
Osteogenic Commitment of Human Periodontal Ligament Cells Is Predetermined by Methylation, Chromatin Accessibility and Expression of Key Transcription Factors


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
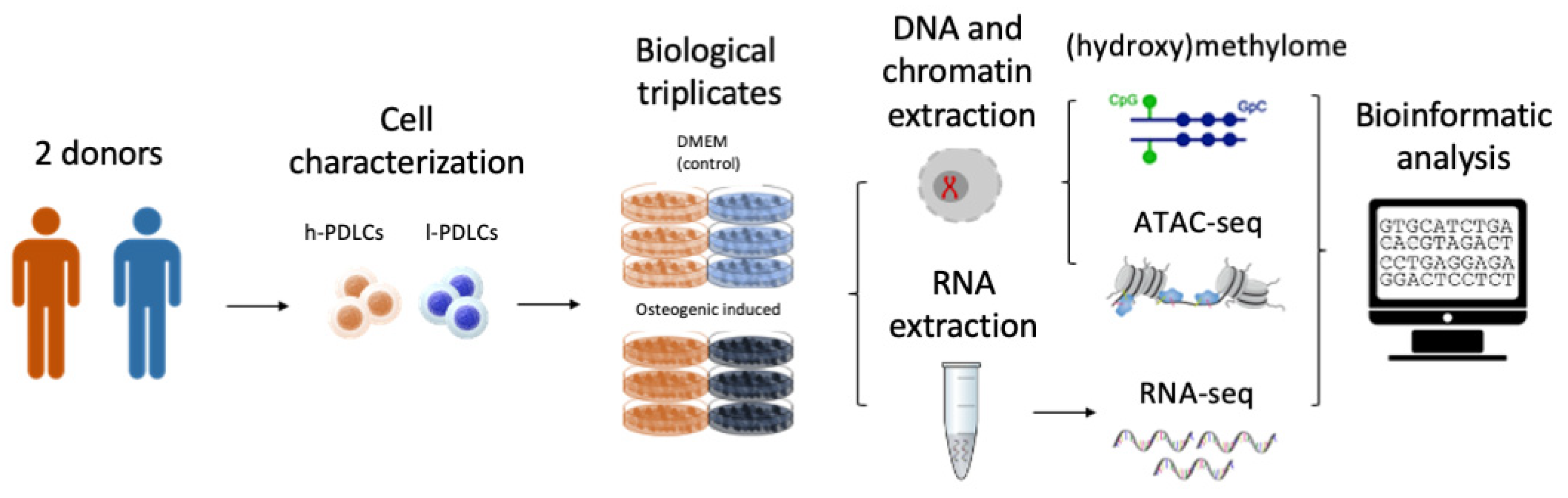
2. Materials and Methods
2.1. Cell Acquisition and Characterization
2.2. Cell Culture and Osteogenic Stimulation
2.3. mRNA Isolation and RNA-Seq
2.4. RNA-Seq Data Processing
2.5. Real Time q-PCR RNA Analysis
2.6. Assay for Transposase-Accessible Chromatin Using Sequencing (ATAC-Seq)
2.7. ATAC-Seq Genome Alignment and Peak Calling
2.8. TF Motif Analysis
2.9. DNA Isolation
2.10. Oxidative Bisulfite Conversion and DNA (Hydroxy)Methylome
2.11. (Hydroxy)Methylation Data Processing
3. Results
3.1. l-PDLCs Are Characterised by Neuronal Rather Than Osteoblastic Cell Pre-Commitment
3.2. Osteogenic Commitment in PDLCs Is Predefined by Chromatin Accessibility and DNA Methylation
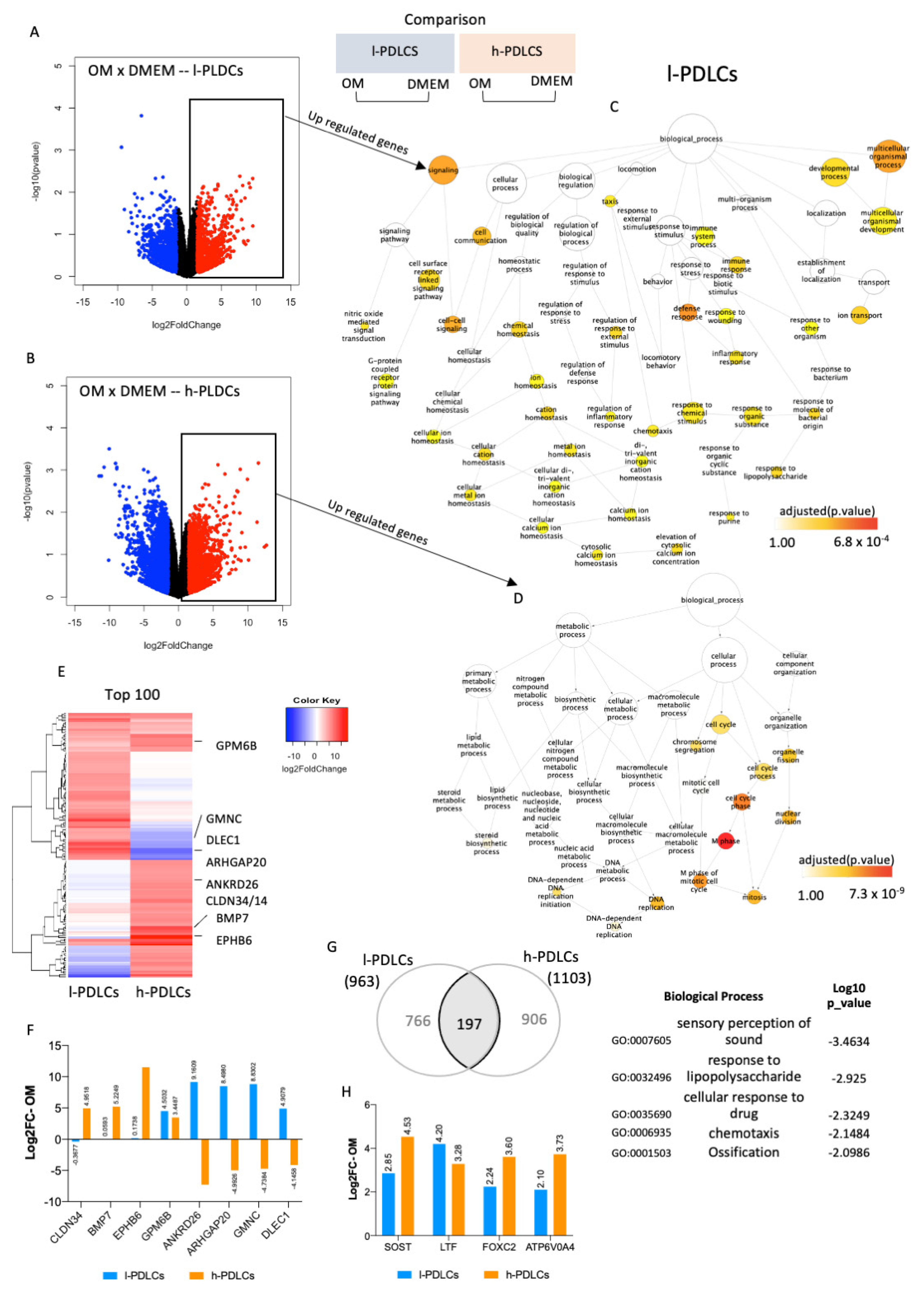
3.3. Genes and Biological Processes Related to Acquisition of Osteoblast Phenotype Are Further Activated in h-PDLCs upon Osteogenic Stimulation
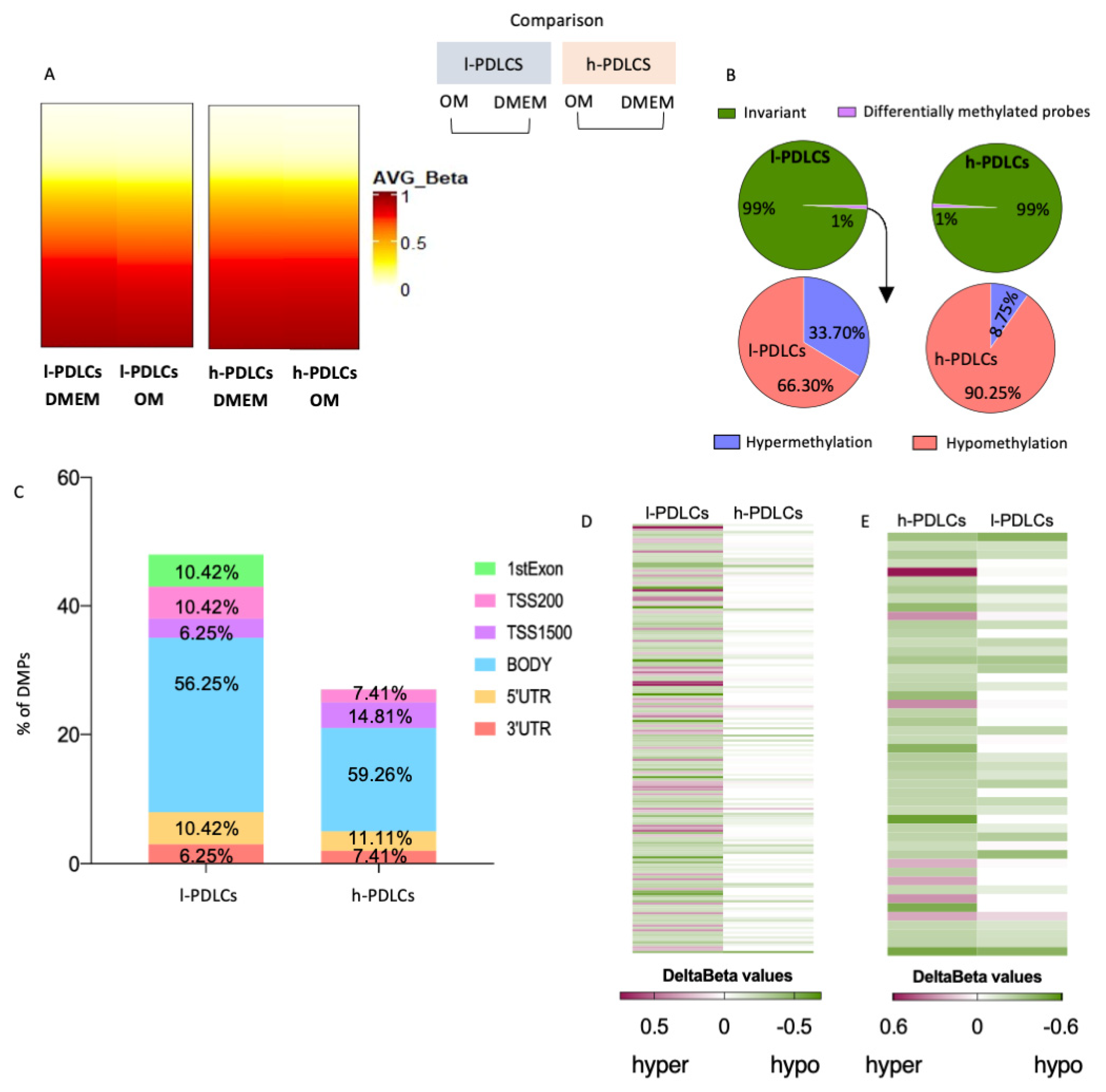
3.4. Transcriptional Changes upon Osteogenic Induction Are Not Accompanied by Major Epigenetic Restructure
3.5. Progression in Osteoblast Phenotype Following Osteogenic Induction Is Limited by Persistent DNA Hypermethylation in l-PDLCs
3.6. Chromatin Accessibility Supports Gene Transcription Related to Osteoblast Function in Stimulated h-PDLCs
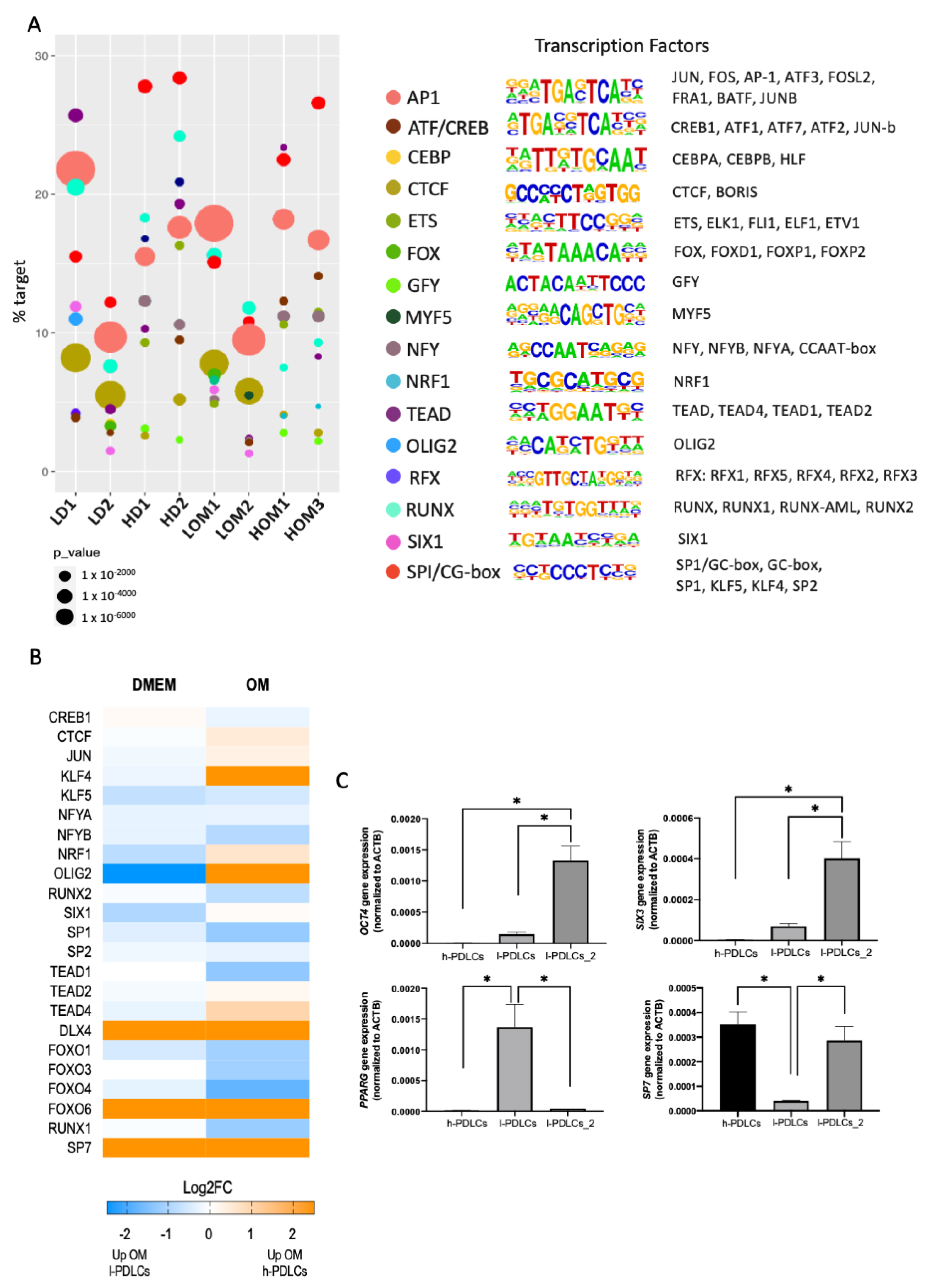
3.7. Non-Osteogenic TFs Are Uniquely Involved in l-PDLCs and Could Be Used to Predict Low Osteogenic Potential
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seo, B.; Miura, M.; Gronthos, S.; Mark Bartold, P.; Batouli, S.; Brahim, J.; Young, M.; Gehron Robey, P.; Wang, C.Y.; Shi, S. Investigation of multipotent postnatal stem cells from human periodontal ligament. Lancet 2004, 364, 149–155. [Google Scholar] [CrossRef]
