
Difenoconazole Induced Damage of Bovine Mammary Epithelial Cells via ER Stress and Inflammatory Response


Abstract

1. Introduction
2. Materials and Methods
2.1. Cell Culture and Chemicals
2.2. Cell Viability Assay
2.3. Spheroid Formation and Treatment
2.4. Flow Cytometry
2.5. Immunocytochemistry
2.6. Quantitative PCR (QPCR)
2.7. Western Blotting
2.8. Statistical Analysis
3. Results
3.1. DIF Induced MAC-T Cell Toxicity and Anti-Proliferation
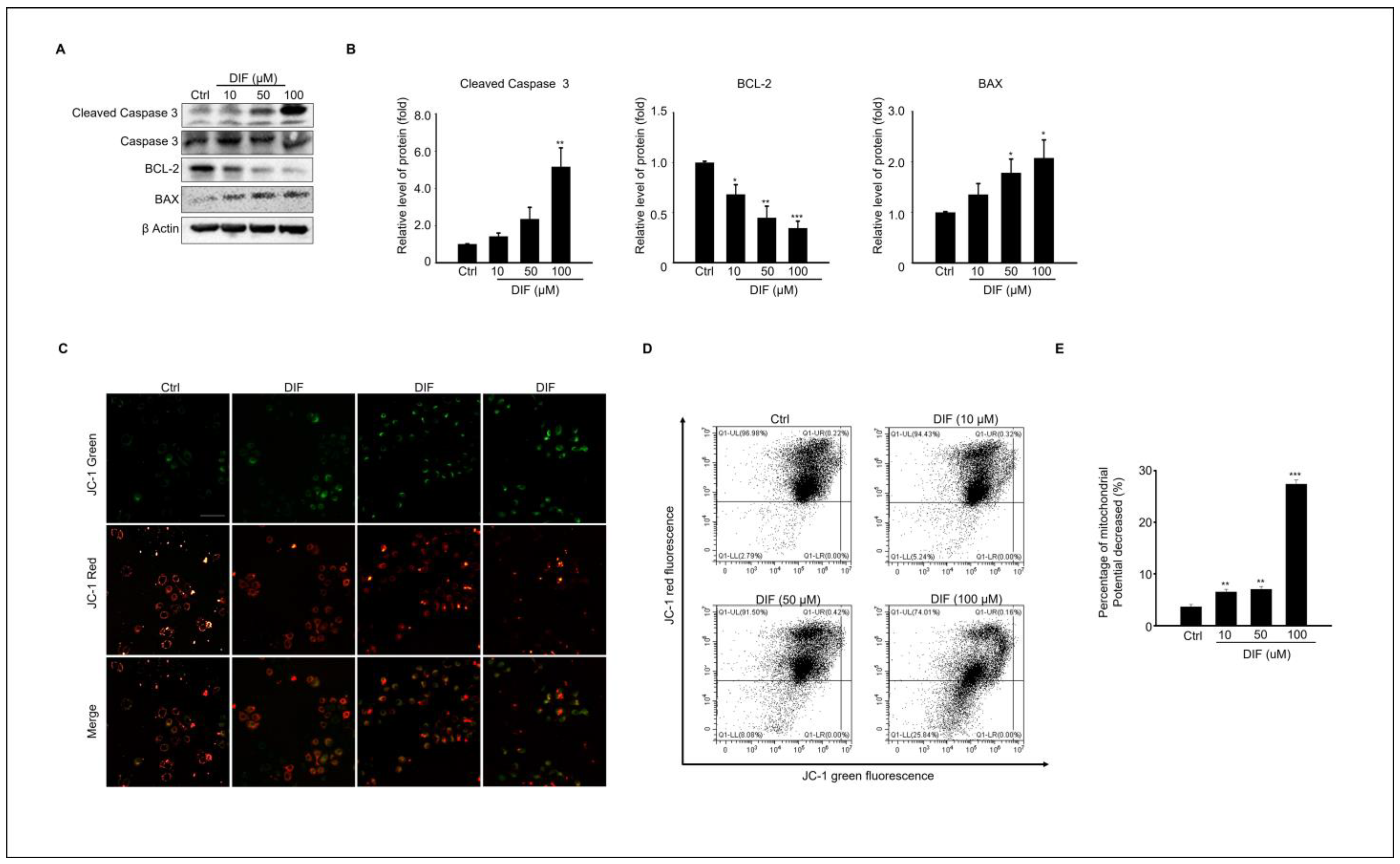
3.2. DIF Induces Apoptosis and Mitochondrial Dysfunction in MAC-T Cells
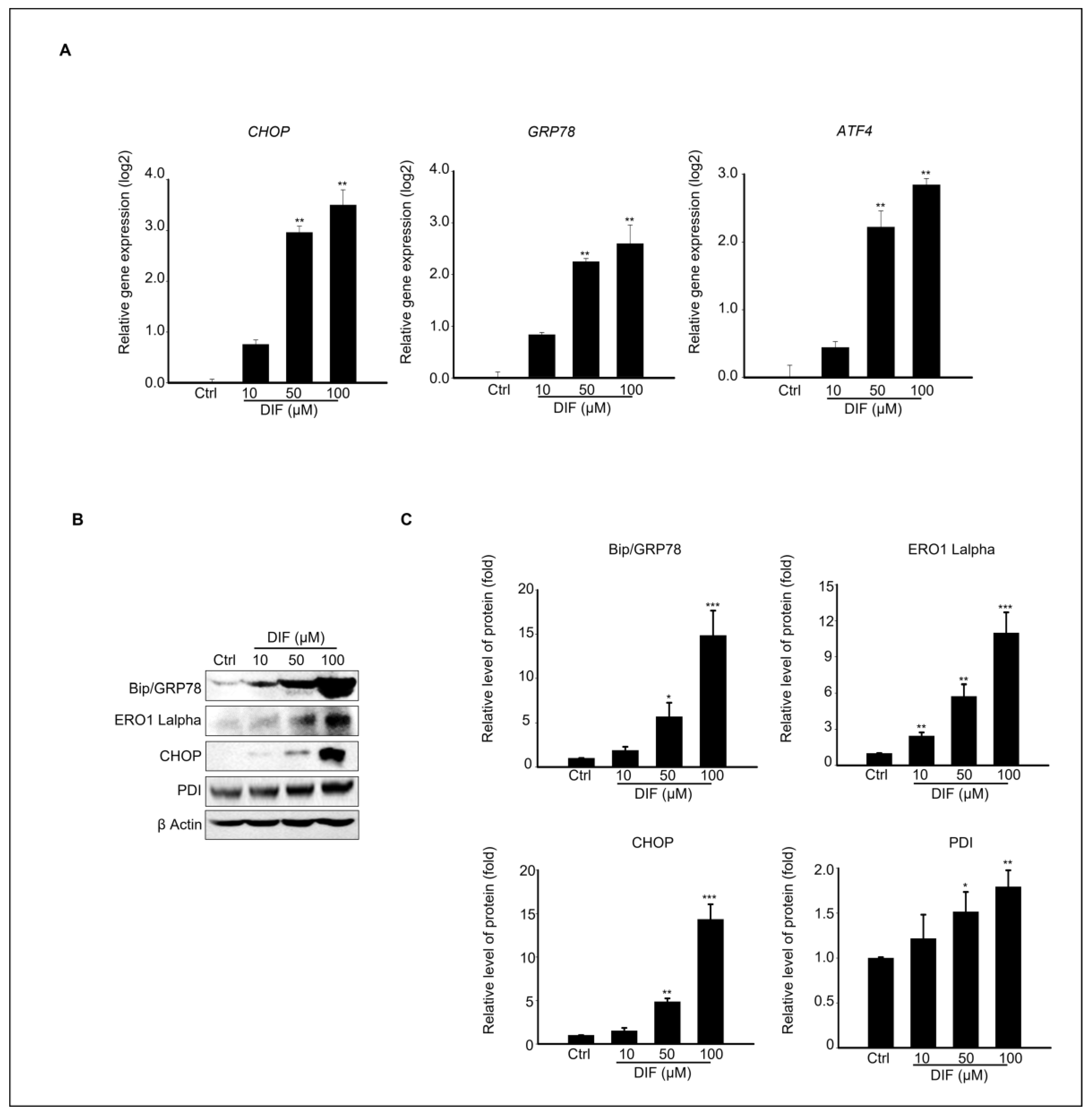
3.3. Endoplasmic Reticulum (ER) Stress-Mediated MAC-T Cell Death Following DIF Treatment
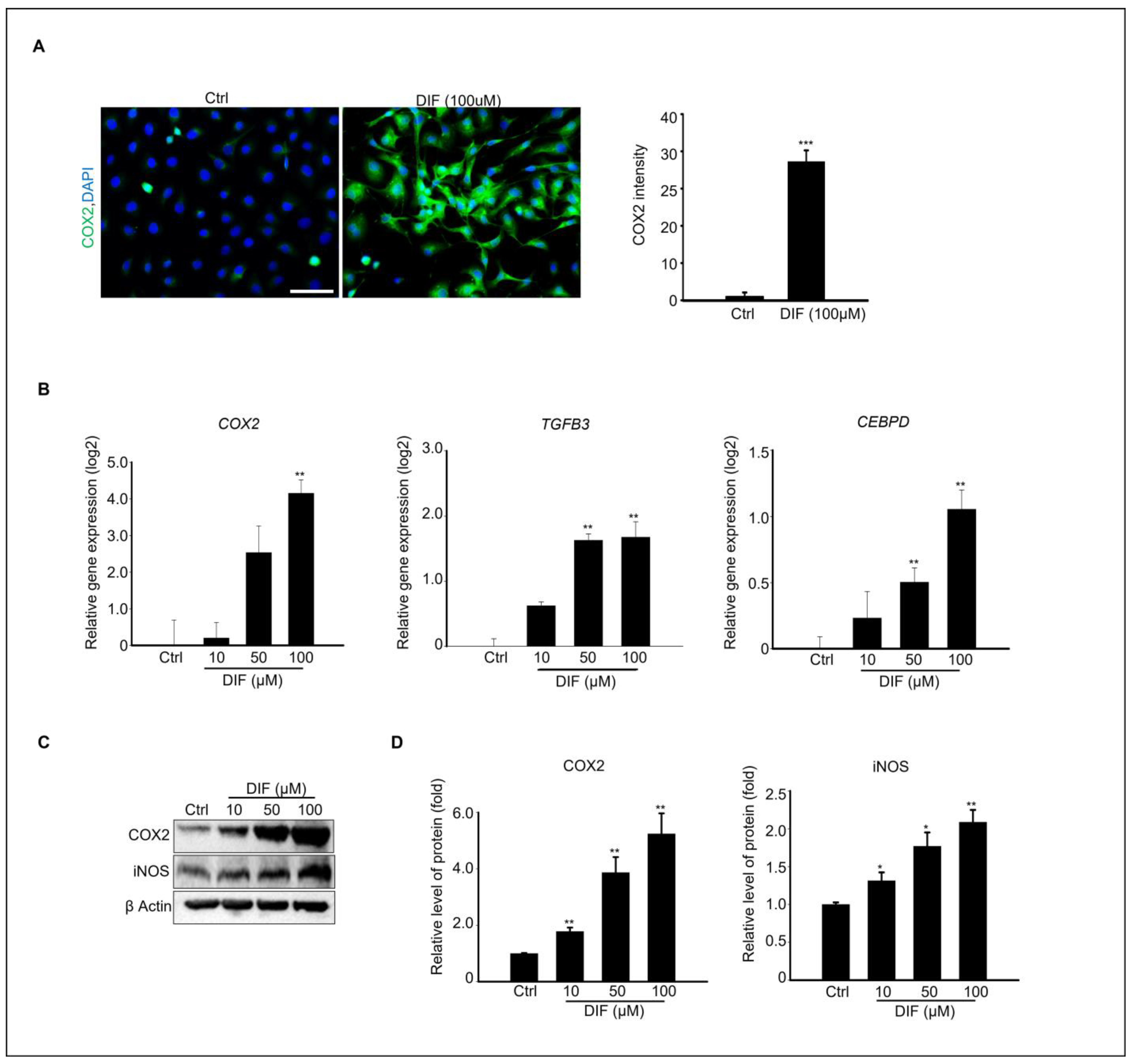
3.4. DIF Induces Inflammatory Responses in MAC-T Cells
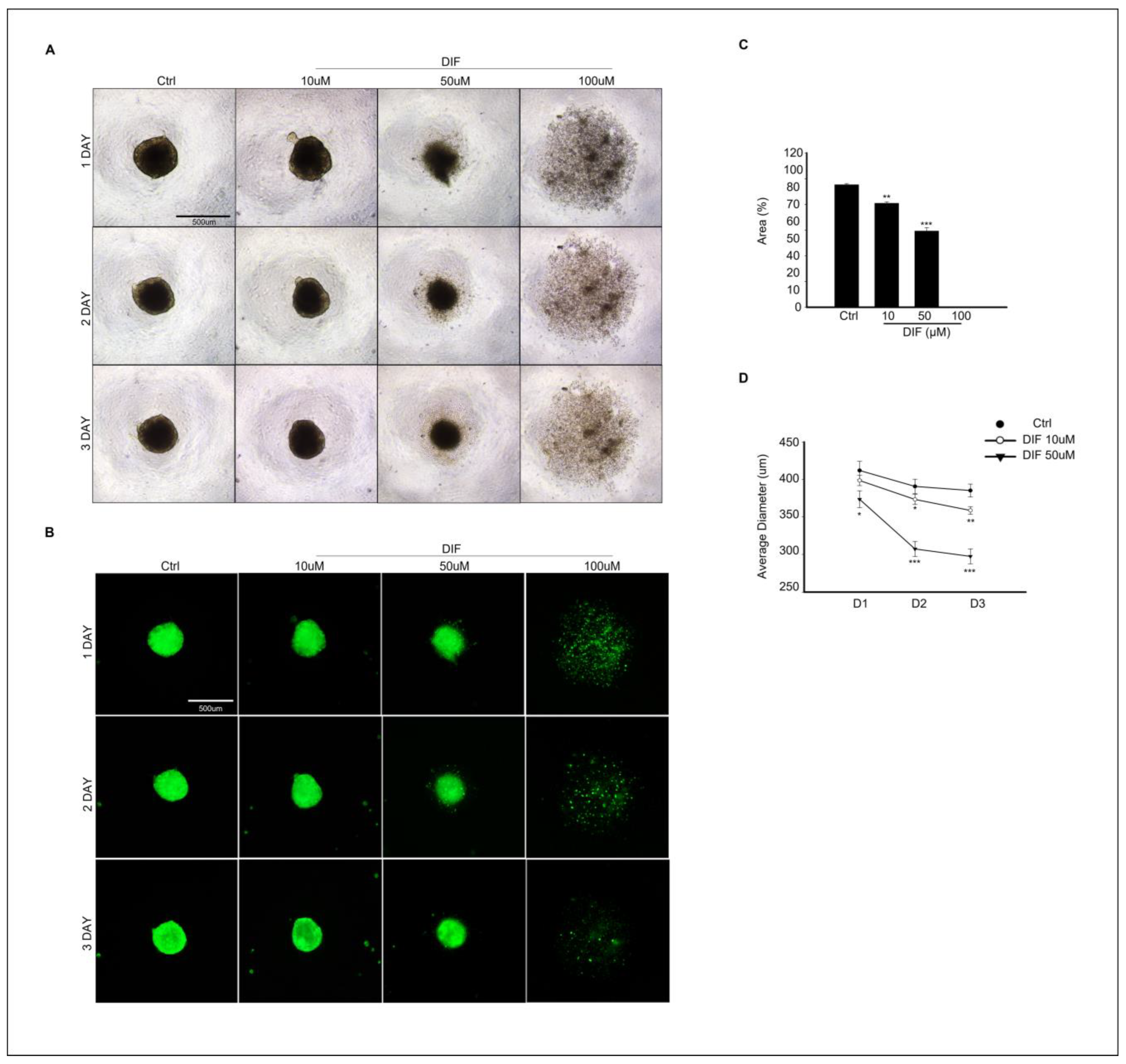
3.5. DIF Treatment Effects on Spheroid Culture of MAC-T Cells
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Machado, F.J.; Santana, F.M.; Lau, D.; Del Ponte, E.M. Quantitative Review of the Effects of Triazole and Benzimidazole Fungicides on Fusarium Head Blight and Wheat Yield in Brazil. Plant Dis. 2017, 101, 1633–1641. [Google Scholar] [CrossRef] [PubMed]
- Rather, I.A.; Sabir, J.S.M.; Asseri, A.H.; Wani, M.Y.; Ahmad, A. Triazole Derivatives Target 14α-Demethylase (LDM) Enzyme in Candida albicans Causing Ergosterol Biosynthesis Inhibition. J. Fungi 2022, 8, 688. [Google Scholar] [CrossRef] [PubMed]
- Brauer, V.S.; Rezende, C.P.; Pessoni, A.M.; De Paula, R.G.; Rangappa, K.S.; Nayaka, S.C.; Gupta, V.K.; Almeida, F. Antifungal Agents in Agriculture: Friends and Foes of Public Health. Biomolecules 2019, 9, 521. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, R.B.; Pettigrove, V.; Rose, G.; Allinson, G.; Wightwick, A.; von der Ohe, P.C.; Shimeta, J.; Kühne, R.; Kefford, B.J. Effects of pesticides monitored with three sampling methods in 24 sites on macroinvertebrates and microorganisms. Environ. Sci. Technol. 2011, 45, 1665–1672. [Google Scholar] [CrossRef]
- Luong, T.T.; Nguyen, T.H.T.; Nguyen, T.D.; Le, V.T.; Pham, T.H.T.; Ho, T.T.; Nguyen, N.L. Degradation of Triazole Fungicides by Plant Growth-Promoting Bacteria from Contaminated Agricultural Soil. J. Microbiol. Biotechnol. 2024, 34, 56–64. [Google Scholar] [CrossRef]
