
Improving Cell Recovery: Freezing and Thawing Optimization of Induced Pluripotent Stem Cells


Abstract
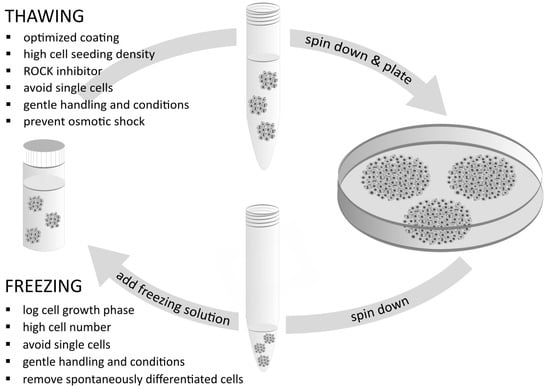
1. Introduction
2. Cryopreservation
2.1. General Guidelines for Working with iPSC during the Cryopreservation Process
2.2. Effect of Passaging Cells as Aggregates or as Single Cells on Cell Recovery after Thawing
2.3. Optimization of Protocol Steps during Cryopreservation
2.4. Logarithmic Cell Growth Phase before Freezing
2.5. Serum-Free Cryopreservation of iPSC
2.6. Cryopreservation with Serum
2.7. DMSO Concentration in the Freezing Medium
2.8. Different Dissociation Solutions Have a Strong Impact on Cryopreservation and Cell Recovery after Thawing
2.9. Economic View on Media, Reagents, and Supplements for the Cryopreservation of iPSC
2.10. Vitrification
3. Thawing of iPSC
3.1. Plasticware
3.2. Cell Attachment, Counting, and Viability
3.3. Preventing the Disruption of Cell Aggregates
3.4. Effects of the ROCK Inhibitor
3.5. Preventing Osmotic Shock during Thawing
3.6. Identifying Problems Causing Insufficient Cell Recovery
3.7. Supplementary Information
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| iPSC | induced pluripotent stem cells |
| ESC | embryonic stem cells |
| log phase | logarithmic cell growth phase (synonym: exponential phase) |
| ROCK inhibitor | Rho-associated, coiled-coil containing protein kinase inhibitor |
| FBS | fetal bovine serum |
| DMSO | dimethyl sulfoxide |
| KSR | knockout serum replacement |
| DMEM/F12 | Dulbecco’s Modified Eagle’s Medium/Ham’s F-12 |
| EDTA | ethylenediaminetetraacetic acid |
| PBS | phosphate-buffered saline |
| SOP | standard operating procedure |
| CVM | CryoLogic vitrification method |
References
- Ohnuki, M.; Takahashi, K. Present and future challenges of induced pluripotent stem cells. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140367. [Google Scholar] [CrossRef] [PubMed]
- Doss, M.X.; Sachinidis, A. Current Challenges of iPSC-Based Disease Modeling and Therapeutic Implications. Cells 2019, 8, 403. [Google Scholar] [CrossRef] [PubMed]
- Drexler, H.G.; Uphoff, C.C. Mycoplasma contamination of cell cultures: Incidence, sources, effects, detection, elimination, prevention. Cytotechnology 2002, 39, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Uphoff, C.C.; Drexler, H.G. Detection of mycoplasma contamination in cell cultures. Curr. Protoc. Mol. Biol. 2014, 2014, 28.4.1–28.4.14. [Google Scholar] [CrossRef]
- Uphoff, C.C.; Drexler, H.G. Elimination of Mycoplasmas from Infected Cell Lines Using Antibiotics. In Cancer Cell Culture; Humana Press: New York, NY, USA, 2011; Volume 731, pp. 105–114. [Google Scholar] [CrossRef]
- Acker, J.P.; McGann, L.E. Protective effect of intracellular ice during freezing? Cryobiology 2003, 46, 197–202. [Google Scholar] [CrossRef]
- Lawson, A.; Ahmad, H.; Sambanis, A. Cytotoxicity effects of cryoprotectants as single-component and cocktail vitrification solutions. Cryobiology 2011, 62, 115–122. [Google Scholar] [CrossRef]
- Fleming Glass, K.K.; Longmire, E.K.; Hubel, A. Optimization of a microfluidic device for diffusion-based extraction of dmso from a cell suspension. Int. J. Heat Mass Transf. 2008, 51, 5749. [Google Scholar] [CrossRef]
- MAZUR, P. Kinetics of Water Loss from Cells at Subzero Temperatures and the Likelihood of Intracellular Freezing. J. Gen. Physiol. 1963, 47, 347. [Google Scholar] [CrossRef]
- Mazur, P. Cryobiology: The freezing of biological systems. Science 1970, 168, 939–949. [Google Scholar] [CrossRef]
- Mazur, P.; Leibo, S.P.; Chu, E.H.Y. A two-factor hypothesis of freezing injury: Evidence from Chinese hamster tissue-culture cells. Exp. Cell Res. 1972, 71, 345–355. [Google Scholar] [CrossRef]
- He, X. Thermostability of Biological Systems: Fundamentals, Challenges, and Quantification. Open Biomed. Eng. J. 2011, 5, 47. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y.; Horiguchi, I.; Kino-oka, M.; Sugiyama, H. Slow freezing process design for human induced pluripotent stem cells by modeling intracontainer variation. Comput. Chem. Eng. 2020, 132, 106597. [Google Scholar] [CrossRef]