- Gay, I.; Chen, S.; MacDougall, M. Isolation and characterization of multipotent human periodontal ligament stem cells. Orthod. Craniofac. Res. 2007, 10, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.T.-J.; Gronthos, S.; Shi, S. Mesenchymal Stem Cells Derived from Dental Tissues vs. Those from Other Sources: Their Biology and Role in Regenerative Medicine. J. Dent. Res. 2009, 88, 792–806. [Google Scholar] [CrossRef] [PubMed]
- Iwata, T.; Yamato, M.; Zhang, Z.; Mukobata, S.; Washio, K.; Ando, T.; Feijen, J.; Okano, T.; Ishikawa, I. Validation of human periodontal ligament-derived cells as a reliable source for cytotherapeutic use. J. Clin. Periodontol. 2010, 37, 1088–1099. [Google Scholar] [CrossRef]
- Assis, R.I.F.; Schmidt, A.G.; Racca, F.; da Silva, R.A.; Zambuzzi, W.F.; Silvério, K.G.; Nociti, F.H.; Pecorari, V.G.; Wiench, M.; Andia, D.C. DNMT1 Inhibitor Restores RUNX2 Expression and Mineralization in Periodontal Ligament Cells. DNA Cell Biol. 2021, 40, 662–674. [Google Scholar] [CrossRef]
- Saito, M.T.; Salmon, C.R.; Amorim, B.R.; Ambrosano, G.M.B.; Casati, M.Z.; Sallum, E.A.; Nociti, F.H.; Silvério, K.G. Characterization of Highly Osteoblast/Cementoblast Cell Clones From a CD105-Enriched Periodontal Ligament Progenitor Cell Population. J. Periodontol. 2014, 85, e205–e211. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.S.; Assis, R.I.F.; Feltran, G.D.; do Rosário Palma, I.C.; Françoso, B.G.; Zambuzzi, W.F.; Andia, D.C.; Silva, R.A. Genome-wide DNA (hydroxy) methylation reveals the individual epigenetic landscape importance on osteogenic phenotype acquisition in periodontal ligament cells. J. Periodontol. 2022, 93, 435–448. [Google Scholar] [CrossRef]
- Assis, R.I.F.; Feltran, G.D.; Silva, M.E.S.; do Rosario Palma, I.C.; Rovai, E.S.; de Miranda, T.B.; Ferreira, M.R.; Zambuzzi, W.F.; Birbrair, A.; Andia, D.C.; et al. Non-coding RNAs repressive role in post-transcriptional processing of RUNX2 during the acquisition of the osteogenic phenotype of periodontal ligament mesenchymal stem cells. Dev. Biol. 2021, 470, 37–48. [Google Scholar] [CrossRef]
