
Housekeeping Gene Stability in Adipose Mesenchymal Stromal Cells Cultivated in Serum/Xeno-Free Media for Osteoarthritis

 ,
, 
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Adipose Mesenchymal Stromal Cells (ASCs) Isolation and Culture
- (i)
- DMEM/F12 + 10% FBS (GE Healthcare, Piscataway, NJ, USA), 1% L-glutamine plus penicillin–streptomycin (PSG) (Life Technologies, Carlsbad, CA, USA). This condition will be named hereafter (F);
- (ii)
- StemPro™ MSC SFM XenoFree (xeno-free, cGMP compliant) (ThermoFisher), 1% PSG. Before seeding, flasks were coated with CELLstart™ Substrate (xeno-free, cGMP compliant) (ThermoFisher) as per manufacturer’s instruction to enhance cell adhesion and growth in absence of serum. Condition is named (X1);
- (iii)
- StemFit® For Mesenchymal Stem Cells (xeno-free) (Amsbio, Cambridge, MA, USA), 1% PSG. Before seeding, flasks were coated with iMatrix-511 expressed in CHO cells for easier translation into GMP (Amsbio) as per manufacturer’s instruction to enhance cell adhesion and growth in absence of serum. iMatrix-511 is comprised of recombinant Laminin-511 E8 protein fragments. Condition is named (X2).
2.3. ASCs Immunophenotype by Flow Cytometry
2.4. RNA Extraction and mRNA Profiling
2.5. Data Analysis
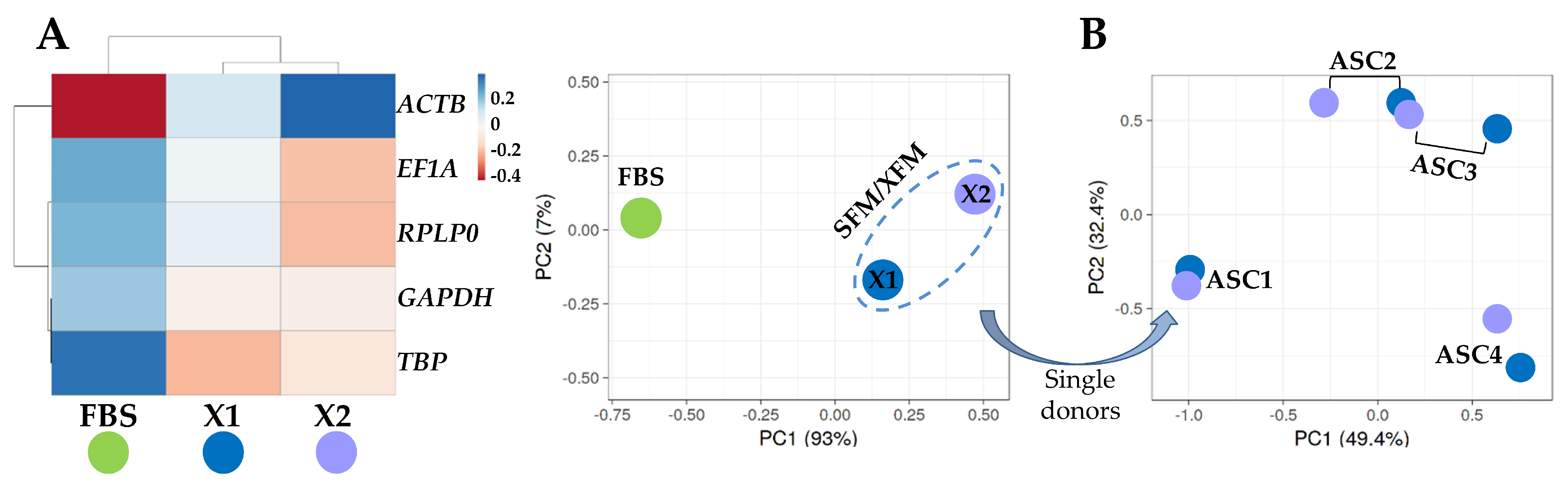
2.6. Hierarchical Clustering and Principal Component Analysis
2.7. Statistical Analyses
3. Results
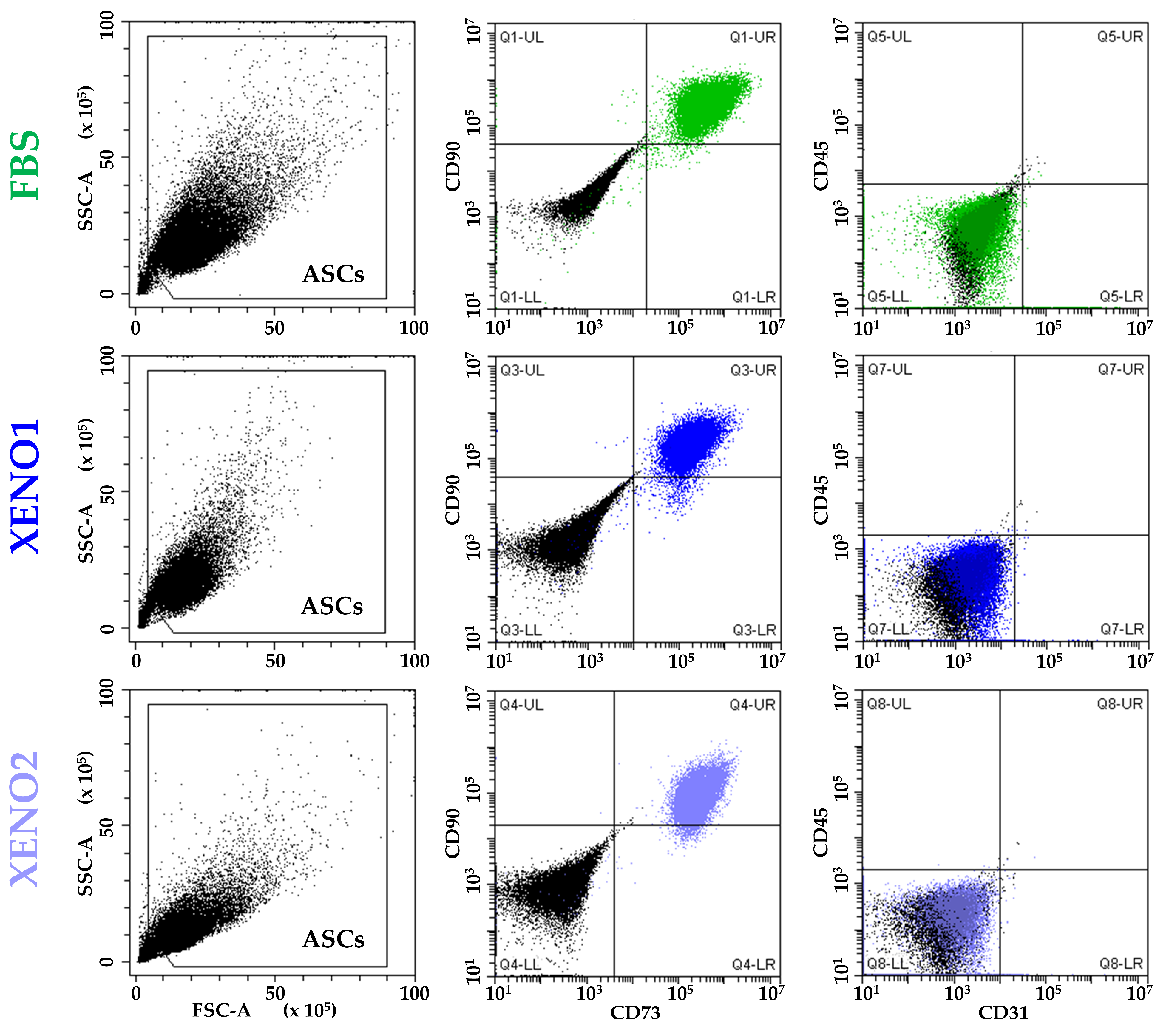
3.1. ASCs Characterization
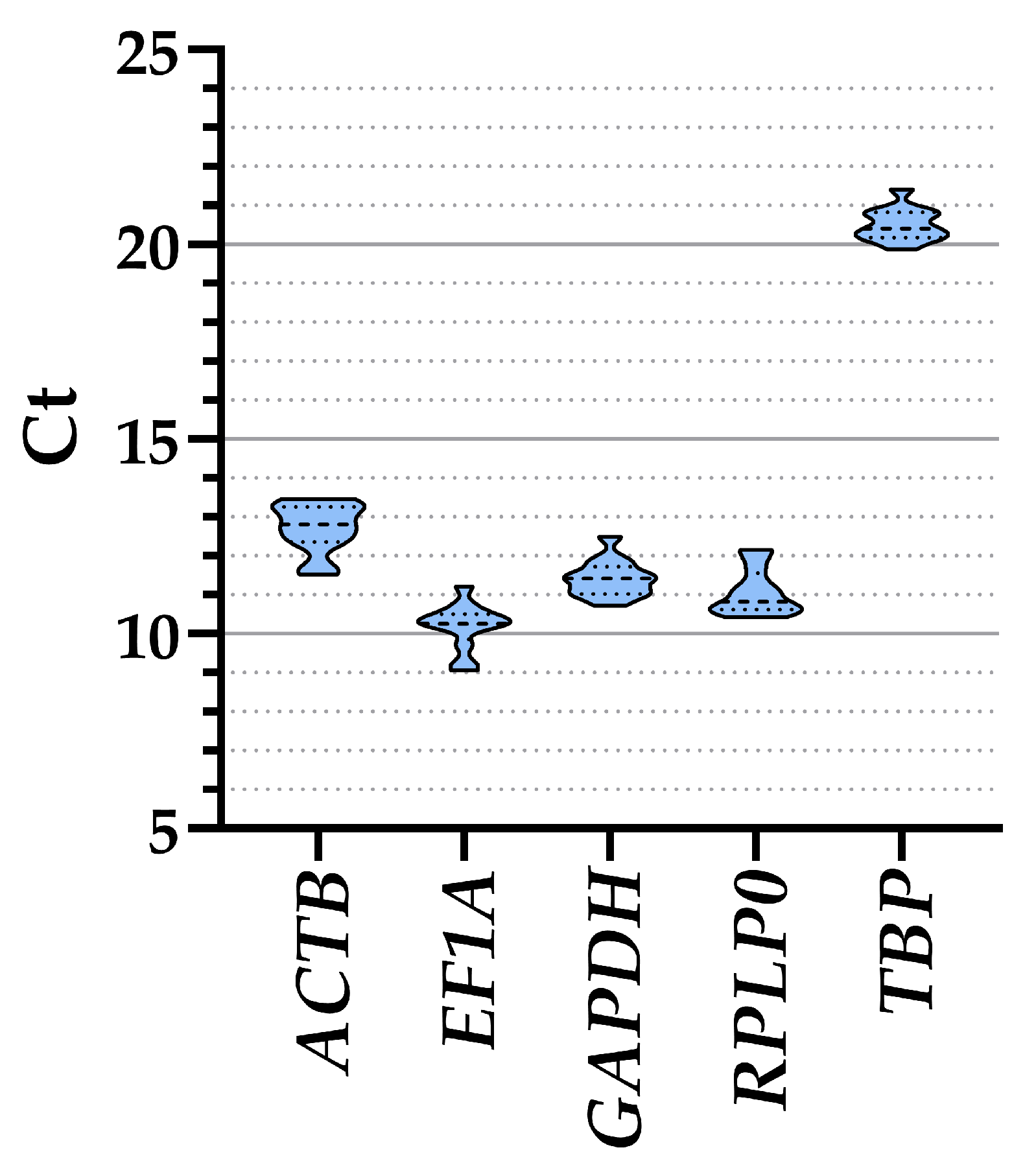
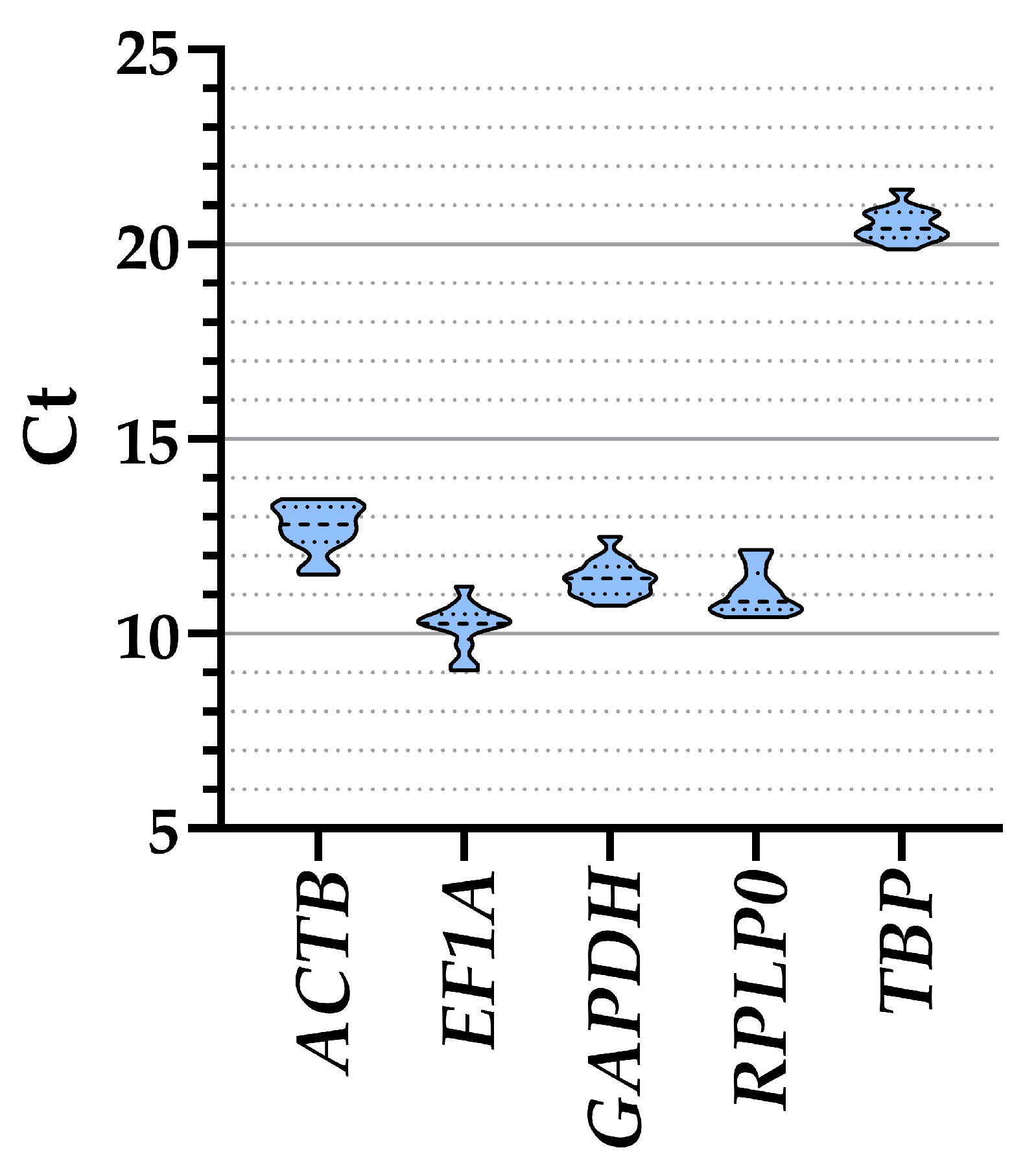
3.2. Candidate HKGs Expression
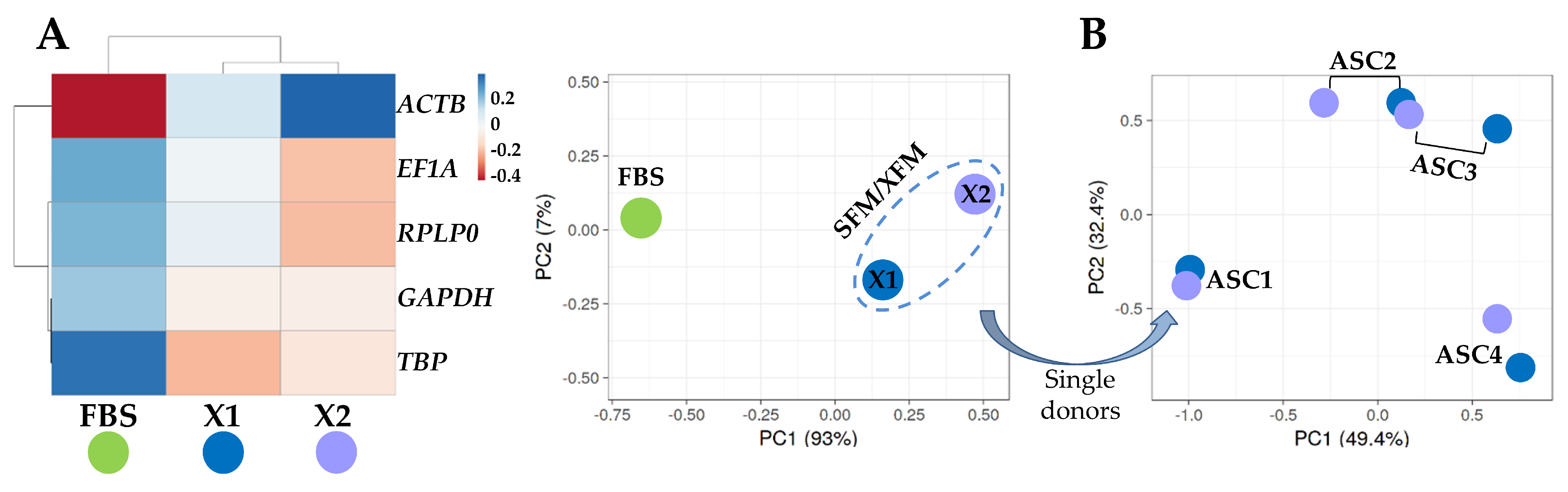
3.3. Analysis of HKGs Stability
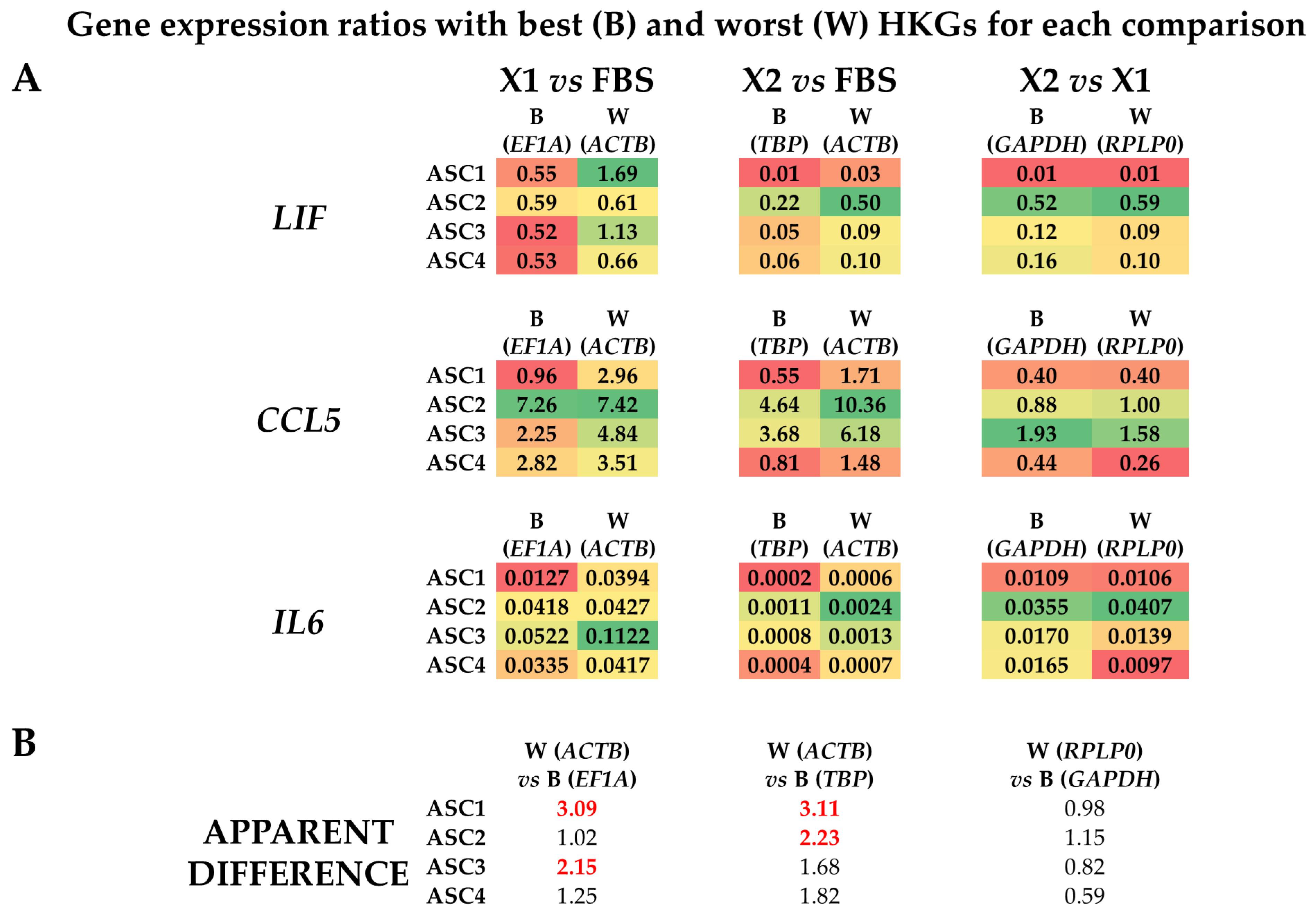
3.4. Effect of HKGs Choice on Target Gene Expression Evaluation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hunter, D.J.; Bierma-Zeinstra, S. Osteoarthritis. Lancet 2019, 393, 1745–1759. [Google Scholar] [CrossRef] [PubMed]
- Martel-Pelletier, J.; Wildi, L.M.; Pelletier, J. Future therapeutics for osteoarthritis. Bone 2012, 51, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Uivaraseanu, B.; Vesa, C.M.; Tit, D.M.; Abid, A.; Maghiar, O.; Maghiar, T.A.; Hozan, C.; Nechifor, A.C.; Behl, T.; Patrascu, J.M.; et al. Therapeutic approaches in the management of knee osteoarthritis (Review). Exp. Ther. Med. 2022, 23, 328. [Google Scholar] [CrossRef] [PubMed]
