
Human Platelet-Rich Plasma Facilitates Angiogenesis to Restore Impaired Uterine Environments with Asherman’s Syndrome for Embryo Implantation and Following Pregnancy in Mice

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. PRP Preparation
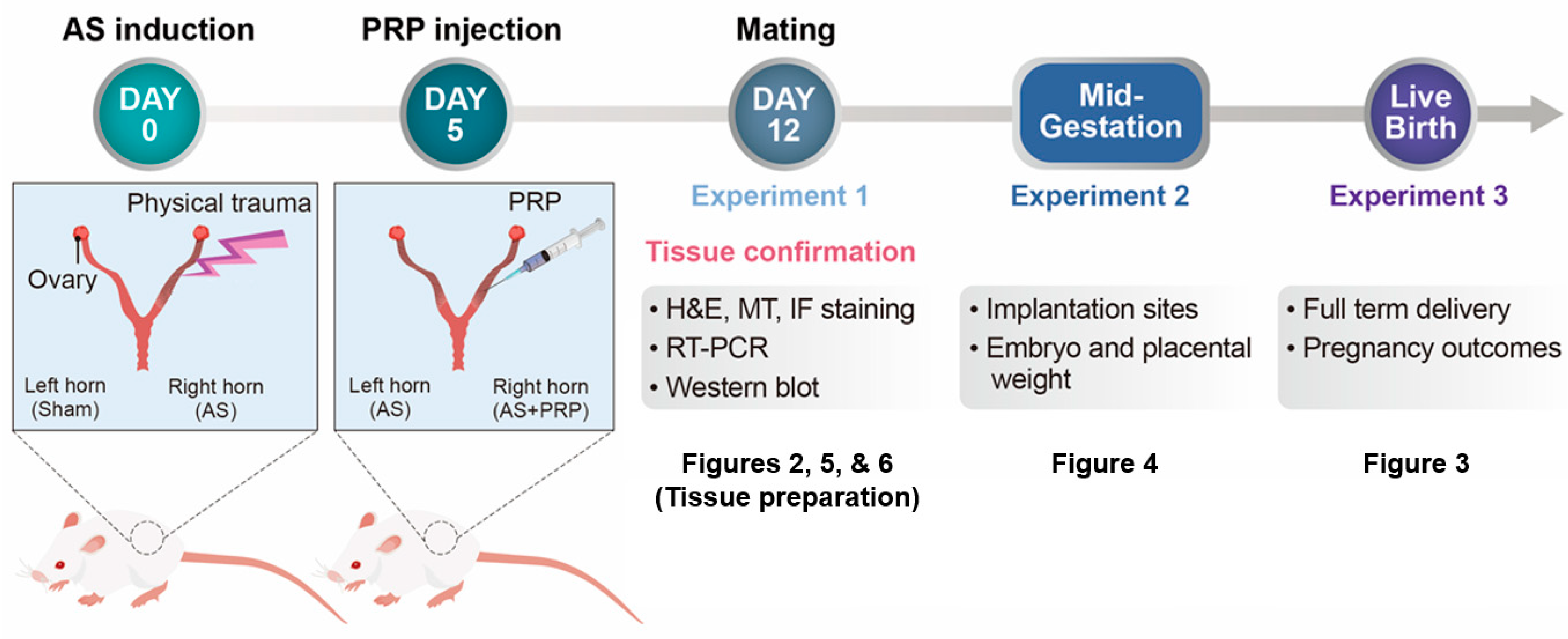
2.3. PRP Therapy to Experimentally Induced Murine Model of AS
2.4. Analyses of Embryo Implantation and Pregnancy Outcomes
2.5. RNA Extraction and Quantitative Real-Time RT-PCR (qRT-PCR)
2.6. Histological Staining
2.7. Western Blot Analyses
2.8. Scratch Wound-Healing Assay
2.9. Statistical Analysis
3. Results
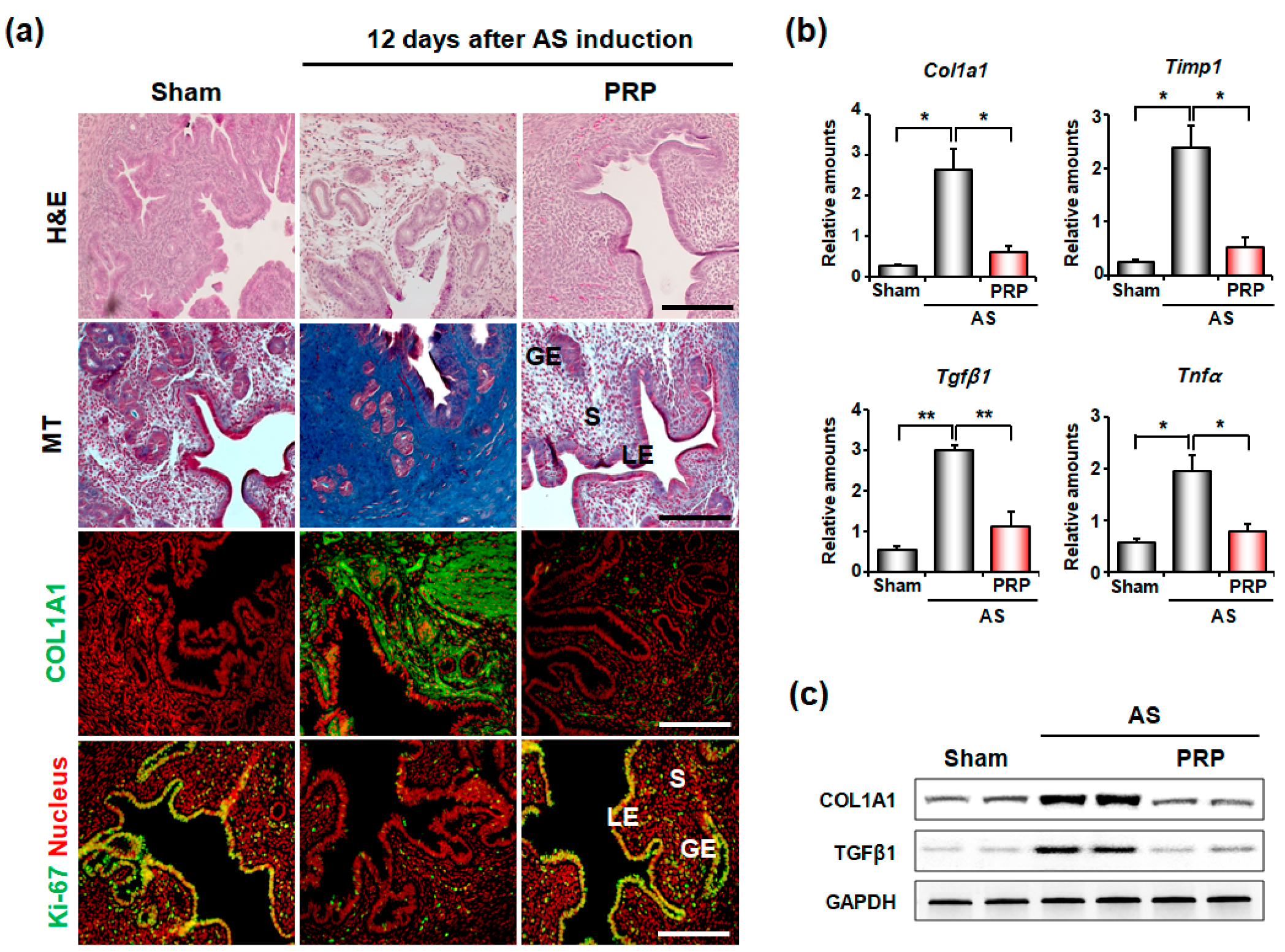
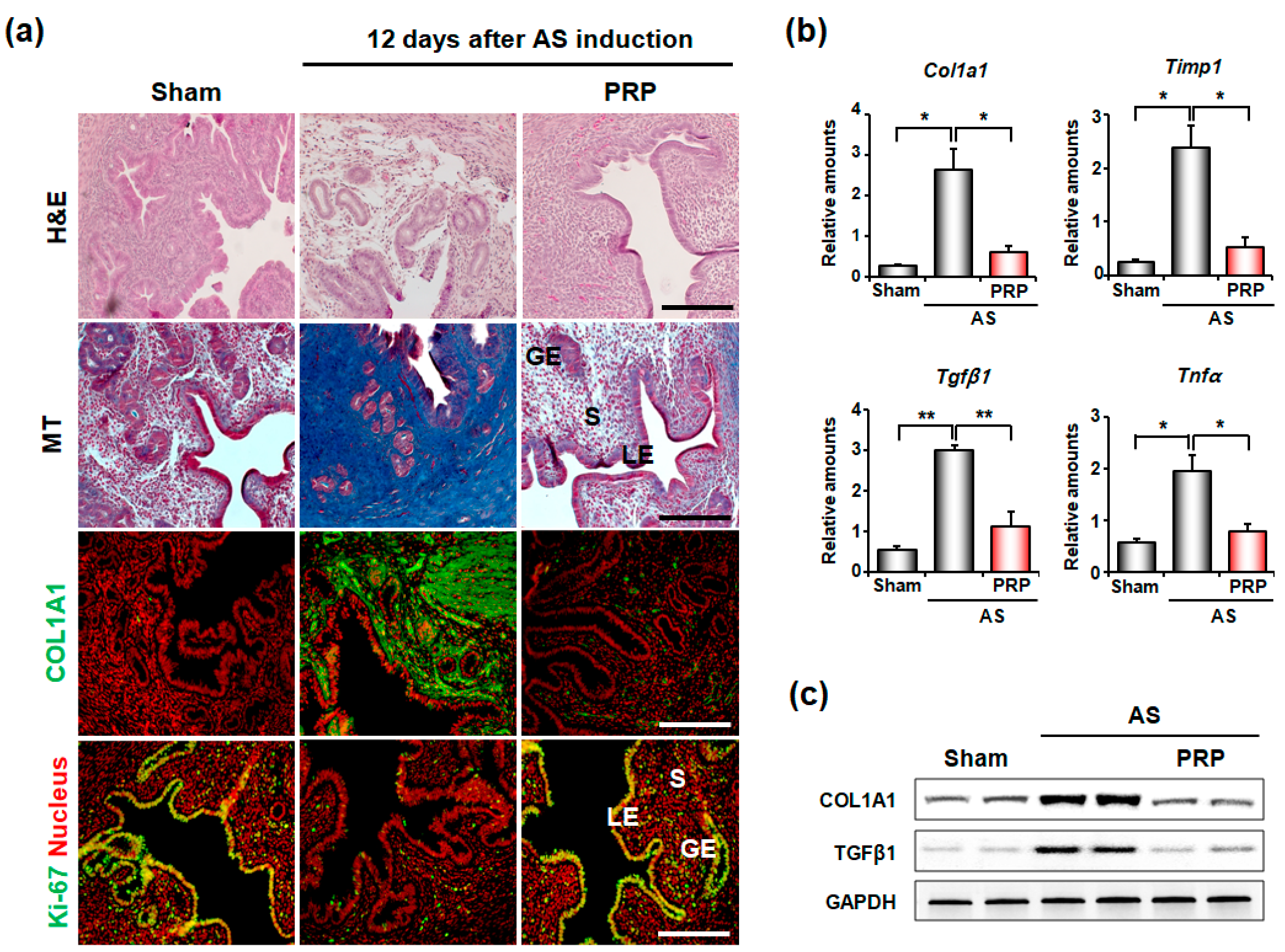
3.1. PRP Significantly Reduces Fibrosis in the Uteri of Mice with AS
3.2. PRP Ameliorates Oligomenorrhea and Rescues Infertility of Mice with AS
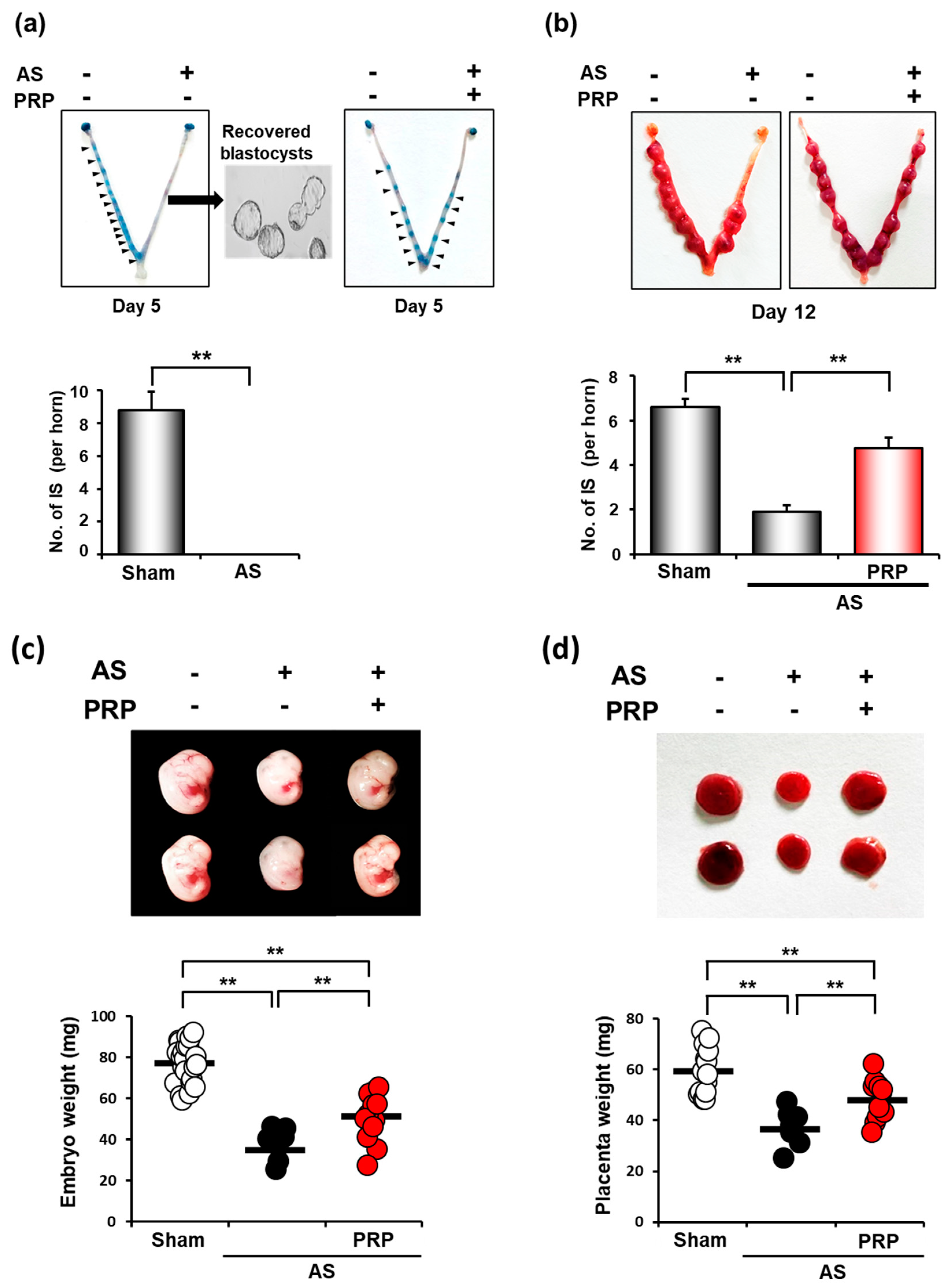
3.3. PRP Improves Poor Uterine Environment for Embryo Implantation Followed by Fetal Development in Mice with AS
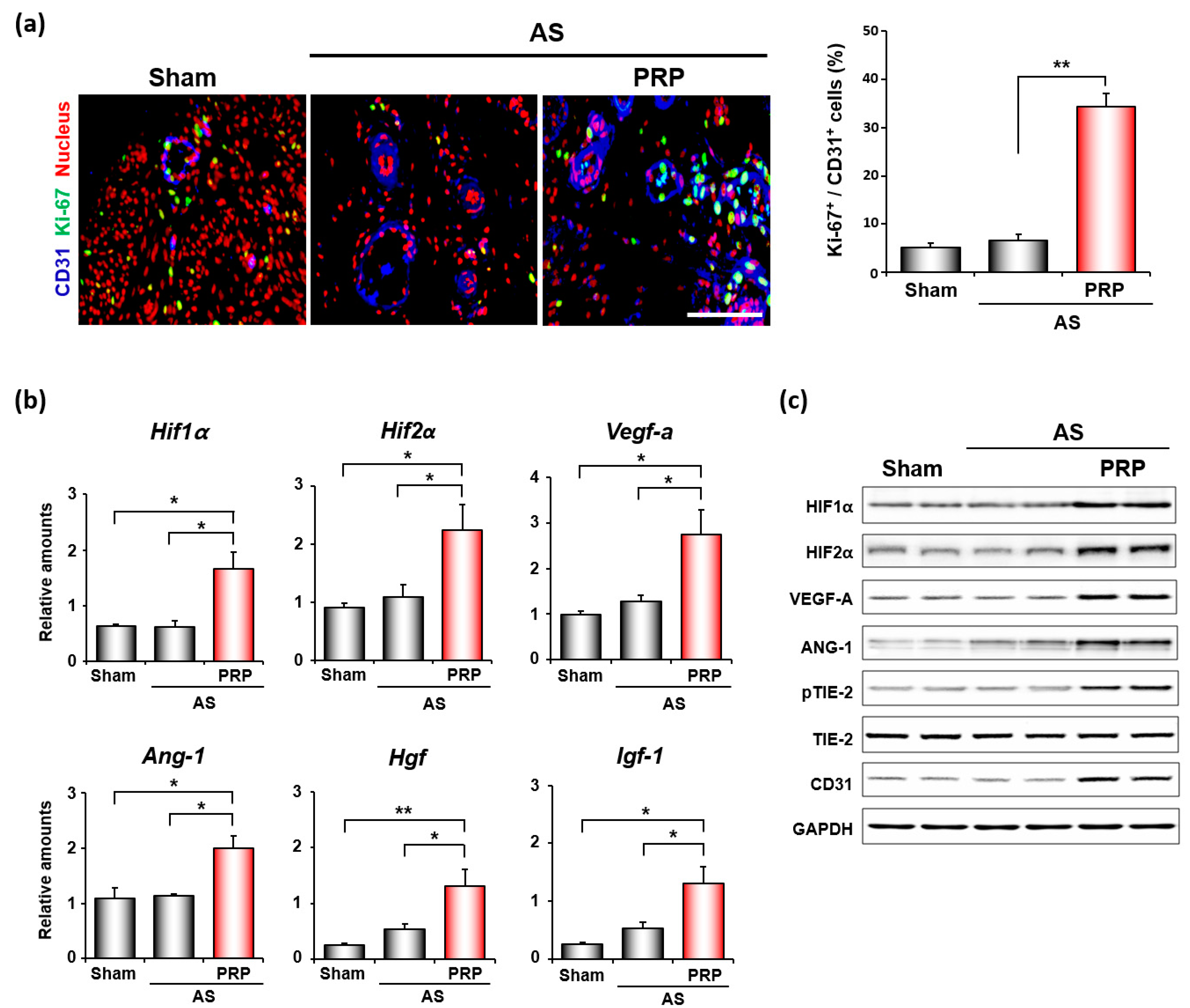
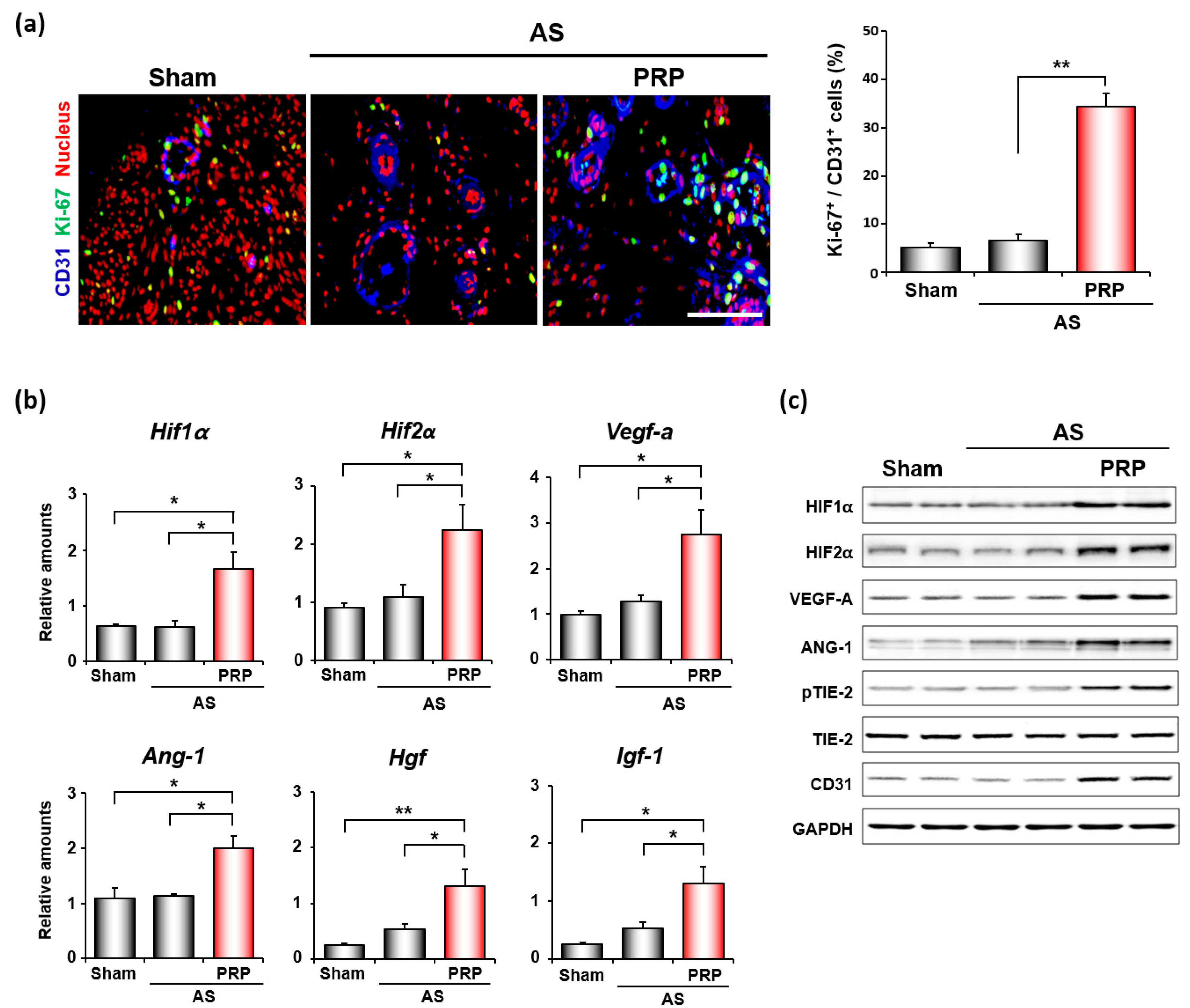
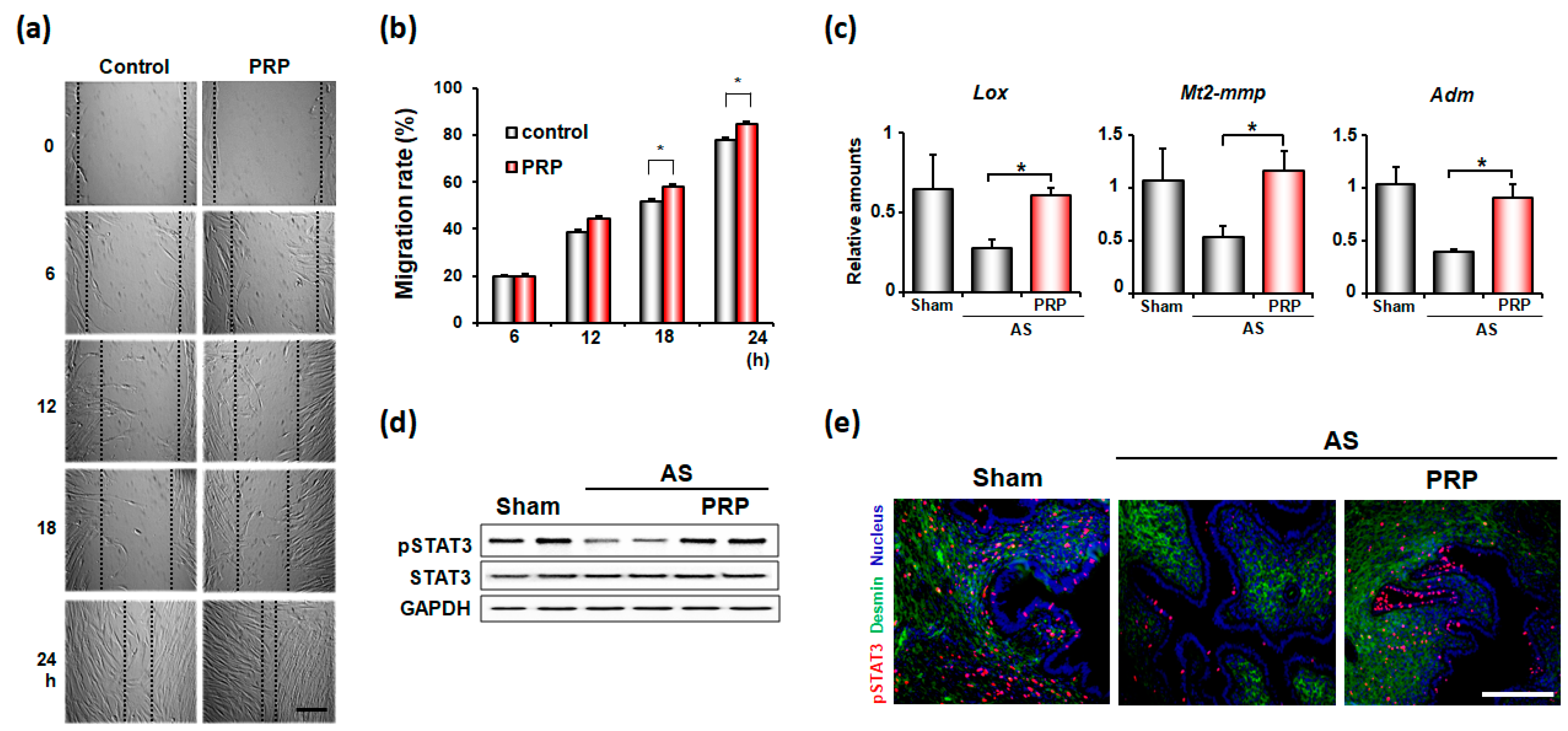
3.4. PRP Promotes Angiogenesis and Cell Migration for Endometrial Regeneration and Remodeling in Mice with AS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asherman, J.G. Amenorrhoea traumatica (atretica). J. Obstet. Gynaecol. Br. Emp. 1948, 55, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Wong, Y.M.; Cheong, Y.; Xia, E.; Li, T.C. Asherman syndrome--one century later. Fertil. Steril. 2008, 89, 759–779. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Liu, Y.; Streuli, I.; Dällenbach, P.; Dubuisson, J.; Ansaldi, Y.; Pluchino, N. Endometrial regeneration in asherman’s syndrome: Clinical and translational evidence of stem cell therapies. Curr. Stem Cell Res. Ther. 2019, 14, 454–459. [Google Scholar] [CrossRef]
- Khan, Z.; Goldberg, J.M. Hysteroscopic management of asherman’s syndrome. J. Minim. Invasive Gynecol. 2018, 25, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, X.; Cabanillas, S.; Cervelló, I.; Arbona, C.; Raga, F.; Ferro, J.; Palmero, J.; Remohí, J.; Pellicer, A.; Simón, C. Autologous cell therapy with cd133+ bone marrow-derived stem cells for refractory asherman’s syndrome and endometrial atrophy: A pilot cohort study. Hum. Reprod. 2016, 31, 1087–1096. [Google Scholar] [CrossRef] [Green Version]