- Vincent, A.; Van Seuningen, I. Epigenetics, stem cells and epithelial cell fate. Differentiation 2009, 78, 99–107. [Google Scholar] [CrossRef]
- Ai, T.; Zhang, J.; Wang, X.; Zheng, X.; Qin, X.; Zhang, Q.; Li, W.; Hu, W.; Lin, J.; Chen, F. DNA methylation profile is associated with the osteogenic potential of three distinct human odontogenic stem cells. Signal. Transduct. Target. Ther. 2018, 3, 1. [Google Scholar] [CrossRef]
- Bird, A. DNA methylation patterns and epigenetic memory. Genes Dev. 2002, 16, 6–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teven, C.M.; Liu, X.; Hu, N.; Tang, N.; Kim, S.H.; Huang, E.; Yang, K.; Li, M.; Gao, J.-L.; Liu, H.; et al. Epigenetic regulation of mesenchymal stem cells: A focus on osteogenic and adipogenic differentiation. Stem Cells Int. 2011, 2011, 201371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F.; Shen, H.; Deng, H.W. Systemic analysis of osteoblast-specific DNA methylation marks reveals novel epigenetic basis of osteoblast differentiation. Bone Rep. 2017, 6, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.A. Functions of DNA methylation: Islands, start sites, gene bodies and beyond. Nat. Rev. Genet. 2012, 13, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Lokk, K.; Modhukur, V.; Rajashekar, B.; Märtens, K.; Mägi, R.; Kolde, R.; Koltšina, M.; Nilsson, T.K.; Vilo, J.; Salumets, A.; et al. DNA methylome profiling of human tissues identifies global and tissue-specific methylation patterns. Genome Biol. 2014, 15, 3248. [Google Scholar] [CrossRef] [Green Version]
- Visone, R.; Bacalini, M.G.; Di Franco, S.; Ferracin, M.; Scavo, E.; Bassi, C.; Saccenti, E.; Nicotra, A.; Grzes, M. DNA methylation of shelf, shore and open sea CpG positions distinguish high microsatellite instability from low or stable microsatellite status colon cancer stem cells. Epigenomics 2019, 11, 587–604. [Google Scholar] [CrossRef] [Green Version]
