
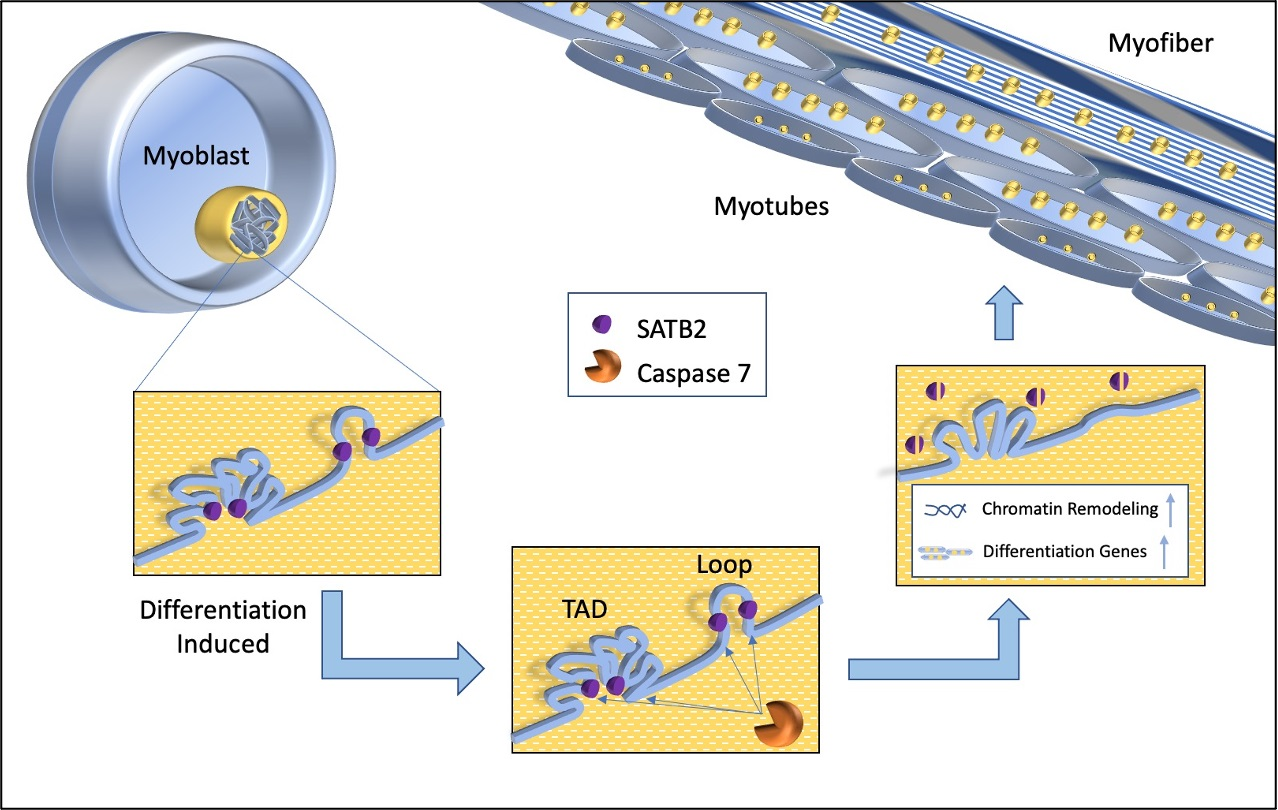
Chromatin Reorganization during Myoblast Differentiation Involves the Caspase-Dependent Removal of SATB2

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Mice and In Vivo Procedures
2.2. Cell Culture
2.3. Protein Extraction and Western Blotting
2.4. Immunofluorescence
2.5. Immunohistochemistry
2.6. Chromatin Immunoprecipitation (ChIP) Assay
2.7. ChIP-Sequencing and Bioinformatics
2.8. RNA-Sequencing and Bioinformatics
2.9. Chromosome Conformation Capture, Hi-C
2.10. Hi-C Data Processing
2.11. Caspase Cleavage Assays
2.12. Caspase Inhibition Assays
2.13. siRNA-Mediated Depletion of SATB2 and Caspase 7 Gene Expression
2.14. Statistical Analysis
3. Results
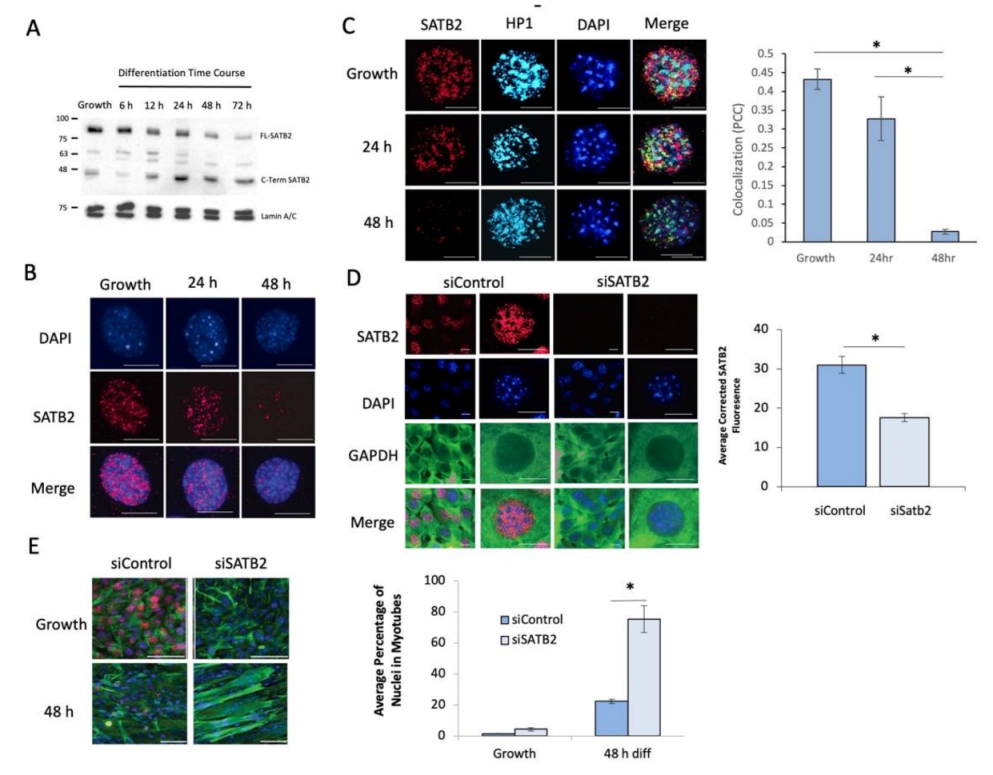
3.1. SATB2 Restrains the Induction of Muscle Cell Differentiation
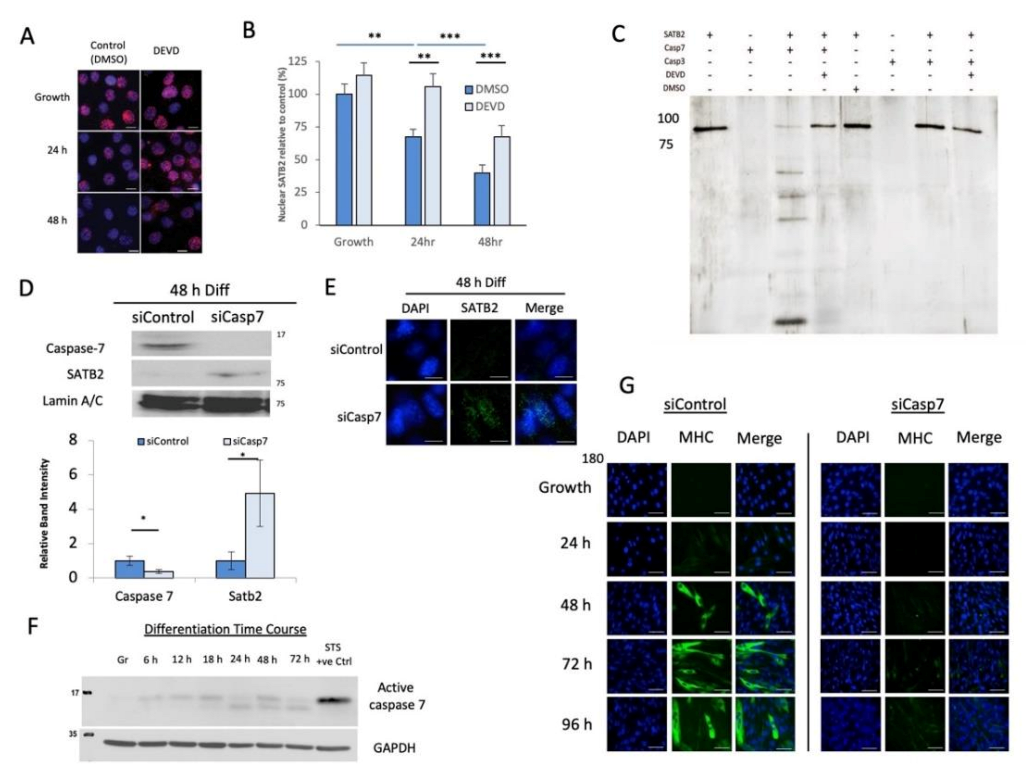
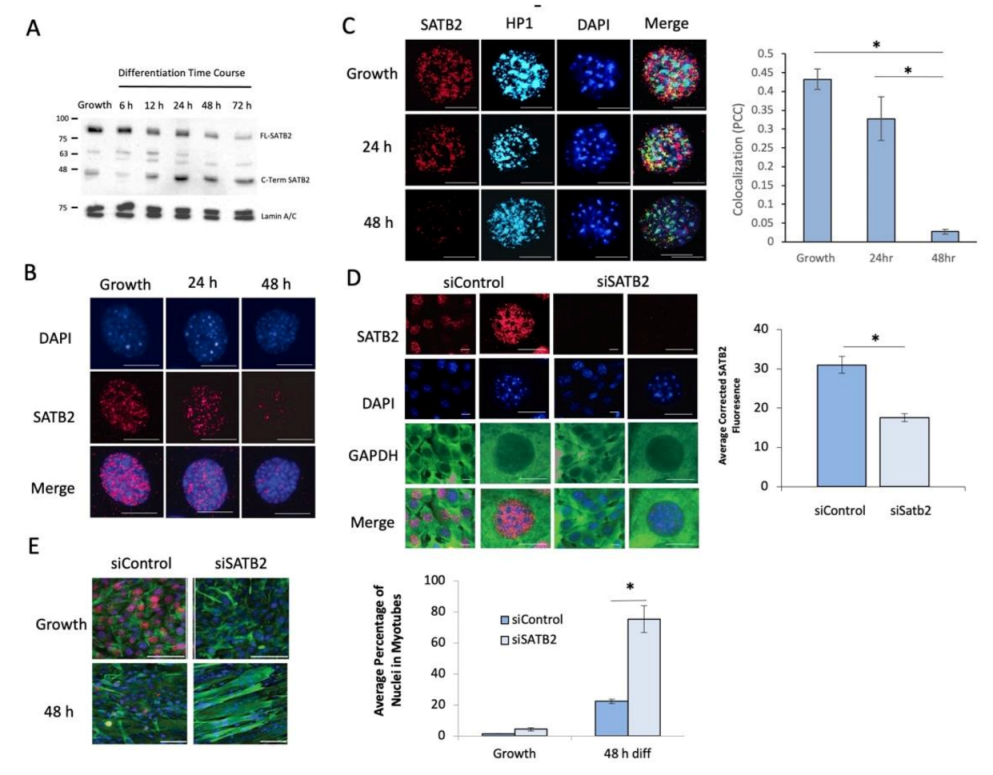
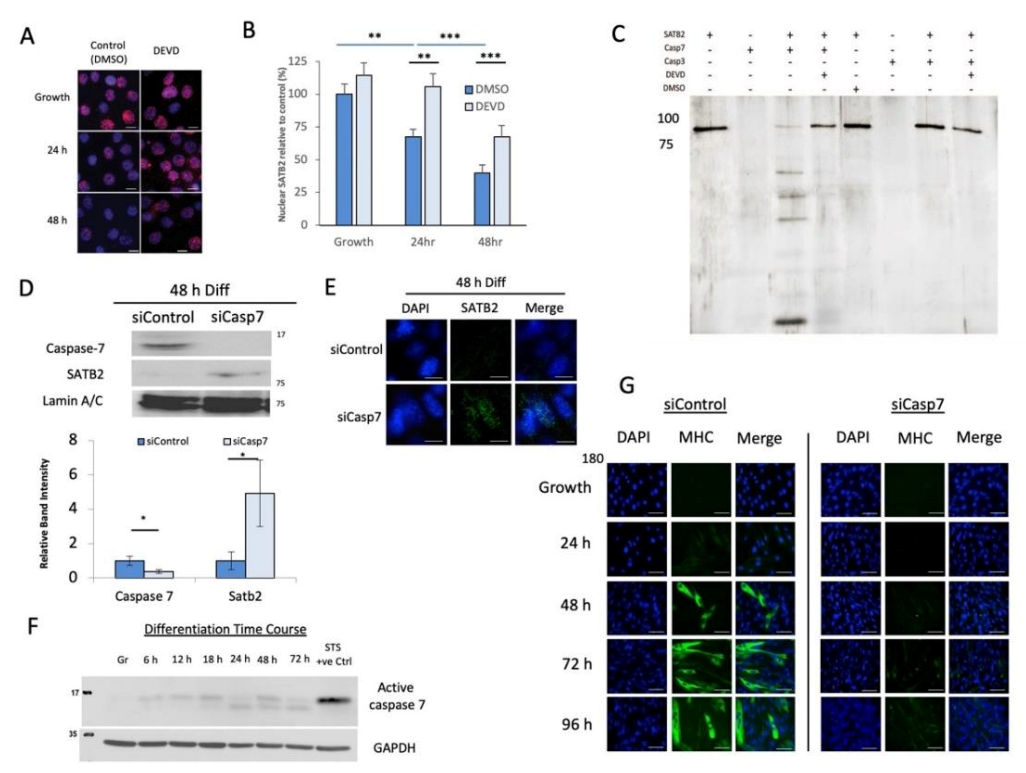
3.2. Caspase 7 Cleaves SATB2 during Early Myogenesis
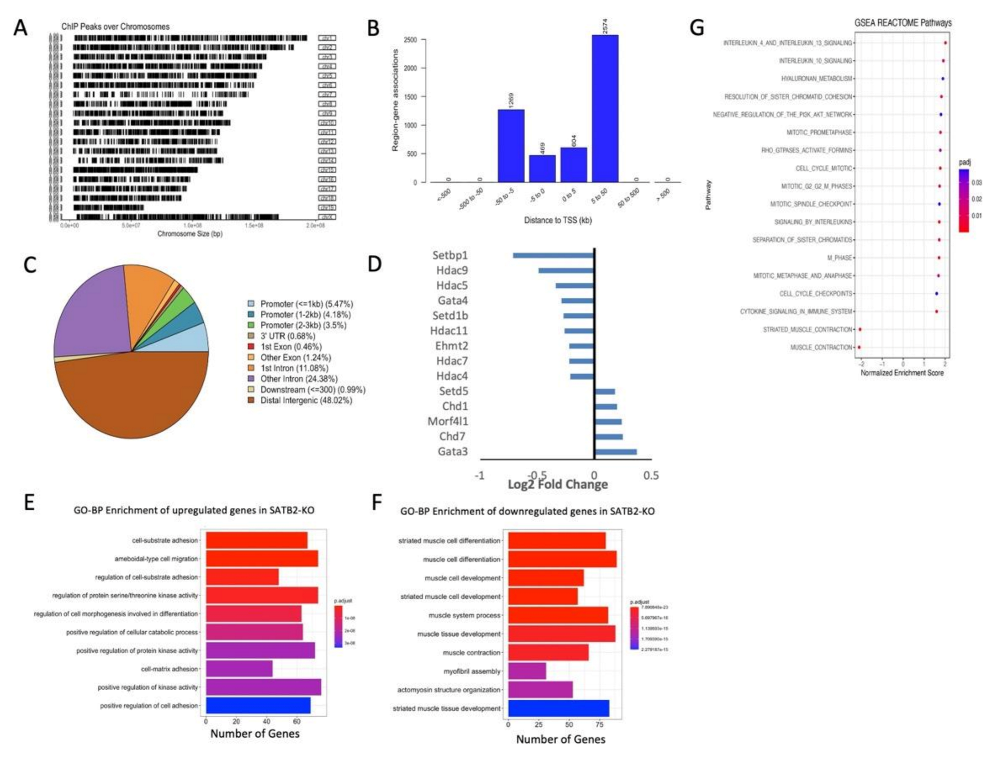
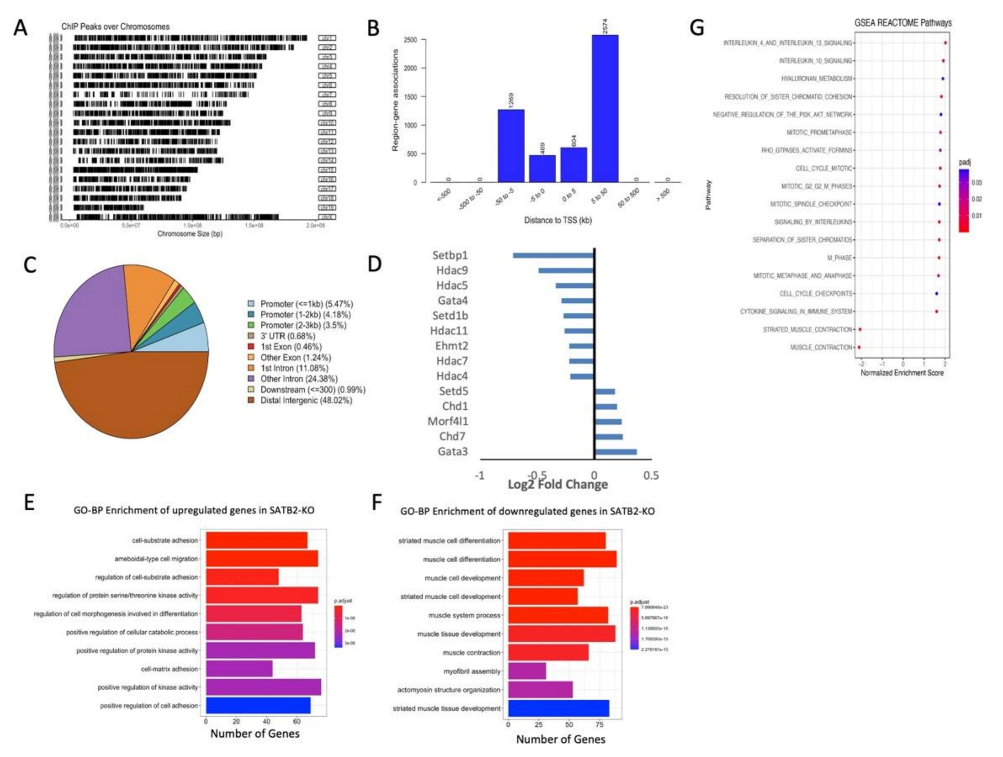
3.3. ChIP- and RNA-seq Data Support the Role of SATB2 in Regulating Muscle Satellite Cell Differentiation and Chromatin Reorganization
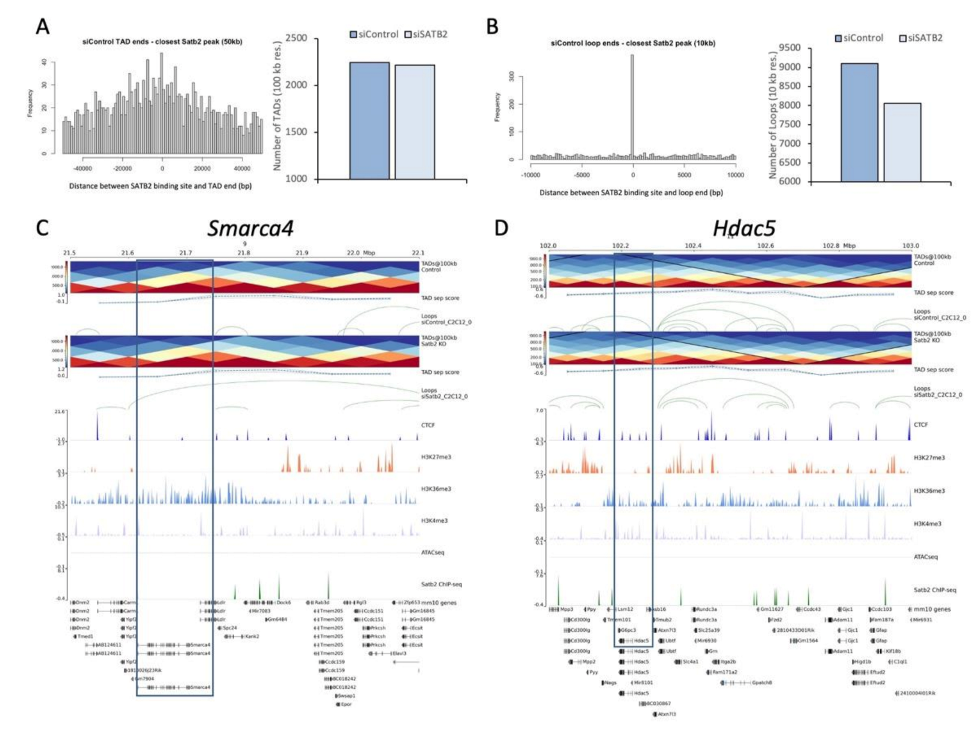
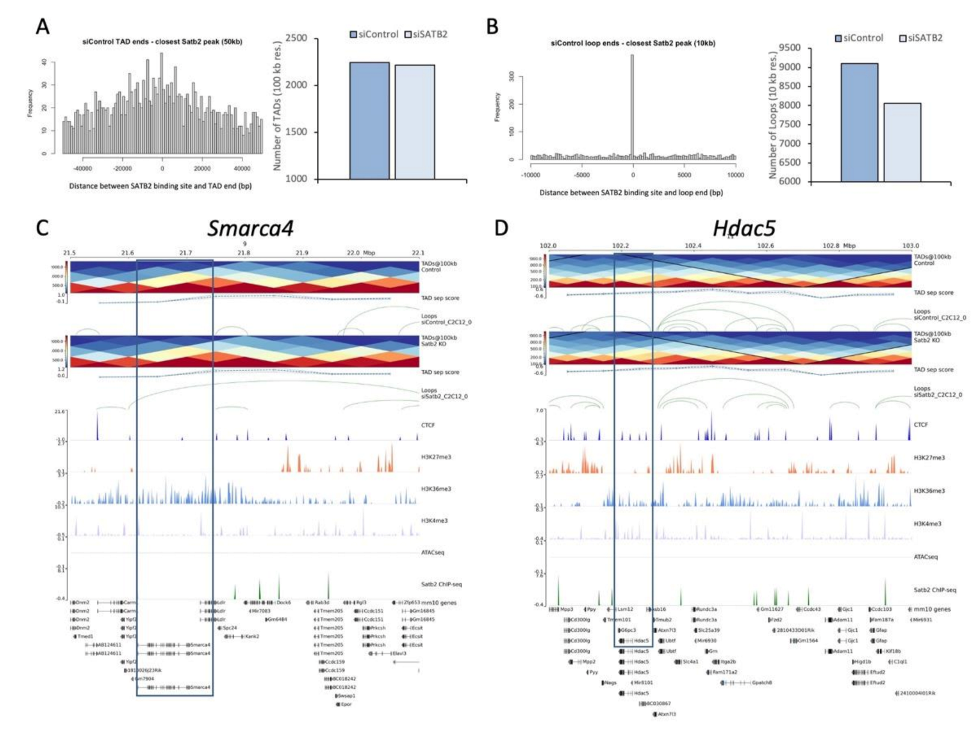
3.4. SATB2 Regulates Chromatin Organization around Genes Related to Myogenesis
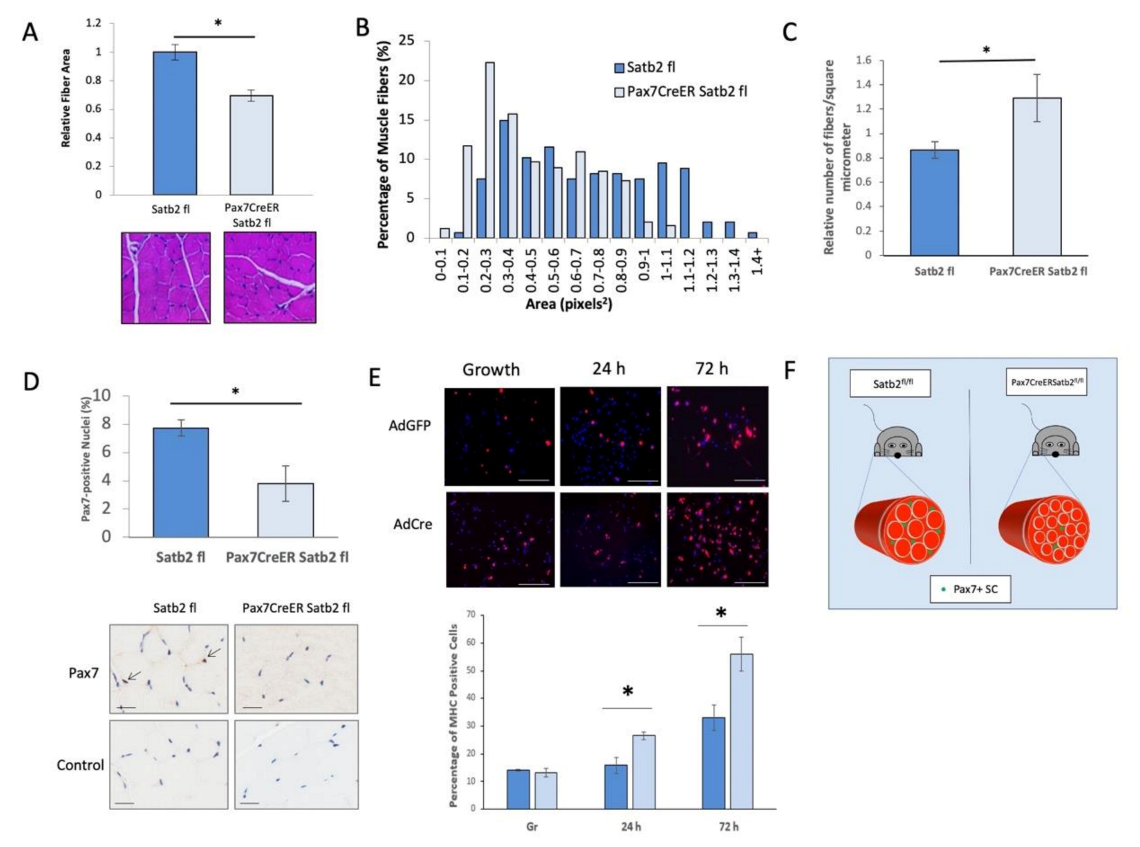
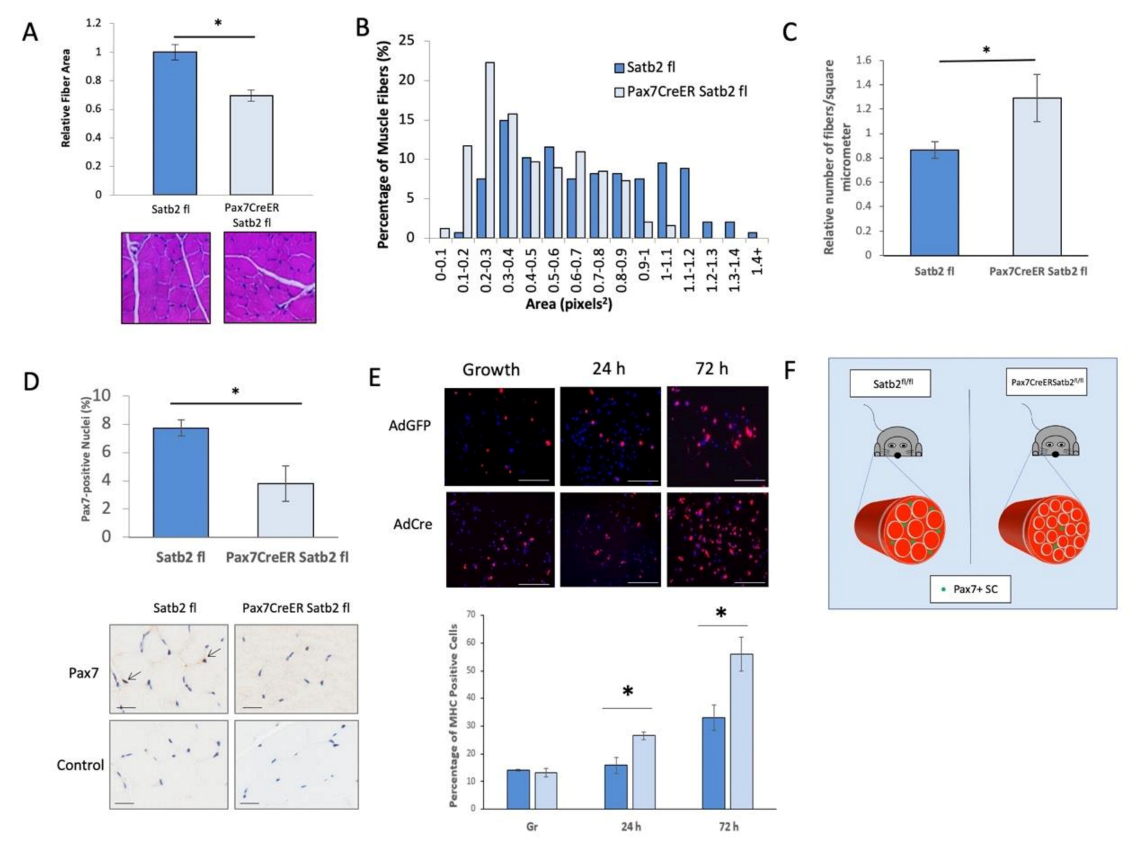
3.5. In Vivo Depletion of SATB2 in Muscle Satellite Cells Decreases Muscle Fiber Area and the Number of Pax7-Expressing Satellite Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Müller, C.; Leutz, A. Chromatin remodeling in development and differentiation. Curr. Opin. Genet. Dev. 2001, 11, 167–174. [Google Scholar] [CrossRef]
- Forcales, S.V.; Albini, S.; Giordani, L.; Malecova, B.; Cignolo, L.; Chemov, A.; Coultinho, P.; Saccone, V.; Consalvi, S.; Williams, R.; et al. Signal-dependent incorporation of MyoD-BAF60c into Brg1-based SWI/SNF chromatin-remodeling complex. EMBO J. 2012, 31, 301–316. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Díaz, E.; Corces, V.G. Architectural proteins: Regulators of 3D genome organization in cell fate. Trends Cell Biol. 2014, 24, 703–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hota, S.K.; Bruneau, B.G. ATP-dependent chromatin remodeling during mammalian development. Development 2016, 143, 2882–2897. [Google Scholar] [CrossRef] [Green Version]
- Doynova, M.D.; Markworth, J.F.; Cameron-Smith, D.; Vickers, M.H.; O’Sullivan, J.M. Linkages between changes in the 3D organization of the genome and transcription during myotube differentiation in vitro. Skelet. Muscle 2017, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Robinson, D.C.L.; Dilworth, J.F. Epigenetic regulation of adult myogenesis. Curr. Top. Dev. Biol. 2018, 126, 235–284. [Google Scholar]