- Mints, M.; Jansson, M.; Sadeghi, B.; Westgren, M.; Uzunel, M.; Hassan, M.; Palmblad, J. Endometrial endothelial cells are derived from donor stem cells in a bone marrow transplant recipient. Hum. Reprod. 2008, 23, 139–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, H.S. Endometrial cells derived from donor stem cells in bone marrow transplant recipients. JAMA 2004, 292, 81–85. [Google Scholar] [CrossRef] [Green Version]
- Strug, M.; Aghajanova, L. Making more womb: Clinical perspectives supporting the development and utilization of mesenchymal stem cell therapy for endometrial regeneration and infertility. J. Pers. Med. 2021, 11, 1364. [Google Scholar] [CrossRef]
- El-Sharkawy, H.; Kantarci, A.; Deady, J.; Hasturk, H.; Liu, H.; Alshahat, M.; Van Dyke, T.E. Platelet-rich plasma: Growth factors and pro- and anti-inflammatory properties. J. Periodontol. 2007, 78, 661–669. [Google Scholar] [CrossRef] [Green Version]
- Bertrand-Duchesne, M.P.; Grenier, D.; Gagnon, G. Epidermal growth factor released from platelet-rich plasma promotes endothelial cell proliferation in vitro. J. Periodontal Res. 2010, 45, 87–93. [Google Scholar] [CrossRef]
- Rolfe, K.J.; Grobbelaar, A.O. The growth receptors and their role in wound healing. Curr. Opin. Investig. Drugs 2010, 11, 1221–1228. [Google Scholar]
- Wang, L.; Chen, Z.; Wang, X. Clinical efficacy and acceptability of panretinal photocoagulation combined with conbercept for patients with proliferative diabetic retinopathy: A protocol for systematic review and meta-analysis. Medicine 2021, 100, e25611. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Lou, X.; Shen, T.; Sun, Y.; Xiao, Y.; Zheng, X.; Wang, X.; Peng, Y.; Guo, Y.; Guo, Y.; et al. Combination of ablative fractional carbon dioxide laser and platelet-rich plasma treatment to improve hypertrophic scars: A retrospective clinical observational study. Burn. Trauma 2021, 9, tkab016. [Google Scholar] [CrossRef] [PubMed]
- Sheth, U.; Dwyer, T.; Smith, I.; Wasserstein, D.; Theodoropoulos, J.; Takhar, S.; Chahal, J. Does platelet-rich plasma lead to earlier return to sport when compared with conservative treatment in acute muscle injuries? A systematic review and meta-analysis. Arthrosc. J. Arthrosc. Relat. Surg. 2018, 34, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Boztug, C.Y.; Karaagac Akyol, T.; Benlice, C.; Koc, M.A.; Doganay Erdogan, B.; Ozcebe, O.I.; Kuzu, M.A.; Akyol, C. Platelet-rich plasma treatment improves postoperative recovery in patients with pilonidal sinus disease: A randomized controlled clinical trial. BMC Surg. 2021, 21, 373. [Google Scholar] [CrossRef] [PubMed]
