

Identification of B Cell Subpopulations with Pro- and Anti-Tumorigenic Properties in an Immunocompetent Mouse Model of Head and Neck Squamous Cell Carcinoma

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
1.1. Understanding the Complex Landscape of HNC: Heterogeneity, Impact, and Prognosis
1.2. Exploring the TME in Orthotopic Grown Tumors
1.3. Tumorigenesis and Immune Evasion in TME
1.4. Unveiling B-Cell Complexity: Shaping Immune Responses and Impact on Tumor Dynamics in HNSCCs
2. Materials and Methods
2.1. SCC VII—Cell Culture
2.2. Animal Model
2.3. Spleen, Lymph Nodes, and Tumor Preparation
2.4. Flow Cytometry
2.5. ELISA
3. Results
3.1. Identification of Potentially Pro- and Anti-Tumorigenic B Cell Populations in a Mouse Model of HNSCC
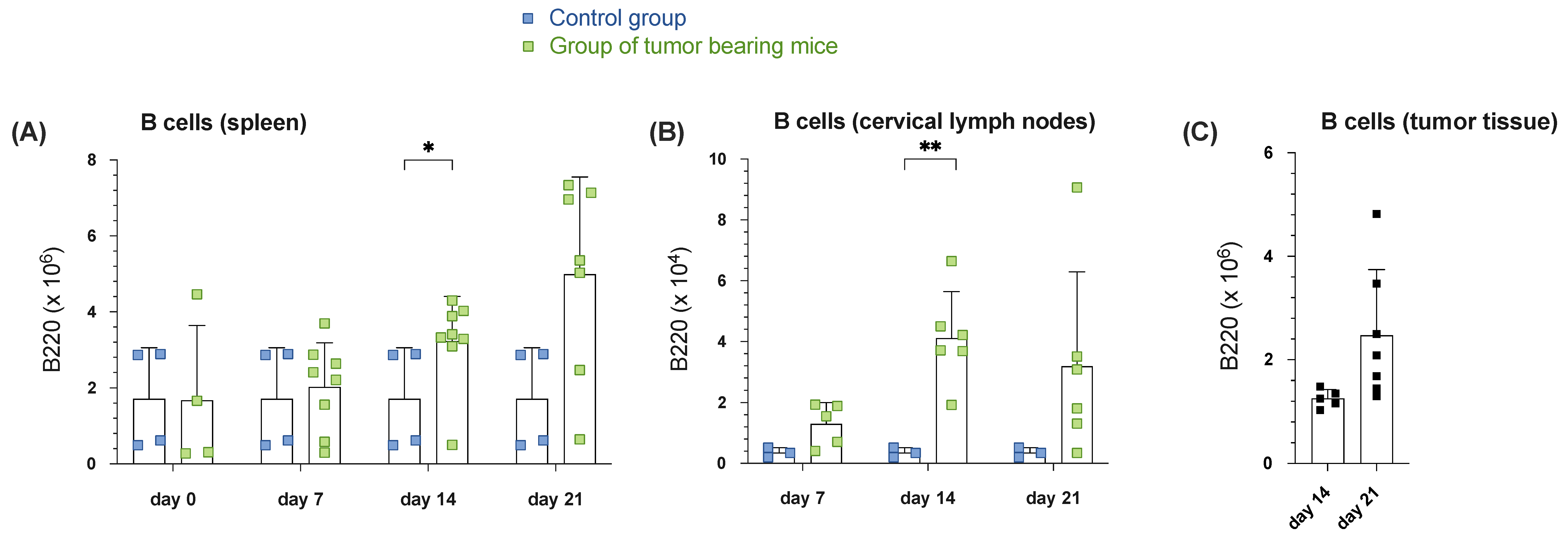
3.1.1. Increased B Cell Numbers in Spleen, Draining Cervical Lymph Nodes and Tumor Tissue During Tumor Progression in Orthotopic HNSCC
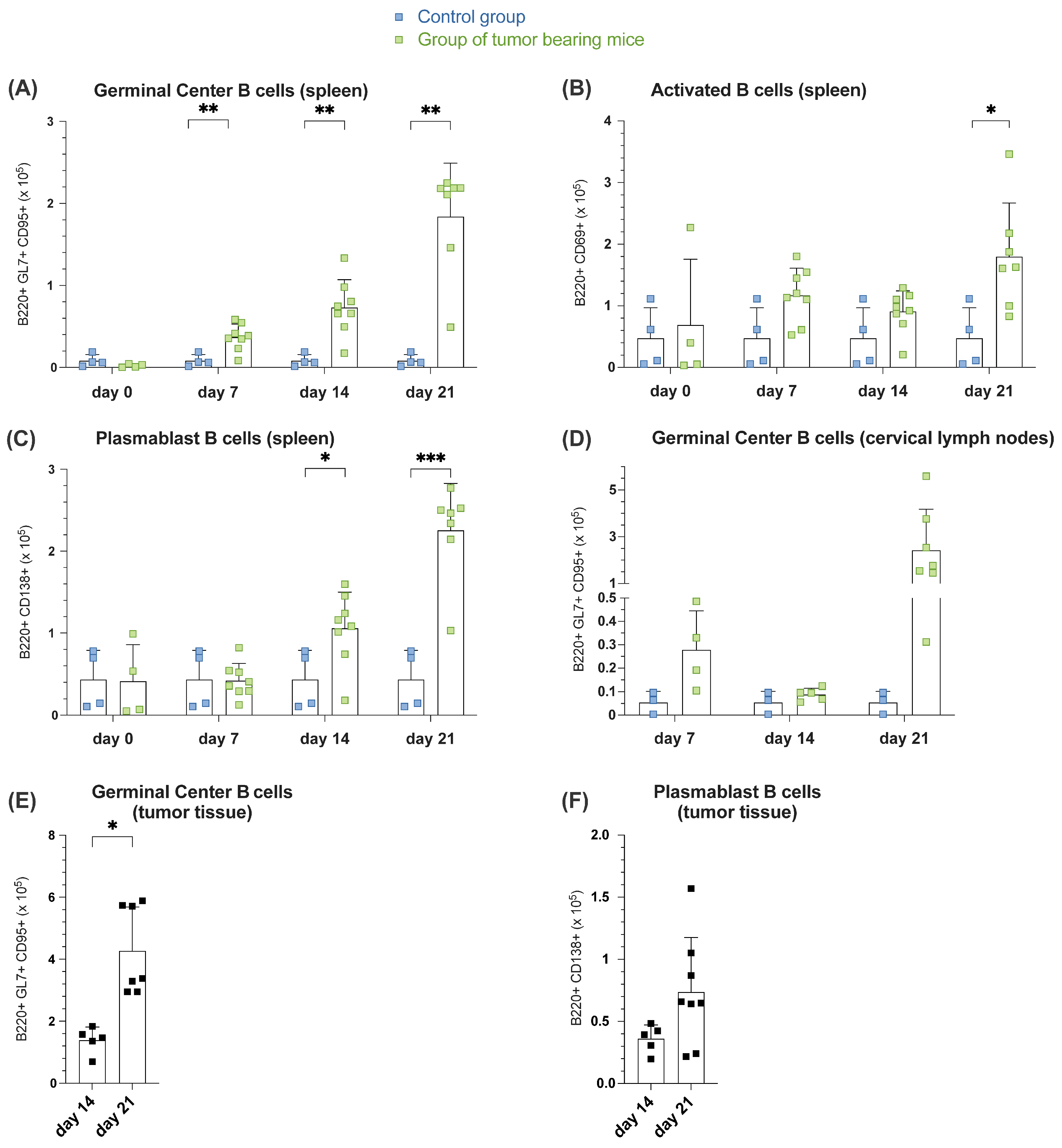
3.1.2. Increased B Cell Activation, GC Formation and Plasmablast Generation During Tumor Formation in Spleen, Lymph Nodes and Tumor Tissue
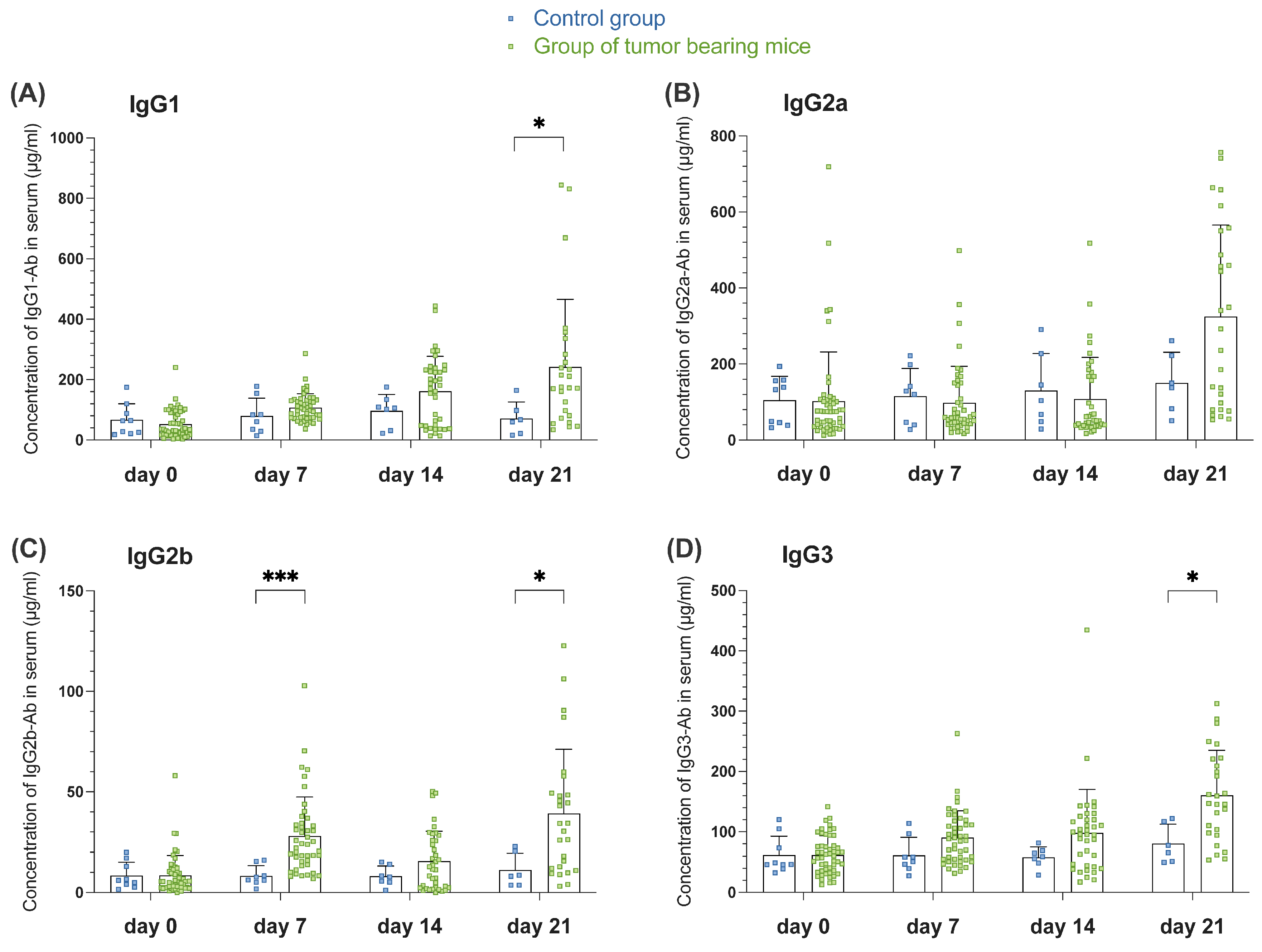
3.1.3. Tumor-Induced GC Reaction Correlates with the Appearance of Switched Igs in Serum of Tumor-Bearing Mice of Orthotopic HNSCC
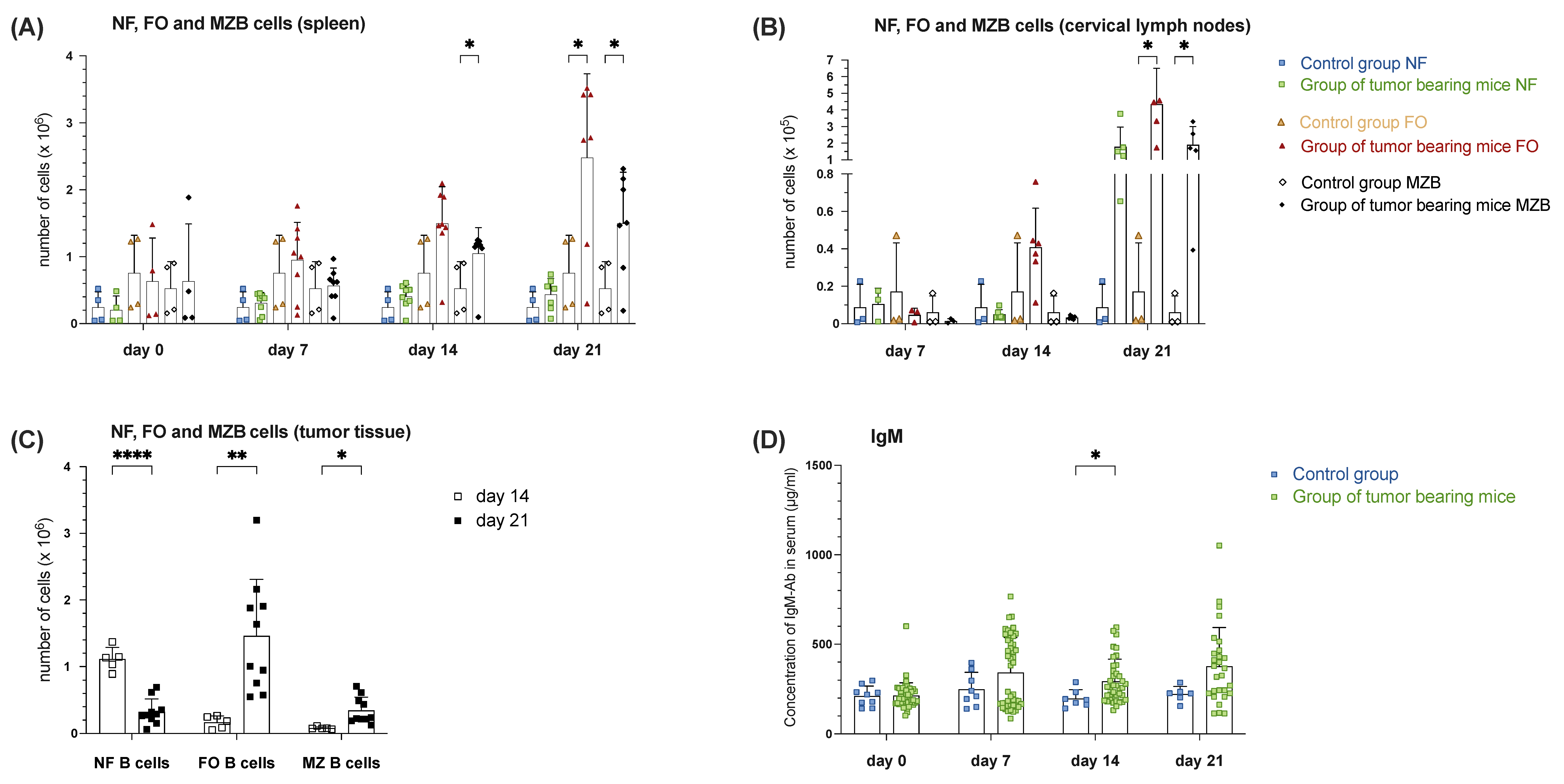
3.1.4. Increased MZ B Cell Population in Spleen, Draining Lymph Nodes and Tumor Tissue During Tumor Progression Correlates with Increased Levels of Serum IgM in Tumor-Bearing Mice of Orthotopic HNSCC
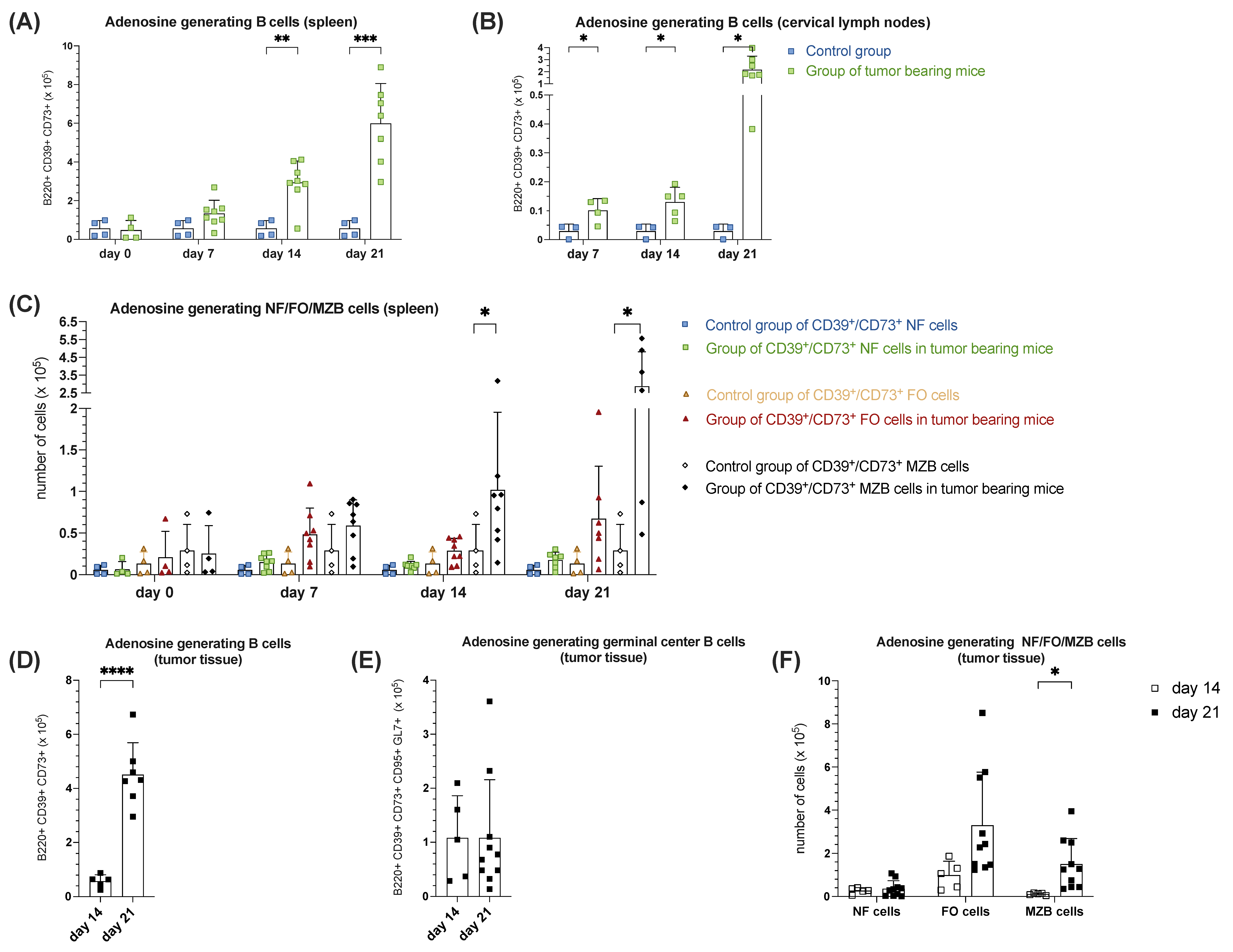
3.1.5. Increased B Cell-Mediated Potential of Ado-Generation During Tumorigenesis in Orthotopic HNSCC
4. Discussion
4.1. Increased B Cell Numbers in Spleen, Draining Lymph Nodes, and Tumor Tissue During Tumor Progression
4.2. GCs and Their Function
4.3. Tumor-Induced GC Reaction Correlates with the Appearance Ig Switched Immunoglobulins in Serum of Tumor Formation
4.4. Increased MZ B Cell Population in Spleen, Draining Lymph Nodes and Tumor Tissue During Tumor Progression Correlates with Increased Levels of Serum IgM in Tumor-Bearing Mice
