
Contribution of Endothelial Laminin-Binding Integrins to Cellular Processes Associated with Angiogenesis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Immunofluorescence Microscopy
2.1.1. Immunostaining
2.1.2. Microscopy
2.1.3. Analysis of Sprouting
2.2. Cell Culture
2.3. Organotypic Culture Assays
2.3.1. Bead-Sprout Assay
2.3.2. Planar Co-Culture
2.4. siRNA
2.5. Inducible shRNA
2.6. Quantitative PCR (qPCR)
2.7. Western Blotting
2.8. Migration
2.9. Animal Experiments
2.10. Statistical Analysis
3. Results
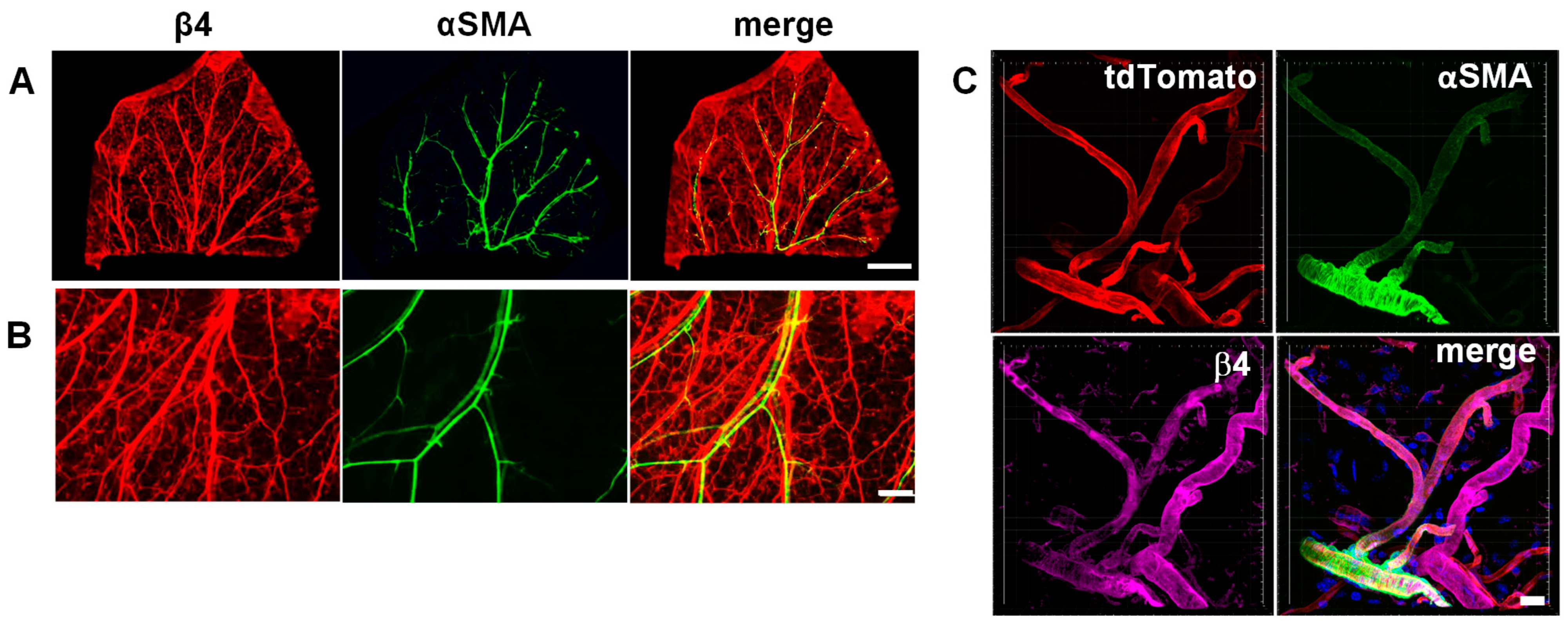
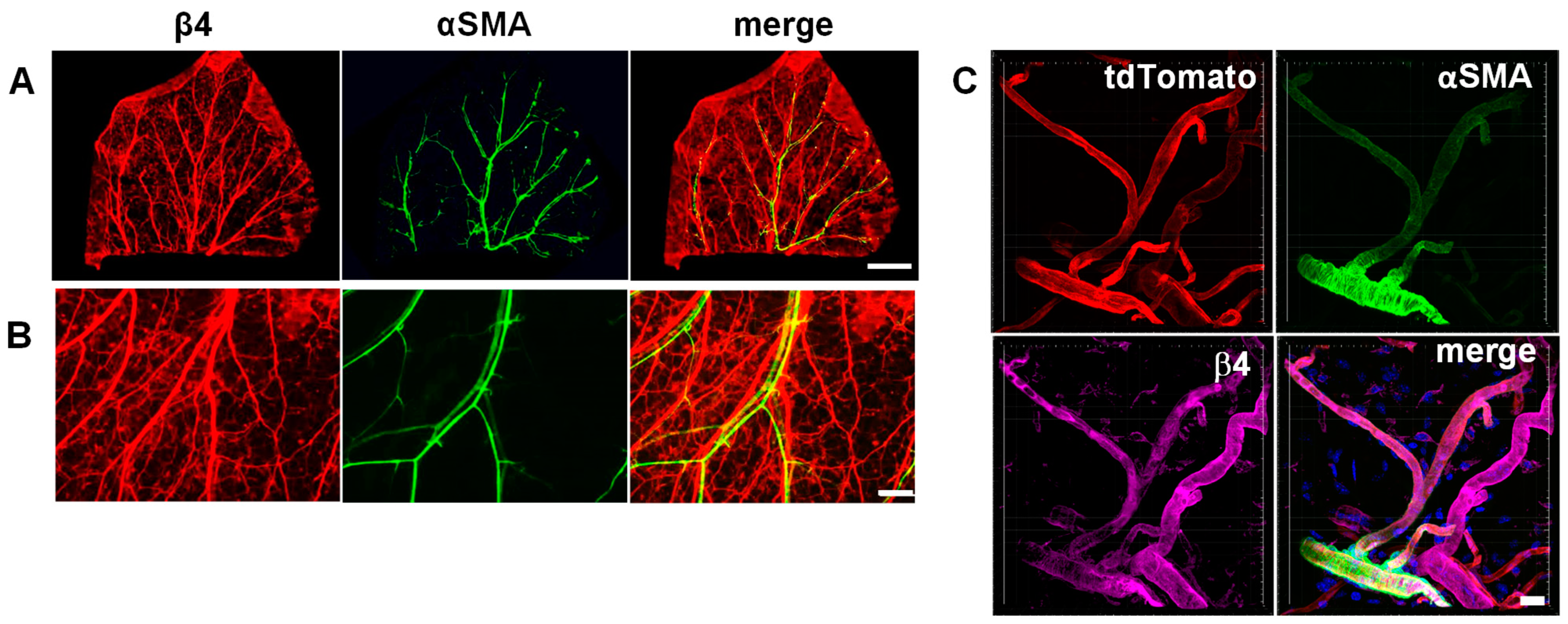
3.1. Integrin α6β4 Is Expressed in Veins and Small Vessels of the Dermis
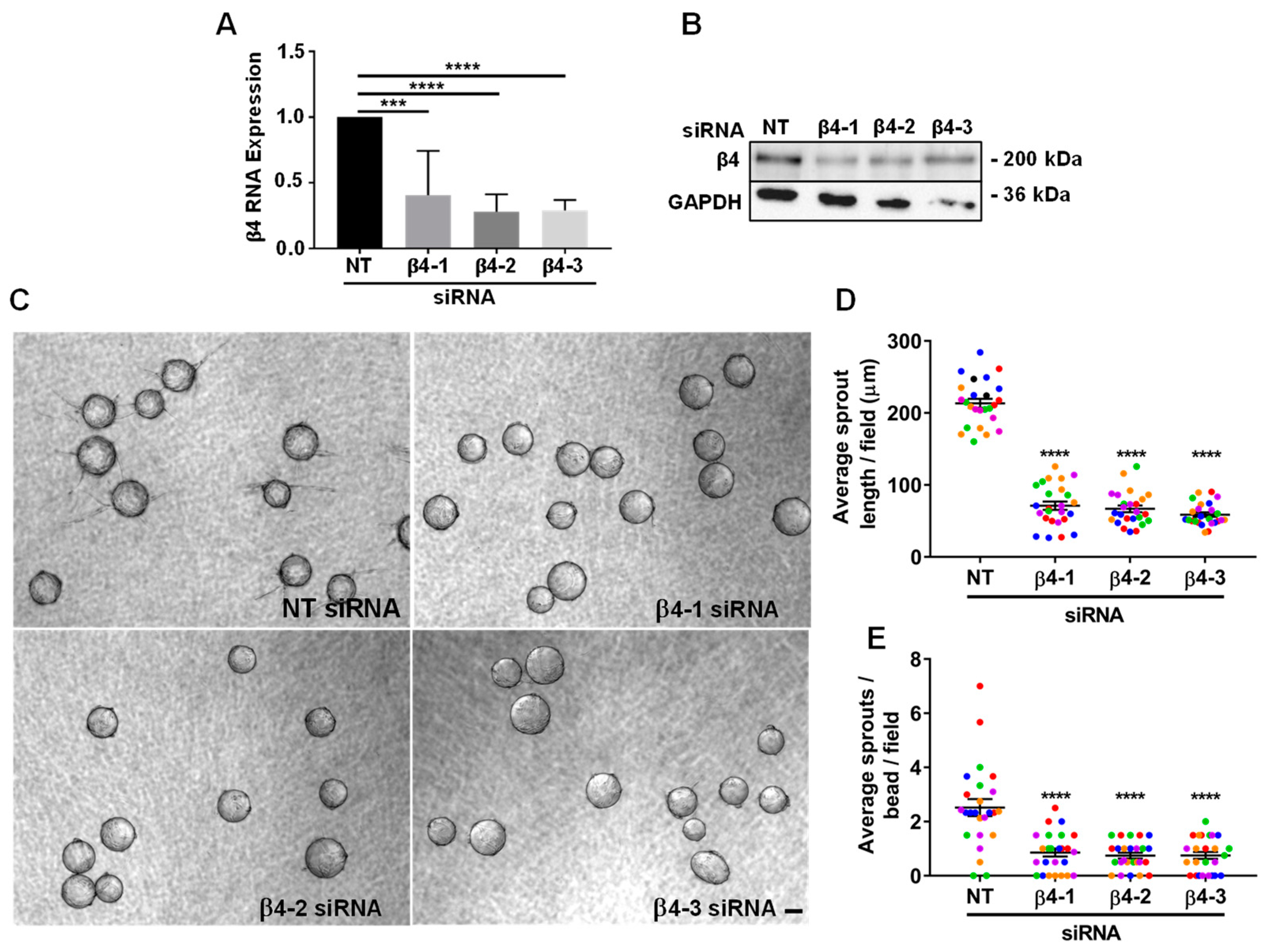
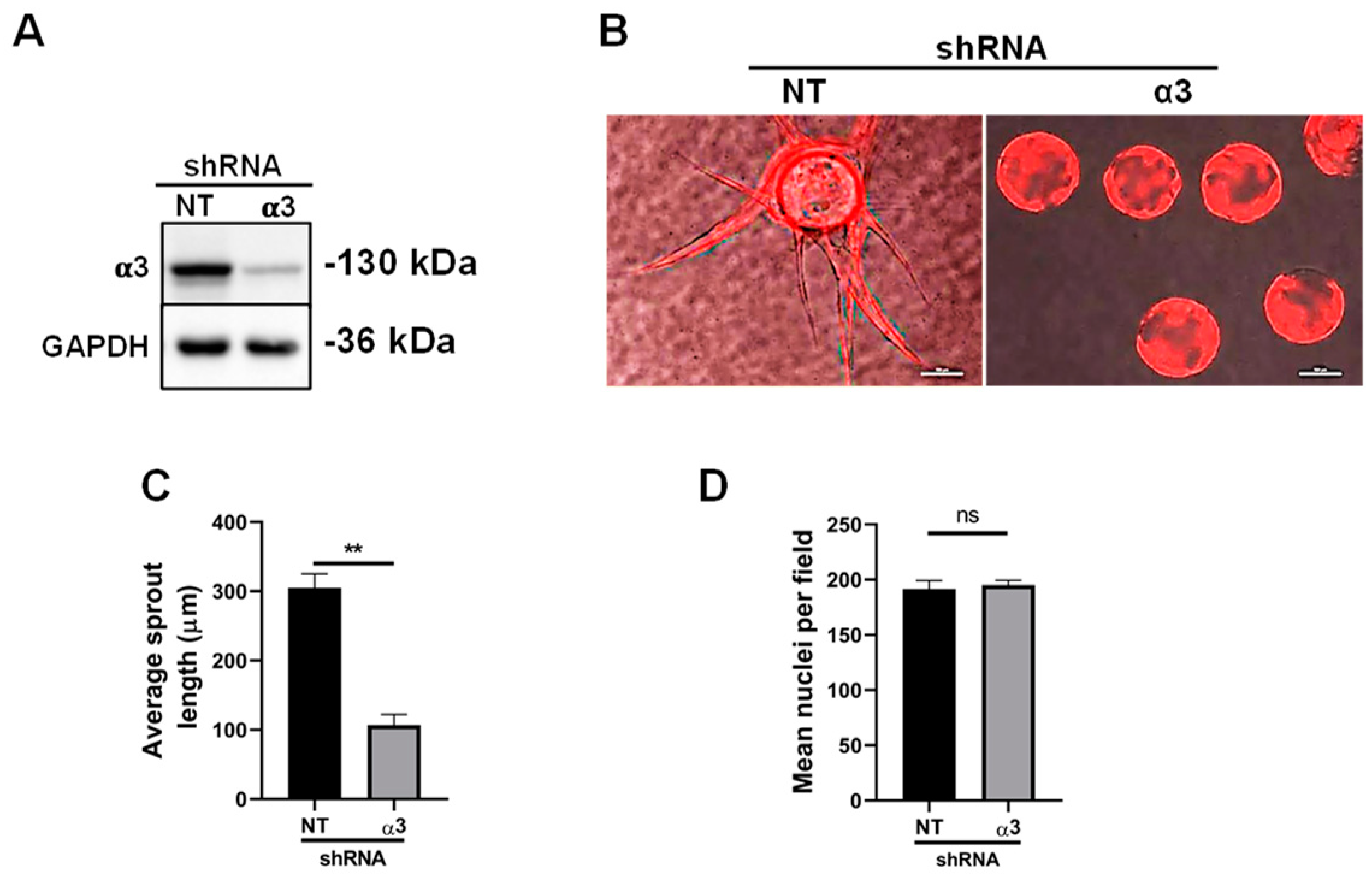
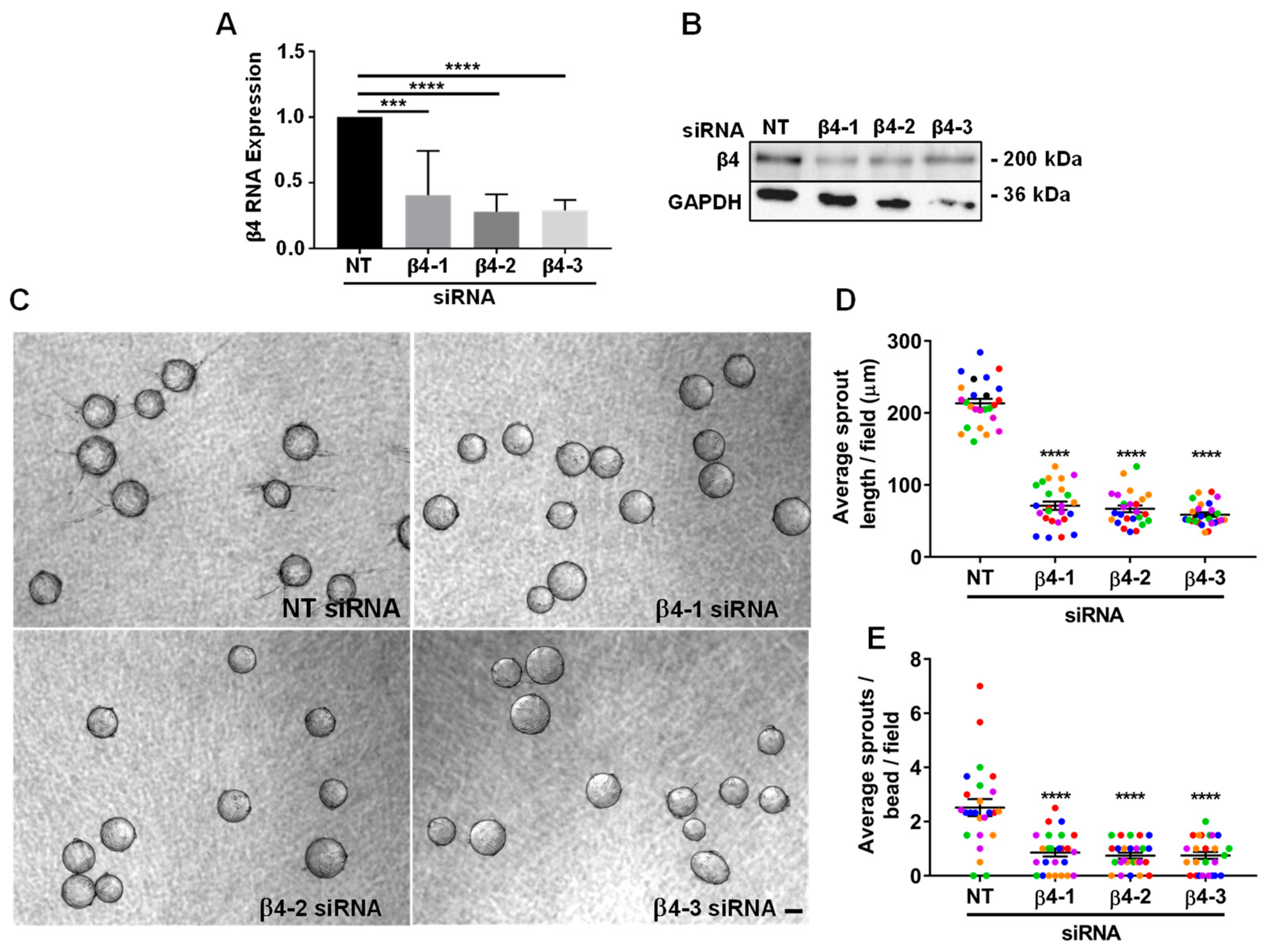
3.2. Integrin α6β4 Promotes Endothelial Morphogenesis
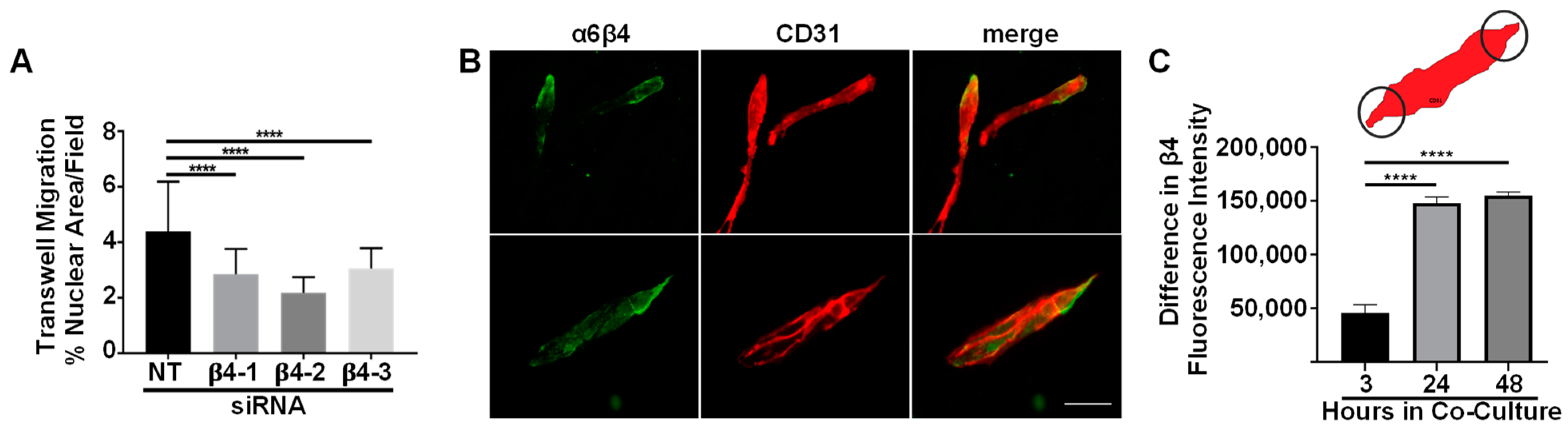
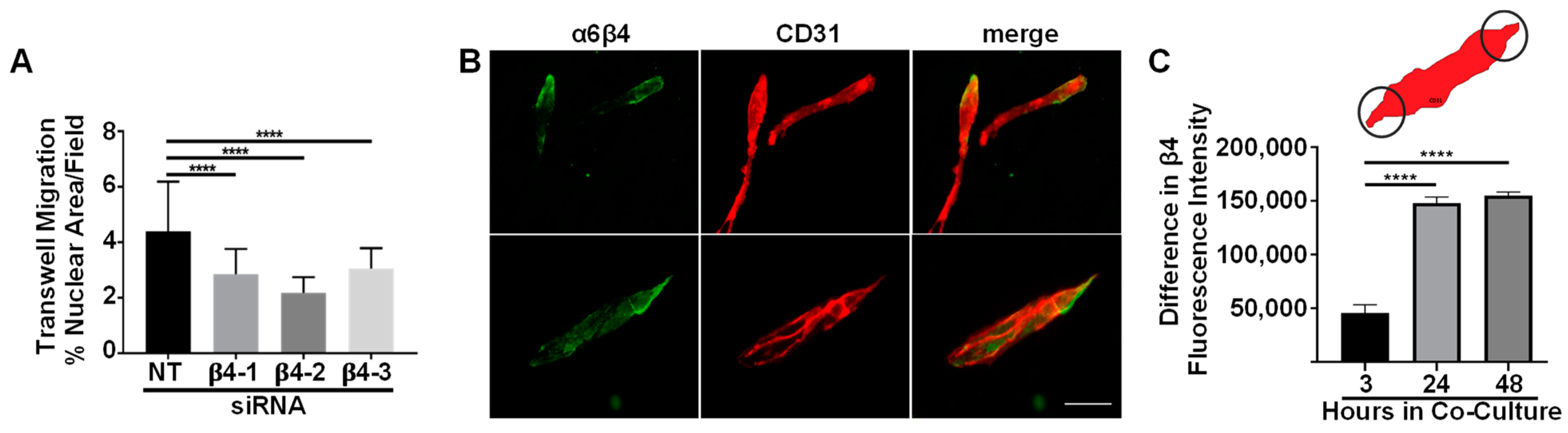
3.3. Integrin α6β4 Promotes Endothelial Migration
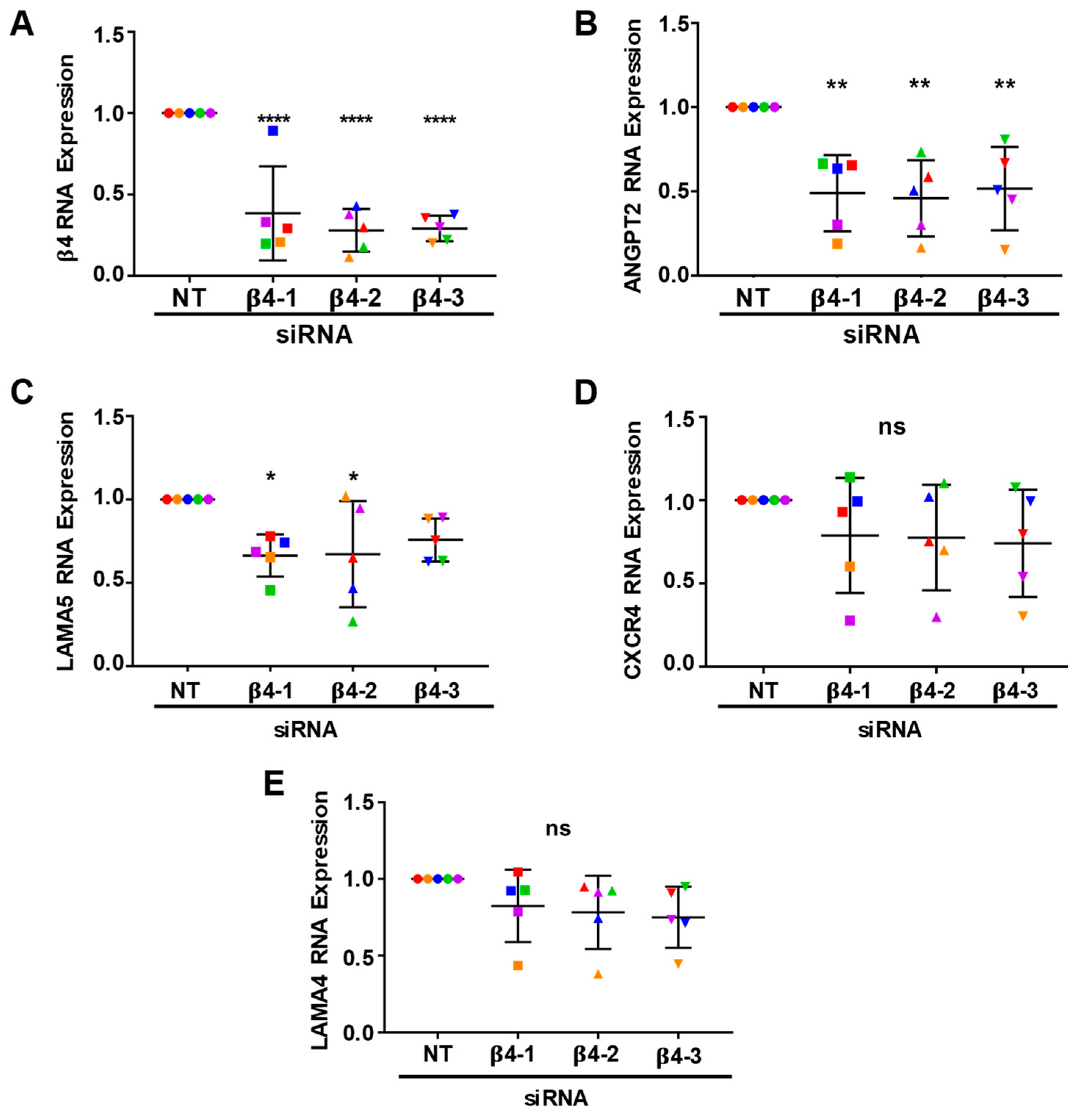
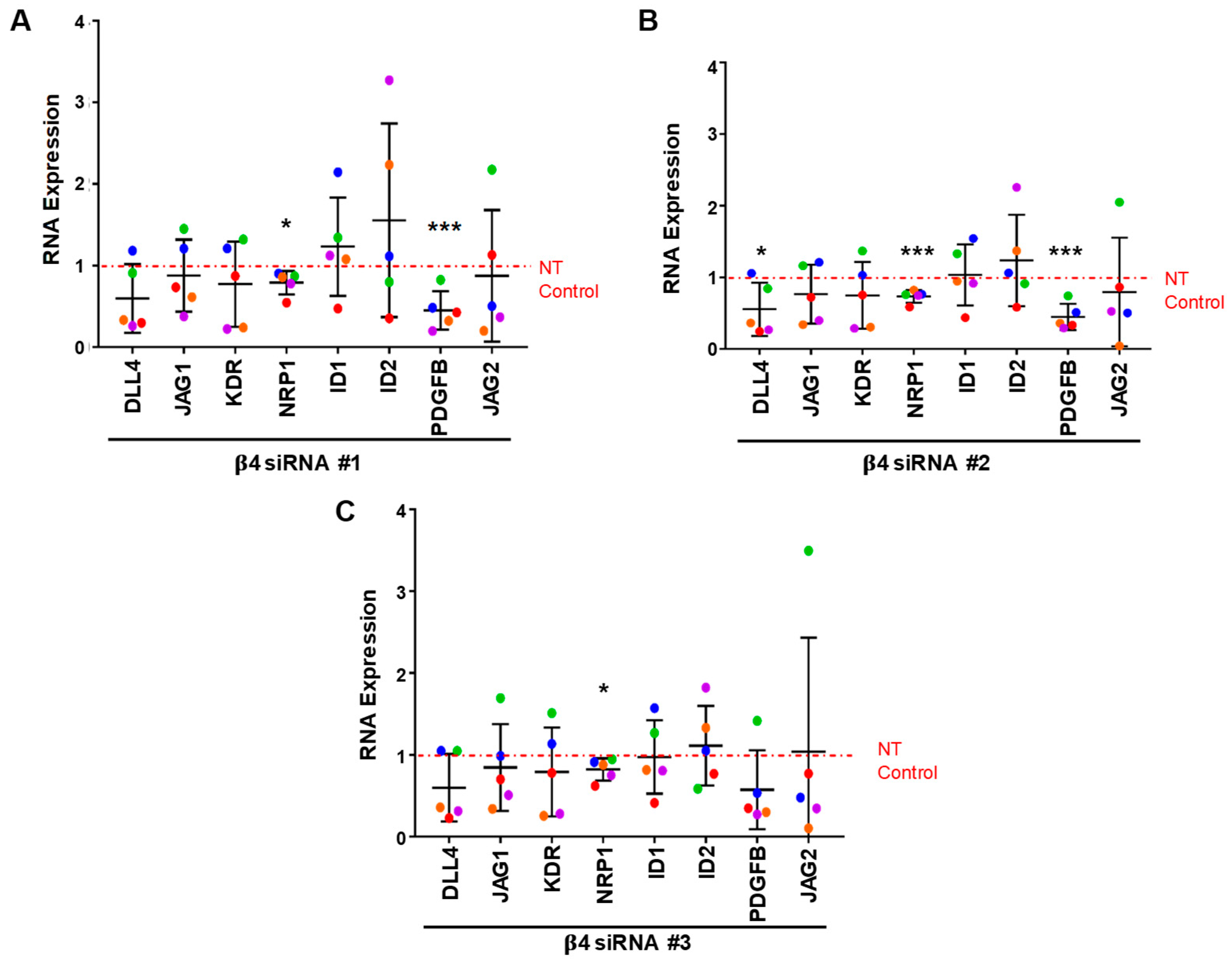
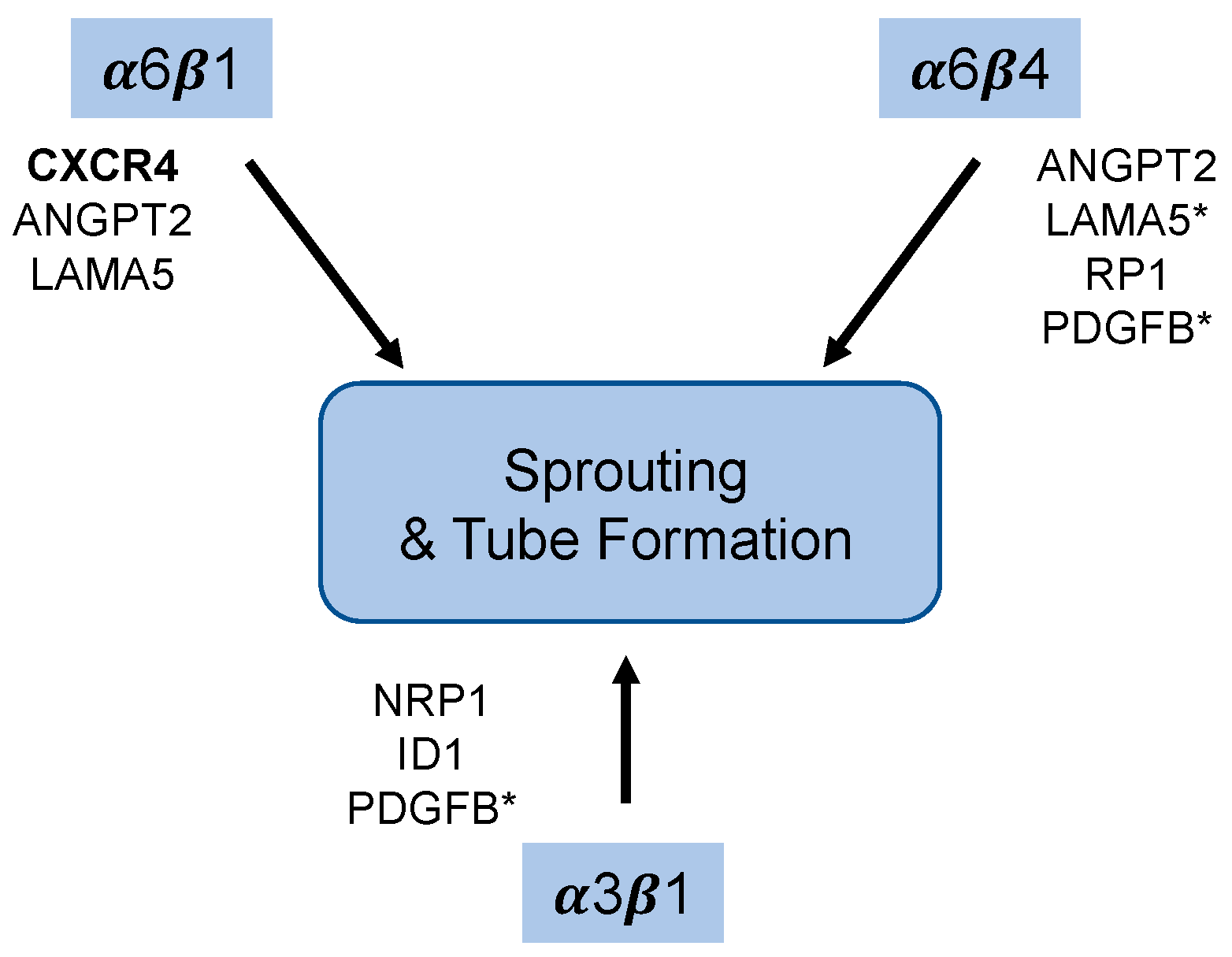
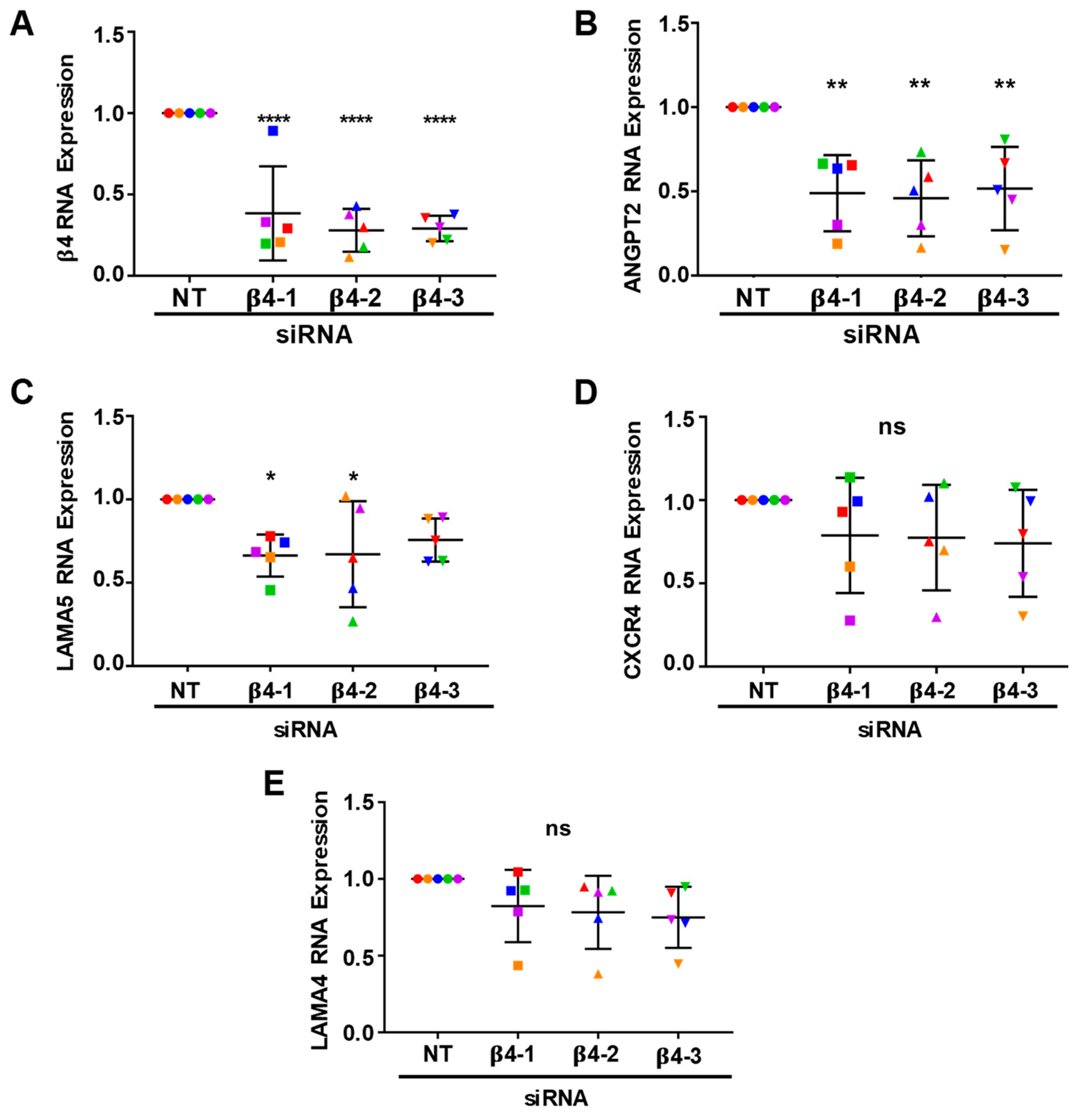
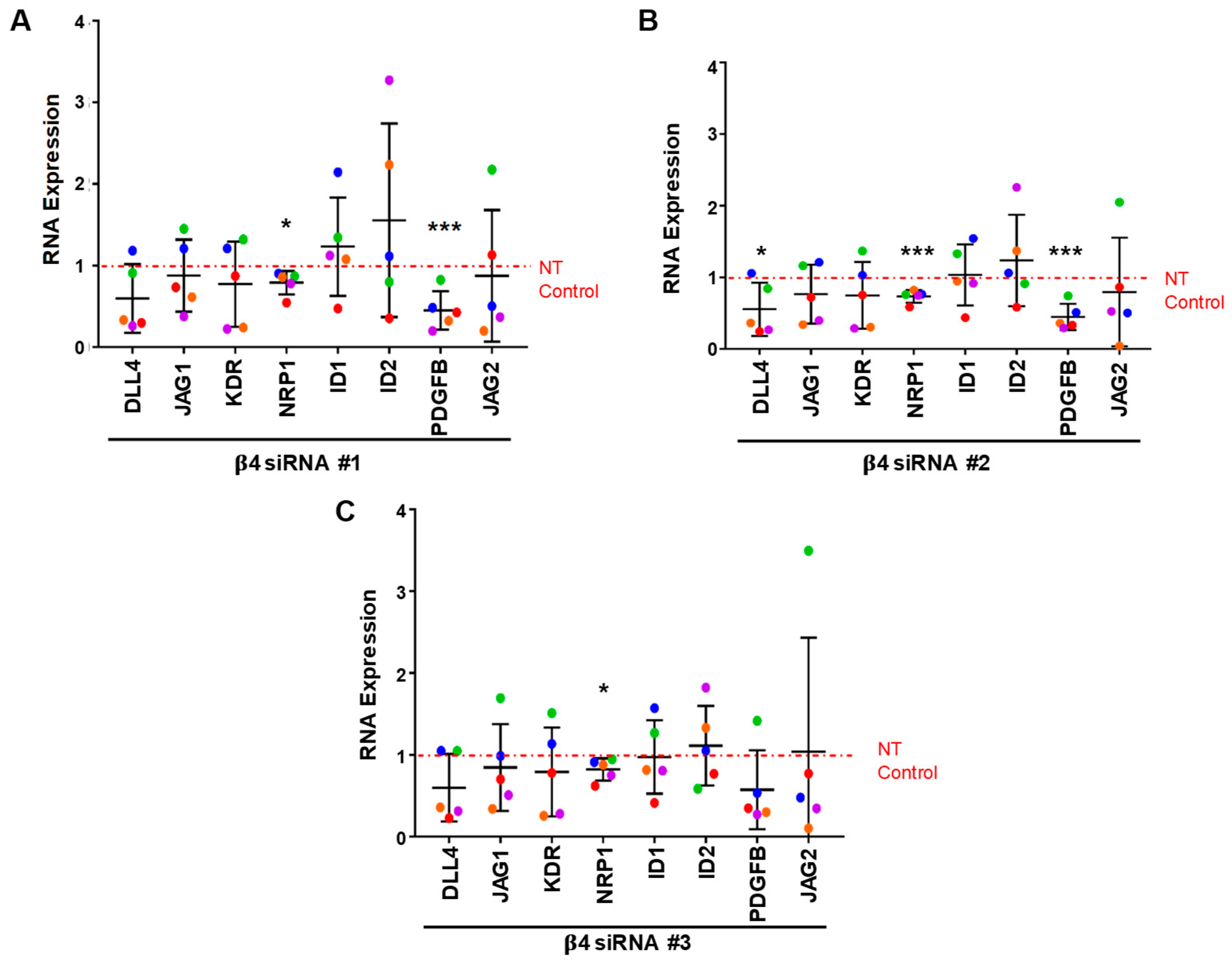
3.4. Integrin α6β4 Promotes the Expression of ANGPT2 and Other Angiogenesis-Associated Genes
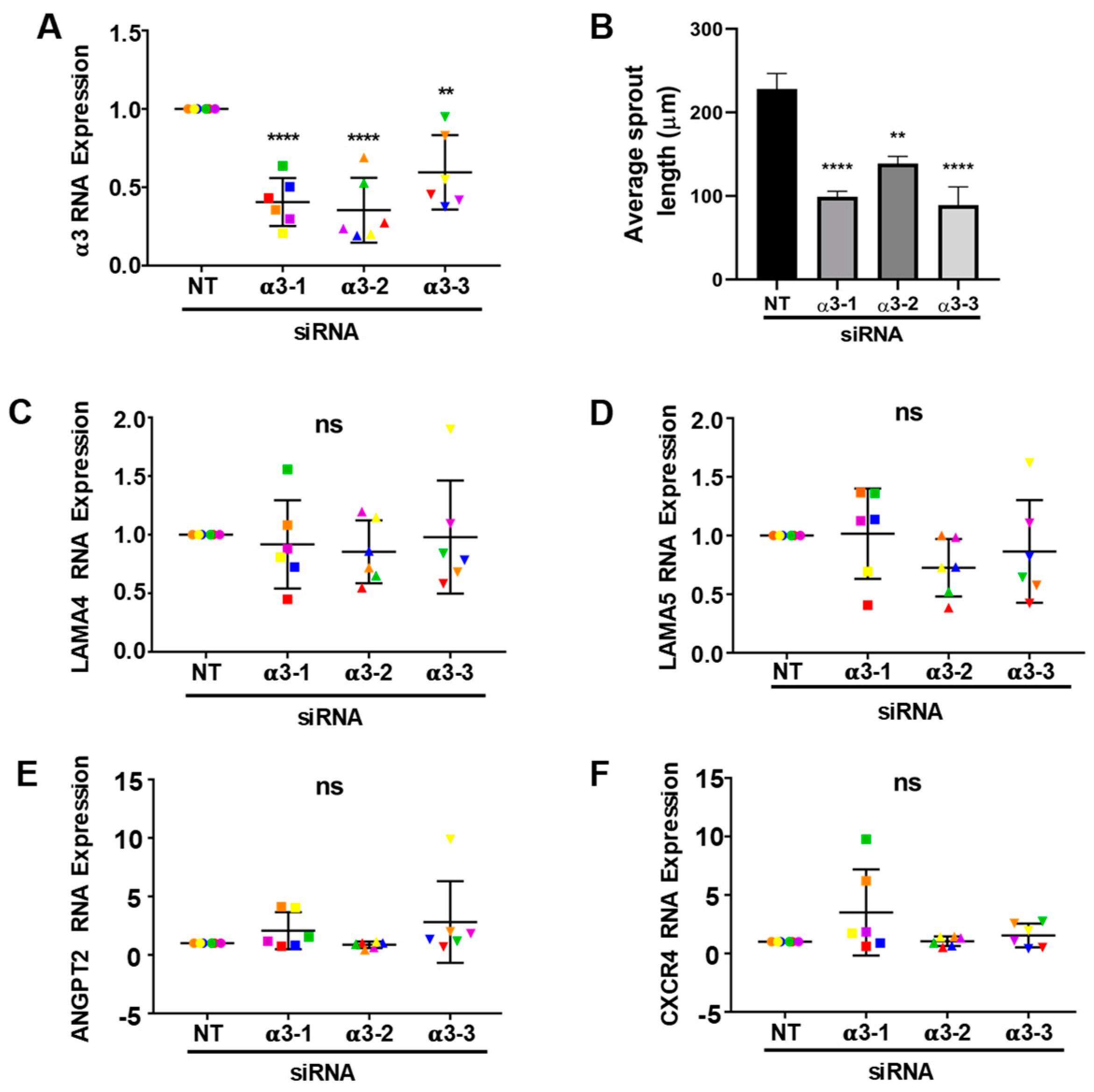
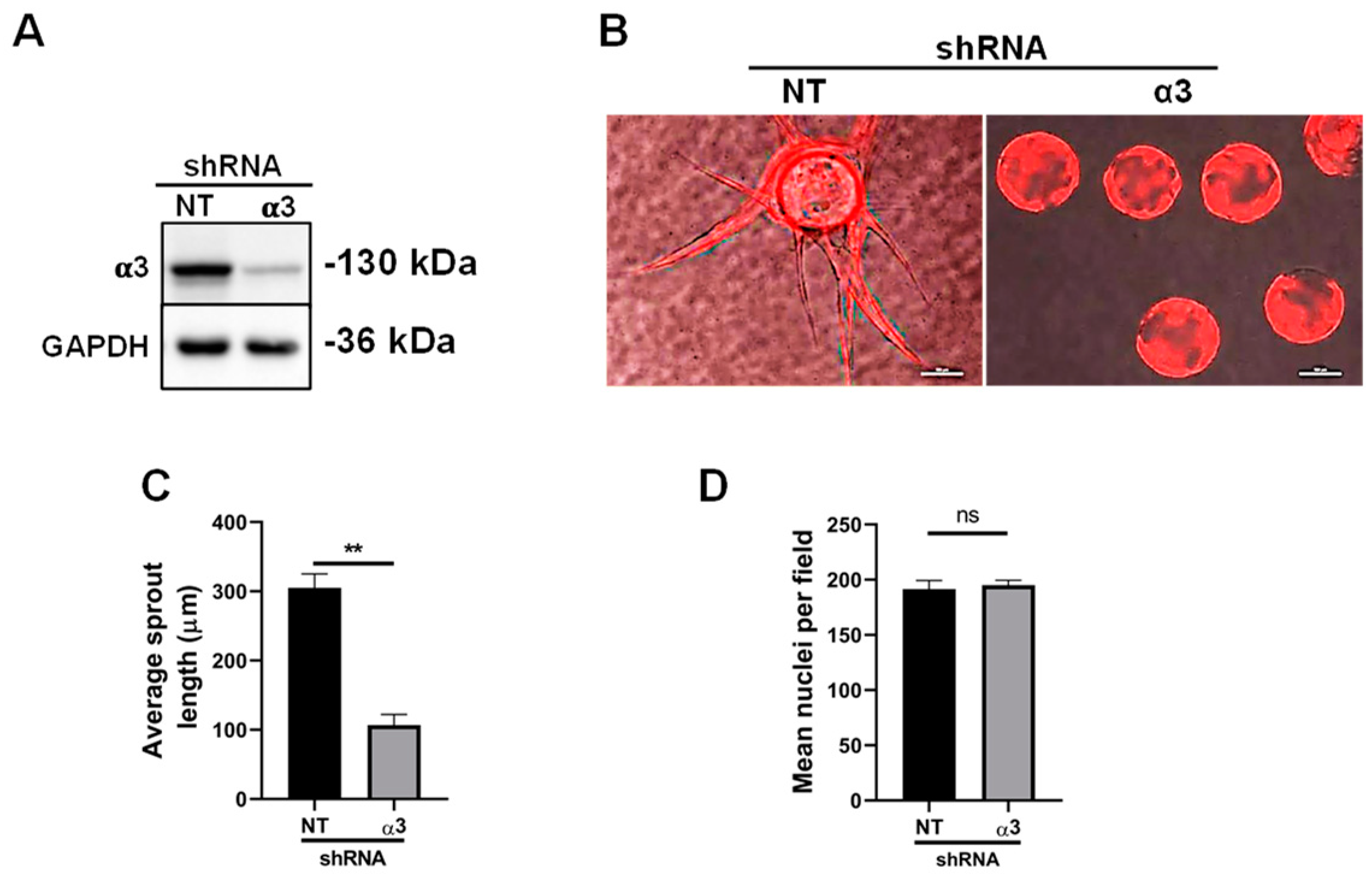
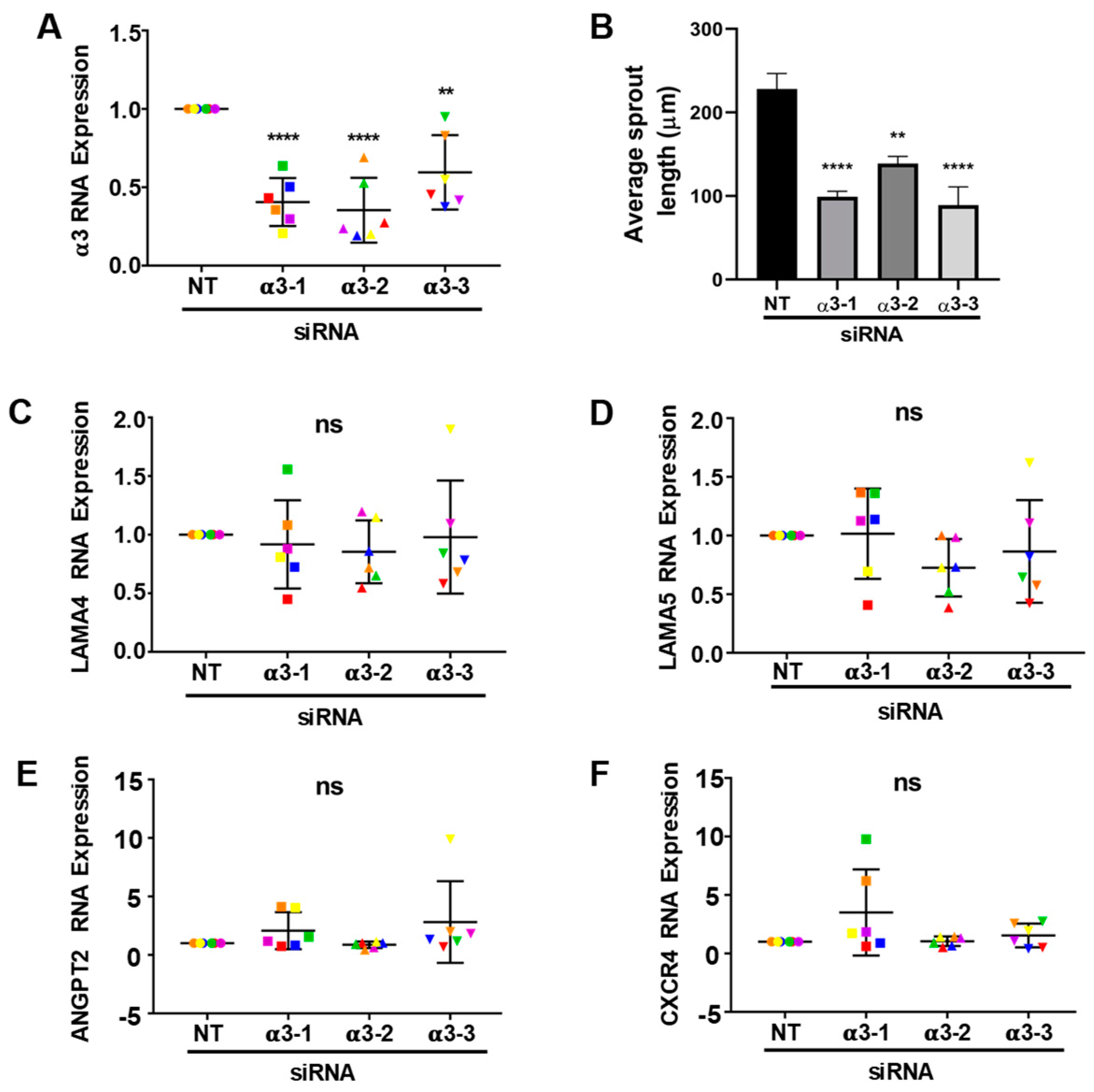
3.5. Integrin α3β1 Plays Overlapping and Distinct Roles during Endothelial Morphogenesis
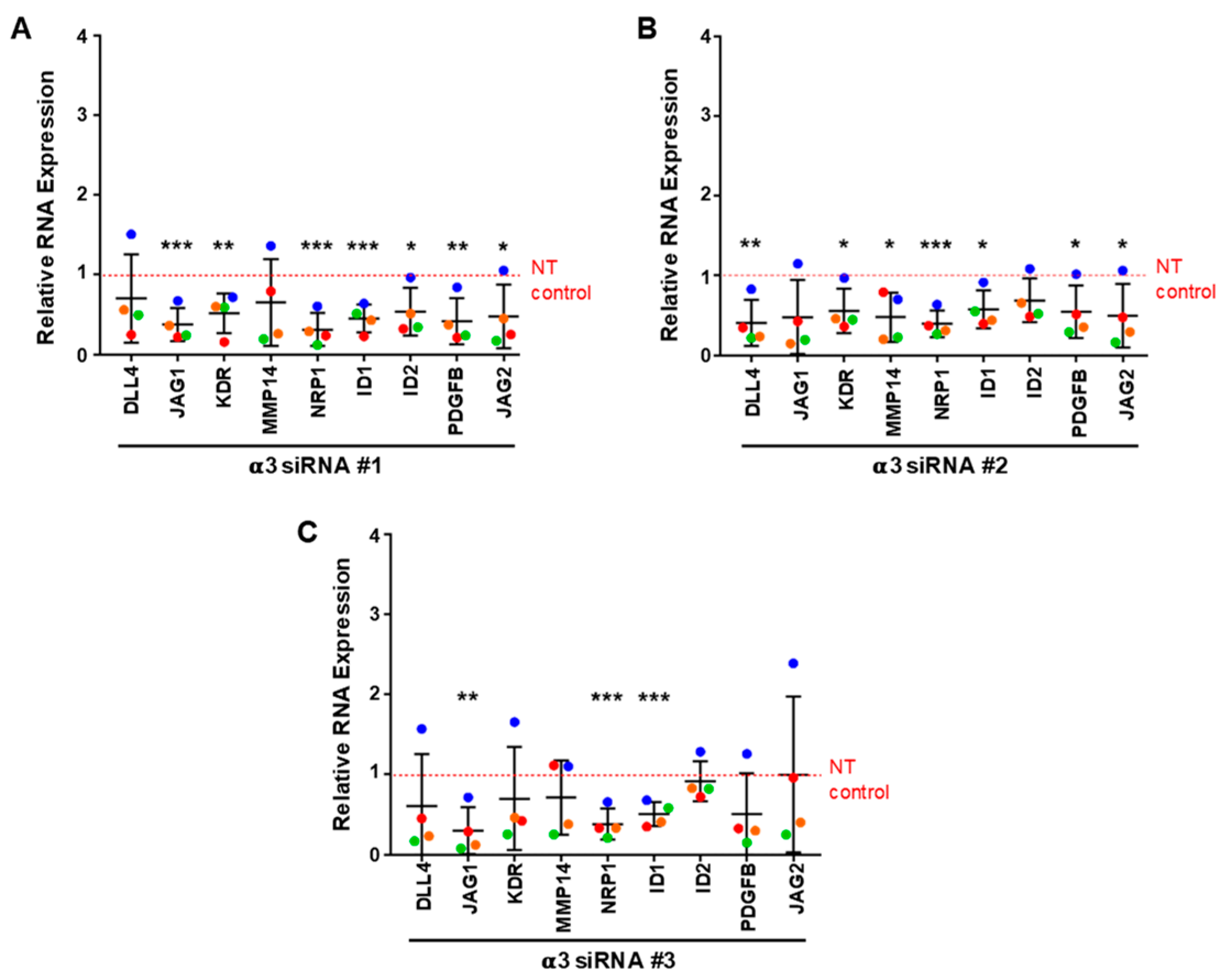
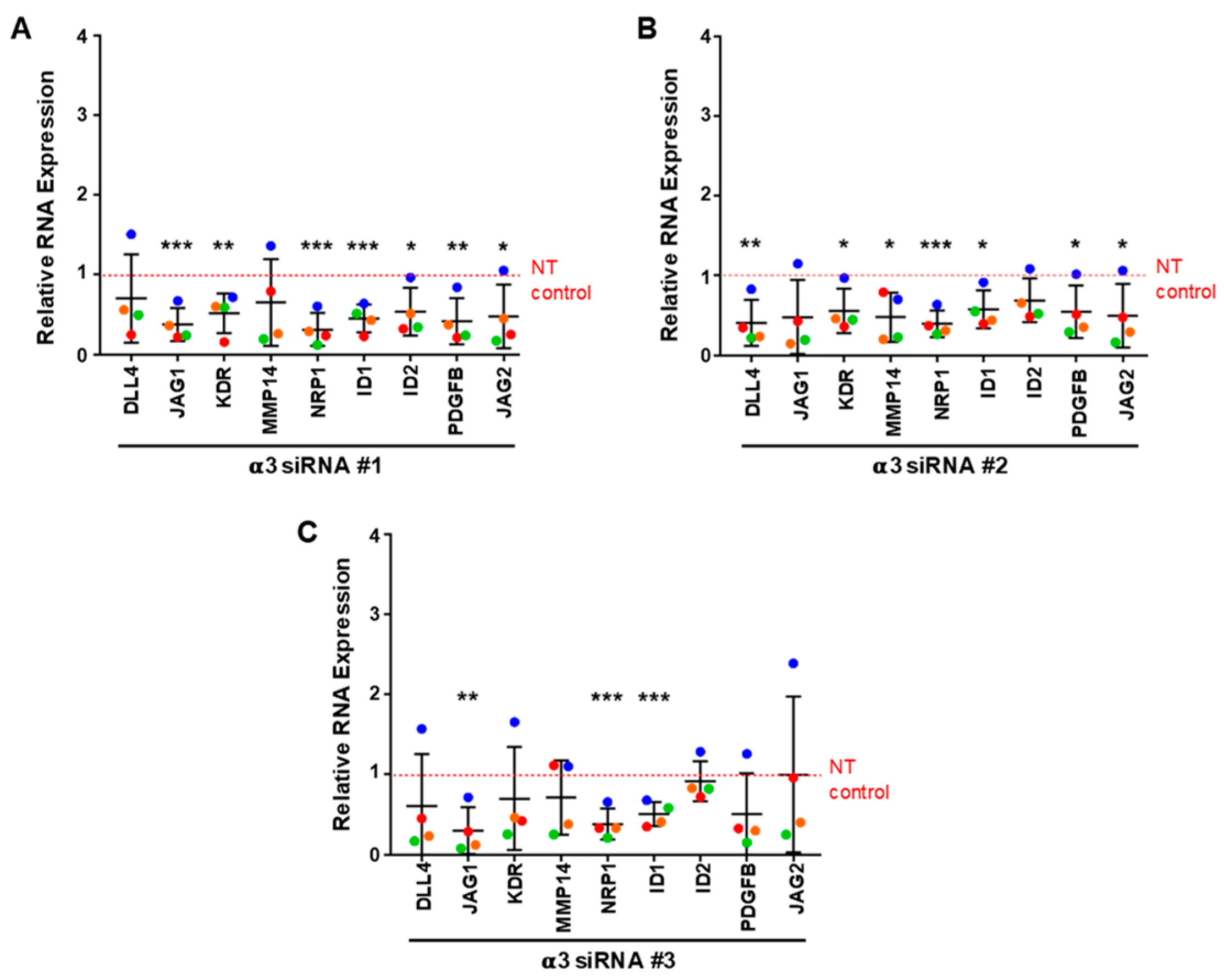
3.6. Integrin α3β1 Contributes to the Expression of a Distinct Set of Angiogenesis-Associated Genes
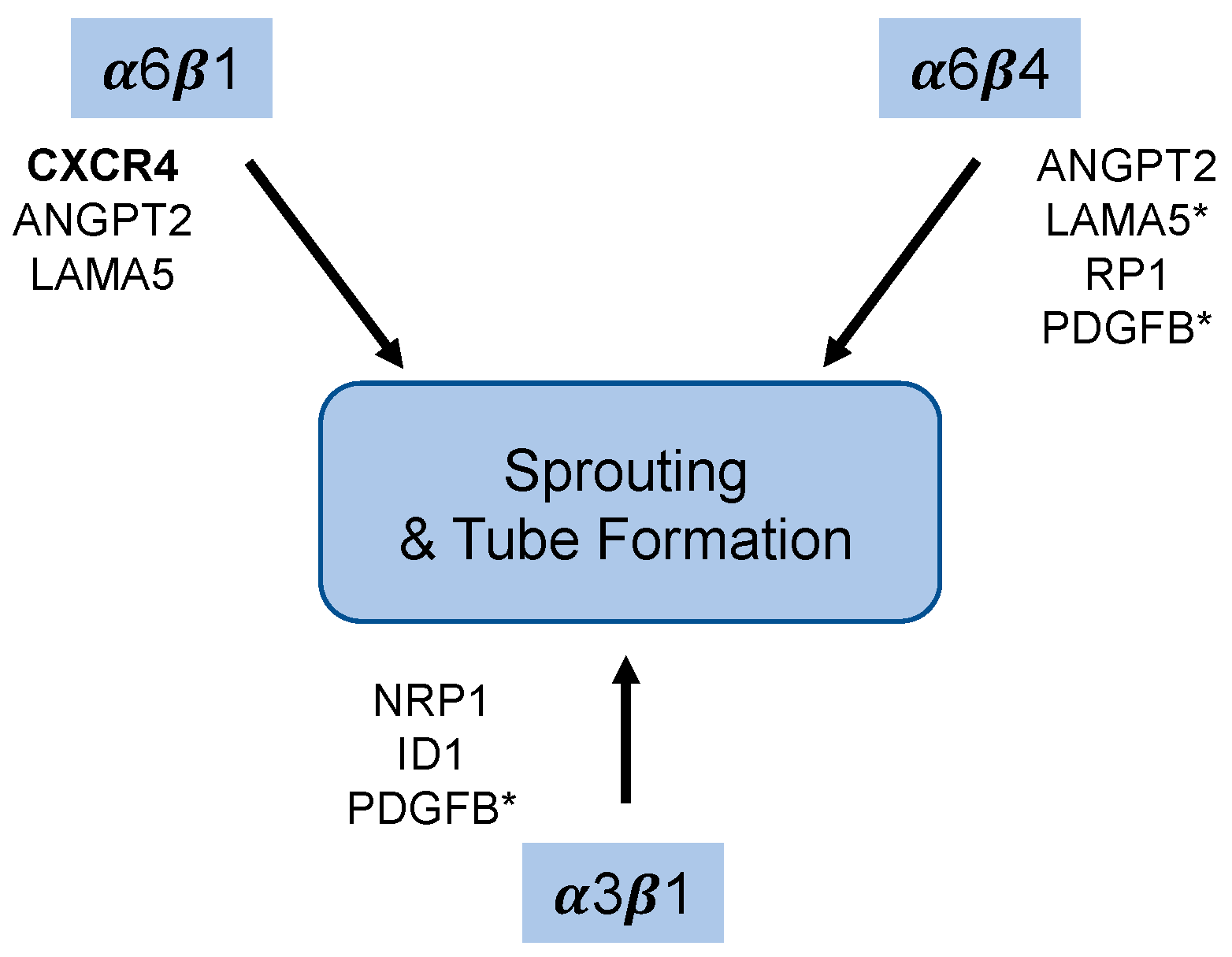
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carmeliet, P. Angiogenesis in health and disease. Nat. Med. 2003, 9, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P. Angiogenesis in life, disease and medicine. Nature 2005, 438, 932–936. [Google Scholar] [CrossRef]
- Senger, D.R.; Davis, G.E. Angiogenesis. Cold Spring Harb. Perspect. Biol. 2011, 3, a005090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eming, S.A.; Brachvogel, B.; Odorisio, T.; Koch, M. Regulation of angiogenesis: Wound healing as a model. Prog. Histochem. Cytochem. 2007, 42, 115–170. [Google Scholar] [CrossRef]
