
Ciliated (FOXJ1+) Cells Display Reduced Ferritin Light Chain in the Airways of Idiopathic Pulmonary Fibrosis Patients

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Samples
2.2. Tissue Dissociation
2.3. Fluorescence-Activated Cell Sorting (FACS)
2.4. Single-Cell RNA Sequencing
2.5. Immunohistochemistry
3. Results
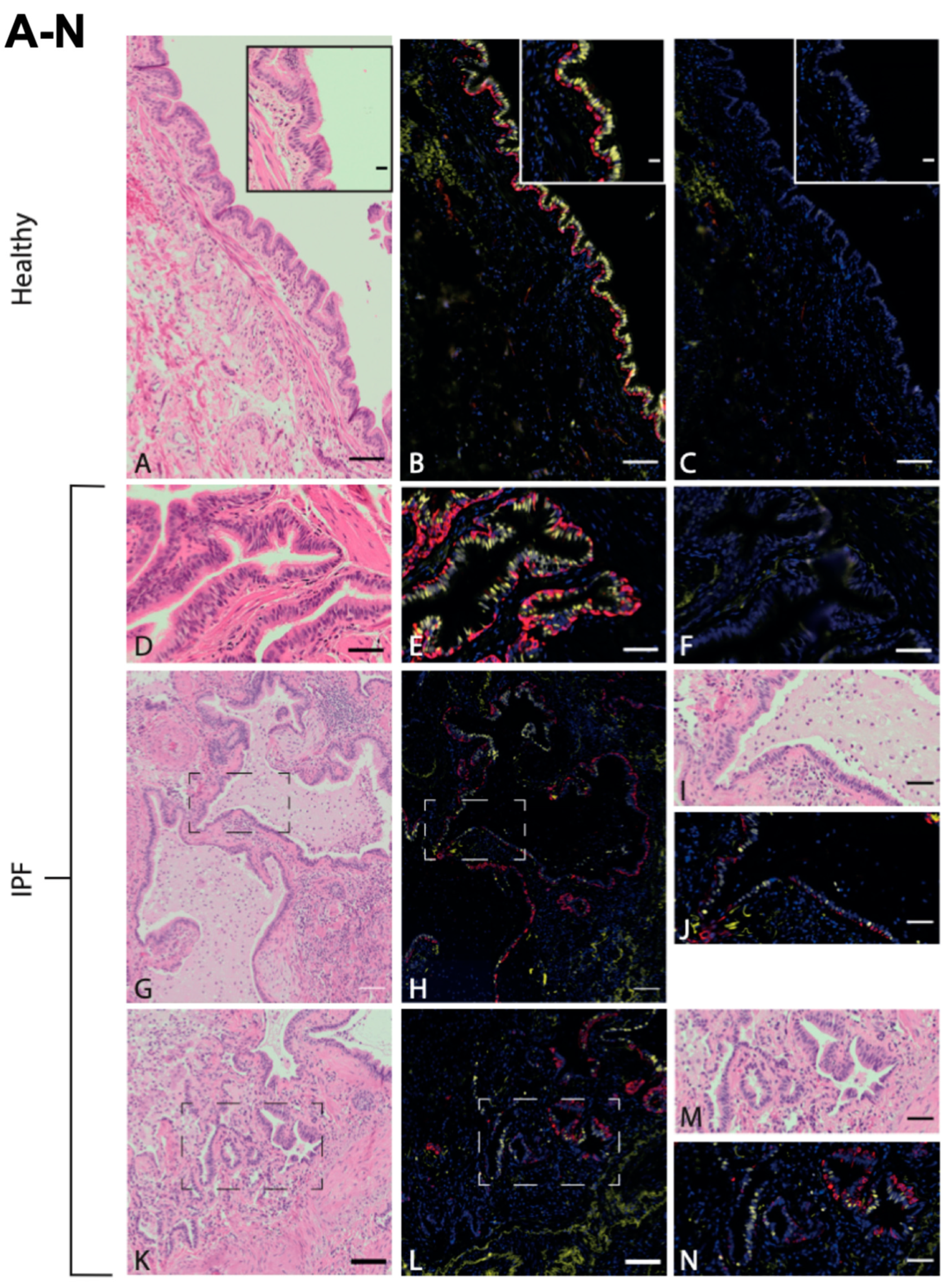
3.1. Immunohistochemistry and Immunofluorescence Show Typical Structures Associated with Disease in Lung Tissue from IPF Patients
3.2. Single-Cell RNA Sequencing Identifies Molecular Differences between Ciliated Cells from IPF Patients and Healthy Control Cells
3.3. Ciliated Cells in IPF Lung Epithelium Exhibit Increased Expression of Ciliated Pathways
3.4. Ciliated Cells in IPF Patients Display Reduced FTL Protein Levels
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ley, B.; Collard, H.R.; King, J.T.E. Clinical Course and Prediction of Survival in Idiopathic Pulmonary Fibrosis. Am. J. Respir. Crit. Care Med. 2011, 183, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Raghu, G.; Weycker, D.; Edelsberg, J.; Bradford, W.Z.; Oster, G. Incidence and prevalence of idiopathic pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 2006, 174, 810–816. [Google Scholar] [CrossRef] [PubMed]
- Wolters, P.J.; Collard, H.R.; Jones, K.D. Pathogenesis of Idiopathic Pulmonary Fibrosis. Annu. Rev. Pathol. 2014, 9, 157–179. [Google Scholar] [CrossRef] [PubMed]
- Raftery, A.E.; Li, N.; Ševčíková, H.; Gerland, P.; Heilig, G.K. Bayesian probabilistic population projections for all countries. Proc. Natl. Acad. Sci. USA 2012, 109, 13915–13921. [Google Scholar] [CrossRef]
- Zuo, W.-L.; Rostami, M.R.; Leblanc, M.; Kaner, R.J.; O’Beirne, S.L.; Mezey, J.G.; Leopold, P.L.; Quast, K.; Visvanathan, S.; Fine, J.S.; et al. Dysregulation of club cell biology in idiopathic pulmonary fibrosis. PLoS ONE 2020, 15, e0237529. [Google Scholar] [CrossRef] [PubMed]
- Seibold, M.A.; Smith, R.W.; Urbanek, C.; Groshong, S.D.; Cosgrove, G.P.; Brown, K.K.; Schwarz, M.I.; Schwartz, D.A.; Reynolds, S.D. The idiopathic pulmonary fibrosis honeycomb cyst contains a mucocilary pseudostratified epithelium. PLoS ONE 2013, 8, e58658. [Google Scholar] [CrossRef]
- Ryu, C.; Homer, R.J.; Herzog, E.L. The Airway in Idiopathic Pulmonary Fibrosis: Protecting the Lung or Promoting Disease? Am. J. Respir. Crit. Care Med. 2016, 193, 1081–1082. [Google Scholar] [CrossRef]
- Bustamante-Marin, X.M.; Ostrowski, L.E. Cilia and Mucociliary Clearance. Cold Spring Harb. Perspect. Biol. 2017, 9, a028241. [Google Scholar] [CrossRef]
- Wiscombe, S.; A Forrest, I.; Simpson, A.J. IPF: Time for the (ciliary) beat generation? Thorax 2013, 68, 1088–1089. [Google Scholar] [CrossRef][Green Version]
- Hogan, B.L.; Barkauskas, C.E.; Chapman, H.A.; Epstein, J.A.; Jain, R.; Hsia, C.C.; Niklason, L.; Calle, E.; Le, A.; Randell, S.H.; et al. Repair and regeneration of the respiratory system: Complexity, plasticity, and mechanisms of lung stem cell function. Cell Stem Cell 2014, 15, 123–138. [Google Scholar] [CrossRef]
