
The Influence of the ACTN3 R577X Genotype on Performance in Brazilian National-Level Decathlon Athletes: A Pilot Study


 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Sampling and Genotyping
2.3. Statistics
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dias, R.G.; Pereira, A.D.C.; Negrão, C.E.; Krieger, J.E. Genetic polymorphisms determining of the physical performance in elite athletes. Rev. Bras. Med. Esporte 2007, 13, 209–216. [Google Scholar] [CrossRef]
- Blanchard, A.; Ohanian, V.; Critchley, D. The structure and function of α-actinin. J. Muscle Res. Cell Motil. 1989, 10, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Pasqua, L.A.; Artioli, G.G.; Pires, F.O.; Bertuzzi, R. ACTN 3 e desempenho esportivo: Um gene candidato ao sucesso em provas de curta e longa duração. Rev. Bras. Cineantropom. Desempenho Hum. 2011, 13, 477–483. [Google Scholar] [CrossRef]
- Dias, R.G. Genetics, human physical performance and gene doping: The common sense versus the scientific reality. Rev. Bras. Med. Esporte 2011, 17, 62–70. [Google Scholar] [CrossRef]
- Mills, M.A.; Yang, N.; Weinberger, R.; Vander Woude, D.L.; Beggs, A.H.; Easteal, S.; North, K.N. Differential expression for the actin-binding proteins, α-actinin-2 and -3, in different species: Implications for the evolution of functional redundancy. Hum. Mol. Genet. 2001, 10, 1335–1346. [Google Scholar] [CrossRef]
- Yang, N.; MacArthur, D.G.; Gulbin, J.P.; Hahn, A.G.; Beggs, A.H.; Easteal, S.; North, K. ACTN3 genotype is associated with human elite athletic performance. Am. J. Hum. Genet. 2003, 73, 627–631. [Google Scholar] [CrossRef]
- Chae, J.H.; Eom, S.-H.; Lee, S.-K.; Jung, J.-H.; Kim, C.-H. Association between Complex ACTN3 and ACE Gene Polymorphisms and Elite Endurance Sports in Koreans: A Case–Control Study. Genes 2024, 15, 1110. [Google Scholar] [CrossRef]
- Niemi, A.K.; Majamaa, K. Mitochondrial DNA and ACTN3 genotypes in Finnish elite endurance and sprint athletes. Eur. J. Hum. Genet. 2005, 13, 965–969. [Google Scholar] [CrossRef]
- Norman, B.; Esbjörnsson, M.; Rundqvist, H.; Österlund, T.; Von Walden, F.; Tesch, P.A. Strength, power, fiber types, and mRNA expression in trained men and women with different ACTN3 R577X genotypes. J. Appl. Physiol. 2009, 106, 959–965. [Google Scholar] [CrossRef]