- Hallmann, R.; Horn, N.; Selg, M.; Wendler, O.; Pausch, F.; Sorokin, L.M. Expression and function of laminins in the embryonic and mature vasculature. Physiol. Rev. 2005, 85, 979–1000. [Google Scholar] [CrossRef] [Green Version]
- Turner, C.J.; Badu-Nkansah, K.; Hynes, R.O. Endothelium-derived fibronectin regulates neonatal vascular morphogenesis in an autocrine fashion. Angiogenesis 2017, 20, 519–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Pumiglia, K.; LaFlamme, S.E. Laminin-511 and alpha6 integrins regulate the expression of CXCR4 to promote endothelial morphogenesis. J. Cell Sci. 2020, 133, jcs246595. [Google Scholar] [CrossRef]
- Song, J.; Zhang, X.; Buscher, K.; Wang, Y.; Wang, H.; Di Russo, J.; Li, L.; Lutke-Enking, S.; Zarbock, A.; Stadtmann, A.; et al. Endothelial Basement Membrane Laminin 511 Contributes to Endothelial Junctional Tightness and Thereby Inhibits Leukocyte Transmigration. Cell Rep. 2017, 18, 1256–1269. [Google Scholar] [CrossRef] [Green Version]
- Avraamides, C.J.; Garmy-Susini, B.; Varner, J.A. Integrins in angiogenesis and lymphangiogenesis. Nat. Rev. Cancer 2008, 8, 604–617. [Google Scholar] [CrossRef] [Green Version]
- Nishiuchi, R.; Takagi, J.; Hayashi, M.; Ido, H.; Yagi, Y.; Sanzen, N.; Tsuji, T.; Yamada, M.; Sekiguchi, K. Ligand-binding specificities of laminin-binding integrins: A comprehensive survey of laminin-integrin interactions using recombinant alpha3beta1, alpha6beta1, alpha7beta1 and alpha6beta4 integrins. Matrix. Biol. 2006, 25, 189–197. [Google Scholar] [CrossRef]
- Kikkawa, Y.; Yu, H.; Genersch, E.; Sanzen, N.; Sekiguchi, K.; Fassler, R.; Campbell, K.P.; Talts, J.F.; Ekblom, P. Laminin isoforms differentially regulate adhesion, spreading, proliferation, and ERK activation of beta1 integrin-null cells. Exp. Cell Res. 2004, 300, 94–108. [Google Scholar] [CrossRef] [PubMed]
- Kreidberg, J.A.; Donovan, M.J.; Goldstein, S.L.; Rennke, H.; Shepherd, K.; Jones, R.C.; Jaenisch, R. Alpha 3 beta 1 integrin has a crucial role in kidney and lung organogenesis. Development 1996, 122, 3537–3547. [Google Scholar] [CrossRef] [PubMed]
- Georges-Labouesse, E.; Messaddeq, N.; Yehia, G.; Cadalbert, L.; Dierich, A.; Le Meur, M. Absence of integrin alpha 6 leads to epidermolysis bullosa and neonatal death in mice. Nat. Genet. 1996, 13, 370–373. [Google Scholar] [CrossRef] [PubMed]