- Hayashi, Y.; Horiguchi, I.; Kino-oka, M.; Sugiyama, H. Model-based assessment of temperature profiles in slow freezing for human induced pluripotent stem cells. Comput. Chem. Eng. 2021, 144, 107150. [Google Scholar] [CrossRef]
- Hunt, C.J. Cryopreservation of Human Stem Cells for Clinical Application: A Review. Transfus. Med. Hemother. 2011, 38, 107–123. [Google Scholar] [CrossRef]
- Ware, C.B.; Nelson, A.M.; Blau, C.A. Controlled-rate freezing of human ES cells. Biotechniques 2005, 38, 879–883. [Google Scholar] [CrossRef]
- Li, R.; Yu, G.; Azarin, S.M.; Hubel, A. Freezing Responses in DMSO-Based Cryopreservation of Human iPS Cells: Aggregates Versus Single Cells. Tissue Eng. Part C. Methods 2018, 24, 289–299. [Google Scholar] [CrossRef]
- Konc, J.; Kanyo, K.; Varga, E.; Kriston, R.; Cseh, S. Oocyte cryopreservation: The birth of the first Hungarian babies from frozen oocytes. J. Assist. Reprod. Genet. 2008, 25, 349. [Google Scholar] [CrossRef][Green Version]
- Paynter, S.J. A rational approach to oocyte cryopreservation. Reprod. Biomed. Online 2005, 10, 578–586. [Google Scholar] [CrossRef]
- Gallardo, M.; Saenz, J.; Risco, R. Human oocytes and zygotes are ready for ultra-fast vitrification after 2 minutes of exposure to standard cpA solutions. Sci. Rep. 2019, 9, 15986. [Google Scholar] [CrossRef]
- T’joen, V.; De Grande, L.; Declercq, H.; Cornelissen, M. An Efficient, Economical Slow-Freezing Method for Large-Scale Human Embryonic Stem Cell Banking. Stem Cells Dev. 2011, 21, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Meneghel, J.; Kilbride, P.; Morris, J.G.; Fonseca, F. Physical events occurring during the cryopreservation of immortalized human T cells. PLoS ONE 2019, 14, e0217304. [Google Scholar] [CrossRef] [PubMed]
- Baust, J.M.; Corwin, W.L.; VanBuskirk, R.; Baust, J.G. Biobanking: The future of cell preservation strategies. In Biobanking in the 21st Century; Springer International Publishing: New York, NY, USA, 2015; pp. 37–53. ISBN 9783319205793. [Google Scholar]
- Baust, J.G.; Gage, A.A.; Klossner, D.; Clarke, D.; Miller, R.; Cohen, J.; Katz, A.; Polascik, T.; Clarke, H.; Baust, J.M. Issues critical to the successful application of cryosurgical ablation of the prostate. Technol. Cancer Res. Treat. 2007, 6, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.; Dou, M.; Mao, J.; Hou, Y.; Liu, S.; Zhao, L.; Lv, J.; Liu, Z.; Wang, Y.; Rao, W.; et al. Strong Hydration Ability of Silk Fibroin Suppresses Formation and Recrystallization of Ice Crystals During Cryopreservation. Biomacromolecules 2021, 23, 478–486. [Google Scholar] [CrossRef]
- Li, W.; Yang, G.; Zhang, A.; Xu, L.X. Numerical Study of Cell Cryo-Preservation: A Network Model of Intracellular Ice Formation. PLoS ONE 2013, 8, e58343. [Google Scholar] [CrossRef]
- Rowley, S.D.; Byrne, D.V. Low-temperature storage of bone marrow in nitrogen vapor-phase refrigerators: Decreased temperature gradients with an aluminum racking system. Transfusion 1992, 32, 750–754. [Google Scholar] [CrossRef]
- Yuan, Y.; Yang, Y.; Tian, Y.; Park, J.; Dai, A.; Roberts, R.M.; Liu, Y.; Han, X. Efficient long-term cryopreservation of pluripotent stem cells at −80 °C. Sci. Rep. 2016, 6, 34476. [Google Scholar] [CrossRef]
- Hartung, O.; Huo, H.; Daley, G.Q.; Schlaeger, T.M. Clump passaging and expansion of human embryonic and induced pluripotent stem cells on mouse embryonic fibroblast feeder cells. Curr. Protoc. Stem Cell Biol. 2010, 14, 1C.10.1–1C.10.15. [Google Scholar] [CrossRef]
- Cruvinel, E.; Ogusuku, I.; Cerioni, R.; Rodrigues, S.; Gonçalves, J.; Góes, M.E.; Alvim, J.M.; Silva, A.C.; de Souza Lino, V.; Boccardo, E.; et al. Long-term single-cell passaging of human iPSC fully supports pluripotency and high-efficient trilineage differentiation capacity. SAGE Open Med. 2020, 8, 205031212096645. [Google Scholar] [CrossRef]
- Baek, S.K.; Cho, Y.S.; Kim, I.S.; Jeon, S.B.; Moon, D.K.; Hwangbo, C.; Choi, J.W.; Kim, T.S.; Lee, J.H. A Rho-Associated Coiled-Coil Containing Kinase Inhibitor, Y-27632, Improves Viability of Dissociated Single Cells, Efficiency of Colony Formation, and Cryopreservation in Porcine Pluripotent Stem Cells. Cell. Reprogram. 2019, 21, 37–50. [Google Scholar] [CrossRef]