- Herman, K.; Gobbi, A. Evidence-Based Approach to Orthobiologics for Osteoarthritis and Other Joint Disorders. Phys. Med. Rehabil. Clin. N. Am. 2023, 34, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhao, Y.; Sun, X.; Xing, Y.; Wang, X.; Yang, Q. Immunomodulation of MSCs and MSC-Derived Extracellular Vesicles in Osteoarthritis. Front. Bioeng. Biotechnol. 2020, 8, 575057. [Google Scholar] [CrossRef]
- Xiang, X.; Zhu, S.; He, H.; Yu, X.; Xu, Y.; He, C. Mesenchymal stromal cell-based therapy for cartilage regeneration in knee osteoarthritis. Stem Cell Res. Ther. 2022, 13, 14. [Google Scholar] [CrossRef]
- Fraser, J.K.; Zhu, M.; Zeni, I.W.; Alfonso, Z. Adipose-derived stem cells. Methods Mol. Biol. 2008, 449, 59–67. [Google Scholar] [CrossRef]
- Song, Y.; Jorgensen, C. Mesenchymal Stromal Cells in Osteoarthritis: Evidence for Structural Benefit and Cartilage Repair. Biomedicines 2022, 10, 1278. [Google Scholar] [CrossRef]
- de Carvalho Carneiro, D.; de Araújo, L.T.; Santos, G.C.; Damasceno, P.K.F.; Vieira, J.L.; Santos, R.R.D.; Barbosa, J.D.V.; Soares, M.B.P. Clinical Trials with Mesenchymal Stem Cell Therapies for Osteoarthritis: Challenges in the Regeneration of Articular Cartilage. Int. J. Mol. Sci. 2023, 24, 9939. [Google Scholar] [CrossRef]
- Czapla, J.; Matuszczak, S.; Kulik, K.; Wiśniewska, E.; Pilny, E.; Jarosz-Biej, M.; Smolarczyk, R.; Sirek, T.; Zembala, M.O.; Zembala, M.; et al. The effect of culture media on large-scale expansion and characteristic of adipose tissue-derived mesenchymal stromal cells. Stem Cell Res. Ther. 2019, 10, 235. [Google Scholar] [CrossRef]
- Nikolits, I.; Nebel, S.; Egger, D.; Kreß, S.; Kasper, C. Towards Physiologic Culture Approaches to Improve Standard Cultivation of Mesenchymal Stem Cells. Cells 2021, 10, 886. [Google Scholar] [CrossRef] [PubMed]
- Bui, H.T.H.; Nguyen, L.T.; Than, U.T.T. Influences of Xeno-Free Media on Mesenchymal Stem Cell Expansion for Clinical Application. Tissue Eng. Regen. Med. 2021, 18, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Cimino, M.; Gonçalves, R.M.; Barrias, C.C.; Martins, M.C.L. Xeno-Free Strategies for Safe Human Mesenchymal Stem/Stromal Cell Expansion: Supplements and Coatings. Stem Cells Int. 2017, 2017, 6597815. [Google Scholar] [CrossRef] [PubMed]