- Bushey, A.M.; Dorman, E.R.; Corces, V.G. Chromatin insulators: Regulatory mechanisms and epigenetic inheritance. Mol. Cell 2008, 32, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Arope, S.; Harraghy, N.; Pjanic, M.; Mermod, N. Molecular characterization of a human matrix attachment region epigenetic regulator. PLoS ONE 2013, 8, e79262. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Hernández, J.M.; Mallappa, C.; Nasipak, B.T.; Oesterreich, S.; Imbalzano, A.N. The scaffold attachment factor b1 (Safb1) regulates myogenic differentiation by facilitating the transition of myogenic gene chromatin from a repressed to an activated state. Nucleic Acid Res. 2013, 41, 5704–5716. [Google Scholar] [CrossRef]
- Savarese, F.; Dávila, A.; Nechanitzky, R.; De La Rosa-Velazquez, I.; Pereira, C.F.; Engelke, R.; Takahashi, K.; Jenuwein, T.; Kohwi-Shigematsu, T.; Fisher, A.G.; et al. Satb1 and Satb2 regulate embryonic stem cell differentiation and Nanog expression. Genes Dev. 2009, 23, 2625–2638. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.; Zhang, P.; Fu, Y.; Ge, J.; Cheng, J.; Yuan, H.; Jiang, H. Roles of SATB2 in site-specific stemness, autophagy and senescence of bone marrow mesenchymal stem cells. J. Cell. Physiol. 2015, 230, 680–690. [Google Scholar] [CrossRef]
- Britanova, O.; Akopov, S.; Lukyanov, S.; Gruss, P.; Tarabykin, V. Novel transcription factor SATB2 interacts with matrix attachment region DNA elements in a tissue-specific manner and demonstrates cell-type-dependent expression in the developing mouse CNS. Eur. J. Neurosci. 2005, 21, 658–668. [Google Scholar] [CrossRef]
- Dobreva, G.; Chahrour, M.; Dautzenberg, M.; Chirivella, L.; Kanzler, B.; Fariñas, I.; Karsenty, G.; Grosschedl, R. SATB2 is a multifunctional determinant of craniofacial patterning and osteoblast differentiation. Cell 2006, 125, 971–986. [Google Scholar] [CrossRef] [Green Version]
- Han, H.J.; Russo, J.; Kohwi, Y.; Kohwi-Shigematsu, T. SATB1 reprogrammes gene expression to promote breast tumour growth and metastasis. Nature 2008, 452, 187–193. [Google Scholar] [CrossRef]
- Leone, D.P.; Heavner, W.E.; Ferenczi, E.A.; Dobreva, G.; Huguendard, J.R.; Grosschedl, R.; McConnell, S.K. Satb2 regulates the differentiation of both callosal and subcerebral projection neurons in the developing cerebral cortex. Cereb. Cortex 2015, 25, 3406–3419. [Google Scholar] [CrossRef] [Green Version]
- Urrutia, G.A.; Ramachandran, H.; Cauchy, P.; Boo, K.; Ramamoorthy, S.; Boller, S.; Dogan, E.; Clapes, T.; Trompouki, E.; Torres-Padilla, M.-E.; et al. ZFP451-mediated SUMOylation of SATB2 drives embryonic stem cell differentiation. Genes Dev. 2021, 35, 1142–1160. [Google Scholar] [CrossRef]