4.5. Limitations of the Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Wang, H.C.; Chan, L.P.; Cho, S.F. Targeting the Immune Microenvironment in the Treatment of Head and Neck Squamous Cell Carcinoma. Front. Oncol. 2019, 9, 1084. [Google Scholar] [CrossRef]
- Peltanova, B.; Raudenska, M.; Masarik, M. Effect of Tumor Microenvironment on Pathogenesis of the Head and Neck Squamous Cell Carcinoma: A Systematic Review. Mol. Cancer 2019, 18, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and Neck Squamous Cell Carcinoma. Nat. Rev. Dis. Primers 2020, 6, 92. [Google Scholar] [CrossRef]
- Thomas, S.J.; Penfold, C.M.; Waylen, A.; Ness, A.R. The Changing Aetiology of Head and Neck Squamous Cell Cancer: A Tale of Three Cancers? Clin. Otolaryngol. 2018, 43, 999–1003. [Google Scholar] [CrossRef] [PubMed]
- Seliger, B.; Massa, C.; Yang, B.; Bethmann, D.; Kappler, M.; Eckert, A.W.; Wickenhauser, C. Immune Escape Mechanisms and Their Clinical Relevance in Head and Neck Squamous Cell Carcinoma. Int. J. Mol. Sci. 2020, 21, 7032. [Google Scholar] [CrossRef] [PubMed]
- Pezzuto, F.; Buonaguro, L.; Caponigro, F.; Ionna, F.; Starita, N.; Annunziata, C.; Buonaguro, F.M.; Tornesello, M.L. Update on Head and Neck Cancer: Current Knowledge on Epidemiology, Risk Factors, Molecular Features and Novel Therapies. Oncology 2015, 89, 125–136. [Google Scholar] [CrossRef]
- Canning, M.; Guo, G.; Yu, M.; Myint, C.; Groves, M.W.; Byrd, J.K.; Cui, Y. Heterogeneity of the Head and Neck Squamous Cell Carcinoma Immune Landscape and Its Impact on Immunotherapy. Front. Cell Dev. Biol. 2019, 7, 52. [Google Scholar] [CrossRef]
- Hoffmann, T.K. Systemic Therapy Strategies for Head-Neck Carcinomas: Current Status. GMS Curr. Top. Otorhinolaryngol. Head. Neck Surg. 2012, 11, Doc03. [Google Scholar] [CrossRef]
- Sano, D.; Myers, J.N. Xenograft Models of Head and Neck Cancers. Head. Neck Oncol. 2009, 1, 32. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Yu, D. Tumor Microenvironment as a Therapeutic Target in Cancer. Pharmacol. Ther. 2021, 221, 107753. [Google Scholar] [CrossRef]
- Jeske, S.S.; Brand, M.; Ziebart, A.; Laban, S.; Doescher, J.; Greve, J.; Jackson, E.K.; Hoffmann, T.K.; Brunner, C.; Schuler, P.J. Adenosine-Producing Regulatory B Cells in Head and Neck Cancer. Cancer Immunol. Immunother. 2020, 69, 1205–1216. [Google Scholar] [CrossRef]
