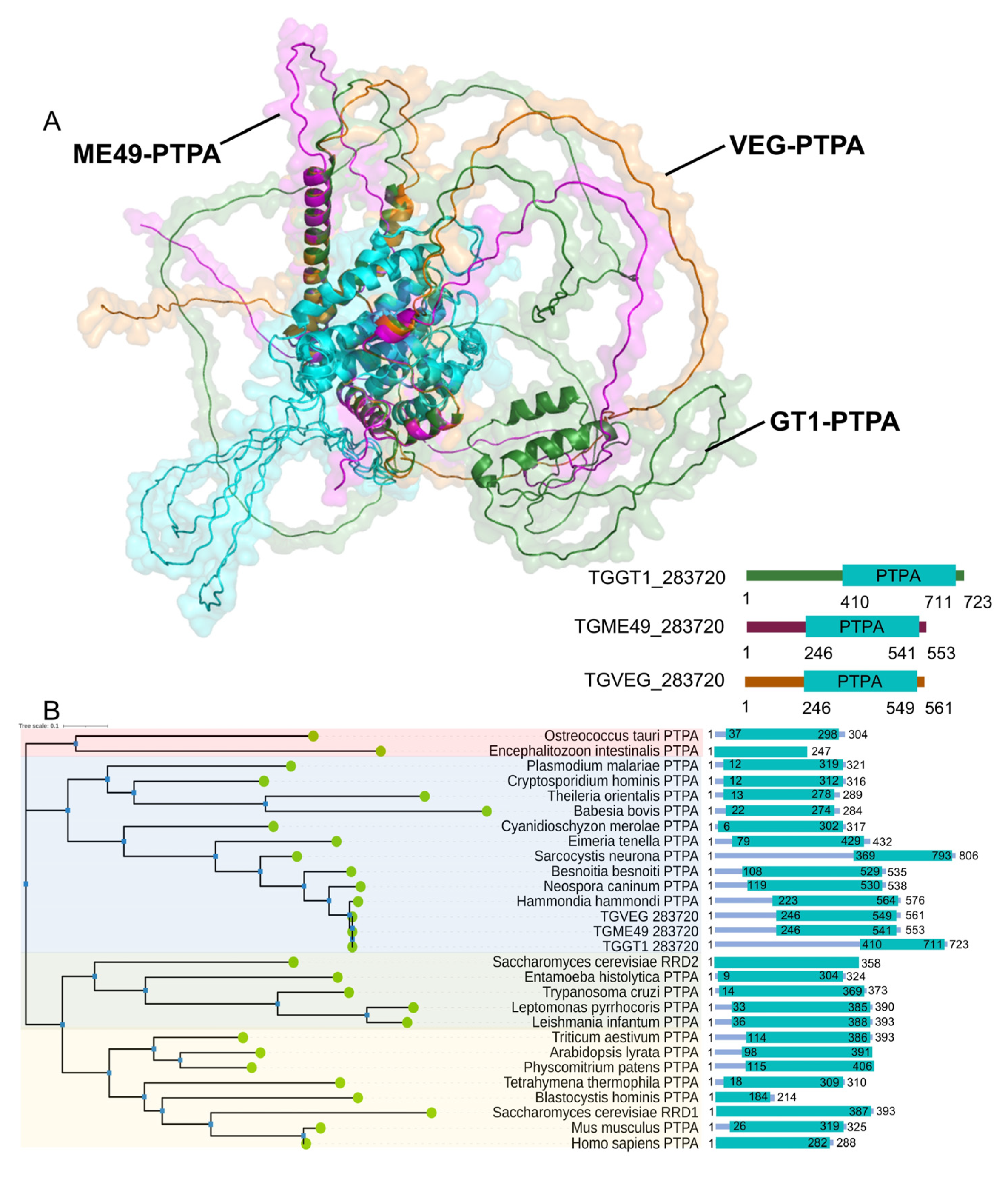
3.1. Toxoplasma Expresses a Homolog of Phosphotyrosyl Phosphatase Activator Protein
3.2. Interaction of PTPA Protein with PP2A A and C Subunits in Toxoplasma gondii
3.3. Conditional Depletion of PTPA Protein Impairs the Lytic Cycle of Toxoplasma gondii
3.4. PTPA Depletion Impairs Tachyzoite-to-Bradyzoite Conversion in Toxoplasma gondii
3.5. PTPA Depletion Triggers Metabolic Dysregulation and Ultrastructural Disintegration in Toxoplasma gondii
3.6. Stage-Specific Proteomic Reveal PTPA’s Role in Toxoplasma gondii Survival and Development
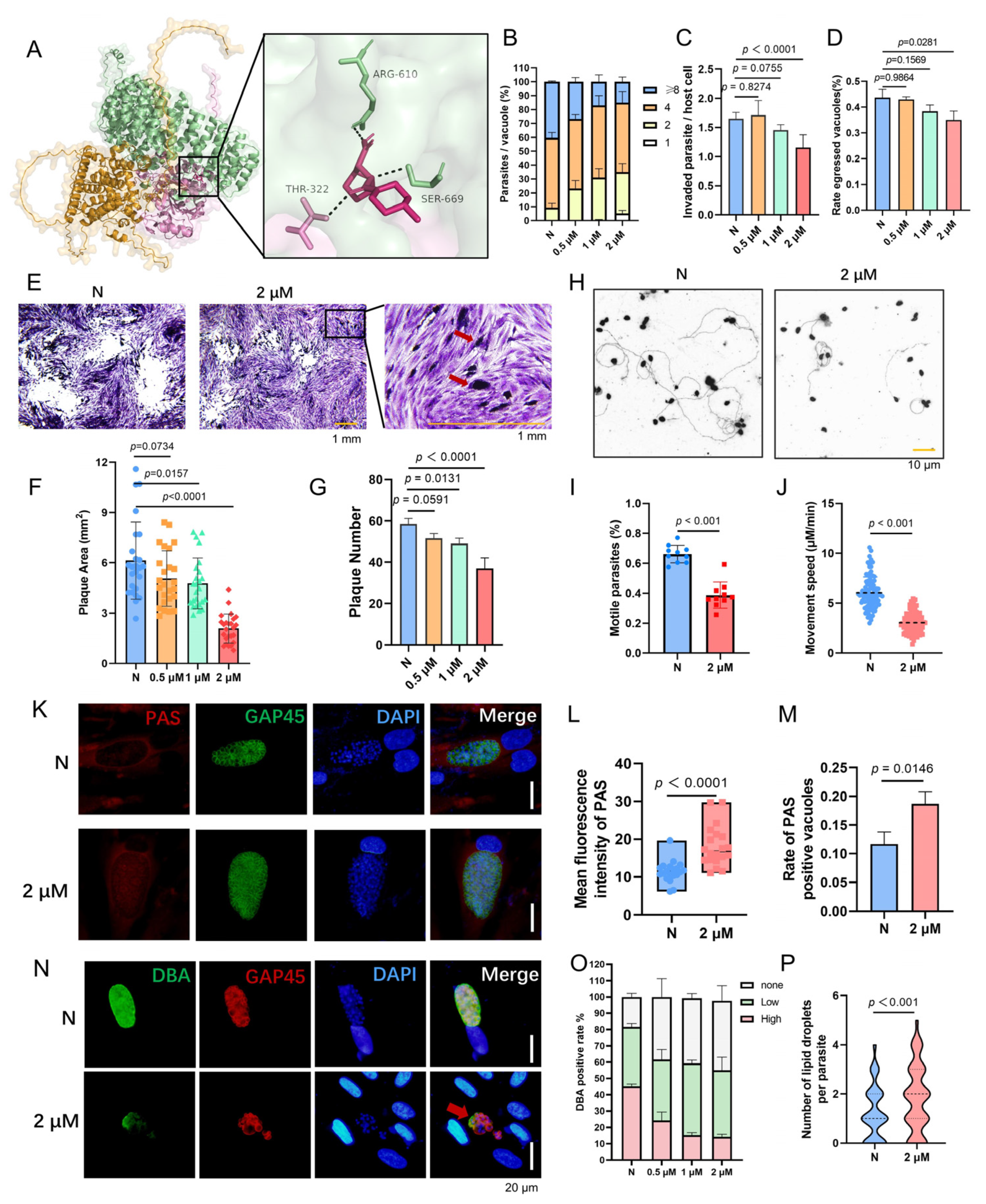
3.7. LB-100 Targets PTPA–PP2A Axis to Block Toxoplasma gondii
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lourido, S. Toxoplasma gondii . Trends Parasitol. 2019, 35, 944–945. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, W.J., Jr.; Jeffers, V. Mechanisms of Toxoplasma gondii persistence and latency. FEMS Microbiol. Rev. 2012, 36, 717–733. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Lindsay, D.S.; Speer, C.A. Structures of Toxoplasma gondii tachyzoites, bradyzoites, and sporozoites and biology and development of tissue cysts. Clin. Microbiol. Rev. 1998, 11, 267–299. [Google Scholar] [CrossRef] [PubMed]
- Soête, M.; Camus, D.; Dubremetz, J.F. Experimental induction of bradyzoite-specific antigen expression and cyst formation by the RH strain of Toxoplasma gondii in vitro. Exp. Parasitol. 1994, 78, 361–370. [Google Scholar] [CrossRef]
- Fox, B.A.; Gigley, J.P.; Bzik, D.J. Toxoplasma gondii lacks the enzymes required for de novo arginine biosynthesis and arginine starvation triggers cyst formation. Int. J. Parasitol. 2004, 34, 323–331. [Google Scholar] [CrossRef]
- Narasimhan, J.; Joyce, B.R.; Naguleswaran, A.; Smith, A.T.; Livingston, M.R.; Dixon, S.E.; Coppens, I.; Wek, R.C.; Sullivan, W.J., Jr. Translation regulation by eukaryotic initiation factor-2 kinases in the development of latent cysts in Toxoplasma gondii. J. Biol. Chem. 2008, 283, 16591–16601. [Google Scholar] [CrossRef]