- Feurle, P.; Abentung, A.; Cera, I.; Wahl, N.; Ablinger, C.; Bucher, M.; Stefan, E.; Sprenger, S.; Teis, D.; Fischer, A.; et al. SATB2-LEMD2 interaction links nuclear shape plasticity to regulation of cognition-related genes. EMBO J. 2021, 40, e103701. [Google Scholar] [CrossRef]
- Bengani, H.; Handley, M.; Alvi, M.; Ibitoye, R.; Lees, M.; Lynch, S.A.; Lam, W.; Fannemel, M.; Nordgren, A.; Malmgren, H.; et al. Clinical and molecular consequences of disease-associated de novo mutations in SATB2. Nature 2017, 19, 900–908. [Google Scholar] [CrossRef] [Green Version]
- Galande, S.; Dickinson, L.A.; Milan, I.S.; Sikorska, M.; Kohwi-Shigematsu, T. SATB1 cleavage by caspase 6 disrupts PDZ domain-mediated dimerization, causing detachment from chromatin early in T-cell apoptosis. Mol. Cell Biol. 2001, 21, 5591–5604. [Google Scholar] [CrossRef] [Green Version]
- Unsain, N.; Barker, P.A. New Views on the Misconstrued: Executioner caspases and their diverse non-apoptotic roles. Neuron 2015, 88, 461–474. [Google Scholar] [CrossRef] [Green Version]
- Bell, R.A.V.; Megeney, L.A. Evolution of caspase-mediated cell death and differentiation: Twins separated at birth. Cell Death Diff. 2017, 24, 1359–1368. [Google Scholar] [CrossRef]
- Fernando, P.; Kelly, J.F.; Balazsi, K.; Slack, R.S.; Megeney, L.A. Caspase 3 activity is required for skeletal muscle differentiation. Proc. Natl. Acad. Sci. USA 2002, 99, 11025–11030. [Google Scholar] [CrossRef] [Green Version]
- Murray, T.V.A.; McMahon, J.M.; Howley, B.A.; Stanley, A.; Ritter, T.; Mohr, A.; Zwacka, R.; Fearnhead, H.O. A non-apoptotic role for caspase-9 in muscle differentiation. J. Cell Sci. 2008, 121, 3786–3793. [Google Scholar] [CrossRef] [Green Version]
- Larsen, B.D.; Rampalli, S.; Burns, L.E.; Brunette, S.; Dilworth, F.J.; Megeney, L.A. Caspase 3/caspase-activated DNase promote cell differentiation by inducing DNA strand breaks. Proc. Natl. Acad. Sci. USA 2010, 107, 4230–4235. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.-G.; He, Q.-Y. ChIPseeker: An R/Bioconductor Package for Chip Peak Annotation, Comparison and Visualization. Bioinformatics 2015, 31, 2382–2383. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Heinz, S.; Benner, C.; Spann, N.; Bertolino, E.; Lin, Y.C.; Laslo, P.; Cheng, J.X.; Murre, C.; Singh, H.; Glass, C.K. Simple Combinations of Lineage-Determining Transcription Factors Prime cis-Regulatory Elements Required for Macrophage and B Cell Identities. Mol. Cell 2010, 38, 576–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimental, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, A.; Ibrahim, J.G.; Love, M.I. Heavy-tailed prior distributions for sequence count data: Removing the noise and preserving large differences. Bioinformatics 2019, 35, 2084–2092. [Google Scholar] [CrossRef]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucl. Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [Green Version]
