
Metabolic Dysfunction-Associated Steatotic Liver Disease and Alcohol-Associated Liver Disease: Liver DNA Methylation Analysis—A Systematic Review

 , , , and
, , , and
Abstract
:1. Introduction
2. Methods
2.1. Search Strategy
2.2. Eligibility Criteria
2.3. Data Collection
2.4. Data Extraction
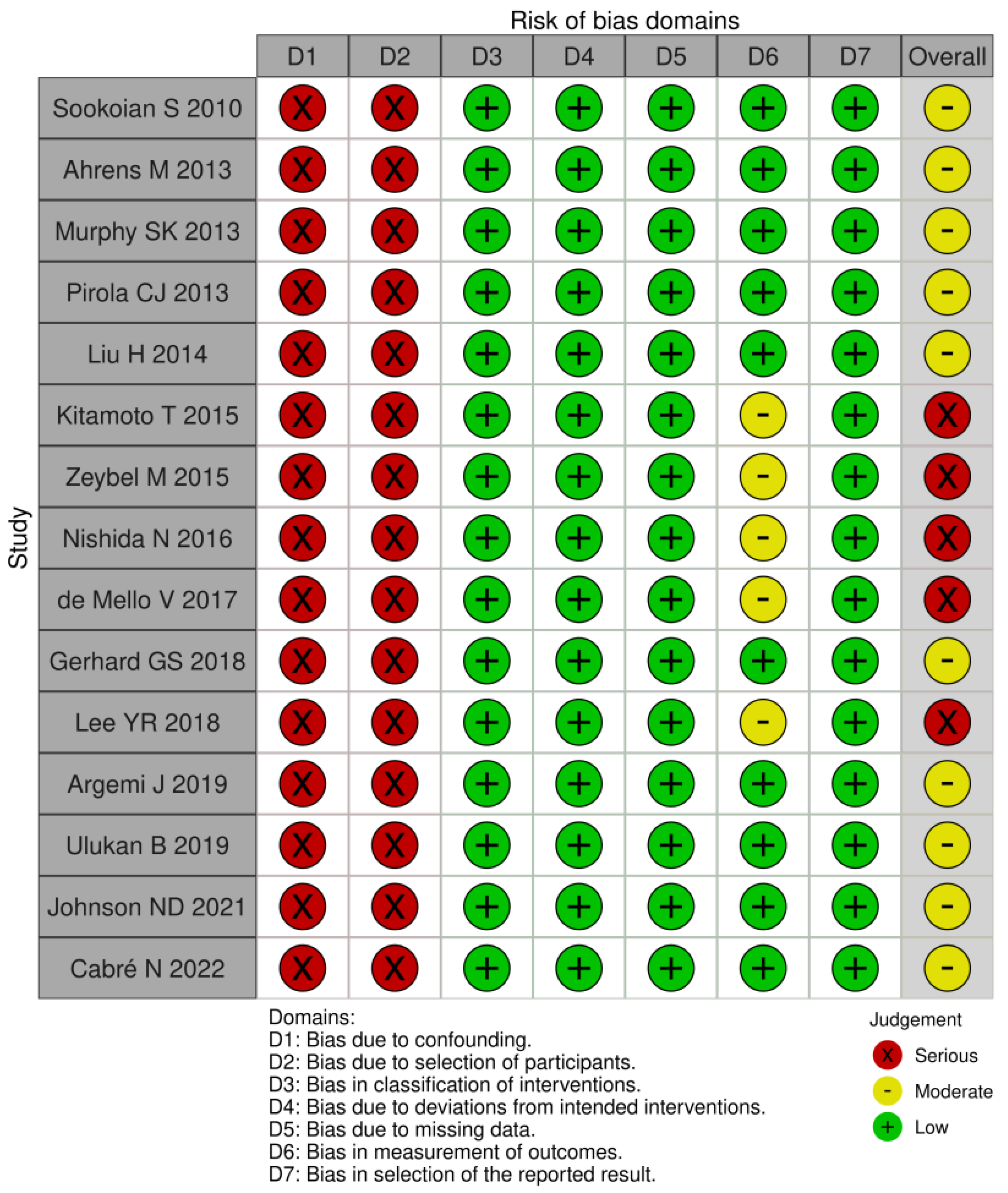
2.5. Risk of Bias Assessment
2.6. Pathway Analysis
3. Results
3.1. Article Selection, Study Characteristics
3.1.1. Article Selection
3.1.2. Study Characteristics
3.2. Differentially Methylated Genes in MASLD, Differentially Methylated Genes in ALD, and Differentially Methylated Genes in MASLD and ALD
3.2.1. Differentially Methylated Genes in MASLD
3.2.2. Differentially Methylated Genes in ALD
3.2.3. Differentially Methylated Genes in MASLD and ALD
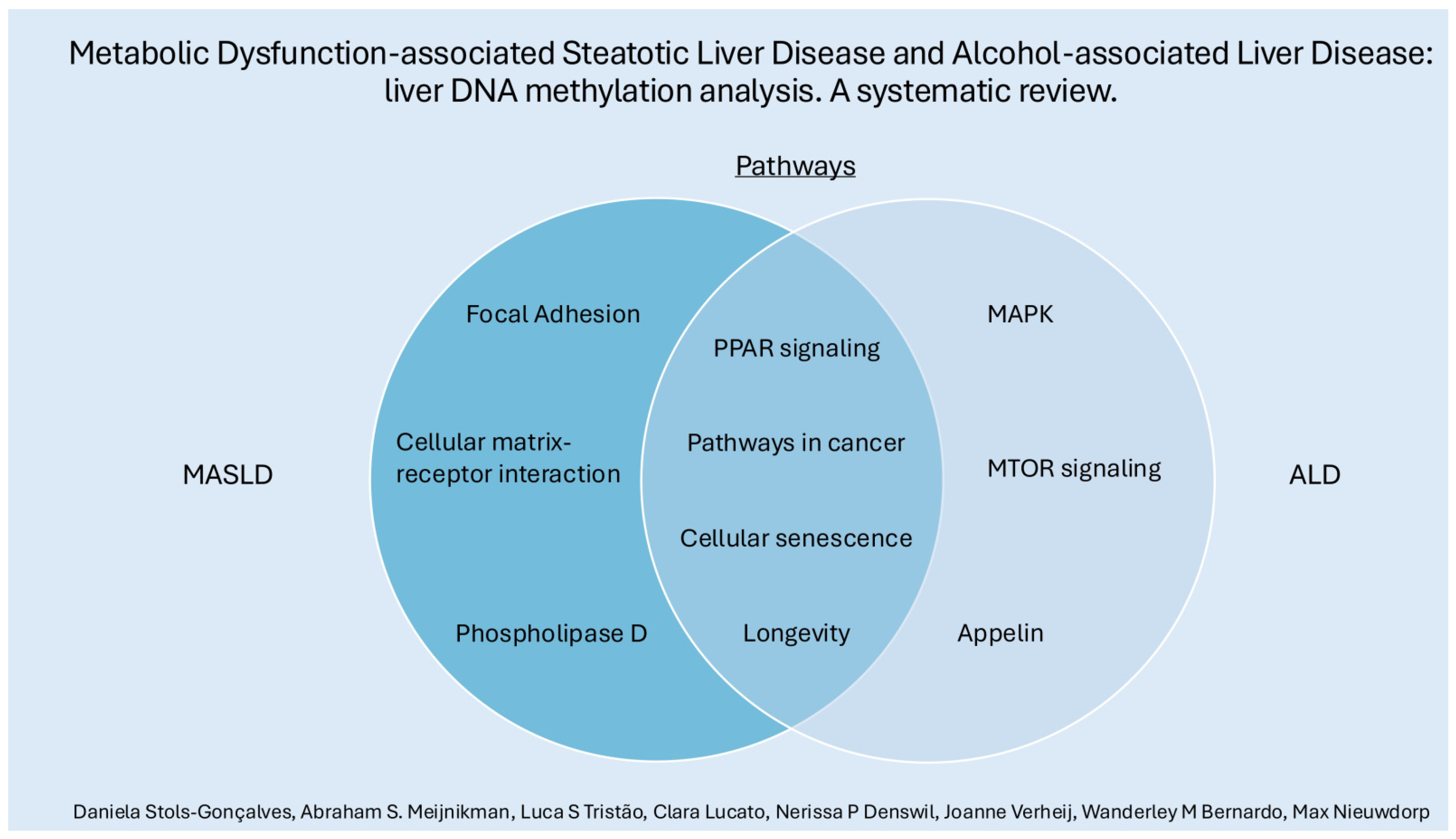
3.3. MASLD Pathways, ALD Pathways, Common Pathways MASLD and ALD
3.3.1. MASLD Pathways
3.3.2. ALD Pathways
3.3.3. Common Pathways in MASLD and ALD
3.4. Risk of Bias Assessment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.-C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Lackner, C.; Stauber, R.E.; Davies, S.; Denk, H.; Dienes, H.P.; Gnemmi, V.; Guido, M.; Miquel, R.; Paradis, V.; Schirmacher, P.; et al. Development and prognostic relevance of a histologic grading and staging system for alcohol-related liver disease. J. Hepatol. 