- Dowling, J.; Yu, Q.C.; Fuchs, E. Beta4 integrin is required for hemidesmosome formation, cell adhesion and cell survival. J. Cell Biol. 1996, 134, 559–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, R.G.; Tavora, B.; Robinson, S.D.; Reynolds, L.E.; Szekeres, C.; Lamar, J.; Batista, S.; Kostourou, V.; Germain, M.A.; Reynolds, A.R.; et al. Endothelial alpha3beta1-integrin represses pathological angiogenesis and sustains endothelial-VEGF. Am. J. Pathol. 2010, 177, 1534–1548. [Google Scholar] [CrossRef]
- Germain, M.; De Arcangelis, A.; Robinson, S.D.; Baker, M.; Tavora, B.; D’Amico, G.; Silva, R.; Kostourou, V.; Reynolds, L.E.; Watson, A.; et al. Genetic ablation of the alpha 6-integrin subunit in Tie1Cre mice enhances tumour angiogenesis. J. Pathol. 2010, 220, 370–381. [Google Scholar] [CrossRef]
- Bouvard, C.; De Arcangelis, A.; Dizier, B.; Galy-Fauroux, I.; Fischer, A.M.; Georges-Labouesse, E.; Helley, D. Tie2-dependent knockout of alpha6 integrin subunit in mice reduces post-ischaemic angiogenesis. Cardiovasc. Res. 2012, 95, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Bouvard, C.; Segaoula, Z.; De Arcangelis, A.; Galy-Fauroux, I.; Mauge, L.; Fischer, A.M.; Georges-Labouesse, E.; Helley, D. Tie2-dependent deletion of alpha6 integrin subunit in mice reduces tumor growth and angiogenesis. Int. J. Oncol. 2014, 45, 2058–2064. [Google Scholar] [CrossRef] [Green Version]
- Seano, G.; Chiaverina, G.; Gagliardi, P.A.; di Blasio, L.; Puliafito, A.; Bouvard, C.; Sessa, R.; Tarone, G.; Sorokin, L.; Helley, D.; et al. Endothelial podosome rosettes regulate vascular branching in tumour angiogenesis. Nat. Cell Biol. 2014, 16, 931–941. [Google Scholar] [CrossRef]
- Welser-Alves, J.V.; Boroujerdi, A.; Tigges, U.; Wrabetz, L.; Feltri, M.L.; Milner, R. Endothelial beta4 integrin is predominantly expressed in arterioles, where it promotes vascular remodeling in the hypoxic brain. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 943–953. [Google Scholar] [CrossRef] [Green Version]
- Welser, J.V.; Halder, S.K.; Kant, R.; Boroujerdi, A.; Milner, R. Endothelial alpha6beta4 integrin protects during experimental autoimmune encephalomyelitis-induced neuroinflammation by maintaining vascular integrity and tight junction protein expression. J. Neuroinflammation 2017, 14, 217. [Google Scholar] [CrossRef]