- Vahle, A.K.; Kerem, A.; Öztürk, E.; Bankfalvi, A.; Lang, S.; Brandau, S. Optimization of an Orthotopic Murine Model of Head and Neck Squamous Cell Carcinoma in Fully Immunocompetent Mice—Role of Toll-like-Receptor 4 Expressed on Host Cells. Cancer Lett. 2012, 317, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Brand, M.; Laban, S.; Theodoraki, M.N.; Doescher, J.; Hoffmann, T.K.; Schuler, P.J.; Brunner, C. Characterization and Differentiation of the Tumor Microenvironment (Tme) of Orthotopic and Subcutaneously Grown Head and Neck Squamous Cell Carcinoma (Hnscc) in Immunocompetent Mice. Int. J. Mol. Sci. 2021, 22, 247. [Google Scholar] [CrossRef] [PubMed]
- Saze, Z.; Schuler, P.J.; Hong, C.S.; Cheng, D.; Jackson, E.K.; Whiteside, T.L. Adenosine Production by Human B Cells and B Cell–Mediated Suppression of Activated T Cells. Blood 2013, 122, 9. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, B.W., Jr.; Cope, K.A.; Johnson, C.S.; Schwartz, M.R. A New Immunocompetent Murine Model for Oral Cancer. Arch. Otolaryngol. Head. Neck Surg. 1997, 123, 20–24. [Google Scholar] [CrossRef]
- Wang, X.; Wu, S.; Wu, W.; Zhang, W.; Li, L.; Liu, Q.; Yan, Z. Candida Albicans Promotes Oral Cancer via IL-17A/IL-17RAMacrophage Axis. mbio 2023, 14, e0044723. [Google Scholar] [CrossRef]
- Smith, L.P.; Thomas, G.R. Animal Models for the Study of Squamous Cell Carcinoma of the Upper Aerodigestive Tract: A Historical Perspective with Review of Their Utility and Limitations. Part A. Chemically-Induced de Novo Cancer, Syngeneic Animal Models of HNSCC, Animal Models of Transplanted Xenogeneic Human Tumors. Int. J. Cancer 2006, 118, 2111–2122. [Google Scholar] [CrossRef]
- Ghiringhelli, F.; Bruchard, M.; Chalmin, F.; Rébé, C. Production of Adenosine by Ectonucleotidases: A Key Factor in Tumor Immunoescape. J. Biomed. Biotechnol. 2012, 2012, 473712. [Google Scholar] [CrossRef]