- Korotkevich, G.; Sukhov, V.; Budin, N.; Shipak, B.; Artyomov, M.N.; Sergushichev, A. Fast gene set enrichment analysis. bioRxiv 2016. [Google Scholar] [CrossRef] [Green Version]
- Durand, N.C.; Shamim, M.S.; Machol, I.; Rao, S.S.P.; Huntley, M.H.; Lander, E.S.; Aiden, E.L. Juicer provides a one-click system for analyzing loop-resolution Hi-C experiments. Cell Syst. 2016, 3, 95–98. [Google Scholar] [CrossRef] [Green Version]
- Asp, P.; Blum, R.; Vethantham, V.; Parisi, F.; Micsinai, M.; Cheng, J.; Bowman, C.; Kluger, Y.; Dynlacht, D. Genome-wide remodeling of the epigenetic landscape during myogenic differentiation. Proc. Natl. Acad. Sci. USA 2011, 108, E149–E158. [Google Scholar] [CrossRef] [Green Version]
- Dick, S.A.; Chang, N.C.; Dumont, N.A.; Bell, R.A.V.; Putinski, C.; Kawabe, Y.; Litchfield, D.W.; Rudnicki, M.A.; Megeney, L.A. Caspase 3 cleavage of Pax7 inhibits self-renewal of satellite cells. Proc. Natl. Acad. Sci. USA 2015, 112, E5246–E5252. [Google Scholar] [CrossRef] [Green Version]
- Al-Khalaf, M.H.; Blake, L.E.; Larsen, B.D.; Bell, R.A.V.; Brunette, S.; Parks, R.J.; Rudnicki, M.A.; McKinnon, P.J.; Dilworth, F.J.; Megeney, L.A. Temporal activation of XRCC1-mediated DNA repair is essential for muscle differentiation. Cell Discov. 2016, 12, 15041. [Google Scholar] [CrossRef] [Green Version]
- Slaymaker, I.M.; Gao, L.; Zetsche, B.; Scott, D.A.; Yan, W.X.; Zhang, F. Rationally engineered Cas9 nucleases with improved specificity. Science 2016, 351, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Wang, T.; Su, Y.; Yin, Y.; Xu, S.; Ma, C.; Han, X. The behavior of SATB1, a MAR-binding protein, in response to apoptosis stimulation. Cell Biol. Int. 2006, 30, 244–247. [Google Scholar] [CrossRef]
- Dudek, A.H.; Pfaff, F.; Bolte, H.; Waguia Kontchou, C.; Schwemmie, M. Partial inactivation of chromatin remodelers SMARCA2 and SMARCA4 in virus-infected cells by caspase-mediated cleavage. J. Virol. 2018, 92, e00343-18. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Chen, Y.; Kim, B.; Wang, H.; Zhao, C.; He, X.; Liu, L.; Liu, W.; Wu, L.M.N.; Mao, M.; et al. Olig2 targets chromatin remodelers to enhancers to initiate oligodendrocyte differentiation. Cell 2013, 152, 248–261. [Google Scholar] [CrossRef] [Green Version]
- Griffin, C.T.; Brennan, J.; Magnuson, T. The chromatin-remodeling enzyme BRG1 plays an essential role in primitive erythropoiesis and vascular development. Development 2008, 135, 493–500. [Google Scholar] [CrossRef] [Green Version]