- Nikolopoulos, S.N.; Blaikie, P.; Yoshioka, T.; Guo, W.; Giancotti, F.G. Integrin beta4 signaling promotes tumor angiogenesis. Cancer Cell 2004, 6, 471–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, D.; Singh, P.; Van De Water, L.; Laflamme, S.E. Dynamic Regulation of Integrin alpha6beta4 During Angiogenesis: Potential Implications for Pathogenic Wound Healing. Adv. Wound Care 2013, 2, 401–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishop, E.T.; Bell, G.T.; Bloor, S.; Broom, I.J.; Hendry, N.F.; Wheatley, D.N. An in vitro model of angiogenesis: Basic features. Angiogenesis 1999, 3, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, A.; Li, Q.F.; Zheng, Q.; Pumiglia, K. Loss of NF1 expression in human endothelial cells promotes autonomous proliferation and altered vascular morphogenesis. PLoS ONE 2012, 7, e49222. [Google Scholar] [CrossRef]
- Nakatsu, M.N.; Hughes, C.C. An optimized three-dimensional in vitro model for the analysis of angiogenesis. Methods Enzymol. 2008, 443, 65–82. [Google Scholar]
- Varney, S.D.; Betts, C.B.; Zheng, R.; Wu, L.; Hinz, B.; Zhou, J.; Van De Water, L. Hic-5 is required for myofibroblast differentiation by regulating mechanically dependent MRTF-A nuclear accumulation. J. Cell Sci. 2016, 129, 774–787. [Google Scholar] [CrossRef] [Green Version]
- Zheng, R.; Longmate, W.M.; DeFreest, L.; Varney, S.; Wu, L.; DiPersio, C.M.; Van De Water, L. Keratinocyte Integrin alpha3beta1 Promotes Secretion of IL-1alpha to Effect Paracrine Regulation of Fibroblast Gene Expression and Differentiation. J. Investig. Dermatol. 2019, 139, 2029–2038.e3. [Google Scholar] [CrossRef]
- He, L.; Vanlandewijck, M.; Mae, M.A.; Andrae, J.; Ando, K.; Del Gaudio, F.; Nahar, K.; Lebouvier, T.; Lavina, B.; Gouveia, L.; et al. Single-cell RNA sequencing of mouse brain and lung vascular and vessel-associated cell types. Sci. Data 2018, 5, 180160. [Google Scholar] [CrossRef] [Green Version]
- Sonnenberg, A.; Linders, C.J.; Daams, J.H.; Kennel, S.J. The alpha 6 beta 1 (VLA-6) and alpha 6 beta 4 protein complexes: Tissue distribution and biochemical properties. J. Cell Sci. 1990, 96 Pt 2, 207–217. [Google Scholar] [CrossRef]