- Wilkat, M.; Bast, H.; Drees, R.; Dünser, J.; Mahr, A.; Azoitei, N.; Marienfeld, R.; Frank, F.; Brhel, M.; Ushmorov, A.; et al. Adenosine Receptor 2B Activity Promotes Autonomous Growth, Migration as Well as Vascularization of Head and Neck Squamous Cell Carcinoma Cells. Int. J. Cancer 2020, 147, 202–217. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, I.; Jeon, D.; Moseman, J.E.; Muralidhar, A.; Potluri, H.K.; McNeel, D.G. Role of B Cells as Antigen Presenting Cells. Front. Immunol. 2022, 13, 954936. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, Z.; Li, J.; Wang, K.; Zhu, W.; Zeng, Y. The Dual Role of B Cells in the Tumor Microenvironment: Implications for Cancer Immunology and Therapy. Int. J. Mol. Sci. 2024, 25, 11825. [Google Scholar] [CrossRef]
- Bao, J.; Betzler, A.C.; Hess, J.; Brunner, C. Exploring the Dual Role of B Cells in Solid Tumors: Implications for Head and Neck Squamous Cell Carcinoma. Front. Immunol. 2023, 14, 1233085. [Google Scholar]
- Sarvaria, A.; Madrigal, J.A.; Saudemont, A. B Cell Regulation in Cancer and Anti-Tumor Immunity. Cell Mol. Immunol. 2017, 14, 662–674. [Google Scholar] [CrossRef]
- Tsou, P.; Katayama, H.; Ostrin, E.J.; Hanash, S.M. The Emerging Role of b Cells in Tumor Immunity. Cancer Res. 2016, 76, 5591–5601. [Google Scholar] [CrossRef]
- Lechner, A.; Schlößer, H.A.; Thelen, M.; Wennhold, K.; Rothschild, S.I.; Gilles, R.; Quaas, A.; Siefer, O.G.; Huebbers, C.U.; Cukuroglu, E.; et al. Tumor-Associated B Cells and Humoral Immune Response in Head and Neck Squamous Cell Carcinoma. Oncoimmunology 2019, 8, 1535293. [Google Scholar] [CrossRef] [PubMed]
- Bowser, J.L.; Blackburn, M.R.; Shipley, G.L.; Molina, J.G.; Dunner, K.; Broaddus, R.R. Loss of CD73-Mediated Actin Polymerization Promotes Endometrial Tumor Progression. J. Clin. Investig. 2016, 126, 220–238. [Google Scholar] [CrossRef]
- Paul, W.E. Immunity; Johns Hopkins University Press: Baltimore, MD, USA, 2015; ISBN 9781421418018. [Google Scholar]
- Ruffin, A.T.; Cillo, A.R.; Tabib, T.; Liu, A.; Onkar, S.; Kunning, S.R.; Lampenfeld, C.; Atiya, H.I.; Abecassis, I.; Kürten, C.H.L.; et al. B Cell Signatures and Tertiary Lymphoid Structures Contribute to Outcome in Head and Neck Squamous Cell Carcinoma. Nat. Commun. 2021, 12, 3349. [Google Scholar] [CrossRef]