- Puente Gonzalo, E.; Alonso Pacheco, L.; Vega Jiménez, A.; Vitale, S.G.; Raffone, A.; Laganà, A.S. Intrauterine infusion of platelet-rich plasma for severe asherman syndrome: A cutting-edge approach. Updates Surg. 2021, 73, 2355–2362. [Google Scholar] [CrossRef]
- Aghajanova, L.; Cedars, M.I.; Huddleston, H.G. Platelet-rich plasma in the management of asherman syndrome: Case report. J. Assist. Reprod. Genet. 2018, 35, 771–775. [Google Scholar] [CrossRef]
- Nazari, L.; Salehpour, S.; Hoseini, S.; Zadehmodarres, S.; Azargashb, E. Effects of autologous platelet-rich plasma on endometrial expansion in patie.ents undergoing frozen-thawed embryo transfer: A double-blind rct. Int. J. Reprod. Biomed. 2019, 17, 443–448. [Google Scholar] [CrossRef]
- Chang, Y.; Li, J.; Wei, L.N.; Pang, J.; Chen, J.; Liang, X. Autologous platelet-rich plasma infusion improves clinical pregnancy rate in frozen embryo transfer cycles for women with thin endometrium. Medicine 2019, 98, e14062. [Google Scholar] [CrossRef]
- Jun, S.M.; Park, M.; Lee, J.Y.; Jung, S.; Lee, J.E.; Shim, S.H.; Song, H.; Lee, D.R. Single cell-derived clonally expanded mesenchymal progenitor cells from somatic cell nuclear transfer-derived pluripotent stem cells ameliorate the endometrial function in the uterus of a murine model with asherman’s syndrome. Cell Prolif. 2019, 52, e12597. [Google Scholar] [CrossRef]
- Alawadhi, F.; Du, H.; Cakmak, H.; Taylor, H.S. Bone marrow-derived stem cell (bmdsc) transplantation improves fertility in a murine model of asherman’s syndrome. PLoS ONE 2014, 9, e96662. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Hong, S.H.; Park, S.H.; Kim, Y.S.; Yang, S.C.; Kim, H.R.; Noh, S.; Na, S.; Lee, H.K.; Lim, H.J.; et al. Perivascular stem cell-derived cyclophilin a improves uterine environment with asherman’s syndrome via hif1α-dependent angiogenesis. Mol. Ther. J. Am. Soc. Gene Ther. 2020, 28, 1818–1832. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, L.; Mou, S.; Zhao, H.; Fang, J.; Xiang, Y.; Zhao, T.; Sha, T.; Ding, J.; Hao, C. Investigation of platelet-rich plasma in increasing proliferation and migration of endometrial mesenchymal stem cells and improving pregnancy outcome of patients with thin endometrium. J. Cell. Biochem. 2019, 120, 7403–7411. [Google Scholar] [CrossRef] [PubMed]
- Nunes, J.P.S.; Dias, A.A.M. Imagej macros for the user-friendly analysis of soft-agar and wound-healing assays. BioTechniques 2017, 62, 175–179. [Google Scholar] [CrossRef] [Green Version]