- Wijk, S.C.; Prabhala, P.; Michaliková, B.; Sommarin, M.; Doyle, A.; Lang, S.; Kanzenbach, K.; Tufvesson, E.; Lindstedt, S.; Leigh, N.D.; et al. Human Primary Basal Cells Display a Continuum of Molecular Phases from Health to Disease in Chronic Obstructive Pulmonary Disease. Am. J. Respir. Cell Mol. Biol. 2021, 65, 103–113. [Google Scholar] [CrossRef]
- Bankhead, P.; Loughrey, M.B.; Fernández, J.A.; Dombrowski, Y.; McArt, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open source software for digital pathology image analysis. Sci. Rep. 2017, 7, 16878. [Google Scholar] [CrossRef] [PubMed]
- Vidal, R.; Miravalle, L.; Gao, X.; Barbeito, A.G.; Baraibar, M.A.; Hekmatyar, S.K.; Widel, M.; Bansal, N.; Delisle, M.B.; Ghetti, B. Expression of a Mutant Form of the Ferritin Light Chain Gene Induces Neurodegeneration and Iron Overload in Transgenic Mice. J. Neurosci. 2008, 28, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.K.; Kim, R.Y.; Brown, A.C.; Donovan, C.; Vanka, K.S.; Mayall, J.R.; Liu, G.; Pillar, A.L.; Jones-Freeman, B.; Xenaki, D.; et al. Critical role for iron accumulation in the pathogenesis of fibrotic lung disease. J. Pathol. 2020, 251, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Kubo, H. Extracellular Vesicles in Lung Disease. Chest 2018, 153, 210–216. [Google Scholar] [CrossRef]
- Li, J.; Han, Y.; Zhang, H.; Qian, Z.; Jia, W.; Gao, Y.; Zheng, H.; Li, B. The m6A demethylase FTO promotes the growth of lung cancer cells by regulating the m6A level of USP7 mRNA. Biochem. Biophys. Res. Commun. 2019, 512, 479–485. [Google Scholar] [CrossRef]
- Ding, Y.; Qi, N.; Wang, K.; Huang, Y.; Liao, J.; Wang, H.; Tan, A.; Liu, L.; Zhang, Z.; Li, J.; et al. FTO Facilitates Lung Adenocarcinoma Cell Progression by Activating Cell Migration Through mRNA Demethylation. OncoTargets Ther. 2020, 13, 1461–1470. [Google Scholar] [CrossRef]
- Yu, X.; Li, Z.; Zheng, H.; Chan, M.T.V.; Wu, W.K.K. NEAT1: A novel cancer-related long non-coding RNA. Cell Prolif. 2017, 50, e12329. [Google Scholar] [CrossRef]
- Yang, I.V.; Coldren, C.D.; Leach, S.M.; Seibold, M.A.; Murphy, E.; Lin, J.; Rosen, R.; Neidermyer, A.J.; McKean, D.F.; Groshong, S.D.; et al. Expression of cilium-associated genes defines novel molecular subtypes of idiopathic pulmonary fibrosis. Thorax 2013, 68, 1114–1121. [Google Scholar] [CrossRef]
- Ali, M.K.; Kim, R.Y.; Karim, R.; Mayall, J.R.; Martin, K.L.; Shahandeh, A.; Abbasian, F.; Starkey, M.R.; Loustaud-Ratti, V.; Johnstone, D.; et al. Role of iron in the pathogenesis of respiratory disease. Int. J. Biochem. Cell Biol. 2017, 88, 181–195. [Google Scholar] [CrossRef]
- Sammarco, M.C.; Ditch, S.; Banerjee, A.; Grabczyk, E. Ferritin L and H subunits are differentially regulated on a post-transcriptional level. J. Biol. Chem. 2008, 283, 4578–4587. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Grier, D.D.; Woo, J.; Ward, M.; Sui, G.; Torti, S.V.; Torti, F.M.; Beaty, M.W. Ferritin H is a novel marker of early erythroid precursors and macrophages. Histopathology 2013, 62, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Mizuno, T.; Sridharan, A.; Du, Y.; Guo, M.; Tang, J.; Wikenheiser-Brokamp, K.A.; Perl, A.-K.T.; Funari, V.A.; Gokey, J.; et al. Single-cell RNA sequencing identifies diverse roles of epithelial cells in idiopathic pulmonary fibrosis. JCI Insight 2016, 1, e90558. [Google Scholar] [CrossRef] [PubMed]
