
Heat Shock Protein SSA1 Enriched in Hypoxic Secretome of Candida albicans Exerts an Immunomodulatory Effect via Regulating Macrophage Function


Abstract
1. Introduction
2. Materials and Methods
2.1. C. albicans Strain and Cell Culture
2.2. Preparation of Normoxic Secretome (NS) and Hypoxic Secretome (HS) from C. albicans
2.3. Mice and Ethical Statement
2.4. Isolation of Mouse Spleen Cells
2.5. Preparation of Mouse Bone Marrow-Derived Macrophages (BMMs)
2.6. Differential Proteomic Analysis of C. albicans NS and HS
2.7. Preparation of Recombinant Ssa1
2.8. Cytokine Assay
2.9. Effect of Ssa1 on Mouse Macrophages Viability
2.10. Differential Proteomic Analysis of Ssa1-Treated and Untreated BMMs
2.11. Gene Expression Analysis of Ssa1-Treated BMMs
2.12. Construction of Ssa1-Deficient Mutant of C. albicans (Ssa1−)
2.13. Infection of Ssa1-Preteated BMMs with C. albicans
2.14. Statistical Analysis
3. Results
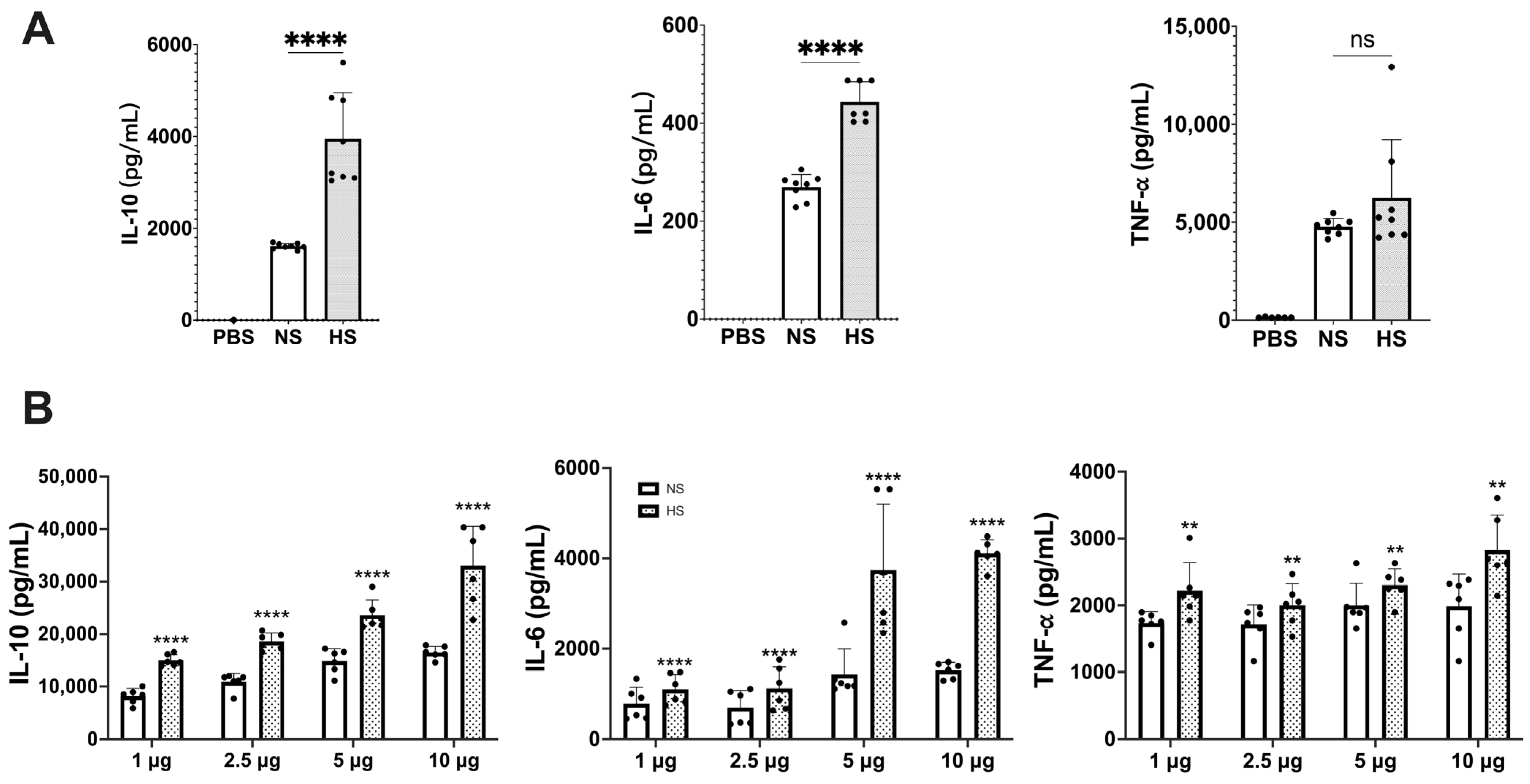
3.1. Cytokine Production in Mouse Macrophages and Spleen Cells Induced by C. albicans NS and HS
3.2. Differential Proteomic Analysis between NS and HS of C. albicans
3.3. Effect of Ssa1 on Cytokine Response and Viability of RAW 264.7 Cells
3.4. Cytokine Production and Protein Expression in Ssa1-Treated BMMs
3.5. Macrophage Polarization in Ssa1-Treated BMMs
3.6. Ssa1 Inhibits the Uptake of C. albicans by BMMs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans Pathogenicity Mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef]
- Bertolini, M.; Ranjan, A.; Thompson, A.; Diaz, P.I.; Sobue, T.; Maas, K.; Dongari-Bagtzoglou, A. Candida albicans Induces Mucosal Bacterial Dysbiosis That Promotes Invasive Infection. PLoS Pathog. 2019, 15, e1007717. [Google Scholar] [CrossRef]
- Qin, Y.; Zhang, L.; Xu, Z.; Zhang, J.; Jiang, Y.-Y.; Cao, Y.; Yan, T. Innate Immune Cell Response upon Candida albicans Infection. Virulence 2016, 7, 512–526. [Google Scholar] [CrossRef]
- Zhao, S.; Shang, A.; Guo, M.; Shen, L.; Han, Y.; Huang, X. The Advances in the Regulation of Immune Microenvironment by Candida albicans and Macrophage Cross-Talk. Front. Microbiol. 2022, 13, 1029966. [Google Scholar] [CrossRef]
- Mori, K.; Naganuma, M.; Mizuno, S.; Suzuki, H.; Kitazume, M.T.; Shimamura, K.; Chiba, S.; Sugita, A.; Matsuoka, K.; Hisamatsu, T.; et al. β-(1,3)-Glucan Derived from Candida albicans Induces Inflammatory Cytokines from Macrophages and Lamina Propria Mononuclear Cells Derived from Patients with Crohn’s Disease. Intest. Res. 2018, 16, 384–392. [Google Scholar] [CrossRef]
- Austermeier, S.; Kasper, L.; Westman, J.; Gresnigt, M.S. I Want to Break Free—Macrophage Strategies to Recognize and Kill Candida albicans, and Fungal Counter-Strategies to Escape. Curr. Opin. Microbiol. 2020, 58, 15–23. [Google Scholar] [CrossRef]
- Zheng, X.-F.; Hong, Y.-X.; Feng, G.-J.; Zhang, G.-F.; Rogers, H.; Lewis, M.A.O.; Williams, D.W.; Xia, Z.-F.; Song, B.; Wei, X.-Q. Lipopolysaccharide-Induced M2 to M1 Macrophage Transformation for IL-12p70 Production Is Blocked by Candida albicans Mediated up-Regulation of EBI3 Expression. PLoS ONE 2013, 8, e63967. [Google Scholar] [CrossRef]
- Kadomoto, S.; Izumi, K.; Mizokami, A. Macrophage Polarity and Disease Control. Int. J. Mol. Sci. 2021, 23, 144. [Google Scholar] [CrossRef]