2021, 75, 810–819. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Golabi, P.; Paik, J.M.; Henry, A.; Van Dongen, C.; Henry, L. The global epidemiology of nonalcoholic fatty liver disease (NAFLD) and nonalcoholic steatohepatitis (NASH): A systematic review. Hepatology 2023, 77, 1335–1347. [Google Scholar] [CrossRef]
- Niu, X.; Zhu, L.; Xu, Y.; Zhang, M.; Hao, Y.; Ma, L.; Li, Y.; Xing, H. Global prevalence, incidence, and outcomes of alcohol related liver diseases: A systematic review and meta-analysis. BMC Public Health 2023, 23, 859. [Google Scholar] [CrossRef]
- Eslam, M.; Sanyal, A.J.; George, J.; International Consensus, P. MAFLD: A Consensus-Driven Proposed Nomenclature for Metabolic Associated Fatty Liver Disease. Gastroenterology 2020, 158, 1999–2014.e1. [Google Scholar] [CrossRef]
- Rinella, M.E.; Lazarus, J.V.; Ratziu, V.; Francque, S.M.; Sanyal, A.J.; Kanwal, F.; Romero, D.; Abdelmalek, M.F.; Anstee, Q.M.; Arab, J.P.; et al. A multisociety Delphi consensus statement on new fatty liver disease nomenclature. J. Hepatol. 2023, 79, 1542–1556. [Google Scholar] [CrossRef]
- Ledford, H. First US drug approved for a liver disease surging around the world. Nature 2024. online ahead of print. [Google Scholar] [CrossRef]