- Tufail, M. Unlocking the Potential of the Tumor Microenvironment for Cancer Therapy. Pathol. Res. Pract. 2023, 251, 154846. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Lu, F.; Lyu, K.; Chang, A.E.; Li, Q. Emerging Concepts Regarding Pro- and Anti Tumor Properties of B Cells in Tumor Immunity. Front. Immunol. 2022, 13, 881427. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.; Rose, N.; Lindner, J.M.; Kim, H.; Gonçalves, A.R.; Callegari, I.; Syedbasha, M.; Kaufmann, L.; Egli, A.; Lindberg, R.L.P.; et al. Antigen Extraction and B Cell Activation Enable Identification of Rare Membrane Antigen Specific Human B Cells. Front. Immunol. 2019, 10, 829. [Google Scholar] [CrossRef]
- Maddur, M.S.; Sharma, M.; Hegde, P.; Stephen-Victor, E.; Pulendran, B.; Kaveri, S.V.; Bayry, J. Human B Cells Induce Dendritic Cell Maturation and Favour Th2 Polarization by Inducing OX-40 Ligand. Nat. Commun. 2014, 5, 4092. [Google Scholar] [CrossRef] [PubMed]
- Germain, C.; Gnjatic, S.; Dieu-Nosjean, M.C. Tertiary Lymphoid Structure-Associated B Cells Are Key Players in Anti-Tumor Immunity. Front. Immunol. 2015, 6, 67. [Google Scholar] [CrossRef]
- Li, Q.; Liu, X.; Wang, D.; Wang, Y.; Lu, H.; Wen, S.; Fang, J.; Cheng, B.; Wang, Z. Prognostic Value of Tertiary Lymphoid Structure and Tumour Infiltrating Lymphocytes in Oral Squamous Cell Carcinoma. Int. J. Oral Sci. 2020, 12, 24. [Google Scholar] [CrossRef] [PubMed]
- Sautès-Fridman, C.; Lawand, M.; Giraldo, N.A.; Kaplon, H.; Germain, C.; Fridman, W.H.; Dieu-Nosjean, M.C. Tertiary Lymphoid Structures in Cancers: Prognostic Value, Regulation, and Manipulation for Therapeutic Intervention. Front. Immunol. 2016, 7, 407. [Google Scholar] [CrossRef]
- Koroleva, E.P.; Fu, Y.X.; Tumanov, A.V. Lymphotoxin in Physiology of Lymphoid Tissues—Implication for Antiviral Defense. Cytokine 2018, 101, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Wirsing, A.M.; Rikardsen, O.G.; Steigen, S.E.; Uhlin-Hansen, L.; Hadler-Olsen, E. Characterisation and Prognostic Value of Tertiary Lymphoid Structures in Oral Squamous Cell Carcinoma. BMC Clin. Pathol. 2014, 14, 38. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Guo, Q.; Zhang, X.; Dong, X.; Liu, W.; Zhang, A.; Li, Y.; Yan, J.; Jia, G.; Zheng, Z.; et al. Oral Cancer-Associated Tertiary Lymphoid Structures: Gene Expression Profile and Prognostic Value. Clin. Exp. Immunol. 2020, 199, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Erazo, L.; Rhodes, J.L.; Marion, V.C.; Kemp, R.A. Tertiary Lymphoid Structures in Cancer—Considerations for Patient Prognosis. Cell Mol. Immunol. 2020, 17, 570–575. [Google Scholar] [CrossRef]