- Wiench, M.; Miranda, T.B.; Hager, G.L. Control of nuclear receptor function by local chromatin structure. FEBS J. 2011, 278, 2211–2230. [Google Scholar] [CrossRef] [Green Version]
- Ponnaluri, V.K.C.; Ehrlich, K.C.; Zhang, G.; Lacey, M.; Johnston, D.; Pradhan, S.; Ehrlich, M. Association of 5-hydroxymethylation and 5-methylation of DNA cytosine with tissue-specific gene expression. Epigenetics 2017, 12, 123–138. [Google Scholar] [CrossRef]
- Ho, Y.-T.; Shimbo, T.; Wijaya, E.; Ouchi, Y.; Takaki, E.; Yamamoto, R.; Kikuchi, Y.; Kaneda, Y.; Tamai, K. Chromatin accessibility identifies diversity in mesenchymal stem cells from different tissue origins. Sci. Rep. 2018, 8, 17765. [Google Scholar] [CrossRef]
- Wiench, M.; John, S.; Baek, S.; Johnson, T.A.; Sung, M.H.; Escobar, T.; Simmons, C.A.; Pearce, K.H.; Biddie, S.C.; Sabo, P.J.; et al. DNA methylation status predicts cell type-specific enhancer activity. EMBO J. 2011, 30, 3028–3039. [Google Scholar] [CrossRef] [Green Version]
- Wiench, M.; Hager, G.L. Expanding horizons for nuclear receptors. EMBO Rep. 2010, 11, 569–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, K.; Moore, J.E.; Zhang, X.; Weng, Z. Genetic and epigenetic features of promoters with ubiquitous chromatin accessibility support ubiquitous transcription of cell-essential genes. Nucleic Acids Res. 2021, 49, 5705–5725. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chen, T.; Sun, W.; Yuan, Z.; Yu, M.; Chen, G.; Guo, W.; Xiao, J.; Tian, W. DNA Demethylation Rescues the Impaired Osteogenic Differentiation Ability of Human Periodontal Ligament Stem Cells in High Glucose. Sci. Rep. 2016, 6, 27447. [Google Scholar] [CrossRef] [PubMed]