- Lindroos, B.; Boucher, S.; Chase, L.; Kuokkanen, H.; Huhtala, H.; Haataja, R.; Vemuri, M.; Suuronen, R.; Miettinen, S. Serum-free, xeno-free culture media maintain the proliferation rate and multipotentiality of adipose stem cells in vitro. Cytotherapy 2009, 11, 958–972. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Pannier, L.; Merino, E.; Marchal, K.; Collado-Vides, J. Effect of genomic distance on coexpression of coregulated genes in E. coli. PLoS ONE 2017, 12, e0174887. [Google Scholar] [CrossRef]
- Sanz-Nogués, C.; O’Brien, T. Current good manufacturing practice considerations for mesenchymal stromal cells as therapeutic agents. Biomater. Biosyst. 2021, 2, 100018. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Yang, W.; Li, Y.; Sun, C. Fetal bovine serum, an important factor affecting the reproducibility of cell experiments. Sci. Rep. 2023, 13, 1942. [Google Scholar] [CrossRef] [PubMed]
- Oeller, M.; Laner-Plamberger, S.; Krisch, L.; Rohde, E.; Strunk, D.; Schallmoser, K. Human Platelet Lysate for Good Manufacturing Practice-Compliant Cell Production. Int. J. Mol. Sci. 2021, 22, 5178. [Google Scholar] [CrossRef]
- Kang, J.Y.; Oh, M.; Joo, H.; Park, H.S.; Chae, D.; Kim, J.; Lee, H.; Oh, I.; Yu, K. Xeno-Free Condition Enhances Therapeutic Functions of Human Wharton’s Jelly-Derived Mesenchymal Stem Cells against Experimental Colitis by Upregulated Indoleamine 2,3-Dioxygenase Activity. J. Clin. Med. 2020, 9, 2913. [Google Scholar] [CrossRef]
- Mark, P.; Kleinsorge, M.; Gaebel, R.; Lux, C.A.; Toelk, A.; Pittermann, E.; David, R.; Steinhoff, G.; Ma, N. Human Mesenchymal Stem Cells Display Reduced Expression of CD105 after Culture in Serum-Free Medium. Stem Cells Int. 2013, 2013, 698076. [Google Scholar] [CrossRef]
- Bobis-Wozowicz, S.; Kmiotek, K.; Kania, K.; Karnas, E.; Labedz-Maslowska, A.; Sekula, M.; Kedracka-Krok, S.; Kolcz, J.; Boruczkowski, D.; Madeja, Z.; et al. Diverse impact of xeno-free conditions on biological and regenerative properties of hUC-MSCs and their extracellular vesicles. J. Mol. Med. 2017, 95, 205–220. [Google Scholar] [CrossRef]
- Winkel, A.; Jaimes, Y.; Melzer, C.; Dillschneider, P.; Hartwig, H.; Stiesch, M.; von der Ohe, J.; Strauss, S.; Vogt, P.M.; Hamm, A.; et al. Cell culture media notably influence properties of human mesenchymal stroma/stem-like cells from different tissues. Cytotherapy 2020, 22, 653–668. [Google Scholar] [CrossRef]