- Pitombeira de Figueirêdo, L.; Athayde, D.B.; Daam, M.A.; van Gestel, C.A.M.; Guerra, G.D.S.; Duarte-Neto, P.J.; Espíndola, E.L.G. Impact of temperature on the toxicity of Kraft 36 EC® (a.s. abamectin) and Score 250 EC® (a.s. difenoconazole) to soil organisms under realistic environmental exposure scenarios. Ecotoxicol. Environ. Saf. 2020, 194, 110446. [Google Scholar] [CrossRef]
- Zhang, H.; Song, J.; Zhang, Z.; Zhang, Q.; Chen, S.; Mei, J.; Yu, Y.; Fang, H. Exposure to fungicide difenoconazole reduces the soil bacterial community diversity and the co-occurrence network complexity. J. Hazard. Mater. 2021, 405, 124208. [Google Scholar] [CrossRef]
- Liu, R.; Li, J.; Zhang, L.; Feng, T.; Zhang, Z.; Zhang, B. Fungicide Difenoconazole Induced Biochemical and Developmental Toxicity in Wheat (Triticum aestivum L.). Plants 2021, 10, 2304. [Google Scholar] [CrossRef]
- Jakl, M.; Ćavar Zeljković, S.; Kovač, I.; Bělonožníková, K.; Jaklová Dytrtová, J. Side effects of triazoles on treated crops. Chemosphere 2021, 277, 130242. [Google Scholar] [CrossRef]
- Mohapatra, S. Dynamics of Difenoconazole and Propiconazole Residues on Pomegranate over 2 years under Field Conditions. Environ. Sci. Pollut. Res. Int. 2016, 23, 5795–5806. [Google Scholar] [CrossRef]
- Mu, X.; Chai, T.; Wang, K.; Zhang, J.; Zhu, L.; Li, X.; Wang, C. Occurrence and Origin of Sensitivity toward Difenoconazole in Zebrafish (Danio reio) during different Life Stages. Aquat. Toxicol. 2015, 160, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Chen, L.; Wu, S.; Lv, L.; Liu, X.; Wang, Q.; Zhao, X. Effects of Difenoconazole on Hepatotoxicity, Lipid Metabolism and Gut Microbiota in Zebrafish (Danio rerio). Environ. Pollut. 2020, 265, 114844. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Deng, P.; Xing, D.; Liu, H.; Shi, F.; Hu, L.; Zou, X.; Nie, H.; Zuo, J.; Zhuang, Z.; et al. Developmental Neurotoxicity of Difenoconazole in Zebrafish Embryos. Toxics 2023, 11, 353. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, X.; Bello, B.K.; Zhang, T.; Yang, H.; Wang, K.; Dong, J. Difenoconazole causes spleen tissue damage and immune dysfunction of carp through oxidative stress and apoptosis. Ecotoxicol. Environ. Saf. 2022, 237, 113563. [Google Scholar] [CrossRef]