- Walsh, P.S.; Metzger, D.A.; Higuchi, R. Chelex® 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material. Biotechniques 1991, 10, 506–513. [Google Scholar] [CrossRef]
- Habibi, A.; Shabani, M.; Rahimi, E.; Fatemi, R.; Najafi, A.; Analoei, H.; Hosseini, M. Relationship between jump test results and acceleration phase of sprint performance in national and regional 100 m sprinters. J. Hum. Kinet. 2010, 23, 29–35. [Google Scholar] [CrossRef]
- Marques, M.C.; González-Badillo, J.J. Relationship between strength parameters and squat jump performance in trained athletes. Motricidade 2011, 7, 43–48. [Google Scholar]
- Vescovi, J.D.; McGuigan, M.R. Relationships between sprinting, agility, and jump ability in female athletes. J. Sports Sci. 2008, 26, 97–107. [Google Scholar] [CrossRef]
- Druzhevskaya, A.M.; Ahmetov, I.I.; Astratenkova, I.V.; Rogozkin, V.A. Association of the ACTN3 R577X polymorphism with power athlete status in Russians. Eur. J. Appl. Physiol. 2008, 103, 631–634. [Google Scholar] [CrossRef]
- Papadimitriou, D.T.; Marakaki, C.; Fretzayas, A.; Nicolaidou, P.; Papadimitriou, A. Negativation of type 1 diabetes-associated autoantibodies to glutamic acid decarboxylase and insulin in children treated with oral calcitriol. J. Diabetes 2013, 5, 344–348. [Google Scholar] [CrossRef]
- Moran, C.N.; Yang, N.; Bailey, M.E.S.; Tsiokanos, A.; Jamurtas, A.; MacArthur, D.G.; North, K.; Pitsiladis, Y.P.; Wilson, R.H. Association analysis of the ACTN3 R577X polymorphism and complex quantitative body composition and performance phenotypes in adolescent Greeks. Eur. J. Hum. Genet. 2007, 15, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Cieszczyk, P.; Eider, J.; Ostanek, M.; Arczewska, A.; Leońska-Duniec, A.; Sawczyn, S.; Ficek, K.; Krupecki, K. Association of the ACTN3 R577X polymorphism in Polish power-orientated athletes. J. Hum. Kinet. 2011, 28, 55–61. [Google Scholar] [CrossRef]
- Rodríguez-Romo, G.; Yvert, T.; de Diego, A.; Santiago, C.; Díaz de Durana, A.L.; Carratalá, V.; Garatachea, N.; Lucia, A. No Association Between ACTN3 R577X Polymorphism and Elite Judo Athletic Status. Int. J. Sports Physiol. Perform. 2013, 8, 579–581. [Google Scholar] [CrossRef]
- Harrison, A.J.; Keane, S.P.; Coglan, J. Force-velocity relationship and stretch-shortening cycle function in sprint and endurance athletes. J. Strength Cond. Res. 2004, 18, 473–479. [Google Scholar]