- Rizzino, A. Stimulating progress in regenerative medicine: Improving the cloning and recovery of cryopreserved human pluripotent stem cells with ROCK inhibitors. Regen. Med. 2010, 5, 799. [Google Scholar] [CrossRef] [PubMed]
- Rivera, T.; Zhao, Y.; Ni, Y.; Wang, J. Human-Induced Pluripotent Stem Cell Culture Methods Under cGMP Conditions. Curr. Protoc. Stem Cell Biol. 2020, 54, e117. [Google Scholar] [CrossRef]
- Trish, E.; Dimos, J.; Eggan, K. Freezing human ES cells. J. Vis. Exp. 2006, 1, e50. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.; Quek, L.E.; Titmarsh, D.; Krömer, J.O.; Kao, L.P.; Nielsen, L.; Wolvetang, E.; Cooper-White, J. Metabolic profiling and flux analysis of MEL-2 human embryonic stem cells during exponential growth at physiological and atmospheric oxygen concentrations. PLoS ONE 2014, 9, e112757. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Chen, G. Cryopreservation of human pluripotent stem cells in defined medium. Curr. Protoc. Stem Cell Biol. 2014, 2014, 1c.17.1–1c.17.13. [Google Scholar] [CrossRef]
- Beers, J.; Gulbranson, D.R.; George, N.; Siniscalchi, L.I.; Jones, J.; Thomson, J.A.; Chen, G. Passaging and colony expansion of human pluripotent stem cells by enzyme-free dissociation in chemically defined culture conditions. Nat. Protoc. 2012, 7, 2029–2040. [Google Scholar] [CrossRef]
- Chatterjee, I.; Li, F.; Kohler, E.E.; Rehman, J.; Malik, A.B.; Wary, K.K. Induced pluripotent stem (iPS) cell culture methods and induction of differentiation into endothelial cells. In Methods in Molecular Biology; Humana Press Inc.: New York, NY, USA, 2016; Volume 1357, pp. 311–327. [Google Scholar]
- Tamm, C.; Galitó, S.P.; Annerén, C. A comparative study of protocols for mouse embryonic stem cell culturing. PLoS ONE 2013, 8, e81156. [Google Scholar] [CrossRef]
- Mummery, C.L.; Zhang, J.; Ng, E.S.; Elliott, D.A.; Elefanty, A.G.; Kamp, T.J. Differentiation of Human ES and iPS Cells to Cardiomyocytes: A Methods Overview. Circ. Res. 2012, 111, 344. [Google Scholar] [CrossRef]
- Morris, C.B. Cryopreservation of animal and human cell lines. Methods Mol. Biol. 1995, 38, 179–187. [Google Scholar] [CrossRef]
- Claassen, D.A.; Desler, M.M.; Rizzino, A. ROCK inhibition enhances the recovery and growth of cryopreserved human embryonic stem cells and human induced pluripotent stem cells. Mol. Reprod. Dev. 2009, 76, 722–732. [Google Scholar] [CrossRef]
- Wang, J.; Hao, J.; Bai, D.; Gu, Q.; Han, W.; Wang, L.; Tan, Y.; Li, X.; Xue, K.; Han, P.; et al. Generation of clinical-grade human induced pluripotent stem cells in Xeno-free conditions. Stem Cell Res. Ther. 2015, 6, 223. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, Y.; Noguchi, H.; Yukawa, H.; Oishi, K.; Matsushita, K.; Iwata, H.; Hayashi, S. Cryopreservation of Induced Pluripotent Stem Cells. Cell Med. 2012, 3, 89. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, M.; Taniguchi, Y.; Senda, S.; Takizawa, N.; Ichisaka, T.; Asano, K.; Morizane, A.; Doi, D.; Takahashi, J.; Nishizawa, M.; et al. A novel efficient feeder-free culture system for the derivation of human induced pluripotent stem cells. Sci. Rep. 2014, 4, 3594. [Google Scholar] [CrossRef] [PubMed]
- Holm, F.; Ström, S.; Inzunza, J.; Baker, D.; Strömberg, A.M.; Rozell, B.; Feki, A.; Bergström, R.; Hovatta, O. An effective serum- and xeno-free chemically defined freezing procedure for human embryonic and induced pluripotent stem cells. Hum. Reprod. 2010, 25, 1271. [Google Scholar] [CrossRef] [PubMed]
- Wagner, K.; Welch, D. Cryopreserving and recovering of human iPS cells using complete knockout serum replacement feeder-free medium. J. Vis. Exp. 2010, 41, e2237. [Google Scholar] [CrossRef]
- Wagner, K.; Welch, D. Feeder-free adaptation, culture and passaging of human IPS cells using complete knockout serum replacement feeder-free medium. J. Vis. Exp. 2010, 41, e2236. [Google Scholar] [CrossRef]
- Chen, G.; Gulbranson, D.R.; Hou, Z.; Bolin, J.M.; Ruotti, V.; Probasco, M.D.; Smuga-Otto, K.; Howden, S.E.; Diol, N.R.; Propson, N.E.; et al. Chemically defined conditions for human iPSC derivation and culture. Nat. Methods 2011, 8, 424–429. [Google Scholar] [CrossRef]
- Wagh, V.; Meganathan, K.; Jagtap, S.; Gaspar, J.A.; Winkler, J.; Spitkovsky, D.; Hescheler, J.; Sachinidis, A. Effects of Cryopreservation on the Transcriptome of Human Embryonic Stem Cells After Thawing and Culturing. Stem Cell Rev. Rep. 2011, 7, 506–517. [Google Scholar] [CrossRef]