- Posch, F.; Silina, K.; Leibl, S.; Mündlein, A.; Moch, H.; Siebenhüner, A.; Samaras, P.; Riedl, J.; Stotz, M.; Szkandera, J.; et al. Maturation of Tertiary Lymphoid Structures and Recurrence of Stage II and III Colorectal Cancer. Oncoimmunology 2018, 7, e1378844. [Google Scholar] [CrossRef]
- Werner, F.; Wagner, C.; Simon, M.; Glatz, K.; Mertz, K.D.; Läubli, H.; Griss, J.; Wagner, S.N. A Standardized Analysis of Tertiary Lymphoid Structures in Human Melanoma: Disease Progression- and Tumor Site-Associated Changes With Germinal Center Alteration. Front. Immunol. 2021, 12, 675146. [Google Scholar] [CrossRef]
- Li, H.; Zhu, S.W.; Zhou, J.J.; Chen, D.R.; Liu, J.; Wu, Z.Z.; Wang, W.Y.; Zhang, M.J.; Sun, Z.J. Tertiary Lymphoid Structure Raises Survival and Immunotherapy in HPV− HNSCC. J. Dent. Res. 2023, 102, 678–688. [Google Scholar] [CrossRef]
- Wang, M.; Zhai, R.; Wang, M.; Zhu, W.; Zhang, J.; Yu, M.; Zhang, W.; Ye, J.; Liu, L. Tertiary Lymphoid Structures in Head and Neck Squamous Cell Carcinoma Improve Prognosis by Recruiting CD8+ T Cells. Mol. Oncol. 2023, 17, 1514–1530. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Meng, X.; Tang, X.; Zou, W.; He, Y. Intratumoral Tertiary Lymphoid Structures Promote Patient Survival and Immunotherapy Response in Head Neck Squamous Cell Carcinoma. Cancer Immunol. Immunother. 2023, 72, 1505–1521. [Google Scholar] [CrossRef] [PubMed]
- Wieland, A.; Patel, M.R.; Cardenas, M.A.; Eberhardt, C.S.; Hudson, W.H.; Obeng, R.C.; Griffith, C.C.; Wang, X.; Chen, Z.G.; Kissick, H.T.; et al. Defining HPV-Specific B Cell Responses in Patients with Head and Neck Cancer. Nature 2021, 597, 274–278. [Google Scholar] [CrossRef]
- Kim, S.S.; Shen, S.; Miyauchi, S.; Dominick Sanders, P.; Franiak-Pietryga, I.; Mell, L.; Silvio Gutkind, J.; Cohen, E.E.W.; Califano, J.A.; Sharabi, A.B. B Cells Improve Overall Survival in HPV-Associated Squamous Cell Carcinomas and Are Activated by Radiation and PD-1 Blockade. Clin. Cancer Res. 2020, 26, 3345–3359. [Google Scholar] [CrossRef]