- Hagmann, S.; Moradi, B.; Frank, S.; Dreher, T.; Kämmerer, P.W.; Richter, W.; Gotterbarm, T. Different culture media affect growth characteristics, surface marker distribution and chondrogenic differentiation of human bone marrow-derived mesenchymal stromal cells. BMC Musculoskelet. Disord. 2013, 14, 223. [Google Scholar] [CrossRef]
- Bronzini, I.; Patruno, M.; Iacopetti, I.; Martinello, T. Influence of temperature, time and different media on mesenchymal stromal cells shipped for clinical application. Vet. J. 2012, 194, 121–123. [Google Scholar] [CrossRef]
- Viganò, M.; Perucca Orfei, C.; de Girolamo, L.; Pearson, J.R.; Ragni, E.; De Luca, P.; Colombini, A. Housekeeping Gene Stability in Human Mesenchymal Stem and Tendon Cells Exposed to Tenogenic Factors. Tissue Eng. Part C Methods 2018, 24, 360–367. [Google Scholar] [CrossRef]
- Curtis, K.M.; Gomez, L.A.; Rios, C.; Garbayo, E.; Raval, A.P.; Perez-Pinzon, M.A.; Schiller, P.C. EF1alpha and RPL13a represent normalization genes suitable for RT-qPCR analysis of bone marrow derived mesenchymal stem cells. BMC Mol. Biol. 2010, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- Ragni, E.; Viganò, M.; Rebulla, P.; Giordano, R.; Lazzari, L. What is beyond a qRT-PCR study on mesenchymal stem cell differentiation properties: How to choose the most reliable housekeeping genes. J. Cell. Mol. Med. 2013, 17, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Bok, E.; Kim, S.; Thakur, G.; Choe, Y.; Oh, S.; Hwang, S.; Ock, S.; Rho, G.; Lee, S.; Lee, W.; et al. Insensitive Effects of Inflammatory Cytokines on the Reference Genes of Synovial Fluid Resident-Mesenchymal Stem Cells Derived from Rheumatoid Arthritis Patients. Int. J. Mol. Sci. 2023, 24, 15159. [Google Scholar] [CrossRef] [PubMed]
- Pham, P.V.; Vu, N.B. Off-the-shelf mesenchymal stem cells from human umbilical cord tissue can significantly improve symptoms in COVID-19 patients: An analysis of evidential relations. World J. Stem Cells 2020, 12, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Goetz, L.H.; Schork, N.J. Personalized medicine: Motivation, challenges, and progress. Fertil. Steril. 2018, 109, 952–963. [Google Scholar] [CrossRef] [PubMed]