- Ha, S.Y.; Jee, B.C.; Suh, C.S.; Kim, H.S.; Oh, S.K.; Kim, S.H.; Moon, S.Y. Cryopreservation of human embryonic stem cells without the use of a programmable freezer. Hum. Reprod. 2005, 20, 1779–1785. [Google Scholar] [CrossRef]
- Fernandes, S.; Khan, N.; Kale, V.; Limaye, L. Catalase incorporation in freezing mixture leads to improved recovery of cryopreserved iPSC lines. Cryobiology 2019, 90, 21–29. [Google Scholar] [CrossRef]
- Schuldiner, M.; Yanuka, O.; Itskovitz-Eldor, J.; Melton, D.A.; Benvenisty, N. Effects of eight growth factors on the differentiation of cells derived from human embryonic stem cells. Proc. Natl. Acad. Sci. USA 2000, 97, 11307–11312. [Google Scholar] [CrossRef] [PubMed]
- Galvao, J.; Davis, B.; Tilley, M.; Normando, E.; Duchen, M.R.; Cordeiro, M.F. Unexpected low-dose toxicity of the universal solvent DMSO. FASEB J. 2014, 28, 1317–1330. [Google Scholar] [CrossRef] [PubMed]
- Vogel, K.G. Effects of hyaluronidase, trypsin, and EDTA on surface composition and topography during detachment of cells in culture. Exp. Cell Res. 1978, 113, 345–357. [Google Scholar] [CrossRef]
- Katkov, I.I.; Kan, N.G.; Cimadamore, F.; Nelson, B.; Snyder, E.Y.; Terskikh, A.V. DMSO-Free Programmed Cryopreservation of Fully Dissociated and Adherent Human Induced Pluripotent Stem Cells. Stem Cells Int. 2011, 2011, 981606. [Google Scholar] [CrossRef]
- Reichard, A.; Asosingh, K. Best Practices for Preparing a Single Cell Suspension from Solid Tissues for Flow Cytometry. Wiley 2018, 95, 219–226. [Google Scholar] [CrossRef]
- Li, X.; Krawetz, R.; Liu, S.; Meng, G.; Rancourt, D.E. ROCK inhibitor improves survival of cryopreserved serum/feeder-free single human embryonic stem cells. Hum. Reprod. 2009, 24, 580–589. [Google Scholar] [CrossRef]
- iPSC-Based Modeling of RAG2 Severe Combined Immunodeficiency Reveals Multiple T Cell Developmental Arrests|Elsevier Enhanced Reader. Available online: https://reader.elsevier.com/reader/sd/pii/S2213671119304497?token=0FE955BBA6552C8E047859237B924974C7D810800B8E49ABCDF49F4FEE7C1BF9152E41278849C1BFFDDCE1EB5F8CF139&originRegion=eu-west-1&originCreation=20220206205918 (accessed on 6 February 2022).
- Kibschull, M.; Mileikovsky, M.; Michael, I.P.; Lye, S.J.; Nagy, A. Human embryonic fibroblasts support single cell enzymatic expansion of human embryonic stem cells in xeno-free cultures. Stem Cell Res. 2011, 6, 70–82. [Google Scholar] [CrossRef]
- Hedges, E.C.; Topp, S.; Shaw, C.E.; Nishimura, A.L. Generation of six induced pluripotent stem cell lines from patients with amyotrophic lateral sclerosis with associated genetic mutations in either FUS or ANXA11. Stem Cell Res. 2021, 52, 102246. [Google Scholar] [CrossRef]
- Beers, J.; Linask, K.L.; Chen, J.A.; Siniscalchi, L.I.; Lin, Y.; Zheng, W.; Rao, M.; Chen, G. A cost-effective and efficient reprogramming platform for large-scale production of integration-free human induced pluripotent stem cells in chemically defined culture. Sci. Rep. 2015, 5, 11319. [Google Scholar] [CrossRef]
- Ludwig, T.E.; Bergendahl, V.; Levenstein, M.E.; Yu, J.; Probasco, M.D.; Thomson, J.A. Feeder-independent culture of human embryonic stem cells. Nat. Methods 2006, 3, 637–646. [Google Scholar] [CrossRef]
- Ludwig, T.E.; Levenstein, M.E.; Jones, J.M.; Berggren, W.T.; Mitchen, E.R.; Frane, J.L.; Crandall, L.J.; Daigh, C.A.; Conard, K.R.; Piekarczyk, M.S.; et al. Derivation of human embryonic stem cells in defined conditions. Nat. Biotechnol. 2006, 24, 185–187. [Google Scholar] [CrossRef] [PubMed]
- Castro-Viñuelas, R.; Sanjurjo-Rodríguez, C.; Piñeiro-Ramil, M.; Rodríguez-Fernández, S.; López-Baltar, I.; Fuentes-Boquete, I.; Blanco, F.J.; Díaz-Prado, S. Tips and tricks for successfully culturing and adapting human induced pluripotent stem cells. Mol. Ther.-Methods Clin. Dev. 2021, 23, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Rienzi, L.; Gracia, C.; Maggiulli, R.; Labarbera, A.R.; Kaser, D.J.; Ubaldi, F.M.; Vanderpoel, S.; Racowsky, C. Oocyte, embryo and blastocyst cryopreservation in art: Systematic review and meta-Analysis comparing slow-freezing versus vitrification to produce evidence for the development of global guidance. Hum. Reprod. Update 2017, 23, 139–155. [Google Scholar] [CrossRef] [PubMed]