- Mu, X.; Chai, T.; Wang, K.; Zhu, L.; Huang, Y.; Shen, G.; Li, Y.; Li, X.; Wang, C. The developmental effect of difenoconazole on zebrafish embryos: A mechanism research. Environ. Pollut. 2016, 212, 18–26. [Google Scholar] [CrossRef]
- Zhang, H.; Qian, M.; Wang, J.; Yang, G.; Weng, Y.; Jin, C.; Li, Y.; Jin, Y. Insights into the effects of difenoconazole on the livers in male mice at the biochemical and transcriptomic levels. J. Hazard. Mater. 2022, 422, 126933. [Google Scholar] [CrossRef]
- Bao, Z.; Wang, W.; Wang, X.; Qian, M.; Jin, Y. Sub-Chronic Difenoconazole Exposure Induced Gut Microbiota Dysbiosis in Mice. Toxics 2022, 10, 34. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, G.; Bao, Z.; Jin, Y.; Wang, J.; Chen, J.; Qian, M. Stereoselective effects of fungicide difenoconazole and its four stereoisomers on gut barrier, microbiota, and glucolipid metabolism in male mice. Sci. Total Environ. 2022, 805, 150454. [Google Scholar] [CrossRef]
- Zheng, X.; Wei, Y.; Chen, J.; Wang, X.; Li, D.; Yu, C.; Hong, Y.; Shen, L.; Long, C.; Wei, G.; et al. Difenoconazole Exposure Induces Retinoic Acid Signaling Dysregulation and Testicular Injury in Mice Testes. Toxics 2023, 11, 328. [Google Scholar] [CrossRef]
- Gupta, U.K.; Kumar, S.; Ray, S.; Gupta, N.K.; Maurya, G. Bioaccumulation of pesticides in fish resulting toxicities in humans through food chain and forensic aspects. Acta Med. Philipp. 2024, 58, 94–98. [Google Scholar]
- Hamilton, D.; Ambrus, A.; Dieterle, R.; Felsot, A.; Harris, C.; Petersen, B.; Racke, K.; Wong, S.S.; Gonzalez, R.; Tanaka, K.; et al. Advisory Committee on Crop Protection Chemistry, Division of Chemistry and the Environment; of the International Union of Pure and Applied Chemistry. Pesticide residues in food--acute dietary exposure. Pest. Manag. Sci. 2004, 60, 311–339. [Google Scholar] [CrossRef] [PubMed]
- Bruinenberg, M.; van Agtmaal, M.; Hoekstra, N.; van Eekeren, N. Residues of pesticides in dairy cow rations and fly treatments reduce the number of Coleoptera in dung. Agric. Ecosyst. Environ. 2023, 344, 108307. [Google Scholar] [CrossRef]