- Condon, S.; Jophlin, L.L. Past, Present, and Future Therapies for Alcohol-associated Hepatitis. Clin. Ther. 2023, 45, 1171–1176. [Google Scholar] [CrossRef]
- Niu, L.; Thiele, M.; Geyer, P.E.; Rasmussen, D.N.; Webel, H.E.; Santos, A.; Gupta, R.; Meier, F.; Strauss, M.; Kjaergaard, M.; et al. Noninvasive proteomic biomarkers for alcohol-related liver disease. Nat. Med. 2022, 28, 1277–1287. [Google Scholar] [CrossRef]
- Eslam, M.; Valenti, L.; Romeo, S. Genetics and epigenetics of NAFLD and NASH: Clinical impact. J. Hepatol. 2018, 68, 268–279. [Google Scholar] [CrossRef]
- Habash, N.W.; Sehrawat, T.S.; Shah, V.H.; Cao, S. Epigenetics of alcohol-related liver diseases. JHEP Rep. 2022, 4, 100466. [Google Scholar] [CrossRef] [PubMed]
- Ling, C.; Ronn, T. Epigenetics in Human Obesity and Type 2 Diabetes. Cell Metab. 2019, 29, 1028–1044. [Google Scholar] [CrossRef] [PubMed]
- Lobbestael, G. DedupEndNote(version1.0.0). Available online: https://github.com/globbestael/DedupEndNote (accessed on 3 January 2024).
- U. o. York. PROSPERO: International Prospective Register of Systematic Reviews. National Institute for Health Research. Available online: https://www.crd.york.ac.uk/prospero/ (accessed on 11 February 2024).