- Ragni, E.; De Luca, P.; Perucca Orfei, C.; Colombini, A.; Viganò, M.; Lugano, G.; Bollati, V.; de Girolamo, L. Insights into Inflammatory Priming of Adipose-Derived Mesenchymal Stem Cells: Validation of Extracellular Vesicles-Embedded miRNA Reference Genes as A Crucial Step for Donor Selection. Cells 2019, 8, 369. [Google Scholar] [CrossRef]
- Wang, J.; Chang, C.; Yang, X.; Zhou, F.; Liu, J.; Feng, Z.; Hu, W. Leukemia inhibitory factor, a double-edged sword with therapeutic implications in human diseases. Mol. Ther. 2023, 31, 331–343. [Google Scholar] [CrossRef]
- Santos, G.C.; Silva, D.N.; Fortuna, V.; Silveira, B.M.; Orge, I.D.; de Santana, T.A.; Sampaio, G.L.; Paredes, B.D.; Santos, R.R.D.; Soares, M.B.P. Leukemia Inhibitory Factor (LIF) Overexpression Increases the Angiogenic Potential of Bone Marrow Mesenchymal Stem/Stromal Cells. Front. Cell Dev. Biol. 2020, 8, 778. [Google Scholar] [CrossRef]
- Jiang, Y.W.; Xiao, Q.; Hu, Z.; Pu, B.; Shu, J.; Yang, Q.; Lao, H.; Hao, J. Tissue levels of leukemia inhibitory factor vary by osteoarthritis grade. Orthopedics 2014, 37, e460–e464. [Google Scholar] [CrossRef]
- Lotz, M.; Moats, T.; Villiger, P.M. Leukemia inhibitory factor is expressed in cartilage and synovium and can contribute to the pathogenesis of arthritis. J. Clin. Investig. 1992, 90, 888–896. [Google Scholar] [CrossRef]
- Zeng, Z.; Lan, T.; Wei, Y.; Wei, X. CCL5/CCR5 axis in human diseases and related treatments. Genes Dis. 2022, 9, 12–27. [Google Scholar] [CrossRef]
- Zhong, W.; Tong, Y.; Li, Y.; Yuan, J.; Hu, S.; Hu, T.; Song, G. Mesenchymal stem cells in inflammatory microenvironment potently promote metastatic growth of cholangiocarcinoma via activating Akt/NF-κB signaling by paracrine CCL5. Oncotarget 2017, 8, 73693–73704. [Google Scholar] [CrossRef]
- Haraden, C.A.; Huebner, J.L.; Hsueh, M.; Li, Y.; Kraus, V.B. 3 Synovial fluid biomarkers associated with osteoarthritis severity reflect macrophage and neutrophil related inflammation. Arthritis Res. Ther. 2019, 21, 146. [Google Scholar] [CrossRef]
- Molnar, V.; Matišić, V.; Kodvanj, I.; Bjelica, R.; Jeleč, Z.; Hudetz, D.; Rod, E.; Čukelj, F.; Vrdoljak, T.; Vidović, D.; et al. Cytokines and Chemokines Involved in Osteoarthritis Pathogenesis. Int. J. Mol. Sci. 2021, 22, 9208. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Wiegertjes, R.; van de Loo, F.A.J.; Blaney Davidson, E.N. A roadmap to target interleukin-6 in osteoarthritis. Rheumatology 2020, 59, 2681–2694. [Google Scholar] [CrossRef]