- Hiran, T.S.; Mazurkiewicz, J.E.; Kreienberg, P.; Rice, F.L.; LaFlamme, S.E. Endothelial expression of the alpha6beta4 integrin is negatively regulated during angiogenesis. J. Cell Sci. 2003, 116, 3771–3781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakatsu, M.N.; Sainson, R.C.; Aoto, J.N.; Taylor, K.L.; Aitkenhead, M.; Perez-del-Pulgar, S.; Carpenter, P.M.; Hughes, C.C. Angiogenic sprouting and capillary lumen formation modeled by human umbilical vein endothelial cells (HUVEC) in fibrin gels: The role of fibroblasts and Angiopoietin-1. Microvasc. Res. 2003, 66, 102–112. [Google Scholar] [CrossRef]
- Newman, A.C.; Nakatsu, M.N.; Chou, W.; Gershon, P.D.; Hughes, C.C. The requirement for fibroblasts in angiogenesis: Fibroblast-derived matrix proteins are essential for endothelial cell lumen formation. Mol. Biol. Cell 2011, 22, 3791–3800. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, B.P.; Ryan, M.C.; Gil, S.G.; Carter, W.G. Deposition of laminin 5 in epidermal wounds regulates integrin signaling and adhesion. Curr. Opin. Cell Biol. 2000, 12, 554–562. [Google Scholar] [CrossRef]
- Frank, D.E.; Carter, W.G. Laminin 5 deposition regulates keratinocyte polarization and persistent migration. J. Cell Sci. 2004, 117, 1351–1363. [Google Scholar] [CrossRef] [Green Version]
- Hamelers, I.H.; Olivo, C.; Mertens, A.E.; Pegtel, D.M.; van der Kammen, R.A.; Sonnenberg, A.; Collard, J.G. The Rac activator Tiam1 is required for (alpha)3(beta)1-mediated laminin-5 deposition, cell spreading, and cell migration. J. Cell Biol. 2005, 171, 871–881. [Google Scholar] [CrossRef] [Green Version]
- Mercurio, A.M.; Rabinovitz, I.; Shaw, L.M. The alpha 6 beta 4 integrin and epithelial cell migration. Curr. Opin. Cell Biol. 2001, 13, 541–545. [Google Scholar] [CrossRef]
- Colburn, Z.T.; Jones, J.C. alpha6beta4 Integrin Regulates the Collective Migration of Epithelial Cells. Am. J. Respir. Cell Mol. Biol. 2017, 56, 443–452. [Google Scholar] [CrossRef] [Green Version]
- Elaimy, A.L.; Wang, M.; Sheel, A.; Brown, C.W.; Walker, M.R.; Amante, J.J.; Xue, W.; Chan, A.; Baer, C.E.; Goel, H.L.; et al. Real-time imaging of integrin beta4 dynamics using a reporter cell line generated by Crispr/Cas9 genome editing. J. Cell Sci. 2019, 132, jcs231241. [Google Scholar] [CrossRef] [Green Version]