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
- Ouzzani, M.; Hammady, H.; Fedorowicz, Z.; Elmagarmid, A. Rayyan-a web and mobile app for systematic reviews. Syst. Rev. 2016, 5, 210. [Google Scholar] [CrossRef] [PubMed]
- Sterne, J.A.; Hernan, M.A.; Reeves, B.C.; Savovic, J.; Berkman, N.D.; Viswanathan, M.; Henry, D.; Altman, D.G.; Ansari, M.T.; Boutron, I.; et al. ROBINS-I: A tool for assessing risk of bias in non-randomised studies of interventions. BMJ 2016, 355, i4919. [Google Scholar] [CrossRef]
- Cabre, N.; Luciano-Mateo, F.; Chapski, D.J.; Baiges-Gaya, G.; Fernandez-Arroyo, S.; Hernandez-Aguilera, A.; Castane, H.; Rodriguez-Tomas, E.; Paris, M.; Sabench, F.; et al. Laparoscopic Sleeve Gastrectomy in Patients with Severe Obesity Restores Adaptive Responses Leading to Nonalcoholic Steatohepatitis. Int. J. Mol. Sci. 2022, 23, 7830. [Google Scholar] [CrossRef]
- Johnson, N.D.; Wu, X.; Still, C.D.; Chu, X.; Petrick, A.T.; Gerhard, G.S.; Conneely, K.N.; DiStefano, J.K. Differential DNA methylation and changing cell-type proportions as fibrotic stage progresses in NAFLD. Clin. Epigenetics 2021, 13, 152. [Google Scholar] [CrossRef]
- Argemi, J.; Latasa, M.U.; Atkinson, S.R.; Blokhin, I.O.; Massey, V.; Gue, J.P.; Cabezas, J.; Lozano, J.J.; Van Booven, D.; Bell, A.; et al. Defective HNF4alpha-dependent gene expression as a driver of hepatocellular failure in alcoholic hepatitis. Nat. Commun. 2019, 10, 3126. [Google Scholar] [CrossRef]
- Ulukan, B.; Yiğit, B.; Yılmaz, O.; Comert, M.; Erkan, M.; Adali, G.; Dayangac, M.; Zeybel, M. FRI-351-DNA methylation pattern of TM6SF2 influences NAFLD progression in genotype-dependent manner. J. Hepatol. 2019, 70, e549. [Google Scholar] [CrossRef]
- Gerhard, G.S.; Malenica, I.; Llaci, L.; Chu, X.; Petrick, A.T.; Still, C.D.; DiStefano, J.K. Differentially methylated loci in NAFLD cirrhosis are associated with key signaling pathways. Clin. Epigenet. 2018, 10, 93. [Google Scholar] [CrossRef]
- Lee, H.Y.R.L. Differential DNA methylation of PPAR-gamma genes in patients with non-alcoholic fatty liver disease. In Proceedings of the 27th Annual Conference of APASL, New Delhi, India, 14–18 March 2018; Volume 12, no. 2. pp. S436–S437. [Google Scholar]
- de Mello, V.D.; Matte, A.; Perfilyev, A.; Mannisto, V.; Ronn, T.; Nilsson, E.; Kakela, P.; Ling, C.; Pihlajamaki, J. Human liver epigenetic alterations in non-alcoholic steatohepatitis are related to insulin action. Epigenetics 2017, 12, 287–295. [Google Scholar] [CrossRef]
- Nishida, N.; Yada, N.; Hagiwara, S.; Sakurai, T.; Kitano, M.; Kudo, M. Unique features associated with hepatic oxidative DNA damage and DNA methylation in non-alcoholic fatty liver disease. J. Gastroenterol. Hepatol. 2016, 31, 1646–1653. [Google Scholar] [CrossRef] [PubMed]