- Gu-Trantien, C.; Loi, S.; Garaud, S.; Equeter, C.; Libin, M.; De Wind, A.; Ravoet, M.; Le Buanec, H.; Sibille, C.; Manfouo-Foutsop, G.; et al. CD4+ Follicular Helper T Cell Infiltration Predicts Breast Cancer Survival. J. Clin. Investig. 2013, 123, 2873–2892. [Google Scholar] [CrossRef]
- Kroeger, D.R.; Milne, K.; Nelson, B.H. Tumor-Infiltrating Plasma Cells Are Associated with Tertiary Lymphoid Structures, Cytolytic T-Cell Responses, and Superior Prognosis in Ovarian Cancer. Clin. Cancer Res. 2016, 22, 3005–3015. [Google Scholar] [CrossRef] [PubMed]
- Wouters, M.C.A.; Nelson, B.H. Prognostic Significance of Tumor-Infiltrating B Cells and Plasma Cells in Human Cancer. Clin. Cancer Res. 2018, 24, 6125–6135. [Google Scholar] [CrossRef]
- Helmink, B.A.; Reddy, S.M.; Gao, J.; Zhang, S.; Basar, R.; Thakur, R.; Yizhak, K.; Sade-Feldman, M.; Blando, J.; Han, G.; et al. B Cells and Tertiary Lymphoid Structures Promote Immunotherapy Response. Nature 2020, 577, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Hiraoka, N.; Ino, Y.; Yamazaki-Itoh, R. Tertiary Lymphoid Organs in Cancer Tissues. Front. Immunol. 2016, 7, 244. [Google Scholar] [CrossRef] [PubMed]
- Barankiewicz, J.; Dosch, H.M.; Cohen, A. Extracellular Nucleotide Catabolism in Human B and T Lymphocytes. The Source of Adenosine Production. J. Biol. Chem. 1988, 263, 7094–7098. [Google Scholar] [CrossRef] [PubMed]
- Basso, K.; Saito, M.; Sumazin, P.; Margolin, A.A.; Wang, K.; Lim, W.K.; Kitagawa, Y.; Schneider, C.; Alvarez, M.J.; Califano, A.; et al. Integrated Biochemical and Computational Approach Identifies BCL6 Direct Target Genes Controlling Multiple Pathways in Normal Germinal Center B Cells. Blood 2010, 115, 975–984. [Google Scholar] [CrossRef]
- Collins, A.M. IgG Subclass Co-Expression Brings Harmony to the Quartet Model of Murine IgG Function. Immunol. Cell Biol. 2016, 94, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Marinkovic, D.; Marinkovic, T. Putative Role of Marginal Zone B Cells in Pathophysiological Processes. Scand. J. Immunol. 2020, 92, e12920. [Google Scholar] [CrossRef] [PubMed]