- Silvério, K.G.; Rodrigues, T.L.; Coletta, R.D.; Benevides, L.; Da Silva, J.S.; Casati, M.Z.; Sallum, E.A.; Nociti, F.H. Mesenchymal Stem Cell Properties of Periodontal Ligament Cells From Deciduous and Permanent Teeth. J. Periodontol. 2010, 81, 1207–1215. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Assis, R.I.F.; Wiench, M.; Silvério, K.G.; da Silva, R.A.; da Silva Feltran, G.; Sallum, E.A.; Casati, M.Z.; Nociti, F.H.; Andia, D.C. RG108 increases NANOG and OCT4 in bone marrow-derived mesenchymal cells through global changes in DNA modifications and epigenetic activation. PLoS ONE 2018, 13, e0207873. [Google Scholar] [CrossRef] [PubMed]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO Summarizes and Visualizes Long Lists of Gene Ontology Terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Buenrostro, J.D.; Giresi, P.G.; Zaba, L.C.; Chang, H.Y.; Greenleaf, W.J. Transposition of native chromatin for fast and sensitive epigenomic profiling of open chromatin, DNA-binding proteins and nucleosome position. Nat. Methods 2013, 10, 1213–1218. [Google Scholar] [CrossRef] [PubMed]
- Corces, M.R.; Buenrostro, J.D.; Wu, B.; Greenside, P.G.; Chan, S.M.; Koenig, J.L.; Snyder, M.P.; Pritchard, J.K.; Kundaje, A.; Greenleaf, W.J.; et al. Lineage-specific and single-cell chromatin accessibility charts human hematopoiesis and leukemia evolution. Nat. Genet. 2016, 48, 1193–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinz, S.; Benner, C.; Spann, N.; Bertolino, E.; Lin, Y.C.; Laslo, P.; Cheng, J.X.; Murre, C.; Singh, H.; Glass, C.K. Simple Combinations of Lineage-Determining Transcription Factors Prime cis-Regulatory Elements Required for Macrophage and B Cell Identities. Mol. Cell 2010, 38, 576–589. [Google Scholar] [CrossRef] [Green Version]
- Aryee, M.J.; Jaffe, A.E.; Corrada-Bravo, H.; Ladd-Acosta, C.; Feinberg, A.P.; Hansen, K.D.; Irizarry, R.A. Minfi: A flexible and comprehensive Bioconductor package for the analysis of Infinium DNA methylation microarrays. Bioinformatics 2014, 30, 1363–1369. [Google Scholar] [CrossRef] [Green Version]