- Hunt, C.J. Cryopreservation: Vitrification and Controlled Rate Cooling. Methods Mol. Biol. 2017, 1590, 41–77. [Google Scholar] [CrossRef]
- Costa, G.M.J.; Avelar, G.F.; Lacerda, S.M.S.N.; Figueiredo, A.F.A.; Tavares, A.O.; Rezende-Neto, J.V.; Martins, F.G.P.; França, L.R. Horse spermatogonial stem cell cryopreservation: Feasible protocols and potential biotechnological applications. Cell Tissue Res. 2017, 370, 489–500. [Google Scholar] [CrossRef]
- Panagiotidis, Y.; Vanderzwalmen, P.; Prapas, Y.; Kasapi, E.; Goudakou, M.; Papatheodorou, A.; Passadaki, T.; Petousis, S.; Nikolettos, N.; Veletza, S.; et al. Open versus closed vitrification of blastocysts from an oocyte-donation programme: A prospective randomized study. Reprod. Biomed. Online 2013, 26, 470–476. [Google Scholar] [CrossRef]
- Seki, S.; Mazur, P. The Dominance of Warming Rate Over Cooling Rate in the Survival of Mouse Oocytes Subjected to a Vitrification Procedure. Cryobiology 2009, 59, 75. [Google Scholar] [CrossRef]
- A review of best practices of rapid-cooling vitrification for oocytes and embryos: A committee opinion. Fertil. Steril. 2021, 115, 305–310. [CrossRef]
- Fahy, G.M.; Wowk, B.; Wu, J.; Phan, J.; Rasch, C.; Chang, A.; Zendejas, E. Cryopreservation of organs by vitrification: Perspectives and recent advances. Cryobiology 2004, 48, 157–178. [Google Scholar] [CrossRef]
- Fahy, G.M.; Wowk, B.; Wu, J.; Paynter, S. Improved vitrification solutions based on the predictability of vitrification solution toxicity. Cryobiology 2004, 48, 22–35. [Google Scholar] [CrossRef]
- Szurek, E.A.; Eroglu, A. Comparison and Avoidance of Toxicity of Penetrating Cryoprotectants. PLoS ONE 2011, 6, e27604. [Google Scholar] [CrossRef] [PubMed]
- Fowler, A.; Toner, M. Cryo-injury and biopreservation. Ann. N. Y. Acad. Sci. 2005, 1066, 119–135. [Google Scholar] [CrossRef] [PubMed]
- Boutron, P. Comparison with the theory of the kinetics and extent of ice crystallization and of the glass-forming tendency in aqueous cryoprotective solutions. Cryobiology 1986, 23, 88–102. [Google Scholar] [CrossRef]
- He, X.; Park, E.Y.H.; Fowler, A.; Yarmush, M.L.; Toner, M. Vitrification by Ultra-fast Cooling at a Low Concentration of Cryoprotectants in a Quartz Microcapillary: A Study Using Murine Embryonic Stem Cells. Cryobiology 2008, 56, 223. [Google Scholar] [CrossRef] [PubMed]
- Kaindl, J.; Meiser, I.; Majer, J.; Sommer, A.; Krach, F.; Katsen-Globa, A.; Winkler, J.; Zimmermann, H.; Neubauer, J.C.; Winner, B. Zooming in on Cryopreservation of hiPSCs and Neural Derivatives: A Dual-Center Study Using Adherent Vitrification. Stem Cells Transl. Med. 2019, 8, 247–259. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, H.; Liu, Z.; Long, P.; Zhao, X.; Li, Q.; Huang, Y.; Ma, Y. Cryopreservation of human induced pluripotent stem cells by using a new CryoLogic vitrification method. Cryobiology 2021, 98, 210–214. [Google Scholar] [CrossRef]
- Xiong, S.; Liu, J.X.; Gao, Y.; Liu, W.W.; Wu, L.H.; Han, W.; Zhang, X.D.; Han, S.B.; Liu, D.Y.; Huang, G.N. Shortened equilibration time can compromise clinical outcomes in human embryo vitrification. Hum. Fertil. 2016, 19, 114–119. [Google Scholar] [CrossRef]
- Vanderzwalmen, P.; Ectors, F.; Grobet, L.; Prapas, Y.; Panagiotidis, Y.; Vanderzwalmen, S.; Stecher, A.; Frias, P.; Liebermann, J.; Zech, N.H. Aseptic vitrification of blastocysts from infertile patients, egg donors and after IVM. Reprod. Biomed. Online 2009, 19, 700–707. [Google Scholar] [CrossRef]
- Wang, W.H.; Meng, L.; Hackett, R.J.; Odenbourg, R.; Keefe, D.L. Limited recovery of meiotic spindles in living human oocytes after cooling-rewarming observed using polarized light microscopy. Hum. Reprod. 2001, 16, 2374–2378. [Google Scholar] [CrossRef]
- Zenzes, M.T.; Bielecki, R.; Casper, R.F.; Leibo, S.P. Effects of chilling to 0 degrees C on the morphology of meiotic spindles in human metaphase II oocytes. Fertil. Steril. 2001, 75, 769–777. [Google Scholar] [CrossRef]