- Sorgo, A.G.; Heilmann, C.J.; Brul, S.; de Koster, C.G.; Klis, F.M. Beyond the Wall: Candida albicans Secret(e)s to Survive. FEMS Microbiol. Lett. 2013, 338, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Gil-Bona, A.; Amador-García, A.; Gil, C.; Monteoliva, L. The External Face of Candida albicans: A Proteomic View of the Cell Surface and the Extracellular Environment. J. Proteom. 2018, 180, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Hruskova-Heidingsfeldova, O. Secreted Proteins of Candida albicans. Front. Biosci. 2008, 13, 7227–7242. [Google Scholar] [CrossRef]
- Klis, F.M.; Brul, S. Adaptations of the Secretome of Candida albicans in Response to Host-Related Environmental Conditions. Eukaryot. Cell 2015, 14, 1165–1172. [Google Scholar] [CrossRef]
- Sorgo, A.G.; Heilmann, C.J.; Dekker, H.L.; Brul, S.; de Koster, C.G.; Klis, F.M. Mass Spectrometric Analysis of the Secretome of Candida albicans. Yeast 2010, 27, 661–672. [Google Scholar] [CrossRef]
- Vaz, C.; Pitarch, A.; Gómez-Molero, E.; Amador-García, A.; Weig, M.; Bader, O.; Monteoliva, L.; Gil, C. Mass Spectrometry-Based Proteomic and Immunoproteomic Analyses of the Candida albicans Hyphal Secretome Reveal Diagnostic Biomarker Candidates for Invasive Candidiasis. J. Fungi 2021, 7, 501. [Google Scholar] [CrossRef]
- Haque, N.; Rahman, M.T.; Abu Kasim, N.H.; Alabsi, A.M. Hypoxic Culture Conditions as a Solution for Mesenchymal Stem Cell Based Regenerative Therapy. Sci. World J. 2013, 2013, 632972. [Google Scholar] [CrossRef]
- Zeitouni, N.E.; Chotikatum, S.; von Köckritz-Blickwede, M.; Naim, H.Y. The Impact of Hypoxia on Intestinal Epithelial Cell Functions: Consequences for Invasion by Bacterial Pathogens. Mol. Cell. Pediatr. 2016, 3, 14. [Google Scholar] [CrossRef]
- Fisher, E.M.; Khan, M.; Salisbury, R.; Kuppusamy, P. Noninvasive Monitoring of Small Intestinal Oxygen in a Rat Model of Chronic Mesenteric Ischemia. Cell Biochem. Biophys. 2013, 67, 451–459. [Google Scholar] [CrossRef]
- Kumamoto, C.A. Inflammation and Gastrointestinal Candida Colonization. Curr. Opin. Microbiol. 2011, 14, 386–391. [Google Scholar] [CrossRef]
- Suzuki, S.; Subsomwong, P.; Narita, K.; Kawai, N.; Ishiai, T.; Teng, W.; Sukchawalit, R.; Nakane, A.; Tasaka, S.; Asano, K. Differential Proteomic Analysis and Pathogenic Effects of Outer Membrane Vesicles Derived from Acinetobacter Baumannii under Normoxia and Hypoxia. PLoS ONE 2023, 18, e0283109. [Google Scholar] [CrossRef]
- Skrzypek, M.S.; Binkley, J.; Binkley, G.; Miyasato, S.R.; Simison, M.; Sherlock, G. The Candida Genome Database (CGD): Incorporation of Assembly 22, Systematic Identifiers and Visualization of High Throughput Sequencing Data. Nucleic Acids Res. 2017, 45, D592–D596. [Google Scholar] [CrossRef]
- Demichev, V.; Messner, C.B.; Vernardis, S.I.; Lilley, K.S.; Ralser, M. DIA-NN: Neural Networks and Interference Correction Enable Deep Proteome Coverage in High Throughput. Nat. Methods 2020, 17, 41–44. [Google Scholar] [CrossRef]
- Vyas, V.K.; Bushkin, G.G.; Bernstein, D.A.; Getz, M.A.; Sewastianik, M.; Barrasa, M.I.; Bartel, D.P.; Fink, G.R. New CRISPR Mutagenesis Strategies Reveal Variation in Repair Mechanisms among Fungi. mSphere 2018, 3, e00154-18. [Google Scholar] [CrossRef]
- Labun, K.; Montague, T.G.; Krause, M.; Torres Cleuren, Y.N.; Tjeldnes, H.; Valen, E. CHOPCHOP v3: Expanding the CRISPR Web Toolbox beyond Genome Editing. Nucleic Acids Res. 2019, 47, W171–W174. [Google Scholar] [CrossRef]
- Borges, T.J.; Wieten, L.; van Herwijnen, M.J.C.; Broere, F.; van der Zee, R.; Bonorino, C.; van Eden, W. The Anti-Inflammatory Mechanisms of Hsp70. Front. Immunol. 2012, 3, 95. [Google Scholar] [CrossRef]