- Enenstein, J.; Kramer, R.H. Confocal microscopic analysis of integrin expression on the microvasculature and its sprouts in the neonatal foreskin. J. Investig. Dermatol. 1994, 103, 381–386. [Google Scholar] [CrossRef] [Green Version]
- Mavria, G.; Vercoulen, Y.; Yeo, M.; Paterson, H.; Karasarides, M.; Marais, R.; Bird, D.; Marshall, C.J. ERK-MAPK signaling opposes Rho-kinase to promote endothelial cell survival and sprouting during angiogenesis. Cancer Cell 2006, 9, 33–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- del Toro, R.; Prahst, C.; Mathivet, T.; Siegfried, G.; Kaminker, J.S.; Larrivee, B.; Breant, C.; Duarte, A.; Takakura, N.; Fukamizu, A.; et al. Identification and functional analysis of endothelial tip cell-enriched genes. Blood 2010, 116, 4025–4033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strasser, G.A.; Kaminker, J.S.; Tessier-Lavigne, M. Microarray analysis of retinal endothelial tip cells identifies CXCR4 as a mediator of tip cell morphology and branching. Blood 2010, 115, 5102–5110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Smet, F.; Segura, I.; De Bock, K.; Hohensinner, P.J.; Carmeliet, P. Mechanisms of vessel branching: Filopodia on endothelial tip cells lead the way. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 639–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenzel, D.; Franco, C.A.; Estrach, S.; Mettouchi, A.; Sauvaget, D.; Rosewell, I.; Schertel, A.; Armer, H.; Domogatskaya, A.; Rodin, S.; et al. Endothelial basement membrane limits tip cell formation by inducing Dll4/Notch signalling in vivo. EMBO Rep. 2011, 12, 1135–1143. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, K.L.; Shaw, L.M.; Mercurio, A.M. Release of cAMP gating by the alpha6beta4 integrin stimulates lamellae formation and the chemotactic migration of invasive carcinoma cells. J. Cell Biol. 1998, 143, 1749–1760. [Google Scholar] [CrossRef] [Green Version]
- Homan, S.M.; Mercurio, A.M.; LaFlamme, S.E. Endothelial cells assemble two distinct alpha6beta4-containing vimentin-associated structures: Roles for ligand binding and the beta4 cytoplasmic tail. J. Cell Sci. 1998, 111 Pt 18, 2717–2728. [Google Scholar] [CrossRef]
- Homan, S.M.; Martinez, R.; Benware, A.; LaFlamme, S.E. Regulation of the association of alpha 6 beta 4 with vimentin intermediate filaments in endothelial cells. Exp. Cell Res. 2002, 281, 107–114. [Google Scholar] [CrossRef]