- Boudebbouz, A.; Boudalia, S.; Bousbia, A.; Habila, S.; Boussadia, M.I.; Gueroui, Y. Pesticide residues levels in raw cow’s milk and health risk assessment across the globe: A systematic review. Sci. Total Environ. 2021, 751, 141830. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.A.; Song, J.; Ham, J.; An, G.; Song, G.; Lim, W. Tetraconazole interrupts mitochondrial function and intracellular calcium levels leading to apoptosis of bovine mammary epithelial cells. Pestic. Biochem. Physiol. 2023, 191, 105366. [Google Scholar] [CrossRef] [PubMed]
- You, H.; An, G.; Lee, H.; Lim, W.; Song, G. Bifenox induces programmed cell death in bovine mammary epithelial cells by impairing calcium homeostasis, triggering ER stress, and altering the signaling cascades of PI3K/AKT and MAPK. Pestic. Biochem. Physiol. 2023, 196, 105626. [Google Scholar] [CrossRef]
- Sung, E.; Park, J.; Lee, H.; Song, G.; Lim, W. Bifenthrin induces cell death in bovine mammary epithelial cells via ROS generation, calcium ion homeostasis disruption, and MAPK signaling cascade alteration. Pestic. Biochem. Physiol. 2023, 196, 105637. [Google Scholar] [CrossRef]
- Lee, W.Y.; Lee, R.; Park, H.J. Tebuconazole Induces ER-Stress-Mediated Cell Death in Bovine Mammary Epithelial Cell Lines. Toxics 2023, 11, 397. [Google Scholar] [CrossRef]
- Wang, C.; Youle, R.J. The role of mitochondria in apoptosis. Annu. Rev. Genet. 2009, 43, 95–118. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Guo, Y.; Tang, J.; Jiang, J.; Chen, Z. New insights into the roles of CHOP-induced apoptosis in ER stress. Acta Biochim. Biophys. Sin. 2014, 46, 629–640. [Google Scholar] [CrossRef]
- Xue, X.; Piao, J.H.; Nakajima, A.; Sakon-Komazawa, S.; Kojima, Y.; Mori, K.; Yagita, H.; Okumura, K.; Harding, H.; Nakano, H. Tumor necrosis factor alpha (TNFalpha) induces the unfolded protein response (UPR) in a reactive oxygen species (ROS)-dependent fashion, and the UPR counteracts ROS accumulation by TNFalpha. J. Biol. Chem. 2005, 280, 33917–33925. [Google Scholar] [CrossRef]
- Kahle, M.; Buerge, I.J.; Hauser, A.; Müller, M.D.; Poiger, T. Azole fungicides: Occurrence and fate in wastewater and surface waters. Environ. Sci. Technol. 2008, 42, 7193–7200. [Google Scholar] [CrossRef] [PubMed]