- Reichenstein, M.; Reich, R.; LeHoux, J.-G.; Hanukoglu, I. Acth induces timp-1 expression and inhibits collagenase in adrenal cortex cells. Mol. Cell. Endocrinol. 2004, 215, 109–114. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Liu, C.; Zhou, D.; Zhang, L. Tgf-β/smad pathway and its regulation in hepatic fibrosis. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2016, 64, 157–167. [Google Scholar] [CrossRef]
- Pilling, D.; Vakil, V.; Cox, N.; Gomer, R.H. Tnf-alpha-stimulated fibroblasts secrete lumican to promote fibrocyte differentiation. Proc. Natl. Acad. Sci. USA 2015, 112, 11929–11934. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Lim, H.; Paria, B.C.; Matsumoto, H.; Swift, L.L.; Morrow, J.; Bonventre, J.V.; Dey, S.K. Cytosolic phospholipase a2alpha is crucial [correction of a2alpha deficiency is crucial] for ‘on-time’ embryo implantation that directs subsequent development. Development 2002, 129, 2879–2889. [Google Scholar] [CrossRef]
- Nakamura, T.; Mizuno, S. The discovery of hepatocyte growth factor (hgf) and its significance for cell biology, life sciences and clinical medicine. Proc. Jpn. Acad. Ser. B. 2010, 86, 588–610. [Google Scholar] [CrossRef] [Green Version]
- Gan, Q.F.; Lim, Y.T.; Foo, C.N.; Yu, C.W.; Woon, C.K.; Cheong, S.K.; Leong, P.P. Incorporating insulin growth factor-1 into regenerative and personalized medicine for cardiovascular disease: A systematic review. Curr. Stem Cell Res. Ther. 2022. [Google Scholar] [CrossRef]
- Fukuhara, S.; Sako, K.; Noda, K.; Zhang, J.; Minami, M.; Mochizuki, N. Angiopoietin-1/tie2 receptor signaling in vascular quiescence and angiogenesis. Histol. Histopathol. 2010, 25, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Alsousou, J.; Ali, A.; Willett, K.; Harrison, P. The role of platelet-rich plasma in tissue regeneration. Platelets 2013, 24, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Park, M.; Paek, J.Y.; Lee, W.S.; Song, H.; Lyu, S.W. Intrauterine infusion of human platelet-rich plasma improves endometrial regeneration and pregnancy outcomes in a murine model of asherman’s syndrome. Front. Physiol. 2020, 11, 105. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Li, J.; Chen, Y.; Wei, L.; Yang, X.; Shi, Y.; Liang, X. Autologous platelet-rich plasma promotes endometrial growth and improves pregnancy outcome during in vitro fertilization. Int. J. Clin. Exp. Med. 2015, 8, 1286–1290. [Google Scholar]