- Colburn, Z.T.; Jones, J.C.R. Complexes of alpha6beta4 integrin and vimentin act as signaling hubs to regulate epithelial cell migration. J. Cell Sci. 2018, 131, jcs214593. [Google Scholar] [CrossRef] [Green Version]
- Carmeliet, P.; Jain, R.K. Molecular mechanisms and clinical applications of angiogenesis. Nature 2011, 473, 298–307. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Bhat, A.; Woodnutt, G.; Lappe, R. Targeting the ANGPT-TIE2 pathway in malignancy. Nat. Rev. Cancer 2010, 10, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Thurston, G.; Daly, C. The complex role of angiopoietin-2 in the angiopoietin-tie signaling pathway. Cold Spring Harb. Perspect. Med. 2012, 2, a006550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kofler, N.M.; Simons, M. Angiogenesis versus arteriogenesis: Neuropilin 1 modulation of VEGF signaling. F1000Prime Rep. 2015, 7, 26. [Google Scholar] [CrossRef] [PubMed]
- Fantin, A.; Lampropoulou, A.; Gestri, G.; Raimondi, C.; Senatore, V.; Zachary, I.; Ruhrberg, C. NRP1 Regulates CDC42 Activation to Promote Filopodia Formation in Endothelial Tip Cells. Cell Rep. 2015, 11, 1577–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benezra, R. Role of Id proteins in embryonic and tumor angiogenesis. Trends Cardiovasc. Med. 2001, 11, 237–241. [Google Scholar] [CrossRef]
- Benezra, R.; Rafii, S.; Lyden, D. The Id proteins and angiogenesis. Oncogene 2001, 20, 8334–8341. [Google Scholar] [CrossRef]
- Volpert, O.V.; Pili, R.; Sikder, H.A.; Nelius, T.; Zaichuk, T.; Morris, C.; Shiflett, C.B.; Devlin, M.K.; Conant, K.; Alani, R.M. Id1 regulates angiogenesis through transcriptional repression of thrombospondin-1. Cancer Cell 2002, 2, 473–483. [Google Scholar] [CrossRef] [Green Version]
- Nishiyama, K.; Takaji, K.; Kataoka, K.; Kurihara, Y.; Yoshimura, M.; Kato, A.; Ogawa, H.; Kurihara, H. Id1 gene transfer confers angiogenic property on fully differentiated endothelial cells and contributes to therapeutic angiogenesis. Circulation 2005, 112, 2840–2850. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, A.; Itoh, F.; Nishiyama, K.; Takezawa, T.; Kurihara, H.; Itoh, S.; Kato, M. Inhibition of endothelial cell activation by bHLH protein E2-2 and its impairment of angiogenesis. Blood 2010, 115, 4138–4147. [Google Scholar] [CrossRef] [Green Version]