- Fortin, J.P.; Triche, T.J., Jr.; Hansen, K.D. Genome analysis Preprocessing, normalization and integration of the Illumina HumanMethylationEPIC array with minfi. Bioinformatics 2017, 33, 558–560. [Google Scholar] [CrossRef]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2—Elegant Graphics for Data Analysis. In Measurement: Interdisciplinary Research and Perspectives, 2nd ed.; Routledge Taylor & Francis Group: Abingdon, UK, 2016; Volume 77, ISBN 9783319242750. [Google Scholar]
- Chen, H.; Boutros, P.C. VennDiagram: A package for the generation of highly-customizable Venn and Euler diagrams in R. BMC Bioinform. 2011, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Huck, K.; Sens, C.; Wuerfel, C.; Zoeller, C.; Nakchbandi, A.I. The Rho GTPase RAC1 in Osteoblasts Controls Their Function. Int. J. Mol. Sci. 2020, 21, 385. [Google Scholar] [CrossRef] [Green Version]
- Américo-Da-Silva, L.; Diaz, J.; Bustamante, M.; Mancilla, G.; Oyarzún, I.; Verdejo, H.E.; Quiroga, C. A new role for HERPUD1 and ERAD activation in osteoblast differentiation and mineralization. FASEB J. 2018, 32, 4681–4695. [Google Scholar] [CrossRef] [Green Version]
- Horiuchi, K.; Tohmonda, T.; Morioka, H. The unfolded protein response in skeletal development and homeostasis. Cell. Mol. Life Sci. 2016, 73, 2851–2869. [Google Scholar] [CrossRef] [PubMed]
- Javed, A.; Chen, H.; Ghori, F.Y. Genetic and Transcriptional Control of Bone Formation. Oral Maxillofac. Surg. Clin. N. Am. 2010, 22, 283–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’haeseleer, P. What are DNA sequence motifs? Nat. Biotechnol. 2006, 24, 423–425. [Google Scholar] [CrossRef] [PubMed]
- Tarkkonen, K.; Hieta, R.; Kytölä, V.; Nykter, M.; Kiviranta, R. Comparative analysis of osteoblast gene expression profiles and Runx2 genomic occupancy of mouse and human osteoblasts in vitro. Gene 2017, 626, 119–131. [Google Scholar] [CrossRef]
- Chen, X.; Li, M.; Yan, J.; Liu, T.; Pan, G.; Yang, H.; Pei, M.; He, F. Alcohol Induces Cellular Senescence and Impairs Osteogenic Potential in Bone Marrow-Derived Mesenchymal Stem Cells. Alcohol Alcohol. 2017, 52, 289–297. [Google Scholar] [CrossRef] [Green Version]