- Tomavo, S.; Boothroyd, J.C. Interconnection between organellar functions, development and drug resistance in the protozoan parasite, Toxoplasma gondii. Int. J. Parasitol. 1995, 25, 1293–1299. [Google Scholar] [CrossRef]
- Ying, Z.; Yin, M.; Zhu, Z.; Shang, Z.; Pei, Y.; Liu, J.; Liu, Q. Iron stress affects the growth and differentiation of Toxoplasma gondii. Int. J. Mol. Sci. 2024, 25, 2493. [Google Scholar] [CrossRef]
- Chao, Y.; Xing, Y.; Chen, Y.; Xu, Y.; Lin, Z.; Li, Z.; Jeffrey, P.D.; Stock, J.B.; Shi, Y. Structure and mechanism of the phosphotyrosyl phosphatase activator. Mol. Cell 2006, 23, 535–546. [Google Scholar] [CrossRef]
- Li, M.; Damuni, Z. Okadaic acid and microcystin-LR directly inhibit the methylation of protein phosphatase 2A by its specific methyltransferase. Biochem. Biophys. Res. Commun. 1994, 202, 1023–1030. [Google Scholar] [CrossRef]
- Van Hoof, C.; Cayla, X.; Bosch, M.; Merlevede, W.; Goris, J. The phosphotyrosyl phosphatase activator of protein phosphatase 2A. A novel purification method, immunological and enzymic characterization. Eur. J. Biochem. 1994, 226, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Jordens, J.; Janssens, V.; Longin, S.; Stevens, I.; Martens, E.; Bultynck, G.; Engelborghs, Y.; Lescrinier, E.; Waelkens, E.; Goris, J.; et al. The protein phosphatase 2A phosphatase activator is a novel peptidyl-prolyl cis/trans-isomerase. J. Biol. Chem. 2006, 281, 6349–6357. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Stanevich, V.; Wlodarchak, N.; Sengupta, R.; Jiang, L.; Satyshur, K.A.; Xing, Y. Structural basis of PP2A activation by PTPA, an ATP-dependent activation chaperone. Cell Res. 2014, 24, 190–203. [Google Scholar] [CrossRef] [PubMed]
- Fellner, T.; Lackner, D.H.; Hombauer, H.; Piribauer, P.; Mudrak, I.; Zaragoza, K.; Juno, C.; Ogris, E. A novel and essential mechanism determining specificity and activity of protein phosphatase 2A (PP2A) in vivo. Genes Dev. 2003, 17, 2138–2150. [Google Scholar] [CrossRef]
- Luo, D.J.; Feng, Q.; Wang, Z.H.; Sun, D.S.; Wang, Q.; Wang, J.Z.; Liu, G.P. Knockdown of phosphotyrosyl phosphatase activator induces apoptosis via mitochondrial pathway and the attenuation by simultaneous tau hyperphosphorylation. J. Neurochem. 2014, 130, 816–825. [Google Scholar] [CrossRef]