- Kitamoto, T.; Kitamoto, A.; Ogawa, Y.; Honda, Y.; Imajo, K.; Saito, S.; Yoneda, M.; Nakamura, T.; Nakajima, A.; Hotta, K. Targeted-bisulfite sequence analysis of the methylation of CpG islands in genes encoding PNPLA3, SAMM50, and PARVB of patients with non-alcoholic fatty liver disease. J. Hepatol. 2015, 63, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Zeybel, M.; Hardy, T.; Robinson, S.M.; Fox, C.; Anstee, Q.M.; Ness, T.; Masson, S.; Mathers, J.C.; French, J.; White, S.; et al. Differential DNA methylation of genes involved in fibrosis progression in non-alcoholic fatty liver disease and alcoholic liver disease. Clin. Epigenet. 2015, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Gong, M.; French, B.A.; Li, J.; Tillman, B.; French, S.W. Mallory-Denk Body (MDB) formation modulates Ufmylation expression epigenetically in alcoholic hepatitis (AH) and non-alcoholic steatohepatitis (NASH). Exp. Mol. Pathol. 2014, 97, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Pirola, C.J.; Gianotti, T.F.; Burgueno, A.L.; Rey-Funes, M.; Loidl, C.F.; Mallardi, P.; Martino, J.S.; Castano, G.O.; Sookoian, S. Epigenetic modification of liver mitochondrial DNA is associated with histological severity of nonalcoholic fatty liver disease. Gut 2013, 62, 1356–1363. [Google Scholar] [CrossRef]
- Ahrens, M.; Ammerpohl, O.; von Schonfels, W.; Kolarova, J.; Bens, S.; Itzel, T.; Teufel, A.; Herrmann, A.; Brosch, M.; Hinrichsen, H.; et al. DNA methylation analysis in nonalcoholic fatty liver disease suggests distinct disease-specific and remodeling signatures after bariatric surgery. Cell Metab. 2013, 18, 296–302. [Google Scholar] [CrossRef]
- Murphy, S.K.; Yang, H.; Moylan, C.A.; Pang, H.; Dellinger, A.; Abdelmalek, M.F.; Garrett, M.E.; Ashley-Koch, A.; Suzuki, A.; Tillmann, H.L.; et al. Relationship between methylome and transcriptome in patients with nonalcoholic fatty liver disease. Gastroenterology 2013, 145, 1076–1087. [Google Scholar] [CrossRef]
- Sookoian, S.; Rosselli, M.S.; Gemma, C.; Burgueno, A.L.; Fernandez Gianotti, T.; Castano, G.O.; Pirola, C.J. Epigenetic regulation of insulin resistance in nonalcoholic fatty liver disease: Impact of liver methylation of the peroxisome proliferator-activated receptor gamma coactivator 1alpha promoter. Hepatology 2010, 52, 1992–2000. [Google Scholar] [CrossRef]
- Fougerat, A.; Montagner, A.; Loiseau, N.; Guillou, H.; Wahli, W. Peroxisome Proliferator-Activated Receptors and Their Novel Ligands as Candidates for the Treatment of Non-Alcoholic Fatty Liver Disease. Cells 2020, 9, 1638. [Google Scholar] [CrossRef]
- Fischer, M.; You, M.; Matsumoto, M.; Crabb, D.W. Peroxisome proliferator-activated receptor alpha (PPARalpha) agonist treatment reverses PPARalpha dysfunction and abnormalities in hepatic lipid metabolism in ethanol-fed mice. J. Biol. Chem. 2003, 278, 27997–28004. [Google Scholar] [CrossRef] [PubMed]
- Francque, S.; Verrijken, A.; Caron, S.; Prawitt, J.; Paumelle, R.; Derudas, B.; Lefebvre, P.; Taskinen, M.R.; Van Hul, W.; Mertens, I.; et al. PPARalpha gene expression correlates with severity and histological treatment response in patients with non-alcoholic steatohepatitis. J. Hepatol. 2015, 63, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Noorolyai, S.; Shajari, N.; Baghbani, E.; Sadreddini, S.; Baradaran, B. The relation between PI3K/AKT signalling pathway and cancer. Gene 2019, 698, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Sun, E.J.; Wankell, M.; Palamuthusingam, P.; McFarlane, C.; Hebbard, L. Targeting the PI3K/Akt/mTOR Pathway in Hepatocellular Carcinoma. Biomedicines 2021, 9, 1639. [Google Scholar] [CrossRef]