- Pelegrin, P.; Barroso-Gutierrez, C.; Surprenant, A. P2X7 Receptor Differentially Couples to Distinct Release Pathways for IL-1beta in Mouse Macrophage. J. Immunol. 2008, 180, 7147–7157. [Google Scholar] [CrossRef]
- Wang, L.-X.; Zhang, S.-X.; Wu, H.-J.; Rong, X.-L.; Guo, J. M2b Macrophage Polarization and Its Roles in Diseases. J. Leukoc. Biol. 2019, 106, 345–358. [Google Scholar] [CrossRef]
- Hasin, N.; Cusack, S.A.; Ali, S.S.; Fitzpatrick, D.A.; Jones, G.W. Global Transcript and Phenotypic Analysis of Yeast Cells Expressing Ssa1, Ssa2, Ssa3 or Ssa4 as Sole Source of Cytosolic Hsp70-Ssa Chaperone Activity. BMC Genom. 2014, 15, 194. [Google Scholar] [CrossRef]
- Guo, M.S.; Gross, C.A. Stress-Induced Remodeling of the Bacterial Proteome. Curr. Biol. 2014, 24, R424–R434. [Google Scholar] [CrossRef]
- Lotz, S.K.; Knighton, L.E.; Nitika; Jones, G.W.; Truman, A.W. Not Quite the SSAme: Unique Roles for the Yeast Cytosolic Hsp70s. Curr. Genet. 2019, 65, 1127–1134. [Google Scholar] [CrossRef]
- Satala, D.; Karkowska-Kuleta, J.; Zelazna, A.; Rapala-Kozik, M.; Kozik, A. Moonlighting Proteins at the Candidal Cell Surface. Microorganisms 2020, 8, 1046. [Google Scholar] [CrossRef] [PubMed]
- Garfoot, A.L.; Shen, Q.; Wüthrich, M.; Klein, B.S.; Rappleye, C.A. The Eng1 β-Glucanase Enhances Histoplasma Virulence by Reducing β-Glucan Exposure. mBio 2016, 7, e01388-15. [Google Scholar] [CrossRef]
- Granger, B.L.; Flenniken, M.L.; Davis, D.A.; Mitchell, A.P.; Cutler, J.E. Yeast Wall Protein 1 of Candida albicans. Microbiology 2005, 151, 1631–1644. [Google Scholar] [CrossRef]
- Esteban, P.F.; Ríos, I.; García, R.; Dueñas, E.; Plá, J.; Sánchez, M.; de Aldana, C.R.V.; Del Rey, F. Characterization of the CaENG1 Gene Encoding an Endo-1,3-Beta-Glucanase Involved in Cell Separation in Candida albicans. Curr. Microbiol. 2005, 51, 385–392. [Google Scholar] [CrossRef]
- Yang, M.; Solis, N.V.; Marshall, M.; Garleb, R.; Zhou, T.; Wang, D.; Swidergall, M.; Pearlman, E.; Filler, S.G.; Liu, H. Control of β-Glucan Exposure by the Endo-1,3-Glucanase Eng1 in Candida albicans Modulates Virulence. PLoS Pathog. 2022, 18, e1010192. [Google Scholar] [CrossRef]
- Sem, X.; Le, G.T.T.; Tan, A.S.M.; Tso, G.; Yurieva, M.; Liao, W.W.P.; Lum, J.; Srinivasan, K.G.; Poidinger, M.; Zolezzi, F.; et al. β-Glucan Exposure on the Fungal Cell Wall Tightly Correlates with Competitive Fitness of Candida Species in the Mouse Gastrointestinal Tract. Front. Cell Infect. Microbiol. 2016, 6, 186. [Google Scholar] [CrossRef]
- Chaffin, W.L. Candida albicans Cell Wall Proteins. Microbiol. Mol. Biol. Rev. 2008, 72, 495–544. [Google Scholar] [CrossRef]
- Gozalbo, D.; Gil-Navarro, I.; Azorín, I.; Renau-Piqueras, J.; Martínez, J.P.; Gil, M.L. The Cell Wall-Associated Glyceraldehyde-3-Phosphate Dehydrogenase of Candida albicans Is Also a Fibronectin and Laminin Binding Protein. Infect. Immun. 1998, 66, 2052–2059. [Google Scholar] [CrossRef]
- Sun, J.N.; Solis, N.V.; Phan, Q.T.; Bajwa, J.S.; Kashleva, H.; Thompson, A.; Liu, Y.; Dongari-Bagtzoglou, A.; Edgerton, M.; Filler, S.G. Host Cell Invasion and Virulence Mediated by Candida albicans Ssa1. PLoS Pathog. 2010, 6, e1001181. [Google Scholar] [CrossRef]
- Zininga, T.; Ramatsui, L.; Shonhai, A. Heat Shock Proteins as Immunomodulants. Molecules 2018, 23, 2846. [Google Scholar] [CrossRef]
- De Maio, A. Extracellular Hsp70: Export and Function. Curr. Protein Pept. Sci. 2014, 15, 225–231. [Google Scholar] [CrossRef]