- Vandomme, A.; Fréville, A.; Cailliau, K.; Kalamou, H.; Bodart, J.F.; Khalife, J.; Pierrot, C. Phosphotyrosyl phosphatase activator of Plasmodium falciparum: Identification of its residues involved in binding to and activation of PP2A. Int. J. Mol. Sci. 2014, 15, 2431–2453. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, W.; Ehsan, M.; Zhang, Y.; Yan, R.; Song, X.; Xu, L.; Zhang, X.; Li, X. Characterization of a phosphotyrosyl phosphatase activator homologue of the parasitic nematode Haemonchus contortus and its immunomodulatory effect on goat peripheral blood mononuclear cells in vitro. Int. J. Parasitol. 2020, 50, 1157–1166. [Google Scholar] [CrossRef]
- Wang, J.L.; Li, T.T.; Elsheikha, H.M.; Liang, Q.L.; Zhang, Z.W.; Wang, M.; Sibley, L.D.; Zhu, X.Q. The protein phosphatase 2A holoenzyme is a key regulator of starch metabolism and bradyzoite differentiation in Toxoplasma gondii. Nat. Commun. 2022, 13, 7560. [Google Scholar] [CrossRef]
- Li, S.; Liu, J.; Zhang, H.; Sun, Z.; Ying, Z.; Wu, Y.; Xu, J.; Liu, Q. Toxoplasma gondii glutathione S-transferase 2 plays an important role in partial secretory protein transport. FASEB J. 2021, 35, e21352. [Google Scholar] [CrossRef]
- Fu, Y.; Brown, K.M.; Jones, N.G.; Moreno, S.N.; Sibley, L.D. Toxoplasma bradyzoites exhibit physiological plasticity of calcium and energy stores controlling motility and egress. eLife 2021, 10, e73011. [Google Scholar] [CrossRef]
- Abramson, J.; Adler, J.; Dunger, J.; Evans, R.; Green, T.; Pritzel, A.; Ronneberger, O.; Willmore, L.; Ballard, A.J.; Bambrick, J.; et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 2024, 630, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, X.; Gan, J.; Chen, S.; Xiao, Z.X.; Cao, Y. CB-Dock2: Improved protein-ligand blind docking by integrating cavity detection, docking and homologous template fitting. Nucleic Acids Res. 2022, 50, W159–W164. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Liu, Y.; Gan, J.; Xiao, Z.X.; Cao, Y. FitDock: Protein-ligand docking by template fitting. Brief. Bioinform. 2022, 23, bbac087. [Google Scholar] [CrossRef] [PubMed]