- Paulo, B.; Paixão, J.; Reis, V.M.M.; Jorge, P.; Paixão, M. 400 M Performance. Methods 2004, 39–45. [Google Scholar]
- MacArthur, D.G.; North, K.N. A gene for speed? The evolution and function of α-actinin-3. BioEssays 2004, 26, 786–795. [Google Scholar] [CrossRef] [PubMed]
- Vincent, B.; De Bock, K.; Ramaekers, M.; Van Den Eede, E.; Van Leemputte, M.; Hespel, P.; Thomis, M.A. ACTN3 (R577X) genotype is associated with fiber type distribution. Physiol. Genom. 2007, 32, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Saunders, C.J.; September, A.V.; Xenophontos, S.L.; Cariolou, M.A.; Anastassiades, L.C.; Noakes, T.D.; Collins, M. No association of the ACTN3 gene R577X polymorphism with endurance performance in Ironman Triathlons. Ann. Hum. Genet. 2007, 71, 777–781. [Google Scholar] [CrossRef] [PubMed]
- Pimenta, E.M.; Coelho, D.B.; Cruz, I.R.; Morandi, R.F.; Veneroso, C.E.; Pussieldi, G.A.; Carvalho, M.R.S.; Silami-Garcia, E.; De Paz Fernández, J.A. The ACTN3 genotype in soccer players in response to acute eccentric training. Eur. J. Appl. Physiol. 2012, 112, 1495–1503. [Google Scholar] [CrossRef]

{kind=link}
| 100 m Race | Long Jump | Shot Put | High Jump | 400 m Race | |||||
|---|---|---|---|---|---|---|---|---|---|
| RES-s | Points | RES-m | Points | RES-m | Points | RES-s | Points | RES-s | Points |
| 10.50 | 975 | 7.39 | 908 | 14.02 | 730 | 1.99 | 794 | 49.67 | 830 |
| 10.51 | 973 | 7.38 | 905 | 14.01 | 729 | 1.98 | 785 | 49.69 | 829 |
| 10.52 | 970 | 7.37 | 903 | 13.99 | 728 | 1.97 | 776 | 49.72 | 828 |
| 10.53 | 968 | 7.36 | 900 | 13.97 | 727 | 1.96 | 767 | 49.74 | 827 |
| 10.54 | 966 | 7.35 | 898 | 13.96 | 726 | 1.95 | 758 | 49.76 | 826 |
| 10.55 | 963 | 7.34 | 896 | 13.94 | 725 | 1.94 | 749 | 49.78 | 825 |
| 10.56 | 961 | 7.33 | 893 | 13.93 | 724 | 1.93 | 740 | 49.80 | 824 |
| 10.57 | 959 | 7.32 | 891 | 13.91 | 723 | 1.92 | 731 | 49.82 | 823 |
| 10.58 | 956 | 7.31 | 888 | 13.89 | 722 | 1.91 | 723 | 49.85 | 822 |
| 10.59 | 954 | 7.30 | 886 | 13.88 | 721 | 1.90 | 714 | 49.87 | 821 |
| 110 w/Obstacle | Disc Launch | Pole Jump | Javelin Throw | 1500 m Race | |||||
|---|---|---|---|---|---|---|---|---|---|
| RES-s | Points | RES-m | Points | RES-m | Points | RES-m | Points | RES-min | Points |
| 14.20 | 949 | 44.16 | 750 | 4.79 | 846 | 60.12 | 740 | 4:39.33 | 685 |
| 14.21 | 948 | 44.11 | 749 | 4.78 | 843 | 60.05 | 739 | 4:39.49 | 684 |
| 14.22 | 946 | 44.06 | 748 | 4.77 | 840 | 59.98 | 738 | 4:39.65 | 683 |