- Liu, C.; Chu, D.; Kalantar-Zadeh, K.; George, J.; Young, H.A.; Liu, G. Cytokines: From Clinical Significance to Quantification. Adv. Sci. 2021, 8, e2004433. [Google Scholar] [CrossRef]
- Opal, S.M.; DePalo, V.A. Anti-Inflammatory Cytokines. Chest 2000, 117, 1162–1172. [Google Scholar] [CrossRef]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The Pro- and Anti-Inflammatory Properties of the Cytokine Interleukin-6. Biochim. Biophys. Acta 2011, 1813, 878–888. [Google Scholar] [CrossRef]
- Barton, B.E.; Shortall, J.; Jackson, J.V. Interleukins 6 and 11 Protect Mice from Mortality in a Staphylococcal Enterotoxin-Induced Toxic Shock Model. Infect. Immun. 1996, 64, 714–718. [Google Scholar] [CrossRef]
- Tilg, H.; Trehu, E.; Atkins, M.B.; Dinarello, C.A.; Mier, J.W. Interleukin-6 (IL-6) as an Anti-Inflammatory Cytokine: Induction of Circulating IL-1 Receptor Antagonist and Soluble Tumor Necrosis Factor Receptor P55. Blood 1994, 83, 113–118. [Google Scholar] [CrossRef]
- Dubey, A.; Izakelian, L.; Ayaub, E.A.; Ho, L.; Stephenson, K.; Wong, S.; Kwofie, K.; Austin, R.C.; Botelho, F.; Ask, K.; et al. Separate Roles of IL-6 and Oncostatin M in Mouse Macrophage Polarization in Vitro and in Vivo. Immunol. Cell Biol. 2018, 96, 257–272. [Google Scholar] [CrossRef]
- Sironi, M.; Martinez, F.O.; D’Ambrosio, D.; Gattorno, M.; Polentarutti, N.; Locati, M.; Gregorio, A.; Iellem, A.; Cassatella, M.A.; Van Damme, J.; et al. Differential Regulation of Chemokine Production by Fcgamma Receptor Engagement in Human Monocytes: Association of CCL1 with a Distinct Form of M2 Monocyte Activation (M2b, Type 2). J. Leukoc. Biol. 2006, 80, 342–349. [Google Scholar] [CrossRef]
- Viola, A.; Munari, F.; Sánchez-Rodríguez, R.; Scolaro, T.; Castegna, A. The Metabolic Signature of Macrophage Responses. Front. Immunol. 2019, 10, 1462. [Google Scholar] [CrossRef]
- Nakanishi, K.; Yoshimoto, T.; Tsutsui, H.; Okamura, H. Interleukin-18 Is a Unique Cytokine That Stimulates Both Th1 and Th2 Responses Depending on Its Cytokine Milieu. Cytokine Growth Factor Rev. 2001, 12, 53–72. [Google Scholar] [CrossRef]
- Detry, S.; Andries, J.; Bloch, Y.; Gabay, C.; Clancy, D.M.; Savvides, S.N. Structural Basis of Human IL-18 Sequestration by the Decoy Receptor IL-18 Binding Protein in Inflammation and Tumor Immunity. J. Biol. Chem. 2022, 298, 101908. [Google Scholar] [CrossRef]
- Dinarello, C.A.; Novick, D.; Kim, S.; Kaplanski, G. Interleukin-18 and IL-18 Binding Protein. Front. Immunol. 2013, 4, 289. [Google Scholar] [CrossRef]
- Zhou, T.; Damsky, W.; Weizman, O.-E.; McGeary, M.K.; Hartmann, K.P.; Rosen, C.E.; Fischer, S.; Jackson, R.; Flavell, R.A.; Wang, J.; et al. IL-18BP Is a Secreted Immune Checkpoint and Barrier to IL-18 Immunotherapy. Nature 2020, 583, 609–614. [Google Scholar] [CrossRef]
- Clark, J.T.; Weizman, O.-E.; Aldridge, D.L.; Shallberg, L.A.; Eberhard, J.; Lanzar, Z.; Wasche, D.; Huck, J.D.; Zhou, T.; Ring, A.M.; et al. IL-18BP Mediates the Balance between Protective and Pathological Immune Responses to Toxoplasma gondii. Cell Rep. 2023, 42, 112147. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, P.; Wang, C.; Han, C.; Meng, J.; Liu, X.; Xu, S.; Li, N.; Wang, Q.; Shi, X.; et al. Immune Responsive Gene 1 (IRG1) Promotes Endotoxin Tolerance by Increasing A20 Expression in Macrophages through Reactive Oxygen Species. J. Biol. Chem. 2013, 288, 16225–16234. [Google Scholar] [CrossRef]