- Yanez-Mo, M.; Barreiro, O.; Gonzalo, P.; Batista, A.; Megias, D.; Genis, L.; Sachs, N.; Sala-Valdes, M.; Alonso, M.A.; Montoya, M.C.; et al. MT1-MMP collagenolytic activity is regulated through association with tetraspanin CD151 in primary endothelial cells. Blood 2008, 112, 3217–3226. [Google Scholar] [CrossRef]
- Sadej, R.; Grudowska, A.; Turczyk, L.; Kordek, R.; Romanska, H.M. CD151 in cancer progression and metastasis: A complex scenario. Lab. Investig. 2014, 94, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, K.; Svenson, K.B.; Longmate, W.M.; Gkirtzimanaki, K.; Sadej, R.; Wang, X.; Zhao, J.; Eliopoulos, A.G.; Berditchevski, F.; Dipersio, C.M. Suppression of integrin alpha3beta1 in breast cancer cells reduces cyclooxygenase-2 gene expression and inhibits tumorigenesis, invasion, and cross-talk to endothelial cells. Cancer Res. 2010, 70, 6359–6367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiPersio, C.M.; Shao, M.; Di Costanzo, L.; Kreidberg, J.A.; Hynes, R.O. Mouse keratinocytes immortalized with large T antigen acquire alpha3beta1 integrin-dependent secretion of MMP-9/gelatinase B. J. Cell Sci. 2000, 113 Pt 16, 2909–2921. [Google Scholar] [CrossRef] [PubMed]
- Iyer, V.; Pumiglia, K.; DiPersio, C.M. Alpha3beta1 integrin regulates MMP-9 mRNA stability in immortalized keratinocytes: A novel mechanism of integrin-mediated MMP gene expression. J. Cell Sci. 2005, 118, 1185–1195. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, H.; LaFlamme, S.E. Contribution of Endothelial Laminin-Binding Integrins to Cellular Processes Associated with Angiogenesis. Cells 2022, 11, 816. https://doi.org/10.3390/cells11050816
Xu H, LaFlamme SE. Contribution of Endothelial Laminin-Binding Integrins to Cellular Processes Associated with Angiogenesis. Cells. 2022; 11(5):816. https://doi.org/10.3390/cells11050816
Chicago/Turabian StyleXu, Hao, and Susan E. LaFlamme. 2022. "Contribution of Endothelial Laminin-Binding Integrins to Cellular Processes Associated with Angiogenesis" Cells 11, no. 5: 816. https://doi.org/10.3390/cells11050816
APA StyleXu, H., & LaFlamme, S. E. (2022). Contribution of Endothelial Laminin-Binding Integrins to Cellular Processes Associated with Angiogenesis. Cells, 11(5), 816. https://doi.org/10.3390/cells11050816