- Carraro, G.; Mulay, A.; Yao, C.; Mizuno, T.; Konda, B.; Petrov, M.; Lafkas, D.; Arron, J.R.; Hogaboam, C.M.; Chen, P.; et al. Single-cell reconstruction of human basal cell diversity in normal and idiopathic pulmonary fibrosis lungs. Am. J. Respir. Crit. Care Med. 2020, 202, 1540–1550. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.M.; Arosio, P. The ferritins: Molecular properties, iron storage function and cellular regulation. Biochim. Biophys. Acta 1996, 1275, 161–203. [Google Scholar] [CrossRef]
- Cassat, J.E.; Skaar, E.P. Iron in infection and immunity. Cell Host Microbe 2013, 13, 509–519. [Google Scholar] [CrossRef]
- Ogger, P.P.; Byrne, A.J. Lung fibrosis enters the iron age. J. Pathol. 2020, 252, 1–3. [Google Scholar] [CrossRef]
- Yu, G.-H.; Fu, L.; Chen, J.; Wei, F.; Shi, W.-X. Decreased expression of ferritin light chain in osteosarcoma and its correlation with epithelial-mesenchymal transition. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 2580–2587. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GO Term | p Value | Genes |
|---|---|---|
| GO:0006614~SRP-dependent cotranslational protein targeting to membrane | 1.56 × 10−18 | RPL30, RPS8, RPLP1, RPL34, RPS6, RPL11, SRP54, RPS3A, RPL10A, RPS4X, RPS14, RPS15A, RPS19, RPL13, RPL29, RPS27A, RPS11, RPL28, RPL39, RPS24, RPS13, RPL19 |
| GO:0006413~translational initiation | 3.00 × 10−17 | RPL30, RPS8, RPLP1, RPL34, RPS6, RPL11, RPS3A, RPL10A, EIF1, RPS4X, RPS14, RPS15A, RPS19, RPL13, RPL29, RPS27A, RPS11, RPL28, EIF3D, RPL39, RPS24, EIF3A, RPS13, RPL19 |
| GO:0000184~nuclear-transcribed mRNA catabolic process, nonsense-mediated decay | 2.60 × 10−16 | RPL30, RPS8, RPLP1, RPL34, RPS6, RPL11, RPS3A, RPL10A, RPS4X, RPS14, RPS15A, RPS19, MAGOH, RPL13, RPL29, RPS27A, RPS11, RPL28, RPL39, RPS24, RPS13, RPL19 |
| GO:0019083~viral transcription | 1.18 × 10−15 | RPL30, RPS8, RPLP1, RPL34, RPS6, RPL11, RPS3A, RPL10A, RPS4X, RPS14, RPS15A, RPS19, RPL13, RPL29, RPS27A, RPS11, RPL28, RPL39, RPS24, RPS13, RPL19 |
| GO:0006364~rRNA processing | 6.37 × 10−13 | RPL30, RPS8, RPLP1, RPL34, RPS6, RPL11, RPS3A, RPL10A, RPS4X, RPS14, EBNA1BP2, RPS15A, RPS19, RPL13, PIH1D2, RPL29, RPS27A, RPS11, RPL28, RPL39, RPS24, RPP38, RPS13, RPL19 |
| GO:0006412~translation | 3.00 × 10−12 | RPL30, RPL34, RPLP1, RPL11, RRBP1, RPL10A, MRPL41, RPS4X, RPS14, RPS15A, RPS19, RPL13, RPS27A, RPS11, RPL39, RPS13, RPL19, RPS8, RPS6, RPS3A, MRPL23, RPL29, RPL28, SLC25A11, RPS24 |
| GO:0043488~regulation of mRNA stability | 6.20 × 10−6 | YTHDF2, SET, PSMA4, PSMA1, PSMD13, APEX1, PSMD2, UBC, PSMD3, RPS27A, YWHAZ |
| GO:0060071~Wnt signaling pathway, planar cell polarity pathway | 1.17 × 10−4 | PSMA4, PSMA1, PSMD13, PSMD2, UBC, PSMD3, PFN1, RPS27A, AP2M1 |