- Tonecka, K.; Braniewska, A.; Pilch, Z.; Sas, Z.; Skorzynski, M.; Manuali, E.; Rygiel, T.P. The CD200 Regulates Inflammation in Mice Independently of TNF-α Production. Int. J. Mol. Sci. 2021, 22, 5358. [Google Scholar] [CrossRef]
- Hoek, R.M.; Ruuls, S.R.; Murphy, C.A.; Wright, G.J.; Goddard, R.; Zurawski, S.M.; Blom, B.; Homola, M.E.; Streit, W.J.; Brown, M.H.; et al. Down-Regulation of the Macrophage Lineage through Interaction with OX2 (CD200). Science 2000, 290, 1768–1771. [Google Scholar] [CrossRef]
- Yunna, C.; Mengru, H.; Lei, W.; Weidong, C. Macrophage M1/M2 Polarization. Eur. J. Pharmacol. 2020, 877, 173090. [Google Scholar] [CrossRef]
- McNamee, E.N.; Ryan, K.M.; Kilroy, D.; Connor, T.J. Noradrenaline Induces IL-1ra and IL-1 Type II Receptor Expression in Primary Glial Cells and Protects against IL-1beta-Induced Neurotoxicity. Eur. J. Pharmacol. 2010, 626, 219–228. [Google Scholar] [CrossRef]
- Harada, H.; Wakabayashi, G.; Takayanagi, A.; Shimazu, M.; Matsumoto, K.; Obara, H.; Shimizu, N.; Kitajima, M. Transfer of the Interleukin-1 Receptor Antagonist Gene into Rat Liver Abrogates Hepatic Ischemia-Reperfusion Injury. Transplantation 2002, 74, 1434–1441. [Google Scholar] [CrossRef]
- Park, E.-M.; Cho, B.-P.; Volpe, B.T.; Cruz, M.O.; Joh, T.H.; Cho, S. Ibuprofen Protects Ischemia-Induced Neuronal Injury via up-Regulating Interleukin-1 Receptor Antagonist Expression. Neuroscience 2005, 132, 625–631. [Google Scholar] [CrossRef]
- Arend, W.P.; Malyak, M.; Guthridge, C.J.; Gabay, C. Interleukin-1 Receptor Antagonist: Role in Biology. Annu. Rev. Immunol. 1998, 16, 27–55. [Google Scholar] [CrossRef]
- Wu, Y.; Zhu, X.; Li, N.; Chen, T.; Yang, M.; Yao, M.; Liu, X.; Jin, B.; Wang, X.; Cao, X. CMRF-35-like Molecule 3 Preferentially Promotes TLR9-Triggered Proinflammatory Cytokine Production in Macrophages by Enhancing TNF Receptor-Associated Factor 6 Ubiquitination. J. Immunol. 2011, 187, 4881–4889. [Google Scholar] [CrossRef]
- Watson, C.K.; Schloesser, D.; Fundel-Clemens, K.; Lerner, C.; Gabler, S.; Baskaran, P.; Wohnhaas, C.T.; Dichtl, S.; Huber, H.J.; Ask, K.; et al. Antifibrotic Drug Nintedanib Inhibits CSF1R to Promote IL-4-Associated Tissue Repair Macrophages. Am. J. Respir. Cell Mol. Biol. 2023, 68, 366–380. [Google Scholar] [CrossRef]
- Qiu, L.-Q.; Cresswell, P.; Chin, K.-C. Viperin Is Required for Optimal Th2 Responses and T-Cell Receptor-Mediated Activation of NF-κB and AP-1. Blood 2009, 113, 3520–3529. [Google Scholar] [CrossRef]
- Zhang, W.; Zhou, Q.; Xu, W.; Cai, Y.; Yin, Z.; Gao, X.; Xiong, S. DNA-Dependent Activator of Interferon-Regulatory Factors (DAI) Promotes Lupus Nephritis by Activating the Calcium Pathway. J. Biol. Chem. 2013, 288, 13534–13550. [Google Scholar] [CrossRef]
- Porta, C.; Rimoldi, M.; Raes, G.; Brys, L.; Ghezzi, P.; Di Liberto, D.; Dieli, F.; Ghisletti, S.; Natoli, G.; De Baetselier, P.; et al. Tolerance and M2 (Alternative) Macrophage Polarization Are Related Processes Orchestrated by P50 Nuclear Factor κB. Proc. Natl. Acad. Sci. USA 2009, 106, 14978–14983. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, W.; Xiong, S. Blockade of Notch1 Signaling Alleviates Murine Lupus via Blunting Macrophage Activation and M2b Polarization. J. Immunol. 2010, 184, 6465–6478. [Google Scholar] [CrossRef]
- Cai, Y.; Zhang, W.; Xiong, S. Mannose-Binding Lectin Blunts Macrophage Polarization and Ameliorates Lupus Nephritis. PLoS ONE 2013, 8, e62465. [Google Scholar] [CrossRef]