- Lyu, F.; Han, F.; Ge, C.; Mao, W.; Chen, L.; Hu, H.; Chen, G.; Lang, Q.; Fang, C. OmicStudio: A composable bioinformatics cloud platform with real-time feedback that can generate high-quality graphs for publication. iMeta 2023, 2, e85. [Google Scholar] [CrossRef]
- Chen, J.; Hu, R.; Zhu, Y.; Shen, G.; Zhang, H. Arabidopsis Phosphotyrosyl phosphatase activator is essential for protein phosphatase 2A holoenzyme assembly and plays important roles in hormone signaling, salt stress response, and plant development. Plant Physiol. 2014, 166, 1519–1534. [Google Scholar] [CrossRef]
- Van Hoof, C.; Janssens, V.; Dinishiotu, A.; Merlevede, W.; Goris, J. Functional analysis of conserved domains in the phosphotyrosyl phosphatase activator. Molecular cloning of the homologues from Drosophila melanogaster and Saccharomyces cerevisiae. Biochemistry. 1998, 37, 12899–12908. [Google Scholar] [CrossRef]
- Uboldi, A.D.; McCoy, J.M.; Blume, M.; Gerlic, M.; Ferguson, D.J.; Dagley, L.F.; Beahan, C.T.; Stapleton, D.I.; Gooley, P.R.; Bacic, A.; et al. Regulation of starch stores by a Ca2+-dependent protein kinase is essential for viable cyst development in Toxoplasma gondii. Cell Host Microbe. 2015, 18, 670–681. [Google Scholar] [CrossRef]
- Sugi, T.; Tu, V.; Ma, Y.; Tomita, T.; Weiss, L.M. Toxoplasma gondii requires glycogen phosphorylase for balancing amylopectin storage and for efficient production of brain cysts. mBio 2017, 8, e01289-17. [Google Scholar] [CrossRef]
- Renaud, E.A.; Maupin, A.J.M.; Bordat, Y.; Graindorge, A.; Berry, L.; Besteiro, S. Iron depletion has different consequences on the growth and survival of Toxoplasma gondii strains. Virulence 2024, 15, 2329566. [Google Scholar] [CrossRef]
- Fan, F.; Xue, L.; Yin, X.; Gupta, N.; Shen, B. AP2XII-1 is a negative regulator of merogony and presexual commitment in Toxoplasma gondii. mBio 2023, 14, e0178523. [Google Scholar] [CrossRef]