- Frim, J.; Mazur, P. Interactions of cooling rate, warming rate, glycerol concentration, and dilution procedure on the viability of frozen-thawed human granulocytes. Cryobiology 1983, 20, 657–676. [Google Scholar] [CrossRef]
- Baharvand, H.; Hosseini Salekdeh, G.; Taei, A.; Mollamohammadi, S. An efficient and easy-to-use cryopreservation protocol for human ES and iPS cells. Nat. Protoc. 2010, 5, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Pakzad, M.; Totonchi, M.; Taei, A.; Seifinejad, A.; Hassani, S.N.; Baharvand, H. Presence of a ROCK inhibitor in extracellular matrix supports more undifferentiated growth of feeder-free human embryonic and induced pluripotent stem cells upon passaging. Stem Cell Rev. Rep. 2010, 6, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Lam, M.T.; Longaker, M.T. Comparison of several attachment methods for human iPS, embryonic and adipose-derived stem cells for tissue engineering. J. Tissue Eng. Regen. Med. 2012, 6 (Suppl. S3), s80–s86. [Google Scholar] [CrossRef]
- Miyazaki, T.; Suemori, H. Slow Cooling Cryopreservation Optimized to Human Pluripotent Stem Cells. Adv. Exp. Med. Biol. 2016, 951, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Martin-Ibañez, R.; Unger, C.; Strömberg, A.; Baker, D.; Canals, J.M.; Hovatta, O. Novel cryopreservation method for dissociated human embryonic stem cells in the presence of a ROCK inhibitor. Hum. Reprod. 2008, 23, 2744–2754. [Google Scholar] [CrossRef] [PubMed]
- Martín-Ibáñez, R.; Strömberg, A.M.; Hovatta, O.; Canals, J.M. Cryopreservation of dissociated human embryonic stem cells in the presence of ROCK inhibitor. Chapter 1, Unit 1C.8. Curr. Protoc. Stem Cell Biol. 2009, 10, 1C.8.1–1C.8.15. [Google Scholar] [CrossRef] [PubMed]
- Hunt, C.J. Technical Considerations in the Freezing, Low-Temperature Storage and Thawing of Stem Cells for Cellular Therapies. Transfus. Med. Hemother. 2019, 46, 134–149. [Google Scholar] [CrossRef]
- Curry, M.R.; Watson, P.F. Osmotic effects on ram and human sperm membranes in relation to thawing injury. Cryobiology 1994, 31, 39–46. [Google Scholar] [CrossRef]
- Culture of Established Induced Pluripotent Stem Cell Lines. Available online: https://www.protocols.io/view/culture-of-established-induced-pluripotent-stem-ce-bgbwjspe (accessed on 6 February 2022).
- Jose, S.S.; Tidu, F.; Burilova, P.; Kepak, T.; Bendickova, K.; Fric, J. The Telomerase Complex Directly Controls Hematopoietic Stem Cell Differentiation and Senescence in an Induced Pluripotent Stem Cell Model of Telomeropathy. Front. Genet. 2018, 9, 345. [Google Scholar] [CrossRef]
- Zeiger, A.S.; Hinton, B.; Van Vliet, K.J. Why the dish makes a difference: Quantitative comparison of polystyrene culture surfaces. Acta Biomater. 2013, 9, 7354–7361. [Google Scholar] [CrossRef] [PubMed]
- Strober, W. Trypan blue exclusion test of cell viability. Appendix 3. Curr. Protoc. Immunol. 2001. [Google Scholar] [CrossRef]
- Chan, L.L.Y.; Rice, W.L.; Qiu, J. Observation and quantification of the morphological effect of trypan blue rupturing dead or dying cells. PLoS ONE 2020, 15, e0227950. [Google Scholar] [CrossRef] [PubMed]
- Moll, G.; Alm, J.J.; Davies, L.C.; Von Bahr, L.; Heldring, N.; Stenbeck-Funke, L.; Hamad, O.A.; Hinsch, R.; Ignatowicz, L.; Locke, M.; et al. Do cryopreserved mesenchymal stromal cells display impaired immunomodulatory and therapeutic properties? Stem Cells 2014, 32, 2430–2442. [Google Scholar] [CrossRef] [PubMed]
- Kovacic, B.; Rosner, M.; Schlangen, K.; Kramer, N.; Hengstschläger, M. DRUGPATH-a novel bioinformatic approach identifies DNA-damage pathway as a regulator of size maintenance in human ESCs and iPSCs. Sci. Rep. 2019, 9, 1897. [Google Scholar] [CrossRef] [PubMed]
- Emmenegger, M.; De Cecco, E.; Hruska-Plochan, M.; Eninger, T.; Schneider, M.M.; Barth, M.; Tantardini, E.; de Rossi, P.; Bacioglu, M.; Langston, R.G.; et al. LAG3 is not expressed in human and murine neurons and does not modulate α-synucleinopathies. EMBO Mol. Med. 2021, 13, e14745. [Google Scholar] [CrossRef]
- Grässer, U.; Bubel, M.; Sossong, D.; Oberringer, M.; Pohlemann, T.; Metzger, W. Dissociation of mono- and co-culture spheroids into single cells for subsequent flow cytometric analysis. Ann. Anat.-Anat. Anzeiger 2018, 216, 1–8. [Google Scholar] [CrossRef]