| GO:0098869~cellular oxidant detoxification | 1.30 × 10−4 | GSTK1, PRDX5, GPX4, GSTP1, GSTA1, CAT, TXN, TXNDC17 |
| GO:0006457~protein folding | 1.63 × 10−4 | CCT3, DNAJA1, FKBP1A, LMAN1, HSP90AA1, ST13, RUVBL2, MLEC, TXN, CCT7, PDIA6, PFDN5 |
| GO:0000398~mRNA splicing, via spliceosome | 2.61 × 10−4 | PRPF40A, LSM7, HNRNPK, SNRPD2, PCBP1, MAGOH, SRSF2, POLR2F, SRSF3, SNRPA1, POLR2H, HNRNPA1, SRSF7 |
| GO:0000302~response to reactive oxygen species | 3.81 × 10−4 | PRDX5, GSTP1, PRDX1, CAT, P4HB, TXN |
| GO:0098609~cell-cell adhesion | 4.62 × 10−4 | DDX3X, ANXA2, RPL34, HDLBP, YWHAZ, CORO1B, MPP7, PCMT1, HNRNPK, PCBP1, PRDX1, PFN1, RPL29, S100A11 |
| GO:0002479~antigen processing and presentation of exogenous peptide antigen via MHC class I, TAP-dependent | 5.15 × 10−4 | PSMA4, PSMA1, PSMD13, PSMD2, PSMD3, HLA-B, B2M |
| GO:0038061~NIK/NF-kappaB signaling | 6.62 × 10−4 | PSMA4, PSMA1, PSMD13, PSMD2, UBC, PSMD3, RPS27A |
| GO:0051436~negative regulation of ubiquitin-protein ligase activity involved in mitotic cell cycle | 9.76 × 10−4 | PSMA4, PSMA1, PSMD13, PSMD2, UBC, PSMD3, RPS27A |
| GO:0006120~mitochondrial electron transport, NADH to ubiquinone | 0.001108056 | NDUFS8, NDUFS7, NDUFA5, NDUFA4, NDUFB11, NDUFS3 |
| GO:0050434~positive regulation of viral transcription | 0.001131143 | POLR2F, RSF1, POLR2H, PFN1, NELFE |
| GO:0090090~negative regulation of canonical Wnt signaling pathway | 0.001306836 | SOX2, PSMA4, PSMA1, PSMD13, PSMD2, UBC, PSMD3, IGFBP2, RPS27A, PFDN5 |
| GO:0051437~positive regulation of ubiquitin-protein ligase activity involved in regulation of mitotic cell cycle transition | 0.001394273 | PSMA4, PSMA1, PSMD13, PSMD2, UBC, PSMD3, RPS27A |
| GO:0043161~proteasome-mediated ubiquitin-dependent protein catabolic process | 0.00169636 | PCNP, PPP2CB, PSMA4, PSMA1, PSMD13, PSMD2, UBC, UBE2D3, PSMD3, FBXO15, RPS27A |
| GO:0031145~anaphase-promoting complex-dependent catabolic process | 0.001703314 | PSMA4, PSMA1, PSMD13, PSMD2, UBC, PSMD3, RPS27A |
| GO:0009060~aerobic respiration | 0.001853146 | CHCHD5, SURF1, PANK2, CAT, UQCRC2 |
| GO:0033209~tumor necrosis factor-mediated signaling pathway | 0.002977448 | PSMA4, PSMA1, PSMD13, PSMD2, UBC, PSMD3, RPS27A, TXNDC17 |
| GO:0090263~positive regulation of canonical Wnt signaling pathway | 0.003270083 | PSMA4, PSMA1, PSMD13, PSMD2, UBC, PSMD3, PIN1, RPS27A |
| GO:1901687~glutathione derivative biosynthetic process | 0.005049175 | GSTK1, GSTP1, GSTA2, GSTA1 |
| GO:0030855~epithelial cell differentiation | 0.005354618 | GSTK1, LGALS3, GSTA2, GSTA1, PGK1, ANXA7 |
| GO:0006283~transcription-coupled nucleotide-excision repair | 0.006767992 | COPS6, UBC, TCEA1, POLR2F, POLR2H, RPS27A |
| GO:0002223~stimulatory C-type lectin receptor signaling pathway | 0.006986379 | PSMA4, PSMA1, PSMD13, PSMD2, UBC, PSMD3, RPS27A |
| GO:0043066~negative regulation of apoptotic process | 0.007011217 | NPM1, ANXA1, DDX3X, GSTP1, RPS6, RPS3A, YWHAZ, DNAJA1, PRDX5, HNRNPK, CAT, UBC, TAX1BP1, RPS27A, SQSTM1, TPT1 |
| GO:0010467~gene expression | 0.007303284 | HNRNPK, PCBP1, POLR2F, POLR2H, HNRNPA1 |