- Eisenberg, E.; Levanon, E.Y. Human housekeeping genes are compact. Trends Genet. 2003, 19, 362–365. [Google Scholar] [CrossRef]
- Fink, T.; Lund, P.; Pilgaard, L.; Rasmussen, J.G.; Duroux, M.; Zachar, V. Instability of standard PCR reference genes in adipose-derived stem cells during propagation, differentiation and hypoxic exposure. BMC Mol. Biol. 2008, 9, 98. [Google Scholar] [CrossRef]
- Nazet, U.; Schröder, A.; Grässel, S.; Muschter, D.; Proff, P.; Kirschneck, C. Housekeeping gene validation for RT-qPCR studies on synovial fibroblasts derived from healthy and osteoarthritic patients with focus on mechanical loading. PLoS ONE 2019, 14, e0225790. [Google Scholar] [CrossRef]
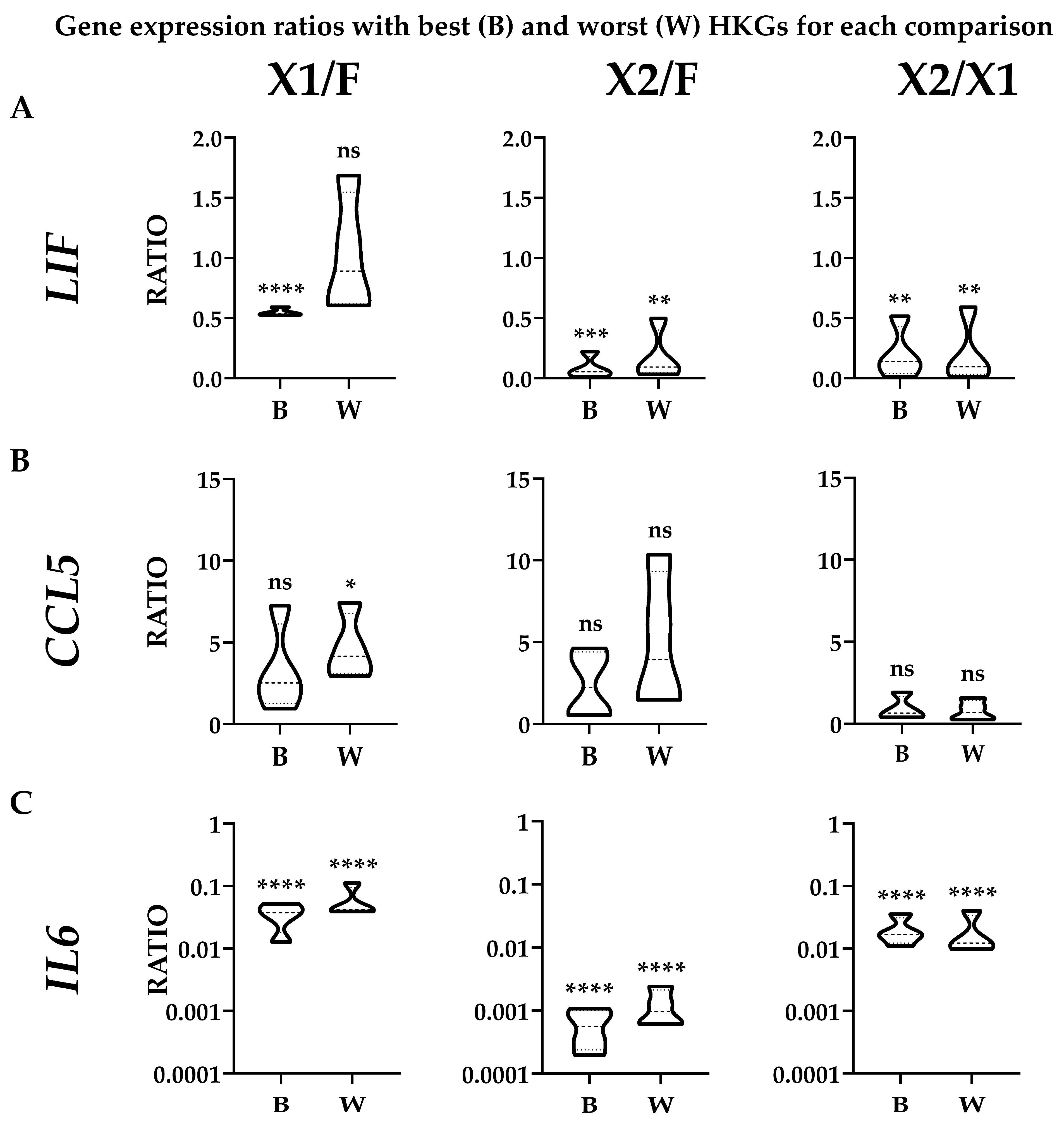

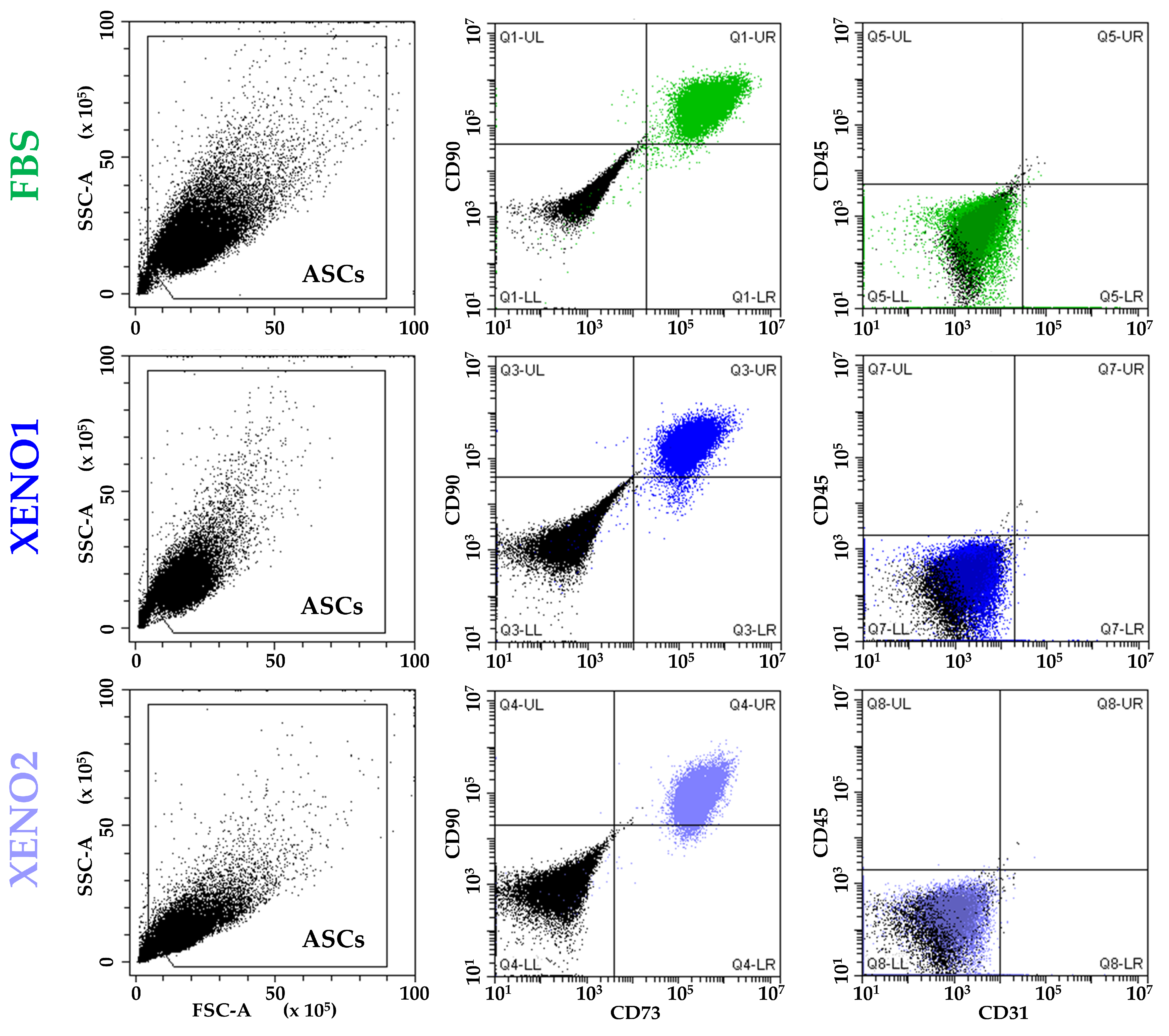{kind=link}
{kind=link}
{kind=link}
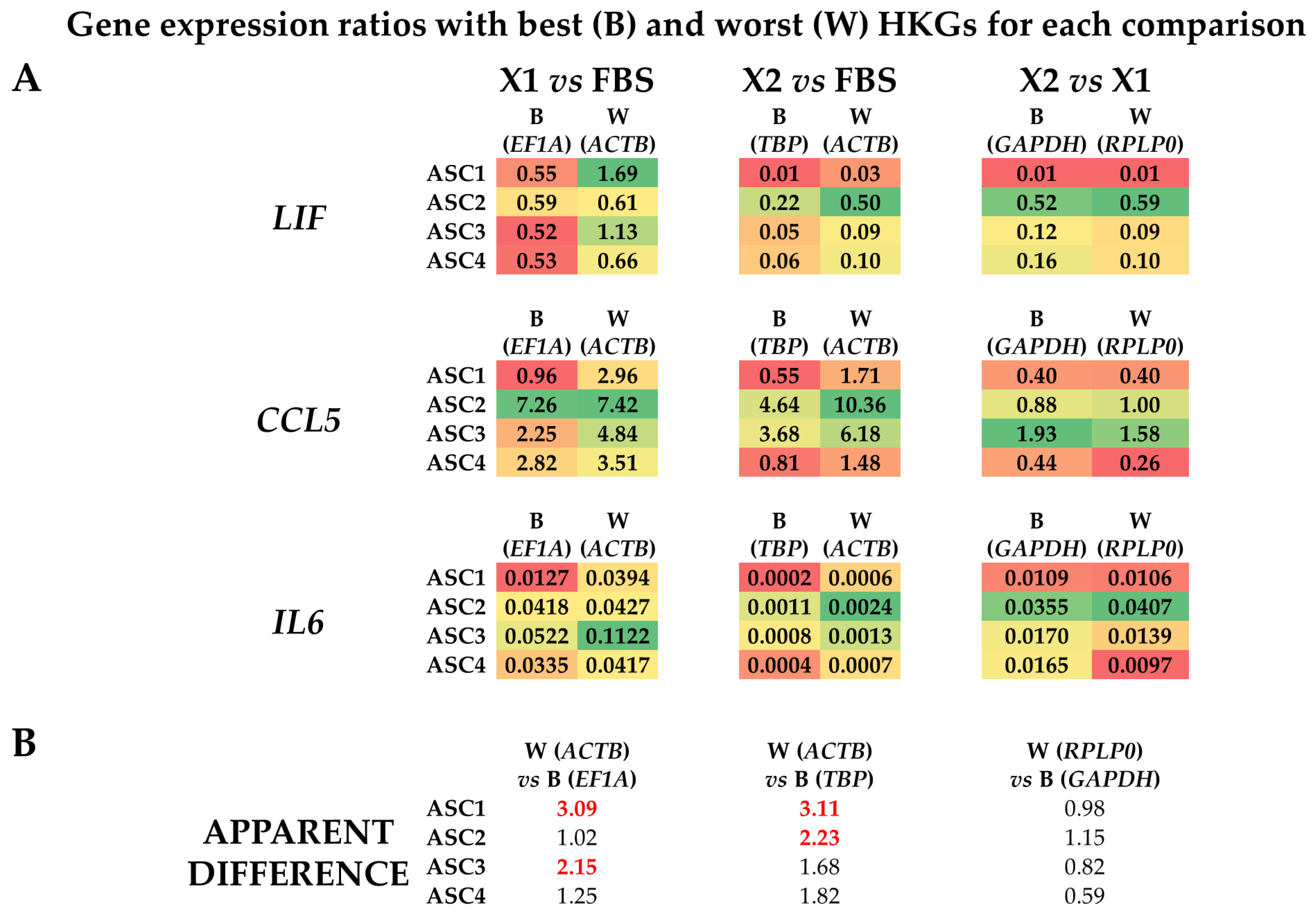{kind=link}
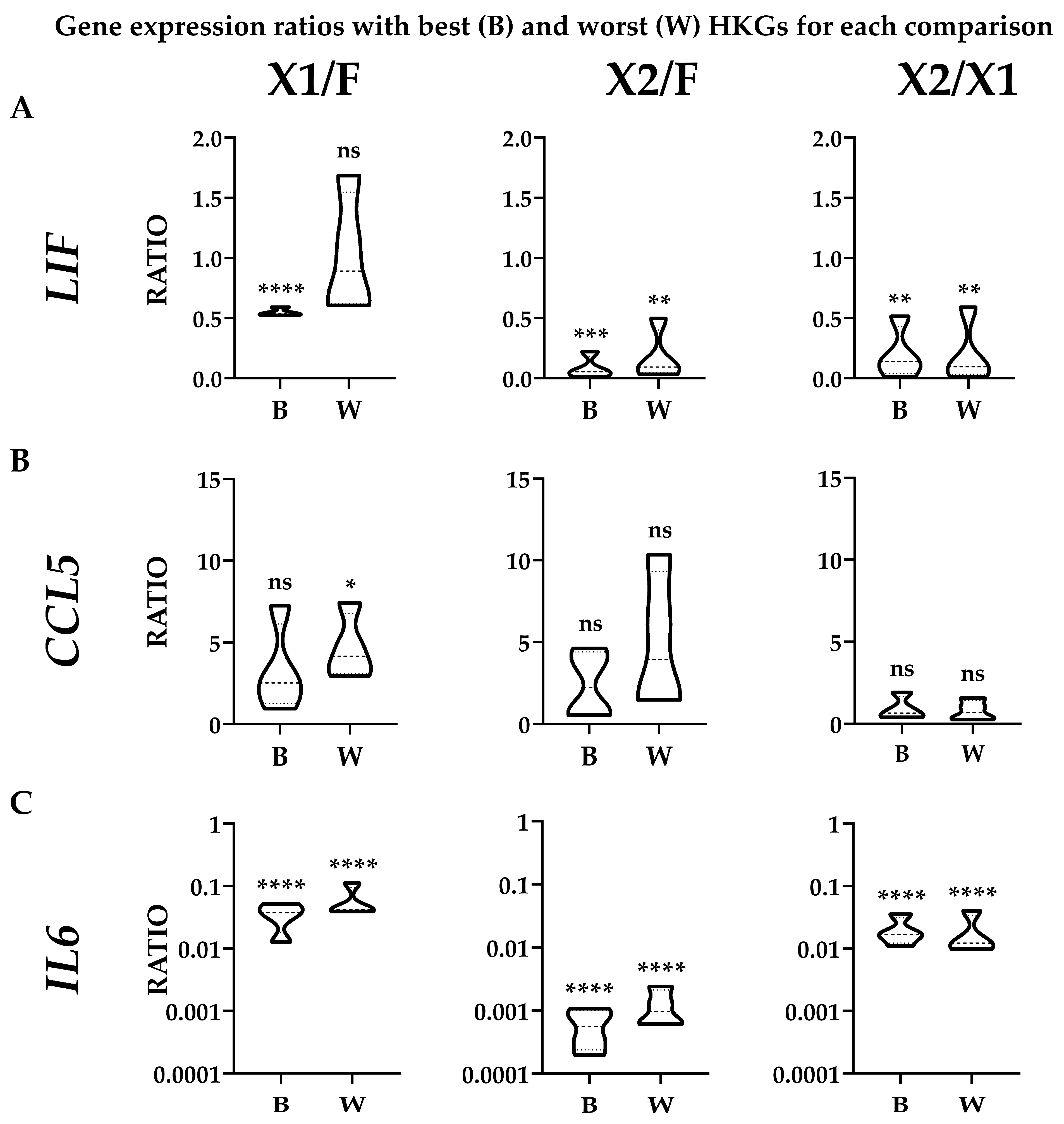{kind=link}
| ACTB | EF1A | GAPDH | RPLP0 | TBP | |
|---|---|---|---|---|---|
| ALL | 12.72 ± 0.18 | 10.19 ± 0.17 | 11.42 ± 0.13 | 11.07 ± 0.17 | 20.51 ± 0.12 |
| FBS | 12.28 ± 0.39 | 10.42 ± 0.27 | 11.59 ± 0.34 | 11.27 ± 0.29 | 20.85 ± 0.18 |
| XENO1 | 12.80 ± 0.16 | 10.18 ± 0.29 | 11.35 ± 0.10 | 11.08 ± 0.30 | 20.28 ± 0.17 |
| XENO2 | 13.09 ± 0.15 | 9.98 ± 0.27 | 11.34 ± 0.16 | 10.85 ± 0.24 | 20.39 ± 0.13 |
| Cond | Geomean | Delta CT | BestKeeper | NormFinder | Genorm | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| FBS | EF1A | 1.19 | EF1A | 0.33 | TBP | 0.30 | EF1A | 0.08 | EF1A|RPLP0 | 0.17 |
| TBP | 1.86 | TBP | 0.40 | EF1A | 0.43 | TBP | 0.20 | |||
| RPLP0 | 2.28 | RPLP0 | 0.41 | RPLP0 | 0.45 | RPLP0 | 0.28 | TBP | 0.23 | |
| ACTB | 4.23 | ACTB | 0.48 | GAPDH | 0.62 | ACTB | 0.36 | ACTB | 0.32 | |
| GAPDH | 4.73 | GAPDH | 0.66 | ACTB | 0.67 | GAPDH | 0.62 | GAPDH | 0.46 | |
| X1 | GAPDH | 1.00 | GAPDH | 0.43 | GAPDH | 0.17 | GAPDH | 0.13 | TBP|GAPDH | 0.26 |
| TBP | 1.86 | TBP | 0.46 | ACTB | 0.24 | TBP | 0.23 | |||
| ACTB | 2.71 | ACTB | 0.55 | TBP | 0.28 | ACTB | 0.38 | ACTB | 0.34 | |
| EF1A | 4.00 | EF1A | 0.60 | EF1A | 0.47 | EF1A | 0.48 | EF1A | 0.43 | |
| RPLP0 | 5.00 | RPLP0 | 0.75 | RPLP0 | 0.50 | RPLP0 | 0.68 | RPLP0 | 0.56 | |