- Lefèvre, L.; Galès, A.; Olagnier, D.; Bernad, J.; Perez, L.; Burcelin, R.; Valentin, A.; Auwerx, J.; Pipy, B.; Coste, A. PPARγ Ligands Switched High Fat Diet-Induced Macrophage M2b Polarization toward M2a Thereby Improving Intestinal Candida Elimination. PLoS ONE 2010, 5, e12828. [Google Scholar] [CrossRef]
- Kobayashi, M.; Nakamura, K.; Cornforth, M.; Suzuki, F. Role of M2b Macrophages in the Acceleration of Bacterial Translocation and Subsequent Sepsis in Mice Exposed to Whole Body [137Cs] γ-Irradiation. J. Immunol. 2012, 189, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, A.; Yang, C.-Q.; Garred, P.; Ma, Y.J. Rapid and Efficient Purification of Functional Collectin-12 and Its Opsonic Activity against Fungal Pathogens. J. Immunol. Res. 2019, 2019, 9164202. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Ohtani, K.; Fukuoh, A.; Yoshizaki, T.; Fukuda, M.; Motomura, W.; Mori, K.; Fukuzawa, J.; Kitamoto, N.; Yoshida, I.; et al. Scavenger Receptor Collectin Placenta 1 (CL-P1) Predominantly Mediates Zymosan Phagocytosis by Human Vascular Endothelial Cells. J. Biol. Chem. 2009, 284, 3956–3965. [Google Scholar] [CrossRef]
- Ohtani, K.; Suzuki, Y.; Eda, S.; Kawai, T.; Kase, T.; Keshi, H.; Sakai, Y.; Fukuoh, A.; Sakamoto, T.; Itabe, H.; et al. The Membrane-Type Collectin CL-P1 Is a Scavenger Receptor on Vascular Endothelial Cells. J. Biol. Chem. 2001, 276, 44222–44228. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Protein Name | p-Value | Fold Change (Hypoxic vs. Normoxic) | Mass (Da) |
|---|---|---|---|---|
| orf19.1672 | Coatomer subunit alpha | 0.01127 | Infinity | 104,312 |
| MLC1 | Mlc1p | 0.03923 | Infinity | 16,444 |
| RPL18 | Ribosomal 60S subunit protein L18A | 0.04962 | Infinity | 20,770 |
| MUQ1 | CTP:phosphoethanolamine cytidylyltransferase | 0.01592 | 9.134821103 | 42,013 |
| ACT1 | Actin | 0.00451 | 8.696141484 | 41,690 |
| ENG1 | Endo-1,3(4)-beta-glucanase 1 | 0.00781 | 7.471613819 | 124,051 |
| SER33 | Phosphoglycerate dehydrogenase | 0.04826 | 7.18653572 | 50,359 |
| YWP1 | Yeast-form wall Protein 1 | 0.01423 | 6.437111487 | 54,278 |
| RNR21 | Ribonucleotide-diphosphate reductase subunit | 0.02691 | 5.305513281 | 47,437 |
| HSP104 | Chaperone ATPase | 0.00311 | 5.208629718 | 99,919 |
| RPL8B | 60S ribosomal protein L8 | 0.01923 | 5.078615072 | 28,498 |
| GCA2 | Gca2p | 0.01544 | 5.017964135 | 105,678 |
| TDH3 | Glyceraldehyde-3-phosphate dehydrogenase | 0.02019 | 4.544732628 | 35,833 |
| EVP1 | Uncharacterized protein | 0.00565 | 4.418480132 | 44,929 |
| orf19.1626 | Deoxyhypusine synthase | 0.0487 | 2.815387167 | 41,634 |
| SSA1 | Heat shock protein SSA1 | 0.01616 | 2.66623664 | 70,324 |
| YPT1 | Rab family GTPase | 0.04738 | 1.705239723 | 23,019 |
| Gene | Protein | Mean (Control) | Mean (Ssa1) | q-Value * | Fold Change |
|---|---|---|---|---|---|
| Rsad2 | Radical S-adenosyl methionine domain-containing protein 2 | 0.000 | 47.557 | 0.001 | |
| Nmes1 | Normal mucosa of esophagus-specific gene 1 protein | 0.000 | 13.452 | 0.027 | |
| Mov10 | Putative helicase MOV-10 | 0.000 | 9.290 | 0.027 | |
| Herc6 | E3 ISG15--protein ligase Herc6 | 0.000 | 9.179 | 0.002 | |
| Ifi205a | Interferon-activable protein 205-A | 0.000 | 7.494 | 0.014 | |
| Il18bp | Interleukin-18-binding protein | 0.000 | 6.862 | 0.005 | |
| Cd86 | T-lymphocyte activation antigen CD86 | 0.000 | 6.152 | 0.018 | |
| Cd40 | Tumor necrosis factor receptor superfamily member 5 | 3.189 | 42.712 | 0.000 | 6.697 |
| Cmpk2 | UMP-CMP kinase 2, mitochondrial | 87.543 | 489.552 | 0.001 | 5.592 |