| GO:0016236~macroautophagy | 0.007564389 | MAP1LC3B, MAP1LC3A, UBC, RPS27A, DYNLL1, SQSTM1 |
| GO:0045454~cell redox homeostasis | 0.007986077 | PRDX5, APEX1, PRDX1, P4HB, TXN, PDIA6 |
| GO:0038095~Fc-epsilon receptor signaling pathway | 0.008269841 | PSMA4, PSMA1, PSMD13, PSMD2, UBC, PSMD3, FOS, CALM1, RPS27A |
| GO:0006521~regulation of cellular amino acid metabolic process | 0.009035625 | PSMA4, PSMA1, PSMD13, PSMD2, PSMD3 |
| GO:0006511~ubiquitin-dependent protein catabolic process | 0.009376141 | PSMA4, PSMA1, PSMD13, USP10, ADRM1, UBE2D3, PSMD3, SQSTM1, HERPUD1 |
| GO:0042493~response to drug | 0.010386588 | TGIF1, HSP90AA1, ABCD3, ANXA1, XRCC5, APEX1, IGFBP2, CAT, SRP54, GNAS, FOS, B2M |
| GO:0006405~RNA export from nucleus | 0.011733373 | MAGOH, SRSF2, SRSF3, HNRNPA1, SRSF7 |
| GO:0032480~negative regulation of type I interferon production | 0.012113648 | UBC, TAX1BP1, PIN1, RPS27A |
| GO:0006368~transcription elongation from RNA polymerase II promoter | 0.012537861 | CCNK, ADRM1, TCEA1, POLR2F, POLR2H, NELFE |
| GO:0000462~maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) | 0.01445558 | RPS14, RPS19, RPS8, RPS24 |
| GO Term | p Value | Genes |
|---|---|---|
| GO:0060271~cilium morphogenesis | 4.02 × 10−24 | CEP126, TTC26, IFT172, CEP164, CCDC28B, RPGR, TCTN2, IQUB, DZIP1L, TEKT3, CEP290, BBS5, DYNC2H1, IFT140, DNAAF1, RFX2, TTC21A, RFX3, IFT80, RPGRIP1L, WDR35, KIF27, DNM2, IFT88, CFAP221, TMEM138, WDPCP, CFAP54, INTU, ARL6, DNAH5, TTBK2, STK36, TMEM67, TTC21B, NPHP3, BBS1, GSN, WDR19, IFT122, FOXJ1, FUZ, TMEM231, AHI1, TMEM17, TTLL3, MKS1, CEP162, DZIP1 |
| GO:0042384~cilium assembly | 2.49 × 10−22 | LAMA5, CEP126, TTC26, INTU, ARL6, DNAH5, IFT172, CEP164, CCDC28B, TTBK2, RPGR, STK36, TMEM67, TCTN2, DZIP1L, BBS9, BBS5, CEP290, DYNC2H1, BBS1, DNAI2, IFT140, WDR19, RAB3IP, RFX2, IFT122, RFX3, FOXJ1, FUZ, IFT80, RPGRIP1L, WDR35, KIF27, TMEM231, AHI1, TMEM17, TTLL3, TMEM138, ALMS1, MKS1, CEP162, WDPCP, CFAP54, DZIP1, CLUAP1 |
| GO:0003341~cilium movement | 6.77 × 10−12 | CCDC39, RSPH4A, DNAH11, DNAH1, DNAI2, CFAP73, DNAH7, DNAAF1, DNAH5, CCDC114, CFAP100, CFAP221, HYDIN, DNAI1, CCDC151, CCDC40 |
| GO:0008380~RNA splicing | 4.28 × 10−7 | RBM25, DDX23, AKAP8L, RNPC3, CCAR2, IWS1, SNRNP70, ZNF326, RBM5, SRSF11, RBM10, RBM38, RBM39, MBNL1, ZRANB2, SCAF11, PRPF4B, THOC2, SCAF1, SON, ZNF638, PRPF3, LUC7L3, RBM20, SRSF4, SREK1, CDK12, CLASRP, TARDBP, SUGP2 |
| GO:0060285~cilium-dependent cell motility | 9.68 × 10−7 | DNAH3, CCDC39, DNAH1, DNAH2, DNAH7, DNAAF2, RFX3, CFAP44 |
| GO:0036159~inner dynein arm assembly | 4.50 × 10−6 | CCDC39, CFAP100, DNAH1, CFAP73, DNAH7, DNAAF1, TEKT2, CCDC40 |
| GO:0061512~protein localization to cilium | 5.66 × 10−6 | BBS1, ARL6, IFT140, TTC21B, IFT122, TTC21A, BBS9, CSNK1D, WDR35 |
| GO:0035721~intraciliary retrograde transport | 9.95 × 10−6 | DYNC2H1, IFT140, TTC21B, WDR19, IFT122, TTC21A, WDR35 |
| GO:0035058~nonmotile primary cilium assembly | 2.37 × 10−5 | BBS1, CEP126, TMEM17, C2CD3, CCDC13, MKS1, FUZ, IFT80, CSNK1D, PIBF1 |
| GO:0035082~axoneme assembly | 4.09 × 10−5 | RSPH4A, RP1, CFAP74, LRGUK, SPEF2, TTLL3, CFAP46, CLUAP1 |