- Teng, M.; Qi, S.; Zhu, W.; Wang, Y.; Wang, D.; Dong, K.; Wang, C. Effects of the bioconcentration and parental transfer of environmentally relevant concentrations of difenoconazole on endocrine disruption in zebrafish (Danio rerio). Environ. Pollut. 2018, 233, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; An, G.; Lim, W.; Song, G. Aclonifen induces bovine mammary gland epithelial cell death by disrupting calcium homeostasis and inducing ROS production. Pestic. Biochem. Physiol. 2022, 181, 105011. [Google Scholar] [CrossRef]
- Vaux, D.L.; Korsmeyer, S.J. Cell death in development. Cell 1999, 96, 245–254. [Google Scholar] [CrossRef]
- Chen, Q.; Kang, J.; Fu, C. The independence of and associations among apoptosis, autophagy, and necrosis. Signal Transduct. Target. Ther. 2018, 3, 18. [Google Scholar] [CrossRef]
- Kashyap, D.; Garg, V.K.; Goel, N. Intrinsic and extrinsic pathways of apoptosis: Role in cancer development and prognosis. Adv. Protein Chem. Struct. Biol. 2021, 125, 73–120. [Google Scholar]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef]
- Popgeorgiev, N.; Jabbour, L.; Gillet, G. Subcellular Localization and Dynamics of the Bcl-2 Family of Proteins. Front. Cell Dev. Biol. 2018, 6, 13. [Google Scholar] [CrossRef]
- Czabotar, P.E.; Garcia-Saez, A.J. Mechanisms of BCL-2 family proteins in mitochondrial apoptosis. Nat. Rev. Mol. Cell Biol. 2023, 24, 732–748. [Google Scholar] [CrossRef]
- Bagkos, G.; Koufopoulos, K.; Piperi, C. A new model for mitochondrial membrane potential production and storage. Med. Hypotheses 2014, 83, 175–181. [Google Scholar] [CrossRef]
- Zorova, L.D.; Popkov, V.A.; Plotnikov, E.Y.; Silachev, D.N.; Pevzner, I.B.; Jankauskas, S.S.; Babenko, V.A.; Zorov, S.D.; Balakireva, A.V.; Juhaszova, M.; et al. Mitochondrial membrane potential. Anal. Biochem. 2018, 552, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Kroemer, G.; Galluzzi, L.; Brenner, C. Mitochondrial membrane permeabilization in cell death. Physiol. Rev. 2007, 87, 99–163. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, P.; Castedo, M.; Susin, S.A.; Zamzami, N.; Hirsch, T.; Macho, A.; Haeffner, A.; Hirsch, F.; Geuskens, M.; Kroemer, G. Mitochondrial permeability transition is a central coordinating event of apoptosis. J. Exp. Med. 1996, 184, 1155–1160. [Google Scholar] [CrossRef]