- Freedman, B.S.; Keeler, E.G.; Lin, L.Y.; Cruz, N.M.; Jing, P.; Liu, Y. Optical tweezers system for live stem cell organization at the single-cell level. Biomed. Opt. Express 2018, 9, 771–779. [Google Scholar] [CrossRef]
- Wei, C.; Lu, X.; Zou, Q.; Gong, S.; Chen, Y.; Lyu, J.; Jia, J. Generation and characterization of a human induced pluripotent stem cell line (XWHNi001-A) derived from an Alzheimer’s disease patient with mutation in the APP gene. Stem Cell Res. 2022, 60, 102690. [Google Scholar] [CrossRef]
- Li, X.; Meng, G.; Krawetz, R.; Liu, S.; Rancourt, D.E. The ROCK inhibitor Y-27632 enhances the survival rate of human embryonic stem cells following cryopreservation. Stem Cells Dev. 2008, 17, 1079–1085. [Google Scholar] [CrossRef]
- Mollamohammadi, S.; Taei, A.; Pakzad, M.; Totonchi, M.; Seifinejad, A.; Masoudi, N.; Baharvand, H. A simple and efficient cryopreservation method for feeder-free dissociated human induced pluripotent stem cells and human embryonic stem cells. Hum. Reprod. 2009, 24, 2468–2476. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhang, P.; Wu, X.; Hu, C.; Wang, P.; Li, X. Rho kinase inhibitor Y-27632 and Accutase dramatically increase mouse embryonic stem cell derivation. Vitr. Cell. Dev. Biol. Anim. 2012, 48, 30–36. [Google Scholar] [CrossRef]
- Maldonado, M.; Luu, R.J.; Ramos, M.E.P.; Nam, J. ROCK inhibitor primes human induced pluripotent stem cells to selectively differentiate towards mesendodermal lineage via epithelial-mesenchymal transition-like modulation. Stem Cell Res. 2016, 17, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Effect of Y-27632 on CCE Colony Formation and Morphological Changes A...|Download Scientific Diagram. Available online: https://www.researchgate.net/figure/Effect-of-Y-27632-on-CCE-colony-formation-and-morphological-changes-A-B-Cells-in_fig9_41487781 (accessed on 10 February 2022).
- Xu, Y.; Ding, W.; Li, S.; Li, C.; Gao, D.; Qiu, B. A single-cell identification and capture chip for automatically and rapidly determining hydraulic permeability of cells. Anal. Bioanal. Chem. 2020, 412, 4537–4548. [Google Scholar] [CrossRef] [PubMed]
- Roffay, C.; Molinard, G.; Kim, K.; Urbanska, M.; Andrade, V.; Barbarasa, V.; Nowak, P.; Mercier, V.; García-Calvo, J.; Matile, S.; et al. Passive coupling of membrane tension and cell volume during active response of cells to osmosis. Proc. Natl. Acad. Sci. USA 2021, 118, e2103228118. [Google Scholar] [CrossRef]
- DeJarnette, J.M.; Barnes, D.A.; Marshall, C.E. Effects of pre-and post-thaw thermal insults on viability characteristics of cryopreserved bovine semen. Theriogenology 2000, 53, 1225–1238. [Google Scholar] [CrossRef]
- Correa, J.R.; Heersche, G.; Zavos, P.M. Sperm membrane functional integrity and response of frozen-thawed bovine spermatozoa during the hypoosmotic swelling test incubation at varying temperatures. Theriogenology 1997, 47, 715–721. [Google Scholar] [CrossRef]
- Correa, J.R.; Zavos, P.M. Frozen-thawed bovine spermatozoa diluted by slow or rapid dilution method: Measurements on occurrence of osmotic shock and sperm viability. Theriogenology 1995, 44, 963–971. [Google Scholar] [CrossRef]
- Correa, J.R.; Rodriguez, M.C.; Patterson, D.J.; Zavos, P.M. Thawing and processing of cryopreserved bovine spermatozoa at various temperatures and their effects on sperm viability, osmotic shock and sperm membrane functional integrity. Theriogenology 1996, 46, 413–420. [Google Scholar] [CrossRef]
- Chen, C.X.Q.; Abdian, N.; Maussion, G.; Thomas, R.A.; Demirova, I.; Cai, E.; Tabatabaei, M.; Beitel, L.K.; Karamchandani, J.; Fon, E.A.; et al. A multistep workflow to evaluate newly generated ipscs and their ability to generate different cell types. Methods Protoc. 2021, 4, 50. [Google Scholar] [CrossRef]
- Brenière-Letuffe, D.; Domke-Shibamiya, A.; Hansen, A.; Eschenhagen, T.; Fehse, B.; Riecken, K.; Stenzig, J. Clonal dynamics studied in cultured induced pluripotent stem cells reveal major growth imbalances within a few weeks. Stem Cell Res. Ther. 2018, 9, 165. [Google Scholar] [CrossRef] [PubMed]
 clone 1,
clone 1,  clone 2,
clone 2,  clone 3,
clone 3,  compromised clone.
clone 1, clone 2, clone 3, compromised clone.
compromised clone.
clone 1, clone 2, clone 3, compromised clone.