| GO:0043484~regulation of RNA splicing | 4.52 × 10−5 | RBM38, CLK1, MBNL1, SON, SNRNP70, CELF1, HNRNPH1, ZNF326, CDK12, CLK4 |
| GO:0006397~mRNA processing | 5.01 × 10−5 | RBM25, CELF1, SRSF1, AKAP8L, CCAR2, IWS1, U2AF2, SNRNP70, ZNF326, SRSF11, RBM10, RBM38, RBM39, MBNL1, ZRANB2, SCAF11, SCAF1, SON, PRPF3, RBM20, SRSF4, SREK1, CDK12, CLASRP, TARDBP, RBM23, SUGP2 |
| GO:0001843~neural tube closure | 7.16 × 10−5 | SDC4, IFT172, SEMA4C, COBL, TSC2, SHROOM3, IFT122, TULP3, FUZ, ARID1A, CELSR1, SPINT1, MKS1, PLXNB2, CLUAP1, PHACTR4 |
| GO:0006468~protein phosphorylation | 7.71 × 10−5 | ULK4, IKBKB, NRBP2, AKAP13, PPP4R1, CDK20, TLK1, NEK3, MAP3K5, STRADA, DAPK1, PRKCD, DYRK1B, CSNK1D, PRPF4B, CSNK1E, PASK, GAK, MAK, SIK3, BIRC6, GAS6, ALPK1, CSNK1G2, CDKL1, CAMK2D, LTK, ATP23, NPR2, PRKCZ, STK3, STK36, PHKG2, ERBB2, STK38, MAP4K4, DMPK, NEK5, CDC42BPG, MOK, BRAF, HIPK1, HIPK3, MAPK10, FER, WNK1, NEK10, AAK1, CDK10, COQ8B, PKN1 |
| GO:0007368~determination of left/right symmetry | 1.20 × 10−4 | DYNC2H1, DNAH11, DNAI2, ARL6, IFT140, DNAH5, FOXJ1, RPGRIP1L, PCSK5, MKS1, NPHP3, DNAI1, CCDC151 |
| GO:0044458~motile cilium assembly | 1.81 × 10−4 | CCDC39, DNAAF3, DNAAF1, DMD, BBOF1, BBS5, CCDC40 |
| GO:0070286~axonemal dynein complex assembly | 1.83 × 10−4 | CCDC39, DNAAF3, DNAAF2, DNAAF1, CCDC151, CCDC40 |
| GO:0098609~cell-cell adhesion | 1.87 × 10−4 | MACF1, PDXDC1, ARHGAP18, CLINT1, BAIAP2L1, MPRIP, PPME1, TRIM29, STK38, MYO6, PACSIN2, EPS8L1, FLNB, EPS8L2, ERC1, LRRFIP1, ARGLU1, SPTBN1, PAK4, SH3GLB2, LYPLA2, IST1, CSNK1D, BAIAP2, RAB11B, HCFC1, ATXN2L, AFDN, EXOC3, ESYT2, TJP2, PLEC, EPHA2, EIF4G1 |
| GO:0051056~regulation of small GTPase mediated signal transduction | 2.47 × 10−4 | ARHGEF12, GDI1, RALGAPA1, RALGAPA2, ARHGEF17, FAM13A, ARAP2, TSC2, ARHGAP18, ARHGAP39, MYO9A, SIPA1L3, ARHGAP24, AKAP13, ABR, RHOT2, RALGAPB, ARHGEF4, SRGAP3, ARHGEF2, SRGAP2 |
| GO:0007224~smoothened signaling pathway | 3.03 × 10−4 | TTC26, IFT172, IFT80, HIPK1, TTBK2, TMEM231, HHAT, TMEM17, TTC21B, TCTN2, IQUB, WDPCP, DZIP1, CLUAP1 |
| GO:0043547~positive regulation of GTPase activity | 4.22 × 10−4 | GDI1, FAM13A, ARHGAP39, SIPA1L3, RPGR, FGF5, AKAP13, HERC2, SYNGAP1, DNM1L, BCAS3, DENND2C, ARHGEF12, ARHGEF17, ELMOD1, ARAP2, TSC2, AGAP9, RASA3, TBC1D20, NRG4, AKAP9, SPATA13, RAPGEF1, ARHGEF4, PKP4, DENND6B, ARHGEF2, ECT2L, LLGL2, TBC1D19, CAMK2D, ARHGAP18, AGAP4, ABR, ALS2CL, ERBB4, ERBB2, EPS8L1, EPS8L2, SRGAP3, SRGAP2, RALGDS, CYTH1, SPTBN1, GIT2, RAB3IP, ARHGAP27, MYO9A, ARHGAP24, PTK2, AFDN, DLG4, ST5, SGSM2, PLXNB1, AGRN |
| GO:0007030~Golgi organization | 6.13 × 10−4 | DYNC2H1, BCAS3, COG7, CSNK1D, SYNE1, VMP1, GAK, TBC1D20, GORASP1, GOLGB1, TRIP11, KIFC3, CLASP1, CLASP2 |
| GO:0018105~peptidyl-serine phosphorylation | 7.64 × 10−4 | SMG1, CAMK2D, DMPK, PRKCD, CSNK1D, CSNK1E, PRKCZ, PKD1, HIPK3, TTBK2, CLK1, GRK2, AKT2, STK38, RICTOR, PKN1, GAS6, CSNK1G2, ATR |
| GO:0008589~regulation of smoothened signaling pathway | 7.89 × 10−4 | INTU, C2CD3, ARL6, IFT140, TTC21B, FUZ, RPGRIP1L |