- Fulda, S.; Gorman, A.M.; Hori, O.; Samali, A. Cellular Stress Responses: Cell Survival and Cell Death. Int. J. Cell Biol. 2010, 2010, 214074. [Google Scholar] [CrossRef]
- Fawcett, T.W.; Martindale, J.L.; Guyton, K.Z.; Hai, T.; Holbrook, N.J. Complexes containing activating transcription factor (ATF)/cAMP-responsiveelement-binding protein (CREB) interact with the CCAAT/enhancer-binding protein (C/EBP)–ATF composite site to regulate Gadd153 expression during the stress response. Biochem. J. 1999, 339, 135–141. [Google Scholar] [CrossRef]
- Sano, R.; Reed, J.C. ER stress-induced cell death mechanisms. Biochim. Biophys. Acta 2013, 1833, 3460–3470. [Google Scholar] [CrossRef]
- Zhang, J.; Guo, J.; Yang, N.; Huang, Y.; Hu, T.; Rao, C. Endoplasmic reticulum stress-mediated cell death in liver injury. Cell Death Dis. 2022, 13, 1051. [Google Scholar] [CrossRef]
- Lin, J.H.; Walter, P.; Yen, T.S. Endoplasmic reticulum stress in disease pathogenesis. Annu. Rev. Pathol. 2008, 3, 399–425. [Google Scholar] [CrossRef]
- Oslowski, C.M.; Urano, F. Measuring ER stress and the unfolded protein response using mammalian tissue culture system. Methods Enzymol. 2011, 490, 71–92. [Google Scholar]
- Ferreiro, E.; Oliveira, C.R.; Pereira, C.M.F. The release of calcium from the endoplasmic reticulum induced by amyloid-beta and prion peptides activates the mitochondrial apoptotic pathway. Neurobiol. Dis. 2008, 30, 331–342. [Google Scholar] [CrossRef]
- Mekahli, D.; Bultynck, G.; Parys, J.B.; De Smedt, H.; Missiaen, L. Endoplasmic-reticulum calcium depletion and disease. Cold Spring Harb. Perspect. Biol. 2011, 3, a004317. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Giorgi, C.; Siviero, R.; Zecchini, E.; Rizzuto, R. Calcium and apoptosis: ER-mitochondria Ca2+ transfer in the control of apoptosis. Oncogene 2008, 27, 6407–6418. [Google Scholar] [CrossRef] [PubMed]
- Walter, P.; Ron, D. The Unfolded Protein Response: From Stress Pathway to Homeostatic Regulation. Science 2011, 334, 1081–1086. [Google Scholar] [CrossRef]