- Albini, S.; Toto, P.C.; Dall’Agnese, A.; Malecova, B.; Cenciarelli, C.; Felsani, A.; Caruso, M.; Bultman, S.J.; Puri, P.L. Brahma is required for cell cycle arrestand late muscle gene expression during skeletal myogenesis. EMBO Rep. 2015, 16, 1037–1050. [Google Scholar] [CrossRef]
- Oda, Y.; Chalkley, R.J.; Burlingame, A.L.; Bikle, D.D. The transcriptional coactivator DRIP/mediator complex is involved in vitamin D receptor function and regulates keritinocyte differentiation. J. Investig. Dermatol. 2010, 130, 2377–2388. [Google Scholar] [CrossRef] [Green Version]
- Sen, R.; Pezoa, S.A.; Shull, L.C.; Hernandez-Lagunas, L.; Niswander, L.A.; Artinger, K.B. Kat2a and Kat2b acetyltransferase activity regulates craniofacial cartilage and bone differentiation in zebrafish and mice. J. Dev. Biol. 2018, 6, 27. [Google Scholar] [CrossRef] [Green Version]
- Hromadkova, L.; Bezdekova, D.; Pala, J.; Schedin-Weiss, S.; Tjernberg, L.O.; Hoschi, C.; Ovespian, S.V. Brain-derived neurotrophic factor (BDNF) promotes molecular polarization and differentiation of immature neuroblastoma cells into definitive neurons. Biochim. Biophys. Acta 2020, 1867, 118737. [Google Scholar] [CrossRef]
- McKinsey, T.A.; Zhang, C.-L.; Lu, J.; Olson, E.N. Signal-dependent nuclear export of a histone deactylase regulates muscle differentiation. Nature 2000, 408, 106–111. [Google Scholar] [CrossRef]
- Wang, L.; Li, W.; Li, K.; Guo, Y.; Liu, D.; Yao, Z.; Lin, X.; Li, S.; Jiang, Z.; Liu, Q.; et al. The oncogenic roles of nuclear receptor coactivator 1 in human esophageal carcinoma. Cancer Med. 2018, 7, 5205–5216. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, Y.; You, J.; Zhang, H.; Zhang, X.; Ye, L. Myosin light-chain kinase contributes to the proliferation and migration of breast cancer cells through cross-talk with activated ERK1/2. Cancer Lett. 2008, 270, 312–327. [Google Scholar] [CrossRef]
- Horsely, V.; Jansen, K.M.; Mills, S.T.; Paviath, G.K. IL-4 acts as a myoblast recruitment factor dueing mammalian muscle growth. Cell 2003, 113, 483–494. [Google Scholar] [CrossRef] [Green Version]
- Poveda-Reyes, S.; Moulisova, V.; Sanmartin-Masia, E.; Quintanilla-Sierra, L.; Salmeron-Sanchez, M.; Ferrer, G.G. Gelatin-hyaluronic acid hydrogels with tuned stiffness counterbalance cellular forces and promote cell differentiation. Macromol. Biosci. 2016, 16, 1311–1324. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, S.D.; Narumiya, S.; Dhawan, J. The RhoA effector mDiaphanous regulates MyoD expression and cell cycle progression via SRF-dependent and SRF-independent pathways. J. Cell Sci. 2007, 120, 3086–3098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishijo, K.; Hosoyama, T.; Bjornson, C.R.R.; Schaffer, B.S.; Prajapati, S.I.; Bahadur, A.N.; Hansen, M.S.; Blandford, M.C.; McGleish, A.T.; Rubin, B.P.; et al. Biomarker system for studying muscle, stem cells, and cancer in vivo. FASEB J. 2009, 23, 2681–2690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seenundun, S.; Rampalli, S.; Liu, Q.C.; Aziz, A.; Palii, C.; Hong, S.; Blais, A.; Brand, M.; Ge, K.; Dilworth, F.J. UTX mediates demethylation of H3K27me3 at muscle-specific genes during myogenesis. EMBO J. 2010, 29, 1401–1411. [Google Scholar] [CrossRef]