| 14.23 | 945 | 44.02 | 747 | 4.76 | 837 | 59.92 | 737 | 4:39.80 | 682 |
| 14.24 | 944 | 43.97 | 746 | 4.75 | 834 | 59.85 | 736 | 4:39.96 | 681 |
| 14.25 | 942 | 43.92 | 745 | 4.74 | 831 | 59.78 | 735 | 4:40.12 | 680 |
| 14.26 | 941 | 43.87 | 744 | 4.73 | 828 | 59.72 | 734 | 4:40.28 | 679 |
| 14.27 | 940 | 43.82 | 743 | 4.72 | 825 | 59.65 | 733 | 4:40.44 | 678 |
| 14.28 | 939 | 43.77 | 742 | 4.71 | 822 | 59.58 | 732 | 4:40.60 | 677 |
| 14.29 | 937 | 43.72 | 741 | 4.70 | 819 | 59.52 | 731 | 4:40.76 | 676 |
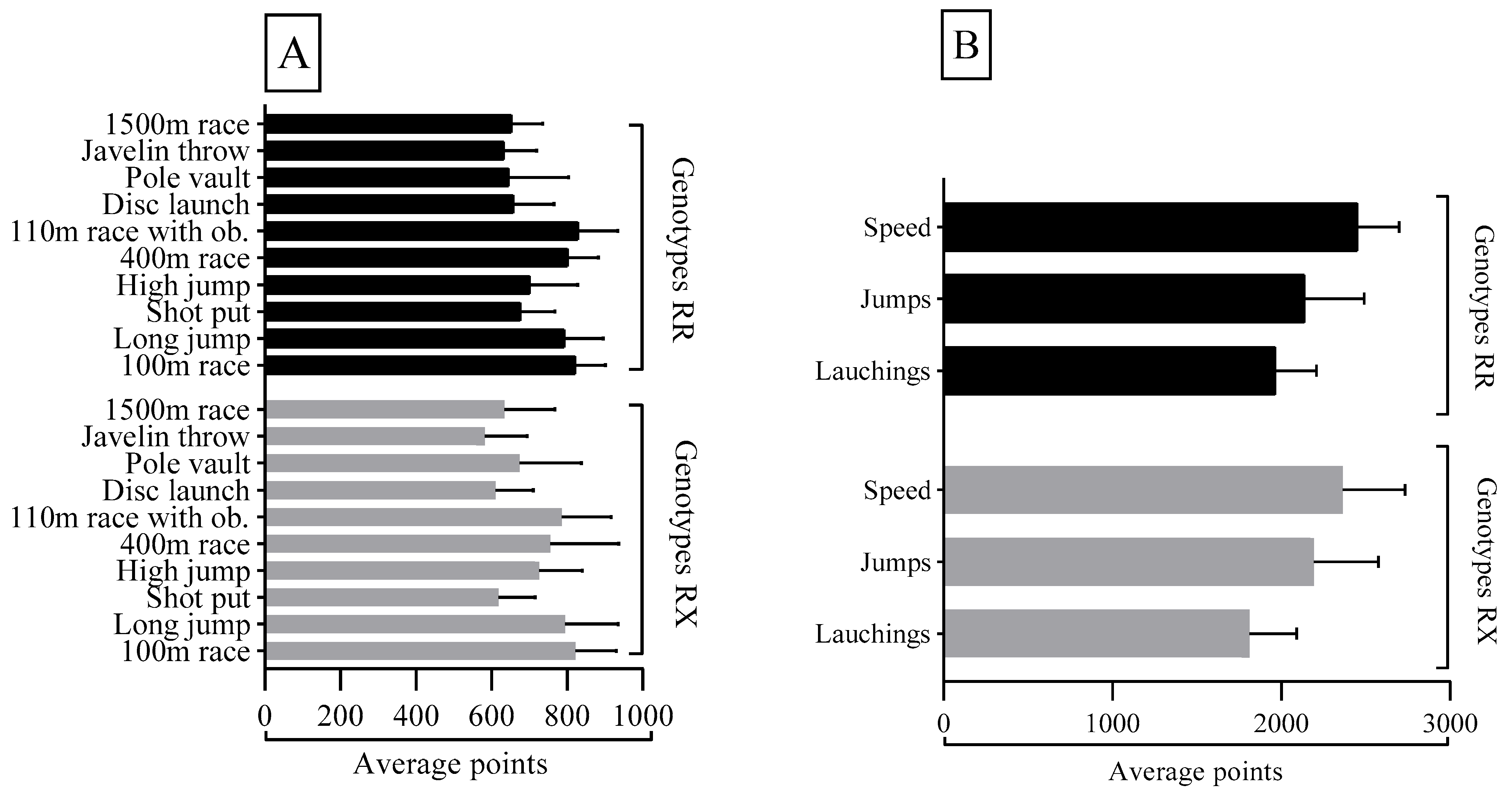
| Set of Events | Sum of Points | Average Points | Fitness Skills |
|---|---|---|---|
| Speed | 2.418 | 806 | Explosive strength, displacement speed, agility |
| Jumps | 2.188 | 729 | Travel speed, explosive force |
| Throwing and Launching | 1.932 | 644 | Maximum strength, explosive strength |
| Medium Distance—1500 m | -- | 633 | Aerobic power |
| Tests | R. 100 m | L.J | S. Put | H.J | C. 400 m | R.110 m w/H | T. of Disc | Pole Jump | T. of the dart | R. 1500 m | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 100 m race | r | 1 | 0.727 ** | 0.375 * | 0.526 ** | 0.759 ** | 0.625 ** | 0.336 | 0.428 * | 0.292 | 0.463 ** |
| p | 0 | 0.038 | 0.002 | 0 | 0 | 0.064 | 0.016 | 0.111 | 0.009 | ||
| 400 m race | r | 0.759 ** | 0.651 ** | 0.581 ** | 0.583 ** | 1 | 0.640 ** | 0.489 ** | 0.612 ** | 0.500 ** | 0.285 |
| p | 0 | 0 | 0.001 | 0.001 | 0 | 0.005 | 0 | 0.004 | 0.12 | ||
| Long jump | r | 0.727 ** | 1 | 0.558 ** | 0.789 ** | 0.651 ** | 0.829 ** | 0.580 ** | 0.623 ** | 0.385 * | 0.328 |
| p | 0 | 0.001 | 0 | 0 | 0 | 0.001 | 0 | 0.033 | 0.071 | ||
| Shot put | r | 0.375 * | 0.558 ** | 1 | 0.583 ** | 0.581 ** | 0.651 ** | 0.736 ** | 0.699 ** | 0.640 ** | 0.035 |
| p | 0.038 | 0.001 | 0.001 | 0.001 | 0 | 0 | 0 | 0 | 0.853 | ||
| High jump | r | 0.526 ** | 0.789 ** | 0.583 ** | 1 | 0.583 ** | 0.745 ** | 0.712 ** | 0.790 ** | 0.397 * | 0.04 |
| p | 0.002 | 0 | 0.001 | 0.001 | 0 | 0 | 0 | 0.027 | 0.832 | ||
| 110 m race with ob. | r | 0.625 ** | 0.829 ** | 0.651 ** | 0.745 ** | 0.640 ** | 1 | 0.675 ** | 0.607 ** | 0.433 * | 0.309 |
| p | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.015 | 0.09 | ||
| Disc launch | r | 0.336 | 0.580 ** | 0.736 ** | 0.712 ** | 0.489 ** | 0.675 ** | 1 | 0.805 ** | 0.608 ** | −0.155 |
| p | 0.064 | 0.001 | 0 | 0 | 0.005 | 0 | 0 | 0 | 0.406 | ||