- Meijnikman, A.S.; Herrema, H.; Scheithauer, T.P.M.; Kroon, J.; Nieuwdorp, M.; Groen, A.K. Evaluating causality of cellular senescence in non-alcoholic fatty liver disease. JHEP Rep. 2021, 3, 100301. [Google Scholar] [CrossRef]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef]
- Ramirez, T.; Longato, L.; Dostalek, M.; Tong, M.; Wands, J.R.; de la Monte, S.M. Insulin resistance, ceramide accumulation and endoplasmic reticulum stress in experimental chronic alcohol-induced steatohepatitis. Alcohol. Alcohol. 2013, 48, 39–52. [Google Scholar] [CrossRef]
- Wickramasinghe, P.B.; Qian, S.; Langley, L.E.; Liu, C.; Jia, L. Hepatocyte Toll-Like Receptor 4 Mediates Alcohol-Induced Insulin Resistance in Mice. Biomolecules 2023, 13, 454. [Google Scholar] [CrossRef]
- de la Monte, S.M.; Yeon, J.E.; Tong, M.; Longato, L.; Chaudhry, R.; Pang, M.Y.; Duan, K.; Wands, J.R. Insulin resistance in experimental alcohol-induced liver disease. J. Gastroenterol. Hepatol. 2008, 23, e477–e486. [Google Scholar] [CrossRef]
- Schrieks, I.C.; Heil, A.L.; Hendriks, H.F.; Mukamal, K.J.; Beulens, J.W. The effect of alcohol consumption on insulin sensitivity and glycemic status: A systematic review and meta-analysis of intervention studies. Diabetes Care 2015, 38, 723–732. [Google Scholar] [CrossRef]
- Kwak, M.; Kim, H.S.; Jiang, Z.G.; Yeo, Y.H.; Trivedi, H.D.; Noureddin, M.; Yang, J.D. MASLD/MetALD and mortality in individuals with any cardio-metabolic risk factor: A population-based study with 26.7 years of follow-up. Hepatology, 2024. [Google Scholar] [CrossRef]
- Fraenkel, E.; Lazurova, I. IGF-1 and IGFBP3 as indirect markers of hepatic insulin resistance and their relation to metabolic syndrome parameters in liver steatosis patients. Endocr. Regul. 2023, 57, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Hammoutene, A.; Rautou, P.E. Role of liver sinusoidal endothelial cells in non-alcoholic fatty liver disease. J. Hepatol. 2019, 70, 1278–1291. [Google Scholar] [CrossRef] [PubMed]
- Auclair, N.; Sane, A.T.; Delvin, E.; Spahis, S.; Levy, E. Phospholipase D as a Potential Modulator of Metabolic Syndrome: Impact of Functional Foods. Antioxid. Redox Signal 2021, 34, 252–278. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Kang, J.H.; Bae, Y.S. The role and regulation of phospholipase D in metabolic disorders. Adv. Biol. Regul. 2024, 91, 100988. [Google Scholar] [CrossRef]
- Hur, J.H.; Park, S.Y.; Dall’Armi, C.; Lee, J.S.; Di Paolo, G.; Lee, H.Y.; Yoon, M.S.; Min, D.S.; Choi, C.S. Phospholipase D1 deficiency in mice causes nonalcoholic fatty liver disease via an autophagy defect. Sci. Rep. 2016, 6, 39170. [Google Scholar] [CrossRef]
- Aroor, A.R.S.; Shukla, S.D. MAP Kinase Signaling in Diverse Effects of Ethanol; Elsevier: Amsterdam, The Netherlands, 2004; Volume 74, pp. 2339–2364. [Google Scholar]