- Zadehmodarres, S.; Salehpour, S.; Saharkhiz, N.; Nazari, L. Treatment of thin endometrium with autologous platelet-rich plasma: A pilot study. JBRA Assist. Reprod. 2017, 21, 54–56. [Google Scholar] [CrossRef]
- Eftekhar, M.; Neghab, N.; Naghshineh, E.; Khani, P. Can autologous platelet rich plasma expand endometrial thickness and improve pregnancy rate during frozen-thawed embryo transfer cycle? A randomized clinical trial. Taiwan. J. Obstet. Gynecol. 2018, 57, 810–813. [Google Scholar] [CrossRef]
- Zhang, S.; Li, P.; Yuan, Z.; Tan, J. Platelet-rich plasma improves therapeutic effects of menstrual blood-derived stromal cells in rat model of intrauterine adhesion. Stem Cell Res. Ther. 2019, 10, 61. [Google Scholar] [CrossRef]
- Jang, H.Y.; Myoung, S.M.; Choe, J.M.; Kim, T.; Cheon, Y.P.; Kim, Y.M.; Park, H. Effects of autologous platelet-rich plasma on regeneration of damaged endometrium in female rats. Yonsei Med. J. 2017, 58, 1195–1203. [Google Scholar] [CrossRef]
- Xu, X.-W.; Zhang, J.; Guo, Z.-W.; Song, M.-M.; Sun, R.; Jin, X.-Y.; Su, J.-D.; Sun, B.-W. A narrative review of research progress on the relationship between hypoxia-inducible factor-2α and wound angiogenesis. Ann. Palliat. Med. 2021, 10, 4882–4888. [Google Scholar] [CrossRef]
- Matsumoto, H.; Nasu, K.; Nishida, M.; Ito, H.; Bing, S.; Miyakawa, I. Regulation of proliferation, motility, and contractility of human endometrial stromal cells by platelet-derived growth factor. J. Clin. Endocrinol. Metab. 2005, 90, 3560–3567. [Google Scholar] [CrossRef]
- Gargett, C.E.; Nguyen, H.P.; Ye, L. Endometrial regeneration and endometrial stem/progenitor cells. Rev. Endocr. Metab. Disord. 2012, 13, 235–251. [Google Scholar] [CrossRef] [PubMed]
- Carlson, M.J.; Thiel, K.W.; Yang, S.; Leslie, K.K. Catch it before it kills: Progesterone, obesity, and the prevention of endometrial cancer. Discov. Med. 2012, 14, 215–222. [Google Scholar] [PubMed]
- Gargett, C.E.; Chan, R.W.S.; Schwab, K.E. Hormone and growth factor signaling in endometrial renewal: Role of stem/progenitor cells. Mol. Cell. Endocrinol. 2008, 288, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Kim, Y.S.; Yoon, J.A.; Yang, S.C.; Park, M.; Seol, D.W.; Lyu, S.W.; Jun, J.H.; Lim, H.J.; Lee, D.R.; et al. Estrogen induces egr1 to fine-tune its actions on uterine epithelium by controlling pr signaling for successful embryo implantation. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2018, 32, 1184–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilcox, A.J.; Baird, D.D.; Weinberg, C.R. Time of implantation of the conceptus and loss of pregnancy. N. Engl. J. Med. 1999, 340, 1796–1799. [Google Scholar] [CrossRef] [PubMed]