- Kim, I.; Xu, W.; Reed, J.C. Cell death and endoplasmic reticulum stress: Disease relevance and therapeutic opportunities. Nat. Rev. Drug Discov. 2008, 7, 1013–1030. [Google Scholar] [CrossRef]
- Almanza, A.; Carlesso, A.; Chintha, C.; Creedican, S.; Doultsinos, D.; Leuzzi, B.; Luís, A.; McCarthy, N.; Montibeller, L.; More, S.; et al. Endoplasmic reticulum stress signalling—From basic mechanisms to clinical applications. FEBS J. 2019, 286, 241–278. [Google Scholar] [CrossRef]
- Chiu, K.W.; Chen, H.Y.; Chen, C.L.; Hsieh, C.P.; Huang, Y.F. Attenuation of Endoplasmic Reticulum Stress Enhances Carvacrol-Induced Apoptosis in Osteosarcoma Cell Lines. Life 2023, 13, 744. [Google Scholar] [CrossRef] [PubMed]
- Rainard, P.; Gilbert, F.B.; Germon, P. Immune defenses of the mammary gland epithelium of dairy ruminants. Front. Immunol. 2022, 13, 1031785. [Google Scholar] [CrossRef]
- Simon, L.S. Role and regulation of cyclooxygenase-2 during inflammation. Am. J. Med. 1999, 106, 37S–42S. [Google Scholar] [CrossRef]
- Surh, Y.J.; Chun, K.S.; Cha, H.H.; Han, S.S.; Keum, Y.S.; Park, K.K.; Lee, S.S. Molecular mechanisms underlying chemopreventive activities of anti-inflammatory phytochemicals: Down-regulation of COX-2 and iNOS through suppression of NF-κB activation. Mutat. Res. 2001, 480–481, 243–268. [Google Scholar] [CrossRef]
- Thomas, B.; Berenbaum, F.; Humbert, L.; Bian, H.; Béréziat, G.; Crofford, L.; Olivier, J.L. Critical role of C/EBPδ and C/EBPβ factors in the stimulation of the cyclooxygenase-2 gene transcription by interleukin-1β in articular chondrocytes. Eur. J. Biochem. 2000, 267, 6798–6809. [Google Scholar] [CrossRef]
- Huynh, M.L.; Fadok, V.A.; Henson, P.M. Phosphatidylserine-dependent ingestion of apoptotic cells promotes TGF-beta1 secretion and the resolution of inflammation. J. Clin. Investig. 2002, 109, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Pasparakis, M.; Vandenabeele, P. Necroptosis and its role in inflammation. Nature 2015, 517, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Haanen, C.; Vermes, I. Apoptosis and inflammation. Mediat. Inflamm. 1995, 4, 5–15. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Manufacturer | Catalog Number | Dilution (WB) | Dilution (ICC) |