- Faralli, H.; Wang, C.; Nakka, K.; Benyoucef, A.; Sebastian, S.; Zhuang, L.; Chu, A.; Palii, C.G.; Liu, C.; Camellato, B.; et al. UTX demethylase activity is required for satellite cell-mediated muscle regeneration. J. Clin. Investig. 2016, 126, 1555–1565. [Google Scholar] [CrossRef] [Green Version]
- Jaitner, C.; Reddy, C.; Abentung, A.; Whittle, N.; Rieder, D.; Delekate, A.; Korte, M.; Jain, G.; Fischer, A.; Sananbenesi, F.; et al. Satb2 determines miRNA expression and long-term memory in the adult central nervous system. eLife 2016, 5, e17361. [Google Scholar] [CrossRef]
- Zhao, X.; Qu, Z.; Tickner, J.; Xu, J.; Dai, K.; Zhang, X. The role of SATB2 in skeletogenesis and human disease. Cytokine Gr. Factor Rev. 2014, 25, 35–44. [Google Scholar] [CrossRef]
- de la Serna, I.L.; Ohkawa, Y.; Imbalzano, A.N. Chromatin remodeling in mammalian differentiation: Lessons from ATP-dependent remodelers. Nat. Rev. Genet. 2006, 7, 461–473. [Google Scholar] [CrossRef]
- Keenen, B.; de la Serna, I.L. Chromatin remodeling in embryonic stem cells: Regulating the balance between pluripotency and differentiation. J. Cell. Physiol. 2008, 219, 1–7. [Google Scholar] [CrossRef]
- Fisher, C.L.; Fisher, A.G. Chromatin states in pluripotent, differentiated, and reprogrammed cells. Curr. Opin. Genet. Dev. 2011, 21, 140–146. [Google Scholar] [CrossRef]
- Chen, T.; Dent, S.Y.R. Chromatin modifiers: Regulators of cellular differentiation. Nat. Rev. Genet. 2014, 15, 93–106. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Li, M.; Gu, L.; Chen, X.; Shi, J.; Zhang, X.; Jiang, C. Chromatin remodeling during in vivo neural stem cells differentiating to neurons in early Drosophila embryos. Cell Death Diff. 2017, 24, 409–420. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, S.M.; Kohwi-Shigematsu, T.; Skalnik, D.G. The matrix attachment region-binding protein SATB1 interacts with multiple elements within the gp91phox promoter and is down-regulated during myeloid differentiation. J. Biol. Chem. 2001, 276, 44472–44480. [Google Scholar] [CrossRef] [Green Version]
- Asanoma, K.; Kubota, K.; Chakraborty, D.; Renaud, S.J.; Wake, N.; Fukushima, K.; Soares, M.J.; Rumi, M.A. SATB homeobox proteins regulate trophoblast stem cell renewal and differentiation. J. Biol. Chem. 2012, 287, 2257–2268. [Google Scholar] [CrossRef] [Green Version]