- Betzler, A.C.; Ezić, J.; Abou Kors, T.; Hoffmann, T.K.; Wirth, T.; Brunner, C. T Cell Specific BOB.1/OBF.1 Expression Promotes Germinal Center Response and T Helper Cell Differentiation. Front. Immunol. 2022, 13, 889564. [Google Scholar] [CrossRef] [PubMed]
- Betzler, A.C.; Fiedler, K.; Hoffmann, T.K.; Fehling, H.J.; Wirth, T.; Brunner, C. BOB.1/OBF.1 Is Required during B-Cell Ontogeny for B-Cell Differentiation and Germinal Center Function. Eur. J. Immunol. 2022, 52, 404–417. [Google Scholar] [CrossRef] [PubMed]
- Doyon-Laliberté, K.; Aranguren, M.; Poudrier, J.; Roger, M. Marginal Zone B-Cell Populations and Their Regulatory Potential in the Context of HIV and Other Chronic Inflammatory Conditions. Int. J. Mol. Sci. 2022, 23, 3372. [Google Scholar] [CrossRef] [PubMed]
- Conter, L.J.; Song, E.; Shlomchik, M.J.; Tomayko, M.M. CD73 Expression Is Dynamically Regulated in the Germinal Center and Bone Marrow Plasma Cells Are Diminished in Its Absence. PLoS ONE 2014, 9, e92009. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sonntag, M.; Stanojevic, S.; Laban, S.; Schuler, P.J.; Hoffmann, T.K.; Brunner, C. Identification of B Cell Subpopulations with Pro- and Anti-Tumorigenic Properties in an Immunocompetent Mouse Model of Head and Neck Squamous Cell Carcinoma. Cells 2025, 14, 20. https://doi.org/10.3390/cells14010020
Sonntag M, Stanojevic S, Laban S, Schuler PJ, Hoffmann TK, Brunner C. Identification of B Cell Subpopulations with Pro- and Anti-Tumorigenic Properties in an Immunocompetent Mouse Model of Head and Neck Squamous Cell Carcinoma. Cells. 2025; 14(1):20. https://doi.org/10.3390/cells14010020
Chicago/Turabian StyleSonntag, Michael, Sandra Stanojevic, Simon Laban, Patrick J. Schuler, Thomas K. Hoffmann, and Cornelia Brunner. 2025. "Identification of B Cell Subpopulations with Pro- and Anti-Tumorigenic Properties in an Immunocompetent Mouse Model of Head and Neck Squamous Cell Carcinoma" Cells 14, no. 1: 20. https://doi.org/10.3390/cells14010020
APA StyleSonntag, M., Stanojevic, S., Laban, S., Schuler, P. J., Hoffmann, T. K., & Brunner, C. (2025). Identification of B Cell Subpopulations with Pro- and Anti-Tumorigenic Properties in an Immunocompetent Mouse Model of Head and Neck Squamous Cell Carcinoma. Cells, 14(1), 20. https://doi.org/10.3390/cells14010020