| GO:0003351~epithelial cilium movement | 7.98 × 10−4 | SPAG17, DNAH1, STK36, DNAI1, KIF27, CCDC40 |
| GO:0060287~epithelial cilium movement involved in determination of left/right asymmetry | 8.68 × 10−4 | CCDC39, DNAAF1, NPHP3, RFX3, CCDC40 |
| GO:0016337~single organismal cell-cell adhesion | 0.001486135 | VEZT, PTPRU, ADGRV1, CTNND1, ICAM2, ICAM5, PKD1, VMP1, ARVCF, DLG1, CDH1, NPHP1, RAPGEF1, FAT1, CTNNB1, PKP4 |
| GO:0036158~outer dynein arm assembly | 0.001676592 | DNAI2, CCDC114, DNAAF1, DNAH5, DNAI1, CCDC151 |
| GO:0007163~establishment or maintenance of cell polarity | 0.001694226 | DLG1, PARD3, FAT1, NPHP3, RPGRIP1L, CLASP1, SYNE2, CLASP2 |
| GO:0003356~regulation of cilium beat frequency | 0.002215945 | CCDC39, DNAH11, DNAAF1, CCDC40 |
| GO:0035469~determination of pancreatic left/right asymmetry | 0.002215945 | CCDC39, DNAAF1, NPHP3, CCDC40 |
| GO:0006351~transcription, DNA-templated | 0.002281724 | HDAC10, WWC1, CTNND1, ATN1, ZBTB20, CCAR2, CCAR1, ZNF83, ZNF606, ZMIZ2, ZNF84, CPNE1, ZNF326, CCNL2, KMT5A, ZNF446, SOX6, ZNF444, TRIM22, DDX17, ZNF440, ZNF19, KMT5C, LMO3, ZNF160, RFX2, PTOV1, RFX3, RFX1, EMSY, SUPT7L, FOXP1, ZNF91, DMTF1, TBL1XR1, MAK, ZNF439, MZF1, SF1, ZNF431, ZNF395, CASZ1, INO80E, CTBP2, ZNF23, PHF21A, NPAS2, SBNO2, ZNF708, ATXN1, NKX2-1, TP53BP1, ARGLU1, GTF2IRD2B, ZNF664, BANP, ZSCAN18, SMAD3, GTF2IRD2, ZBTB16, NR2F1, CNOT10, FOXN3, MED13L, MOV10, GON4L, BCL6, CNOT1, MAFK, MMS19, CNOT9, ZNF254, CREBZF, PHF3, DIDO1, ZNF253, KDM5C, ZNF493, CHD9, CHD8, CDCA7L, CHD7, AKAP8L, ZNF44, EFCAB6, CHD2, ZNF280D, SIN3A, CHMP1A, ZNF644, LRRFIP1, ZNF763, PELP1, NCOA2, BCAS3, RBM14, PAWR, IL16, ARID1A, ZFP90, SREBF2, NCOR2, ELF2, ZNF638, CRY2, CRY1, BDP1, GTF3C1, GTF3C3, KDM3B, ATF6B, SFSWAP, SATB1, CXXC1, ZNF518A, ZBTB44, ZNF69, IWS1, HDAC7, RXRA, MTA1, ERBB4, POLR2B, FAM120B, ZNF506, ERBB2, TP53INP1, E2F3, STAT6, APBB1, ZNF621, ZNF586, E2F8, HES4, MLXIP, SPEN, RBM39, KDM4B, EYA1, PCGF3, SAMD4B, ZNF76, HIPK1, YY1AP1, ZNF33B, NFIB, CTNNB1, ZNF611, PKN1, LPIN1, PAXBP1, SSBP3, MPHOSPH8 |
| GO:0016477~cell migration | 0.002504521 | LAMA5, SDC4, TNK2, WWC1, USP33, SORBS2, DGKZ, PRKCZ, PTPRF, ABI2, ERBB4, PEAK1, TAOK2, SPATA13, FAT1, ABL1, PIP5K1A, PLXNB1, ELMO3, GAS6, EPHA2, PAK4 |
| GO:0046777~protein autophosphorylation | 0.002504521 | DDR1, SMG1, CAMK2D, DAPK1, PASK, CLK4, MAPK15, PTK2, IGF1R, CLK1, FER, STK33, WNK1, ERBB4, PEAK1, MAK, ERBB2, AAK1, ABL1, CDK12, CSNK1G2, ATR |
| GO:0006897~endocytosis | 0.002600443 | TNK2, USP33, INPPL1, TSC2, AP2A1, CSNK1D, CSNK1E, CLINT1, C9ORF72, DNM2, MYO6, AAK1, ANKFY1, ESYT2, DNM1L, RAB5A, ATP9B, CSNK1G2, BCL2L1 |
| GO:0001701~in utero embryonic development | 0.003188547 | SLC34A2, MBNL1, SMAD3, C2CD3, PRKCSH, ANKRD11, CHD8, WDR19, CHD7, SRSF1, RPGRIP1L, PKD1, FOXP1, MUC1, RXRA, C6, SCO2, SIN3A, MYH9, CTNNB1, SOX6, BCL2L1, WDTC1 |
| GO:0010506~regulation of autophagy | 0.003466465 | VMP1, DAPK1, USP33, TP53INP1, PSAP, ABL1, ITPR1, EP300, HSPB1, FBXL2 |
| GO:0071910~determination of liver left/right asymmetry | 0.004225947 | CCDC39, DNAAF1, NPHP3, CCDC40 |