- March, C.M. Management of asherman’s syndrome. Reprod. Biomed. Online 2011, 23, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Baradwan, S.; Baradwan, A.; Bashir, M.; Al-Jaroudi, D. The birth weight in pregnant women with asherman syndrome compared to normal intrauterine cavity: A case-control study. Medicine 2018, 97, e11797. [Google Scholar] [CrossRef]
- Kim, H.; Shin, J.E.; Koo, H.S.; Kwon, H.; Choi, D.H.; Kim, J.H. Effect of autologous platelet-rich plasma treatment on refractory thin endometrium during the frozen embryo transfer cycle: A pilot study. Front. Endocrinol. 2019, 10, 61. [Google Scholar] [CrossRef] [Green Version]
- Kusumi, M.; Ihana, T.; Kurosawa, T.; Ohashi, Y.; Tsutsumi, O. Intrauterine administration of platelet-rich plasma improves embryo implantation by increasing the endometrial thickness in women with repeated implantation failure: A single-arm self-controlled trial. Reprod. Med. Biol. 2020, 19, 350–356. [Google Scholar] [CrossRef]
- Dogra, Y.; Singh, N.; Vanamail, P. Autologous platelet-rich plasma optimizes endometrial thickness and pregnancy outcomes in women with refractory thin endometrium of varied aetiology during fresh and frozen-thawed embryo transfer cycles. JBRA Assist. Reprod. 2022, 26, 13–21. [Google Scholar] [CrossRef]
- Sherer, D.M.; Abulafia, O. Angiogenesis during implantation, and placental and early embryonic development. Placenta 2001, 22, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Maybin, J.A.; Murray, A.A.; Saunders, P.T.K.; Hirani, N.; Carmeliet, P.; Critchley, H.O.D. Hypoxia and hypoxia inducible factor-1α are required for normal endometrial repair during menstruation. Nat. Commun. 2018, 9, 295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Yi, H.; Li, T.C.; Wang, Y.; Wang, H.; Chen, X. Role of vascular endothelial growth factor (vegf) in human embryo implantation: Clinical implications. Biomolecules 2021, 11, 253. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Chen, C.D.; Chou, C.H.; Wen, W.F.; Tsao, P.N.; Lee, H.; Chen, S.U. Intentional endometrial injury increases embryo implantation potentials through enhanced endometrial angiogenesis. Biol. Reprod. 2019, 100, 381–389. [Google Scholar] [CrossRef]