|---|---|---|---|---|
| β-actin | Santa Cruz Biotech | SC-47778 | 1:1000 | |
| Bcl-2 | Cell Signaling | 15071S | 1:2000 | |
| Cleaved-caspase 3 | Cell Signaling | 9661S | 1:2000 | |
| Caspase 3 | Santa Cruz | SC-22171 | 1:2000 | |
| BAX | Santa Cruz | SC-7480 | 1:2000 | |
| BIP/GRP78 | Cell Signaling | 3177T | 1:2000 | |
| ERO-La | Cell Signaling | 3264T | 1:2000 | |
| PDI | Cell Signaling | 3501S | 1:2000 | |
| CHOP | Cell Signaling | 2895S | 1:2000 | |
| iNOS | Cell Signaling | 13120 | 1:2000 | |
| COX2 | Cell Signaling | 12282 | 1:2000 | 1:200 |
| Ki 67 | Abcam | Ab15580 | 1:200 |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| GAPDH | 5′-GGGTCATCATCTCTGCACCT-3′ | 5′-GGTCATAAGTCCCTCCACGA-3′ |
| COX2 | 5′-TCCTGAAACCCACTCCCAACA-3′ | 5′-TGGGCAGTCATCAGGCACAG-3′ |
| Tgfb3 | 5′-TCTGGGGCGACTTAAGAAGA-3′ | 5′-ATTGCGGAAGCAGTAATTGG-3′ |
| CEBPD | 5′-ATCGACTTCAGCGCCTACAT-3′ | 5′-TGTGGTTGCTGTTGAAGAGG-3′ |
| CHOP | 5′-GCAACGCATGAAGGAGAAAG-3′ | 5′-AACCATCCGGTCAATCAGAG-3′ |
| Grp78 | 5′-TGGCTGGAAAGTCACCAAG-3′ | 5′-GTCTGCTGCTTCCTCCTCAC-3′ |
| ATF4 | 5′-GCTGTGGATTGGTTGGTCTC-3′ | 5′-AGCTCATCTGGCATGGTTTC-3′ |
| Nrf2 | 5′-AGGACATGGATTTGATTGAC-3′ | 5′-TACCTGGGAGTAGTTGGCA-3′ |
| HO-1 | 5′-GGTGATGGCGTCTTTGTACC-3′ | 5′-GCAGCTCCTCTGGGAAGTAG-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Na, M.-J.; Lee, W.-Y.; Park, H.-J. Difenoconazole Induced Damage of Bovine Mammary Epithelial Cells via ER Stress and Inflammatory Response. Cells 2024, 13, 1715. https://doi.org/10.3390/cells13201715
Na M-J, Lee W-Y, Park H-J. Difenoconazole Induced Damage of Bovine Mammary Epithelial Cells via ER Stress and Inflammatory Response. Cells. 2024; 13(20):1715. https://doi.org/10.3390/cells13201715
Chicago/Turabian StyleNa, Myoung-Jun, Won-Young Lee, and Hyun-Jung Park. 2024. "Difenoconazole Induced Damage of Bovine Mammary Epithelial Cells via ER Stress and Inflammatory Response" Cells 13, no. 20: 1715. https://doi.org/10.3390/cells13201715
APA StyleNa, M.-J., Lee, W.-Y., & Park, H.-J. (2024). Difenoconazole Induced Damage of Bovine Mammary Epithelial Cells via ER Stress and Inflammatory Response. Cells, 13(20), 1715. https://doi.org/10.3390/cells13201715