| GO:0060294~cilium movement involved in cell motility | 0.004225947 | DNAH1, CFAP46, CFAP54, GAS8 |
| GO:0007018~microtubule-based movement | 0.004348348 | DYNC2H1, DNAH3, DNAH12, DNAH2, DNAH10, DNAH5, DNAH6, DNHD1, AP2A1, KIF27, KIF19, KIF13A, KIFC3 |
| GO:0043001~Golgi to plasma membrane protein transport | 0.005604612 | BBS1, MACF1, RAB31, KIF13A, GCC2, ANK3, SPTBN1 |
| GO:0043984~histone H4-K16 acetylation | 0.006667143 | KMT2A, KANSL1, KANSL1L, OGT, MSL1, HCFC1 |
| GO:0034453~microtubule anchoring | 0.006890861 | CCDC187, FGFR1OP, GCC2, CLASP1, CLASP2 |
| GO:0032956~regulation of actin cytoskeleton organization | 0.008098045 | LRP1, PRKCD, ABL1, ARHGAP18, RICTOR, CELSR1, BAIAP2, GPM6B, CLASP2 |
| GO:0021591~ventricular system development | 0.00917602 | TTC21B, MBOAT7, HYDIN, AK8, KIF27 |
| GO:0090630~activation of GTPase activity | 0.009840864 | TBC1D8B, BCAS3, RALGAPB, RALGAPA1, AKT2, RALGAPA2, SGSM3, SGSM2, TBC1D8, PIP5K1A, FOXJ1, EPHA2 |
| GO:0051301~cell division | 0.0122476 | ANKLE2, NUMA1, CDCA7L, CEP164, KATNB1, PMF1, POGZ, CNTRL, CHMP1A, CDK20, ZNF207, NEK3, MAP4, KMT5A, CLASP1, CLASP2, DIS3L2, LMLN, LIG1, IST1, SPICE1, ATAD3B, CHFR, CCNA1, MAU2, STAG2, PPP2R2D, CDK10, ANAPC4, MAPRE3, BIRC6, ARHGEF2, CDK13, LLGL2 |
| GO:0000226~microtubule cytoskeleton organization | 0.012507657 | FER, SON, DST, RNF19A, ULK4, TACC2, TTL, PRKCZ, PTK2, CLASP1, CLASP2 |
| GO:0001947~heart looping | 0.013018715 | CCDC39, AHI1, SMAD3, C2CD3, IFT172, DNAAF1, NPHP3, CLUAP1, BBS5, CCDC40 |
| GO:0045880~positive regulation of smoothened signaling pathway | 0.014858592 | DYNC2H1, POR, INTU, STK36, IFT172, IFT80 |
| GO:0030010~establishment of cell polarity | 0.014858592 | WWC1, RICTOR, NEK3, PRKCZ, PKD1, PTK2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wijk, S.C.; Prabhala, P.; Löfdahl, A.; Nybom, A.; Lang, S.; Brunnström, H.; Bjermer, L.; Westergren-Thorsson, G.; Magnusson, M. Ciliated (FOXJ1+) Cells Display Reduced Ferritin Light Chain in the Airways of Idiopathic Pulmonary Fibrosis Patients. Cells 2022, 11, 1031. https://doi.org/10.3390/cells11061031
Wijk SC, Prabhala P, Löfdahl A, Nybom A, Lang S, Brunnström H, Bjermer L, Westergren-Thorsson G, Magnusson M. Ciliated (FOXJ1+) Cells Display Reduced Ferritin Light Chain in the Airways of Idiopathic Pulmonary Fibrosis Patients. Cells. 2022; 11(6):1031. https://doi.org/10.3390/cells11061031
Chicago/Turabian StyleWijk, Sofia C., Pavan Prabhala, Anna Löfdahl, Annika Nybom, Stefan Lang, Hans Brunnström, Leif Bjermer, Gunilla Westergren-Thorsson, and Mattias Magnusson. 2022. "Ciliated (FOXJ1+) Cells Display Reduced Ferritin Light Chain in the Airways of Idiopathic Pulmonary Fibrosis Patients" Cells 11, no. 6: 1031. https://doi.org/10.3390/cells11061031
APA StyleWijk, S. C., Prabhala, P., Löfdahl, A., Nybom, A., Lang, S., Brunnström, H., Bjermer, L., Westergren-Thorsson, G., & Magnusson, M. (2022). Ciliated (FOXJ1+) Cells Display Reduced Ferritin Light Chain in the Airways of Idiopathic Pulmonary Fibrosis Patients. Cells, 11(6), 1031. https://doi.org/10.3390/cells11061031