| Pole jump | r | 0.428 * | 0.623 ** | 0.699 ** | 0.790 ** | 0.612 ** | 0.607 ** | 0.805 ** | 1 | 0.546 ** | 0.037 |
| p | 0.016 | 0 | 0 | 0 | 0 | 0 | 0 | 0.001 | 0.842 | ||
| Javelin throw | r | 0.292 | 0.385 * | 0.640 ** | 0.397 * | 0.500 ** | 0.433 * | 0.608 ** | 0.546 ** | 1 | −0.129 |
| p | 0.111 | 0.033 | 0 | 0.027 | 0.004 | 0.015 | 0 | 0.001 | 0.49 | ||
| 1500 m race | r | 0.463 ** | 0.328 | 0.035 | 0.04 | 0.285 | 0.309 | −0.155 | 0.037 | −0.129 | 1 |
| p | 0.009 | 0.071 | 0.853 | 0.832 | 0.12 | 0.09 | 0.406 | 0.842 | 0.49 | ||
| Set of Events | Variable | Speed | Jumps | Launches |
|---|---|---|---|---|
| Speed | r | 1 | 0.786 | 0.634 |
| p-value | <0.01 | <0.01 | ||
| N | 31 | 31 | 31 | |
| Jumps | r | 0.786 | 1 | 0.747 |
| p-value | <0.01 | <0.01 | ||
| N | 31 | 31 | 31 | |
| Launching and Throws | r | 0.634 | 0.747 | 1 |
| p-value | <0.01 | <0.01 | ||
| N | 31 | 31 | 31 |
| Genotype Frequency | Allele Frequency | |||
|---|---|---|---|---|
| R577R (%) | R577X (%) | X577X (%) | 577R (%) | 577X (%) |
| 16 (51.6%) | 15 (48.4%) | 0 (0.0%) | 76 | 24 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nunes, J.R.d.A.; Ceylan, H.I.; de Almeida-Neto, P.F.; Murawska-Ciałowicz, E.; Bragazzi, N.L.; de Assis, G.G. The Influence of the ACTN3 R577X Genotype on Performance in Brazilian National-Level Decathlon Athletes: A Pilot Study. Cells 2025, 14, 782. https://doi.org/10.3390/cells14110782
Nunes JRdA, Ceylan HI, de Almeida-Neto PF, Murawska-Ciałowicz E, Bragazzi NL, de Assis GG. The Influence of the ACTN3 R577X Genotype on Performance in Brazilian National-Level Decathlon Athletes: A Pilot Study. Cells. 2025; 14(11):782. https://doi.org/10.3390/cells14110782
Chicago/Turabian StyleNunes, Jose Ricardo de Assis, Halil Ibrahim Ceylan, Paulo F. de Almeida-Neto, Eugenia Murawska-Ciałowicz, Nicola Luigi Bragazzi, and Gilmara Gomes de Assis. 2025. "The Influence of the ACTN3 R577X Genotype on Performance in Brazilian National-Level Decathlon Athletes: A Pilot Study" Cells 14, no. 11: 782. https://doi.org/10.3390/cells14110782
APA StyleNunes, J. R. d. A., Ceylan, H. I., de Almeida-Neto, P. F., Murawska-Ciałowicz, E., Bragazzi, N. L., & de Assis, G. G. (2025). The Influence of the ACTN3 R577X Genotype on Performance in Brazilian National-Level Decathlon Athletes: A Pilot Study. Cells, 14(11), 782. https://doi.org/10.3390/cells14110782