- Melgar-Lesmes, P.; Perramon, M.; Jimenez, W. Roles of the Hepatic Endocannabinoid and Apelin Systems in the Pathogenesis of Liver Fibrosis. Cells 2019, 8, 1311. [Google Scholar] [CrossRef]
- Stankevic, E.; Israelsen, M.; Juel, H.B.; Madsen, A.L.; Angquist, L.; Aldiss, P.S.J.; Torp, N.; Johansen, S.; Hansen, C.D.; Hansen, J.K.; et al. Binge drinking episode causes acute, specific alterations in systemic and hepatic inflammation-related markers. Liver Int. 2023, 43, 2680–2691. [Google Scholar] [CrossRef]
- Ilieva, M.; Dao, J.; Miller, H.E.; Madsen, J.H.; Bishop, A.J.R.; Kauppinen, S.; Uchida, S. Systematic Analysis of Long Non-Coding RNA Genes in Nonalcoholic Fatty Liver Disease. Noncoding RNA 2022, 8, 56. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Study Number | Author | Year | Patient Population | Sample Size | Age (Year) | Sex (Fem/Male) | BMI | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | B | A | B | A | B | A | B | A | B | |||
| 1 | Cabré N | 2022 | Obese NASH (NAS ≥ 5) | Obese non-NASH (NAS ≤ 2) | 8 | 8 | Matched but not specified for this subgroup | Matched but not specified for this subgroup | Matched but not specified for this subgroup | |||
| 2 | Johnson ND | 2021 | G3 = grade 3 fib G3–4 = grade 3–4 fib G4 = grade 4 fib | G0 = no fibrosis | G3 n = 55 G3–4 n = 36 G4 n = 28 | 206 | G3 49.5 ± 10.0 G3/4 50.4 ± 8.1 G4 53.8 ± 9.5 | (All 48.6 ± 11.5) G0 47.2 ± 12.4 | G3 61.8% G3/4 80.5% G4 78.5% | (All 77%) G0 80.5% | G3 48.6 ± 8.0 G3/4 46.5 ± 9.7 G4 48.1 ± 12.1 | All 47.1 ± 9.1 G0 46.6 ± 8.8 |
| 3 | Argemi J | 2019 | AH | Normal liver | 6 | 5 | - | - | - | |||
| 4 | Ulukan B | 2019 | NASH | NAFLD F0 | Cirrhotic NASH (f4) n = 22 (F1/F2 F3) n = 24 | 9 | - | - | - | |||
| 5 | Gerhard GS | 2018 | NAFLD cirrhosis | Normal liver | 11 | 15 | 47.1 ± 6.3 | 51.3 ± 8.2 | 100% women | 41.0 ± 4.6 | 44.4 ± 6.6 | |
| 6 | Lee YR | 2018 | NAFLD | Normal liver | 54 | 18 | - | - | - | |||
| 7 | de Mello V | 2017 | Obese NASH | Obese Normal liver Obese SS | 26 | Normal n = 35 SS n = 34 | NASH = 51.3 ± 7.9 | Nl = 50.7 ± 7.0 SS = 46.9 ± 7.6 | 13/13 | Nl (11/24) SS (10/24) | NASH 43.4 ± 6.4 | Nl 42.4 ± 6.1 SS 43.5 ± 4.7 |
| 8 | Nishida N | 2016 | NAFLD | HCC or normal liver | 65 | 16 | 54.3 (50.7–57.9) | 40/25 | - | - | 29.3 (28.0–30.6) | |
| 9 | Kitamoto T | 2015 | advanced fibrosis (F2–4) | Mild fibrosis (F0–1) | 29 | 36 | - | - | - | |||
| 10a | Zeybel M | 2015 | advanced fibrosis (F2–4) | NAFLD, minimal fibrosis | 9 | 8 | 51.88 (40.43–63.32) | 60.56 (54.5–66.61) | 100% male | 35 (30–43.2) | 36 (30–46) | |
| 10b | Zeybel M | 2015 | ALD | Normal liver | 10 | 17 | 63.31 (67.94–54.71) | 46.3 (39.09–57.27) | 10 male (58%) | 7 male (70%) | 24.2 (19–32.5) | |
| 11 | Hui Liu | 2014 | NASH | Alcoholic hepatitis Normal liver | AH 3–5 NASH 3–5 | 3 | - | - | - | |||
| 12 | Pirola CJ | 2013 | NAFLD (simple steatosis and NASH) | Near normal histology | 22 | Non-MS almost nl n = 18 Steatosis n = 23 | NASH 48 ± 9.2 | Nl 48.3 ± 8.7 Stea 51.6 ± 10.5 | NASH 12/10 | Fem/male Nl 10/8 Steat 14/9 | NASH 31.2 ± 6 | Nl 25.3 ± 4.1 Steat 31.6 ± 5 |