| X2 | GAPDH | 1.32 | GAPDH | 0.44 | TBP | 0.22 | GAPDH | 0.14 | RPLP0|GAPDH | 0.29 |
| TBP | 2.00 | TBP | 0.55 | ACTB | 0.24 | TBP | 0.36 | |||
| ACTB | 2.71 | ACTB | 0.57 | GAPDH | 0.30 | ACTB | 0.43 | ACTB | 0.43 | |
| RPLP0 | 2.83 | RPLP0 | 0.59 | RPLP0 | 0.41 | RPLP0 | 0.48 | TBP | 0.49 | |
| EF1A | 5.00 | EF1A | 0.67 | EF1A | 0.46 | EF1A | 0.58 | EF1A | 0.56 | |
| X1/X2 | GAPDH | 1.00 | GAPDH | 0.43 | GAPDH | 0.23 | GAPDH | 0.18 | TBP|GAPDH | 0.36 |
| TBP | 1.68 | TBP | 0.49 | TBP | 0.25 | TBP | 0.28 | |||
| ACTB | 3.00 | ACTB | 0.56 | ACTB | 0.30 | ACTB | 0.42 | ACTB | 0.39 | |
| EF1A | 4.23 | EF1A | 0.61 | RPLP0 | 0.46 | EF1A | 0.51 | EF1A | 0.48 | |
| RPLP0 | 4.73 | RPLP0 | 0.65 | EF1A | 0.47 | RPLP0 | 0.56 | RPLP0 | 0.55 | |
| FBS/X1 | EF1A | 1.32 | EF1A | 0.51 | TBP | 0.37 | EF1A | 0.29 | EF1A|TBP | 0.33 |
| TBP | 1.41 | TBP | 0.53 | GAPDH | 0.39 | TBP | 0.33 | |||
| GAPDH | 2.71 | GAPDH | 0.56 | EF1A | 0.43 | GAPDH | 0.38 | GAPDH | 0.43 | |
| RPLP0 | 4.00 | RPLP0 | 0.61 | RPLP0 | 0.49 | RPLP0 | 0.47 | RPLP0 | 0.52 | |
| ACTB | 5.00 | ACTB | 0.67 | ACTB | 0.55 | ACTB | 0.56 | ACTB | 0.58 | |
| FBS/X2 | TBP | 1.00 | TBP | 0.56 | TBP | 0.32 | TBP | 0.30 | EF1A|TBP | 0.38 |
| EF1A | 2.00 | EF1A | 0.58 | EF1A | 0.40 | GAPDH | 0.37 | |||
| GAPDH | 3.13 | RPLP0 | 0.59 | GAPDH | 0.46 | RPLP0 | 0.37 | RPLP0 | 0.47 | |
| RPLP0 | 3.22 | GAPDH | 0.59 | RPLP0 | 0.48 | EF1A | 0.38 | GAPDH | 0.50 | |
| ACTB | 5.00 | ACTB | 0.82 | ACTB | 0.62 | ACTB | 0.74 | ACTB | 0.63 | |
| ALL | TBP | 1.41 | GAPDH | 0.54 | TBP | 0.34 | GAPDH | 0.28 | EF1A|TBP | 0.39 |
| GAPDH | 1.57 | TBP | 0.54 | GAPDH | 0.36 | TBP | 0.31 | |||
| EF1A | 2.28 | EF1A | 0.57 | EF1A | 0.43 | EF1A | 0.39 | GAPDH | 0.45 | |
| RPLP0 | 4.00 | RPLP0 | 0.63 | RPLP0 | 0.48 | RPLP0 | 0.46 | RPLP0 | 0.52 | |
| ACTB | 5.00 | ACTB | 0.73 | ACTB | 0.50 | ACTB | 0.63 | ACTB | 0.60 | |
| Cond | Geomean | Delta CT | BestKeeper | NormFinder | Genorm | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| ASC1 | TBP | 1.73 | TBP | 0.47 | RPLP0 | 0.20 | TBP | 0.13 | EF1A|GAPDH | 0.14 |
| RPLP0 | 2.00 | RPLP0 | 0.52 | ACTB | 0.35 | RPLP0 | 0.13 | |||
| EF1A | 2.45 | EF1A | 0.56 | TBP | 0.37 | EF1A | 0.44 | TBP | 0.30 | |
| GAPDH | 2.99 | GAPDH | 0.59 | EF1A | 0.59 | GAPDH | 0.50 | RPLP0 | 0.38 | |