| Acod1 | Cis-aconitate decarboxylase | 14.565 | 91.405 | 0.027 | 5.230 |
| Oasl1 | 2′-5′-oligoadenylate synthase-like protein 1 | 13.636 | 65.679 | 0.003 | 4.014 |
| Cd200 | OX-2 membrane glycoprotein | 0.365 | 8.688 | 0.002 | 3.968 |
| Gbp4 | Guanylate-binding protein 4 | 19.279 | 74.645 | 0.001 | 3.872 |
| Gbp1 | Guanylate-binding protein 1 | 18.157 | 68.355 | 0.000 | 3.765 |
| Bst2 | Bone marrow stromal antigen 2 | 5.948 | 26.738 | 0.031 | 3.746 |
| Ifit1 | Interferon-induced protein with tetratricopeptide repeats 1 | 18.342 | 68.266 | 0.001 | 3.722 |
| Isg15 | Ubiquitin-like protein ISG15 | 112.471 | 411.178 | 0.039 | 3.656 |
| Phf11 | PHD finger protein 11 | 1.918 | 13.647 | 0.030 | 3.557 |
| Gbp5 | Guanylate-binding protein 5 | 9.073 | 47.393 | 0.014 | 3.482 |
| Ifit2 | Interferon-induced protein with tetratricopeptide repeats 2 | 25.488 | 83.357 | 0.007 | 3.270 |
| Ifit3 | Interferon-induced protein with tetratricopeptide repeats 3 | 22.823 | 88.089 | 0.004 | 3.216 |
| Ptgs2 | Prostaglandin G/H synthase 2 | 1.107 | 20.992 | 0.038 | 3.161 |
| Iigp1 | Interferon-inducible GTPase 1 | 8.602 | 31.608 | 0.002 | 3.062 |
| Cd274 | Programmed cell death 1 ligand 1 | 3.463 | 19.962 | 0.024 | 2.883 |
| Gbp2 | Guanylate-binding protein 2 | 11.134 | 36.508 | 0.007 | 2.733 |
| H2-T23 | H-2 class I histocompatibility antigen, D-37 alpha chain | 7.209 | 29.348 | 0.006 | 2.714 |
| Nt5c3a | Cytosolic 5’-nucleotidase 3A | 9.823 | 26.521 | 0.002 | 2.700 |
| Slc7a2 | Cationic amino acid transporter 2 | 0.832 | 12.178 | 0.004 | 2.439 |
| Sp110 | Sp110 nuclear body protein | 11.318 | 27.038 | 0.005 | 2.389 |
| Tapbp | Tapasin | 165.935 | 380.833 | 0.006 | 2.295 |
| H2-Q8 | H-2 class I histocompatibility antigen, Q8 alpha chain | 0.888 | 12.071 | 0.002 | 2.265 |
| Slc15a3 | Solute carrier family 15 member 3 | 4.673 | 20.979 | 0.014 | 2.245 |
| Icam1 | Intercellular adhesion molecule 1 | 31.871 | 70.840 | 0.007 | 2.223 |
| Isg20 | Interferon-stimulated gene 20 kDa protein | 1.486 | 18.469 | 0.029 | 2.071 |
| Trex1 | Three-prime repair exonuclease 1 | 22.766 | 45.811 | 0.038 | 2.012 |
| Daxx | Death domain-associated protein 6 | 3.396 | 19.414 | 0.020 | 1.906 |
| Rnaset2a; Rnaset2b | Ribonuclease T2-A | 2.402 | 9.037 | 0.038 | 1.881 |
| Sdc1 | Syndecan-1 | 2.376 | 12.898 | 0.025 | 1.810 |
| H2-L | H-2 class I histocompatibility antigen, L-D alpha chain | 217.562 | 379.962 | 0.002 | 1.746 |
| Tap1 | Antigen peptide transporter 1 | 20.946 | 36.059 | 0.013 | 1.721 |
| Serpina3k | Serine protease inhibitor A3K | 0.414 | 4.245 | 0.010 | 1.708 |
| Clic4 | Chloride intracellular channel protein 4 | 59.976 | 101.831 | 0.006 | 1.698 |
| Clec4e | C-type lectin domain family 4 member E | 0.873 | 8.762 | 0.007 | 1.672 |
| Il1rn | Interleukin-1 receptor antagonist protein | 22.857 | 37.083 | 0.018 | 1.622 |
| Tap2 | Antigen peptide transporter 2 | 37.013 | 55.981 | 0.026 | 1.513 |
| Nfkb2 | Nuclear factor NF-kappa-B p100 subunit | 26.719 | 39.959 | 0.039 | 1.496 |
| Chmp4b | Charged multivesicular body protein 4b | 88.550 | 131.179 | 0.046 | 1.481 |
| Sema4d | Semaphorin-4D | 0.549 | 4.822 | 0.019 | 1.463 |
| Plxna1 | Plexin-A1 | 23.996 | 34.045 | 0.026 | 1.419 |
| Rnf114 | E3 ubiquitin-protein ligase RNF114 | 13.082 | 18.431 | 0.031 | 1.409 |
| Irgm1 | Immunity-related GTPase family M protein 1 | 62.808 | 87.582 | 0.034 | 1.394 |
| Mthfd2 | Bifunctional methylenetetrahydrofolate dehydrogenase/cyclohydrolase, mitochondrial | 1.540 | 11.974 | 0.027 | 1.296 |