| 13 | Ahrens | 2013 | NASH | No adv Normal control “Healthy” obese Steatosis | 15 | Obese healthy n = 18 Steat n = 12 Normal control n = 18 | 44 (41–50) | 46 (37–49) 47 (40–50) 51 (44–72) | Male % 0 | Male % 42 27 50 | 45 (42–49) | 50 (47–55) 49 (44–56) 24 (21–26) |
| 14 | Murphy SK | 2013 | Advanced NAFLD F3–F4 | Mild NAFDL F0–F1 | 23 | 33 | Advanced 51.7 ± 10.3 | Mild = 51.5 ± 10.3 | Advanced 5/18 | Mild 12/21 | Advanced 33.8 (31.3–41.9) | Mild 32.8 (28.4– 40.4) |
| 15 | Sookoian S | 2010 | NAFLD | NAFLD Control (no histological fatty changes) | 63 | 11 | NAFLD 50.3 ± 9.9 | 49.46 ± 10.2 | NAFLD 32/31 | Control 5/6 | NAFLD 31.9 ± 5.6 | Control 27.9 ± 5.9 |
| Pathways | Number of Genes with Changes in Methylation | |||
|---|---|---|---|---|
| Hypo ALD | Hyper ALD | Hypo MASLD | Hyper MASLD | |
| AMPK signaling pathway | 4 | 3 | ||
| Amoebiasis | 3 | 6 | ||
| Growth hormone synthesis, secretion and action | 4 | 3 | ||
| Human papillomavirus infection | 5 | 6 | ||
| Lipid and atherosclerosis | 3 | 4 | 3 | |
| Longevity-regulating pathway | 4 | 3 | ||
| Metabolic pathways | 3 | 34 | 6 | 9 |
| MicroRNAs in cancer | 3 | 3 | 4 | |
| Neutrophil extracellular trap formation | 3 | 3 | ||
| Pathogenic Escherichia coli infection | 3 | 3 | ||
| Pathways in cancer * | 7 | 6 | 11 | 7 |
| PI3K Akt signaling pathway | 3 | 3 | 7 | |
| Regulation of actin cytoskeleton | 5 | 3 | ||
| Thermogenesis | 5 | 3 | ||
| Tight junction | 3 | 4 | ||
| Transcriptional dysregulation in cancer | 3 | 3 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stols-Gonçalves, D.; Meijnikman, A.S.; Tristão, L.S.; Santos, C.L.d.; Denswil, N.P.; Verheij, J.; Bernardo, W.M.; Nieuwdorp, M. Metabolic Dysfunction-Associated Steatotic Liver Disease and Alcohol-Associated Liver Disease: Liver DNA Methylation Analysis—A Systematic Review. Cells 2024, 13, 1893. https://doi.org/10.3390/cells13221893
Stols-Gonçalves D, Meijnikman AS, Tristão LS, Santos CLd, Denswil NP, Verheij J, Bernardo WM, Nieuwdorp M. Metabolic Dysfunction-Associated Steatotic Liver Disease and Alcohol-Associated Liver Disease: Liver DNA Methylation Analysis—A Systematic Review. Cells. 2024; 13(22):1893. https://doi.org/10.3390/cells13221893
Chicago/Turabian StyleStols-Gonçalves, Daniela, Abraham S. Meijnikman, Luca Schiliró Tristão, Clara Lucato dos Santos, Nerissa P. Denswil, Joanne Verheij, Wanderley M. Bernardo, and Max Nieuwdorp. 2024. "Metabolic Dysfunction-Associated Steatotic Liver Disease and Alcohol-Associated Liver Disease: Liver DNA Methylation Analysis—A Systematic Review" Cells 13, no. 22: 1893. https://doi.org/10.3390/cells13221893
APA StyleStols-Gonçalves, D., Meijnikman, A. S., Tristão, L. S., Santos, C. L. d., Denswil, N. P., Verheij, J., Bernardo, W. M., & Nieuwdorp, M. (2024). Metabolic Dysfunction-Associated Steatotic Liver Disease and Alcohol-Associated Liver Disease: Liver DNA Methylation Analysis—A Systematic Review. Cells, 13(22), 1893. https://doi.org/10.3390/cells13221893