- Addison, W.N.; Hall, K.C.; Kokabu, S.; Matsubara, T.; Fu, M.M.; Gori, F.; Baron, R. Zfp423 Regulates Skeletal Muscle Regeneration and Proliferation. Mol. Cell. Biol. 2019, 39, e00447-18. [Google Scholar] [CrossRef] [Green Version]
- Matalova, E.; Lesot, H.; Svandova, E.; Vanden Berghe, T.; Sharpe, P.T.; Healy, C.; Vandenabeele, P.; Tucker, A.S. Caspase-7 participates in differentiation of cells forming dental hard tissues. Dev. Growth Differ. 2013, 55, 615–621. [Google Scholar] [CrossRef]
- Svandova, E.; Vesela, B.; Tucker, A.S.; Matalova, E. Activation of pro-apoptotic caspases in non-apoptotic cells during odontogenesis and related osteogenesis. Front. Physiol. 2018, 9, 174. [Google Scholar] [CrossRef] [Green Version]
- Zhivotovsky, B.; Samali, A.; Gahm, A.; Orrenius, S. Caspases: Their intracellular localization and translocation during apoptosis. Cell Death Diff. 1999, 6, 644–651. [Google Scholar] [CrossRef]
- Lee, Y.B.; Colley, S.; Norman, M.; Biamonti, G.; Uney, J.B. SAFB re-distribution marks steps of the apoptotic process. Exp. Cell Res. 2007, 313, 3914–3923. [Google Scholar] [CrossRef]
- Gotzmann, J.; Vicek, S.; Foisner, R. Caspase-mediated cleavage of the chromosome-binding domain of lamina-associated polypeptide 2α. J. Cell Sci. 2000, 113, 3769–3780. [Google Scholar] [CrossRef]
- Narwade, N.; Patel, S.; Alam, A.; Chattopadhyay, S.; Mittal, S.; Kulkarni, A. Mapping of scaffold/matrix attachment regions in human genome: A data mining exercise. Nucleic Acids Res. 2019, 47, 7247–7261. [Google Scholar] [CrossRef] [Green Version]
- Yasui, D.; Miyano, M.; Cai, S.; Varga-Weisz, P.; Kohwi-Shigematsu, T. SATB1 targets chromatin remodeling to regulate genes over long distances. Nature 2002, 419, 641–645. [Google Scholar] [CrossRef]
- Galande, S.; Purbey, P.K.; Notani, D.; Kumar, P. The third dimension of gene regulation: Organization of dynamic chromatin loopscape by SATB. Curr. Opin. Genet. Dev. 2007, 17, 408–414. [Google Scholar] [CrossRef]
- Cai, S.; Lee, C.C.; Kohwi-Shigematsu, T. SATB1 packages densely looped, transcriptionally active chromatin for coordinated expression of cytokine genes. Nat. Genet. 2006, 38, 1278–1288. [Google Scholar] [CrossRef]
- Kumar, P.; Bischof, O.; Purbey, P.K.; Notani, D.; Urlaub, H.; Dejean, A.; Galande, S. Functional interaction between PML and SATB1 regulates chromatin-loop architecture and transcription of the MHC class I locus. Nat. Cell Biol. 2007, 9, 45–56. [Google Scholar]
- Bouwman, B.A.M.; de Laat, W. Getting the genome in shape: The formation of loops, domains and compartments. Genome Biol. 2015, 16, 154. [Google Scholar] [CrossRef] [Green Version]