| Hspa5 | Endoplasmic reticulum chaperone BiP | 402.346 | 512.956 | 0.030 | 1.275 |
| Src | Proto-oncogene tyrosine-protein kinase Src | 35.885 | 45.608 | 0.039 | 1.271 |
| Aldh1b1 | Aldehyde dehydrogenase X, mitochondrial | 20.697 | 26.002 | 0.022 | 1.256 |
| Gyg1 | Glycogenin-1 | 43.732 | 54.653 | 0.013 | 1.250 |
| Oat | Ornithine aminotransferase, mitochondrial | 45.390 | 56.139 | 0.039 | 1.237 |
| Tlr2 | Toll-like receptor 2 | 20.473 | 25.201 | 0.040 | 1.231 |
| Nampt | Nicotinamide phosphoribosyltransferase | 102.911 | 125.617 | 0.027 | 1.221 |
| Tgm2 | Protein-glutamine gamma-glutamyltransferase 2 | 59.923 | 71.772 | 0.020 | 1.198 |
| Pdia3 | Protein disulfide-isomerase A3 | 427.181 | 508.180 | 0.013 | 1.190 |
| Dlat | Dihydrolipoyllysine-residue acetyltransferase component of pyruvate dehydrogenase complex, mitochondrial | 82.697 | 98.320 | 0.038 | 1.189 |
| Hnrnpm | Heterogeneous nuclear ribonucleoprotein M | 150.866 | 174.278 | 0.033 | 1.155 |
| Sec11a | Signal peptidase complex catalytic subunit SEC11A | 23.882 | 27.462 | 0.038 | 1.150 |
| Hsp90ab1 | Heat shock protein HSP 90-beta | 811.190 | 928.346 | 0.024 | 1.144 |
| Hsp90b1 | Endoplasmin | 904.794 | 1006.715 | 0.024 | 1.113 |
| Capn2 | Calpain-2 catalytic subunit | 107.346 | 90.642 | 0.010 | 0.844 |
| Gpaa1 | Glycosylphosphatidylinositol anchor attachment 1 protein | 22.174 | 17.066 | 0.042 | 0.770 |
| Cpt1a | Carnitine O-palmitoyltransferase 1, liver isoform | 37.425 | 28.763 | 0.003 | 0.769 |
| Lrp1 | Prolow-density lipoprotein receptor-related protein 1 | 102.596 | 77.790 | 0.003 | 0.758 |
| Aqr | RNA helicase aquarius | 16.361 | 12.218 | 0.048 | 0.747 |
| Ddb1 | DNA damage-binding protein 1 | 30.754 | 22.739 | 0.038 | 0.739 |
| Ubxn6 | UBX domain-containing protein 6 | 14.477 | 10.133 | 0.038 | 0.700 |
| Rpl22l1 | 60S ribosomal protein L22-like 1 | 27.692 | 19.173 | 0.040 | 0.692 |
| Glg1 | Golgi apparatus protein 1 | 34.516 | 22.497 | 0.009 | 0.652 |
| Apoe | Apolipoprotein E | 192.407 | 96.859 | 0.024 | 0.503 |
| Cd300ld3 | CMRF35-like molecule 3 | 17.868 | 8.905 | 0.006 | 0.498 |
| Lpl | Lipoprotein lipase | 35.613 | 13.226 | 0.027 | 0.446 |
| Csf1r | Macrophage colony-stimulating factor 1 receptor | 43.767 | 15.612 | 0.027 | 0.428 |
| Colec12 | Collectin-12 | 22.477 | 8.953 | 0.002 | 0.398 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teng, W.; Subsomwong, P.; Narita, K.; Nakane, A.; Asano, K. Heat Shock Protein SSA1 Enriched in Hypoxic Secretome of Candida albicans Exerts an Immunomodulatory Effect via Regulating Macrophage Function. Cells 2024, 13, 127. https://doi.org/10.3390/cells13020127
Teng W, Subsomwong P, Narita K, Nakane A, Asano K. Heat Shock Protein SSA1 Enriched in Hypoxic Secretome of Candida albicans Exerts an Immunomodulatory Effect via Regulating Macrophage Function. Cells. 2024; 13(2):127. https://doi.org/10.3390/cells13020127
Chicago/Turabian StyleTeng, Wei, Phawinee Subsomwong, Kouji Narita, Akio Nakane, and Krisana Asano. 2024. "Heat Shock Protein SSA1 Enriched in Hypoxic Secretome of Candida albicans Exerts an Immunomodulatory Effect via Regulating Macrophage Function" Cells 13, no. 2: 127. https://doi.org/10.3390/cells13020127
APA StyleTeng, W., Subsomwong, P., Narita, K., Nakane, A., & Asano, K. (2024). Heat Shock Protein SSA1 Enriched in Hypoxic Secretome of Candida albicans Exerts an Immunomodulatory Effect via Regulating Macrophage Function. Cells, 13(2), 127. https://doi.org/10.3390/cells13020127