- Plaisier, M.; Dennert, I.; Rost, E.; Koolwijk, P.; van Hinsbergh, V.W.; Helmerhorst, F.M. Decidual vascularization and the expression of angiogenic growth factors and proteases in first trimester spontaneous abortions. Hum. Reprod. 2009, 24, 185–197. [Google Scholar] [CrossRef] [Green Version]
- Aghajanova, L.; Houshdaran, S.; Balayan, S.; Manvelyan, E.; Irwin, J.C.; Huddleston, H.G.; Giudice, L.C. In vitro evidence that platelet-rich plasma stimulates cellular processes involved in endometrial regeneration. J. Assist. Reprod. Genet. 2018, 35, 757–770. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.; Li, P.; Wang, Q.; Li, Y.; Li, X.; Zhao, D.; Xu, X.; Kong, L. Autologous menstrual blood-derived stromal cells transplantation for severe asherman’s syndrome. Hum. Reprod. 2016, 31, 2723–2729. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Escudero, J.; Moreno, V.; Martín-Alonso, M.; Hernández-Riquer, M.V.; Feinberg, T.; Colmenar, Á.; Calvo, E.; Camafeita, E.; Martínez, F.; Oudhoff, M.J. E-cadherin cleavage by mt2-mmp regulates apical junctional signaling and epithelial homeostasis in the intestine. J. Cell Sci. 2017, 130, 4013–4027. [Google Scholar] [CrossRef] [Green Version]
- Hotary, K.; Li, X.Y.; Allen, E.; Stevens, S.L.; Weiss, S.J. A cancer cell metalloprotease triad regulates the basement membrane transmigration program. Genes Dev. 2006, 20, 2673–2686. [Google Scholar] [CrossRef] [Green Version]
- Erler, J.T.; Bennewith, K.L.; Cox, T.R.; Lang, G.; Bird, D.; Koong, A.; Le, Q.T.; Giaccia, A.J. Hypoxia-induced lysyl oxidase is a critical mediator of bone marrow cell recruitment to form the premetastatic niche. Cancer Cell 2009, 15, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Hikage, F.; Atkins, S.; Kahana, A.; Smith, T.J.; Chun, T.H. Hif2a-lox pathway promotes fibrotic tissue remodeling in thyroid-associated orbitopathy. Endocrinology 2019, 160, 20–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Zhou, Y.; Wang, L.; Zhang, J.; Wu, H.; Xiong, J.; Zhang, J.; Tian, Y.; Wang, C.; Wu, H. Transcriptional upregulation of mt2-mmp in response to hypoxia is promoted by hif-1α in cancer cells. Mol. Carcinog. 2011, 50, 770–780. [Google Scholar] [CrossRef] [PubMed]
- Karli, P.; Bildircin, F.D.; Ozdemir, A.Z.; Avci, B. Higher adrenomedullin levels during implantation are associated with successful pregnancy. Future Sci. OA 2020, 6, FSO453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, L.; Hirota, Y.; Saito-Fujita, T.; Takeda, N.; Tanaka, T.; Hiraoka, T.; Akaeda, S.; Fujita, H.; Shimizu-Hirota, R.; Igaue, S.; et al. Hif2alpha in the uterine stroma permits embryo invasion and luminal epithelium detachment. J. Clin. Investig. 2018, 128, 3186–3197. [Google Scholar] [CrossRef] [Green Version]