| ACTB | 3.98 | ACTB | 1.01 | GAPDH | 0.65 | ACTB | 1.00 | ACTB | 0.63 | |
| ASC2 | GAPDH | 1.73 | GAPDH | 0.32 | TBP | 0.05 | GAPDH | 0.11 | RPLP0|TBP | 0.13 |
| TBP | 2.00 | EF1A | 0.33 | RPLP0 | 0.09 | EF1A | 0.15 | |||
| RPLP0 | 2.06 | RPLP0 | 0.38 | GAPDH | 0.24 | RPLP0 | 0.30 | GAPDH | 0.29 | |
| EF1A | 2.83 | TBP | 0.40 | EF1A | 0.29 | TBP | 0.33 | EF1A | 0.31 | |
| ACTB | 5.00 | ACTB | 0.50 | ACTB | 0.42 | ACTB | 0.46 | ACTB | 0.39 | |
| ASC3 | TBP | 1.19 | GAPDH | 0.35 | TBP | 0.07 | TBP | 0.04 | TBP|GAPDH | 0.08 |
| GAPDH | 1.41 | TBP | 0.37 | GAPDH | 0.13 | GAPDH | 0.04 | |||
| EF1A | 3.00 | EF1A | 0.38 | EF1A | 0.19 | EF1A | 0.08 | EF1A | 0.15 | |
| RPLP0 | 4.00 | RPLP0 | 0.59 | RPLP0 | 0.21 | RPLP0 | 0.55 | RPLP0 | 0.27 | |
| ACTB | 5.00 | ACTB | 0.86 | ACTB | 0.70 | ACTB | 0.85 | ACTB | 0.51 | |
| ASC4 | ACTB | 1.32 | ACTB | 0.32 | GAPDH | 0.17 | ACTB | 0.07 | ACTB|GAPDH | 0.15 |
| GAPDH | 1.73 | EF1A | 0.34 | RPLP0 | 0.21 | EF1A | 0.16 | |||
| EF1A | 2.63 | GAPDH | 0.39 | ACTB | 0.24 | GAPDH | 0.30 | EF1A | 0.29 | |
| RPLP0 | 3.36 | RPLP0 | 0.45 | EF1A | 0.44 | RPLP0 | 0.38 | RPLP0 | 0.35 | |
| TBP | 5.00 | TBP | 0.46 | TBP | 0.57 | TBP | 0.42 | TBP | 0.39 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ragni, E.; Piccolo, S.; De Luca, P.; Taiana, M.; Grieco, G.; de Girolamo, L. Housekeeping Gene Stability in Adipose Mesenchymal Stromal Cells Cultivated in Serum/Xeno-Free Media for Osteoarthritis. Cells 2024, 13, 167. https://doi.org/10.3390/cells13020167
Ragni E, Piccolo S, De Luca P, Taiana M, Grieco G, de Girolamo L. Housekeeping Gene Stability in Adipose Mesenchymal Stromal Cells Cultivated in Serum/Xeno-Free Media for Osteoarthritis. Cells. 2024; 13(2):167. https://doi.org/10.3390/cells13020167
Chicago/Turabian StyleRagni, Enrico, Simona Piccolo, Paola De Luca, Michela Taiana, Giulio Grieco, and Laura de Girolamo. 2024. "Housekeeping Gene Stability in Adipose Mesenchymal Stromal Cells Cultivated in Serum/Xeno-Free Media for Osteoarthritis" Cells 13, no. 2: 167. https://doi.org/10.3390/cells13020167
APA StyleRagni, E., Piccolo, S., De Luca, P., Taiana, M., Grieco, G., & de Girolamo, L. (2024). Housekeeping Gene Stability in Adipose Mesenchymal Stromal Cells Cultivated in Serum/Xeno-Free Media for Osteoarthritis. Cells, 13(2), 167. https://doi.org/10.3390/cells13020167