- Wang, J.L.; Li, T.T.; Zhang, N.Z.; Wang, M.; Sun, L.X.; Zhang, Z.W.; Fu, B.Q.; Elsheikha, H.M.; Zhu, X.Q. The transcription factor AP2XI-2 is a key negative regulator of Toxoplasma gondii merogony. Nat. Commun. 2024, 15, 793. [Google Scholar] [CrossRef] [PubMed]
- Mazhar, S.; Taylor, S.E.; Sangodkar, J.; Narla, G. Targeting PP2A in cancer: Combination therapies. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Swingle, M.; Ni, L.; Honkanen, R.E. Small-molecule inhibitors of ser/thr protein phosphatases: Specificity, use and common forms of abuse. Methods Mol. Biol. 2007, 365, 23–38. [Google Scholar] [PubMed]
- Ho, W.S.; Feldman, M.J.; Maric, D.; Amable, L.; Hall, M.D.; Feldman, G.M.; Ray-Chaudhury, A.; Lizak, M.J.; Vera, J.C.; Robison, R.A.; et al. PP2A inhibition with LB100 enhances cisplatin cytotoxicity and overcomes cisplatin resistance in medulloblastoma cells. Oncotarget 2016, 7, 12447–12463. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Yang, Y.; Shi, Y.; Chen, X.; Yang, Y.; Pan, L.; Du, Z.; Sun, H.; Yao, C.; Ma, G.; et al. PP2Acα-B′/PR61 Holoenzyme of Toxoplasma gondii is required for the amylopectin metabolism and proliferation of tachyzoites. Microbiol. Spectr. 2023, 11, e0010423. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ying, Z.; Wu, Y.; Pei, Y.; Shang, Z.; Liu, J.; Liu, Q. PTPA Governs Stress-Responsive Differentiation and Metabolic Homeostasis in Toxoplasma gondii. Cells 2025, 14, 835. https://doi.org/10.3390/cells14110835
Ying Z, Wu Y, Pei Y, Shang Z, Liu J, Liu Q. PTPA Governs Stress-Responsive Differentiation and Metabolic Homeostasis in Toxoplasma gondii. Cells. 2025; 14(11):835. https://doi.org/10.3390/cells14110835
Chicago/Turabian StyleYing, Zhu, Yuntong Wu, Yanqun Pei, Zheng Shang, Jing Liu, and Qun Liu. 2025. "PTPA Governs Stress-Responsive Differentiation and Metabolic Homeostasis in Toxoplasma gondii" Cells 14, no. 11: 835. https://doi.org/10.3390/cells14110835
APA StyleYing, Z., Wu, Y., Pei, Y., Shang, Z., Liu, J., & Liu, Q. (2025). PTPA Governs Stress-Responsive Differentiation and Metabolic Homeostasis in Toxoplasma gondii. Cells, 14(11), 835. https://doi.org/10.3390/cells14110835