- Marini, F.; Cianferotti, L.; Brandi, M. Epigenetic Mechanisms in Bone Biology and Osteoporosis: Can They Drive Therapeutic Choices? Int. J. Mol. Sci. 2016, 17, 1329. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Gao, J.; Liang, J.; Dai, W.; Wang, Z.; Xia, M.; Chen, T.; Huang, S.; Na, J.; Xu, L.; et al. HDAC6 inactivates Runx2 promoter to block osteogenesis of bone marrow stromal cells in age-related bone loss of mice. Stem Cell Res. Ther. 2021, 12, 484. [Google Scholar] [CrossRef]
- Van Amerongen, R. Alternative Wnt pathways and receptors. Cold Spring Harb. Perspect. Biol. 2012, 4, a007914. [Google Scholar] [CrossRef] [Green Version]
- Rauch, A.; Haakonsson, A.K.; Madsen, J.G.S.; Larsen, M.; Forss, I.; Madsen, M.R.; Van Hauwaert, E.L.; Wiwie, C.; Jespersen, N.Z.; Tencerova, M.; et al. Osteogenesis depends on commissioning of a network of stem cell transcription factors that act as repressors of adipogenesis. Nat. Genet. 2019, 51, 716–727. [Google Scholar] [CrossRef]
- Bueno, C.; Martínez-Morga, M.; Martínez, S. Non-proliferative neurogenesis in human periodontal ligament stem cells. Sci. Rep. 2019, 9, 18038. [Google Scholar] [CrossRef] [Green Version]
- Zaret, K.S. Pioneer Transcription Factors Initiating Gene Network Changes. Annu. Rev. Genet. 2020, 54, 367–385. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chu, A.; Chakroun, I.; Islam, U.; Blais, A. Cooperation between myogenic regulatory factors and SIX family transcription factors is important for myoblast differentiation. Nucleic Acids Res. 2010, 38, 6857–6871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francetic, T.; Li, Q. Skeletal myogenesis and Myf5 activation. Transcription 2011, 2, 109–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, C.; Pan, T.; Zhang, N.; Guo, J.; Yang, B.; Zhang, D.; Li, J.; Xu, K.; Meng, Z.; He, H. Sp1 promotes dental pulp stem cell osteoblastic differentiation through regulating noggin. Mol. Cell. Probes 2020, 50, 101504. [Google Scholar] [CrossRef] [PubMed]