- Filippova, D.; Patro, R.; Duggal, G.; Kingsford, C. Identification of alternative topological domains in chromatin. Algorithms Mol. Biol. 2014, 9, 14. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; He, B.; Tan, K. Identifying topologically associating domains and subdomains by Gaussian mixture model and proportion test. Nat. Commun. 2017, 8, 535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips-Cremins, J.E.; Sauria, M.E.G.; Sanyal, A.; Gerasimova, T.I.; Lajoie, B.R.; Bell, J.S.K.; Ong, C.-T.; Hookway, T.A.; Guo, C.; Sun, Y.; et al. Architectural protein subclasses shape 3D organization of genomes during lineage commitment. Cell 2013, 153, 1281–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maass, P.G.; Barutcu, A.R.; Weiner, C.L.; Rinn, J.L. Inter-chromosomal contact properties in live-cell imagin and in Hi-C. Mol. Cell 2018, 69, P1039–P1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, K.; Ma, J. Revealing Hi-C subcompartments by imputing inter-chromosomal chromatin interactions. Nat. Commun. 2019, 10, 5069. [Google Scholar] [CrossRef] [Green Version]
- Abbas, A.; He, X.; Niu, J.; Zhou, B.; Zhu, G.; Ma, T.; Song, J.; Gao, J.; Zhang, M.Q.; Zeng, J. Integrating Hi-C and FISH data for modeling of the 3D organization of chromosomes. Nat. Commun. 2019, 10, 2049. [Google Scholar] [CrossRef] [Green Version]
- Oh, H.J.; Aguilar, R.; Kesner, B.; Lee, H.-G.; Kriz, A.J.; Chu, H.-P.; Lee, J.T. Jpx RNA regulates CTCF anchor site selection and formation of chromosome loops. Cell 2021, 184, P6157–P6173. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bell, R.A.V.; Al-Khalaf, M.H.; Brunette, S.; Alsowaida, D.; Chu, A.; Bandukwala, H.; Dechant, G.; Apostolova, G.; Dilworth, F.J.; Megeney, L.A. Chromatin Reorganization during Myoblast Differentiation Involves the Caspase-Dependent Removal of SATB2. Cells 2022, 11, 966. https://doi.org/10.3390/cells11060966
Bell RAV, Al-Khalaf MH, Brunette S, Alsowaida D, Chu A, Bandukwala H, Dechant G, Apostolova G, Dilworth FJ, Megeney LA. Chromatin Reorganization during Myoblast Differentiation Involves the Caspase-Dependent Removal of SATB2. Cells. 2022; 11(6):966. https://doi.org/10.3390/cells11060966
Chicago/Turabian StyleBell, Ryan A. V., Mohammad H. Al-Khalaf, Steve Brunette, Dalal Alsowaida, Alphonse Chu, Hina Bandukwala, Georg Dechant, Galina Apostolova, F. Jeffrey Dilworth, and Lynn A. Megeney. 2022. "Chromatin Reorganization during Myoblast Differentiation Involves the Caspase-Dependent Removal of SATB2" Cells 11, no. 6: 966. https://doi.org/10.3390/cells11060966
APA StyleBell, R. A. V., Al-Khalaf, M. H., Brunette, S., Alsowaida, D., Chu, A., Bandukwala, H., Dechant, G., Apostolova, G., Dilworth, F. J., & Megeney, L. A. (2022). Chromatin Reorganization during Myoblast Differentiation Involves the Caspase-Dependent Removal of SATB2. Cells, 11(6), 966. https://doi.org/10.3390/cells11060966