- Hiraoka, T.; Hirota, Y.; Fukui, Y.; Gebril, M.; Kaku, T.; Aikawa, S.; Hirata, T.; Akaeda, S.; Matsuo, M.; Haraguchi, H.; et al. Differential roles of uterine epithelial and stromal stat3 coordinate uterine receptivity and embryo attachment. Sci. Rep. 2020, 10, 15523. [Google Scholar] [CrossRef]
- Choi, Y.; Kim, H.R.; Lim, E.J.; Park, M.; Yoon, J.A.; Kim, Y.S.; Kim, E.K.; Shin, J.E.; Kim, J.H.; Kwon, H.; et al. Integrative analyses of uterine transcriptome and micrornaome reveal compromised lif-stat3 signaling and progesterone response in the endometrium of patients with recurrent/repeated implantation failure (rif). PLoS ONE 2016, 11, e0157696. [Google Scholar] [CrossRef] [Green Version]
- Hiraoka, T.; Hirota, Y.; Saito-Fujita, T.; Matsuo, M.; Egashira, M.; Matsumoto, L.; Haraguchi, H.; Dey, S.K.; Furukawa, K.S.; Fujii, T.; et al. Stat3 accelerates uterine epithelial regeneration in a mouse model of decellularized uterine matrix transplantation. JCI Insight 2016, 1, e87591. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.K.; Yoon, J.A.; Yoon, S.Y.; Park, M.; Lee, W.S.; Lyu, S.W.; Song, H. Human Platelet-Rich Plasma Facilitates Angiogenesis to Restore Impaired Uterine Environments with Asherman’s Syndrome for Embryo Implantation and Following Pregnancy in Mice. Cells 2022, 11, 1549. https://doi.org/10.3390/cells11091549
Kim MK, Yoon JA, Yoon SY, Park M, Lee WS, Lyu SW, Song H. Human Platelet-Rich Plasma Facilitates Angiogenesis to Restore Impaired Uterine Environments with Asherman’s Syndrome for Embryo Implantation and Following Pregnancy in Mice. Cells. 2022; 11(9):1549. https://doi.org/10.3390/cells11091549
Chicago/Turabian StyleKim, Min Kyoung, Jung Ah Yoon, Sook Young Yoon, Mira Park, Woo Sik Lee, Sang Woo Lyu, and Haengseok Song. 2022. "Human Platelet-Rich Plasma Facilitates Angiogenesis to Restore Impaired Uterine Environments with Asherman’s Syndrome for Embryo Implantation and Following Pregnancy in Mice" Cells 11, no. 9: 1549. https://doi.org/10.3390/cells11091549
APA StyleKim, M. K., Yoon, J. A., Yoon, S. Y., Park, M., Lee, W. S., Lyu, S. W., & Song, H. (2022). Human Platelet-Rich Plasma Facilitates Angiogenesis to Restore Impaired Uterine Environments with Asherman’s Syndrome for Embryo Implantation and Following Pregnancy in Mice. Cells, 11(9), 1549. https://doi.org/10.3390/cells11091549