- Bozec, A.; Bakiri, L.; Jimenez, M.; Schinke, T.; Amling, M.; Wagner, E.F. Fra-2/AP-1 controls bone formation by regulating osteoblast differentiation and collagen production. J. Cell Biol. 2010, 190, 1093–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tai, P.W.L.; Wu, H.; van Wijnen, A.J.; Stein, G.S.; Stein, J.L.; Lian, J.B. Genome-wide DNase hypersensitivity, and occupancy of RUNX2 and CTCF reveal a highly dynamic gene regulome during MC3T3 pre-osteoblast differentiation. PLoS ONE 2017, 12, e0188056. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Assis, R.I.F.; Racca, F.; Ferreira, R.S.; Ruiz, K.G.S.; da Silva, R.A.; Clokie, S.J.H.; Wiench, M.; Andia, D.C. Osteogenic Commitment of Human Periodontal Ligament Cells Is Predetermined by Methylation, Chromatin Accessibility and Expression of Key Transcription Factors. Cells 2022, 11, 1126. https://doi.org/10.3390/cells11071126
Assis RIF, Racca F, Ferreira RS, Ruiz KGS, da Silva RA, Clokie SJH, Wiench M, Andia DC. Osteogenic Commitment of Human Periodontal Ligament Cells Is Predetermined by Methylation, Chromatin Accessibility and Expression of Key Transcription Factors. Cells. 2022; 11(7):1126. https://doi.org/10.3390/cells11071126
Chicago/Turabian StyleAssis, Rahyza I. F., Francesca Racca, Rogério S. Ferreira, Karina G. S. Ruiz, Rodrigo A. da Silva, Samuel J. H. Clokie, Malgorzata Wiench, and Denise C. Andia. 2022. "Osteogenic Commitment of Human Periodontal Ligament Cells Is Predetermined by Methylation, Chromatin Accessibility and Expression of Key Transcription Factors" Cells 11, no. 7: 1126. https://doi.org/10.3390/cells11071126
APA StyleAssis, R. I. F., Racca, F., Ferreira, R. S., Ruiz, K. G. S., da Silva, R. A., Clokie, S. J. H., Wiench, M., & Andia, D. C. (2022). Osteogenic Commitment of Human Periodontal Ligament Cells Is Predetermined by Methylation, Chromatin Accessibility and Expression of Key Transcription Factors. Cells, 11(7), 1126. https://doi.org/10.3390/cells11071126