{kind=link}
{kind=link}
| Factors | Not Optimized | Optimized | Critical Steps | References |
|---|---|---|---|---|
| Cell growth phase | Cells have already entered the stationary phase. | Freeze cells during the log growth phase, approx. 2–4 days after passaging. | With the goal of reaching a sufficiently high cell number (high confluency), it may happen that the cells are unintentionally grown for too many days and thus have already entered the stationary phase. | [34,35,36] |
| Cell number | Cell number too low. | Let cells grow up to approx. 70–80% confluency. Freeze iPSC from one well of a six-well plate in one ml of freezing solution (one cryovial). | While the iPSC number is growing, make sure that the cells are still within the log growth phase upon freezing. If necessary, whenever cells have already entered the stationary phase, split 1:2–1:4 and freeze 2–4 days later. | [36,37] |
| Cell aggregate size | Cell aggregates are disrupted, resulting in single cells or cell aggregates that are too small. | Avoid harsh pipetting. Frequently, a cell aggregate size of 50–200 µm is recommended. | From harvesting until the final steps of freezing, the cell aggregates can be disrupted in many of these steps. Therefore, all steps should be carried out gently *. | [37], STEMCELL Technologies website |
| Differentiated cells in the iPSC culture | Too many spontaneously differentiated cells appear near or within the iPSC colonies. | Remove differentiated cells manually under the microscope or by short incubation times with an EDTA-based dissociation reagent on a regular basis and directly before freezing. | The number of differentiated cells can vary from clone to clone. iPSC clones which very rapidly differentiate spontaneously should be discarded. Those with an acceptable (low) number of differentiated cells should be manually cleaned on a regular basis (differentiated cells should be removed under a microscope before freezing). Some lots of FBS (e.g., not ESC-qualified FBS) possibly induce differentiation; thus, it may be considered to withdraw FBS after thawing (provided that the freezing solution contained FBS). | [33,38,39,40] |
| Factors | Not Optimized | Optimized | Critical Steps | References |
|---|---|---|---|---|
| Coating | Wrong or expired coating substance. | Use good quality Matrigel™ or another suitable coating substance. | Thaw and aliquot Matrigel™ according to SOP on ice *. | [87], Corning and BD Biosciences websites. |
| Cell number for seeding | Cell number too low, resulting in loss of cell-cell contacts. | Assure high cell density upon seeding. | Try to reach a high cell density **. If necessary, by seeding the given number of cells (e.g., thawed cells from one cryovial) onto a smaller surface (smaller well). | [37,88] |
| ROCK inhibitor | ROCK inhibitor is missing in the medium. | Add 10 µM ROCK inhibitor. | ROCK inhibitor is helpful for cell attachment and survival, especially if other factors are not optimized (e.g., disrupted cell aggregates, low cell seeding density). If cells are completely singularized, adding ROCK inhibitor is very important for cell survival. | [31,32,37,42,89,90] |
| Cell aggregate size | Cell aggregates are disrupted, resulting in single cells or cell aggregates that are too small. | Avoid harsh pipetting. Use ROCK inhibitor, especially if aggregates are significantly smaller than approx. 50 µm. | During the whole thawing and seeding process, the cell aggregates can be disrupted. Therefore, all steps should be carried out gently. *** | [37], STEMCELL Technologies website |
| Osmotic shock | The medium is first poured into a 15 mL conical tube, then all the thawed cells from the cryovial are added suddenly and at once to the medium in the tube. | Put the thawed cells in a 15 mL conical tube first, then add medium slowly in a dropwise manner. | A sudden change in the osmolarity of the freezing solution around the cells may cause a rapid stream of water across the membranes of the cells. This may stress the cells, making them more prone to die. Avoiding this kind of stress can contribute to better cell survival. | [91,92], European Bank for induced pluripotent stem cells (EBiSC) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uhrig, M.; Ezquer, F.; Ezquer, M. Improving Cell Recovery: Freezing and Thawing Optimization of Induced Pluripotent Stem Cells. Cells 2022, 11, 799. https://doi.org/10.3390/cells11050799
Uhrig M, Ezquer F, Ezquer M. Improving Cell Recovery: Freezing and Thawing Optimization of Induced Pluripotent Stem Cells. Cells. 2022; 11(5):799. https://doi.org/10.3390/cells11050799
Chicago/Turabian StyleUhrig, Markus, Fernando Ezquer, and Marcelo Ezquer. 2022. "Improving Cell Recovery: Freezing and Thawing Optimization of Induced Pluripotent Stem Cells" Cells 11, no. 5: 799. https://doi.org/10.3390/cells11050799
APA StyleUhrig, M., Ezquer, F., & Ezquer, M. (2022). Improving Cell Recovery: Freezing and Thawing Optimization of Induced Pluripotent Stem Cells. Cells, 11(5), 799. https://doi.org/10.3390/cells11050799