
Post-mortem Findings of Inflammatory Cells and the Association of 4-Hydroxynonenal with Systemic Vascular and Oxidative Stress in Lethal COVID-19 †

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Immunohistochemistry
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Roche, L.; Mesta, F. Oxidative Stress as Key Player in Severe Acute Respiratory Syndrome Coronavirus (SARS-CoV) Infection. Arch. Med. Res. 2020, 51, 384–387. [Google Scholar] [CrossRef] [PubMed]
- LaForge, M.; Elbim, C.; Frère, C.; Hémadi, M.; Massaad, C.; Nuss, P.; Benoliel, J.-J.; Becker, C. Tissue damage from neutrophil-induced oxidative stress in COVID-19. Nat. Rev. Immunol. 2020, 20, 515–516. [Google Scholar] [CrossRef] [PubMed]
- Jaganjac, M.; Čačev, T.; Čipak, A.; Kapitanovic, S.; Trošelj, K.G.; Zarković, N. Even stressed cells are individuals: Second messengers of free radicals in pathophysiology of cancer. Croat. Med. J. 2012, 53, 304–309. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Expósito, M.J.; Martinez-Martos, M.J.R.-E.A.J.M. The Delicate Equilibrium between Oxidants and Antioxidants in Brain Glioma. Curr. Neuropharmacol. 2019, 17, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Sharif-Askari, N.S.; Sharif-Askari, F.S.; Mdkhana, B.; Alsayed, H.A.H.; Alsafar, H.; Alrais, Z.F.; Hamid, Q.; Halwani, R. Upregulation of oxidative stress gene markers during SARS-COV-2 viral infection. Free Radic. Biol. Med. 2021, 172, 688–698. [Google Scholar] [CrossRef]
- Kim, D.-H.; Ahn, H.-S.; Go, H.-J.; Kim, D.-Y.; Kim, J.-H.; Lee, J.-B.; Park, S.-Y.; Song, C.-S.; Lee, S.-W.; Ha, S.-D.; et al. Hemin as a novel candidate for treating COVID-19 via heme oxygenase-1 induction. Sci. Rep. 2021, 11, 21462. [Google Scholar] [CrossRef]
- Su, W.-L.; Lin, C.-P.; Hang, H.-C.; Wu, P.-S.; Cheng, C.-F.; Chao, Y.-C. Desaturation and heme elevation during COVID-19 infection: A potential prognostic factor of heme oxygenase-1. J. Microbiol. Immunol. Infect. 2021, 54, 113–116. [Google Scholar] [CrossRef]
- Baier, M.J.; Wagner, S.; Hupf, J.; Evert, K.; Evert, M.; Sossalla, S.; Jungbauer, C.; Maier, L.S.; Neef, S.; Mustroph, J. Cardiac iron overload promotes cardiac injury in patients with severe COVID-19. Infection 2021, 11, 1–6. [Google Scholar] [CrossRef]
- Morrissey, S.M.; Geller, A.E.; Hu, X.; Tieri, D.; Ding, C.; Klaes, C.K.; Cooke, E.A.; Woeste, M.R.; Martin, Z.C.; Chen, O.; et al. A specific low-density neutrophil population correlates with hypercoagulation and disease severity in hospitalized COVID-19 patients. JCI Insight 2021, 6. [Google Scholar] [CrossRef]
- Abais, J.M.; Xia, M.; Zhang, Y.; Boini, K.M.; Li, P.-L. Redox Regulation of NLRP3 Inflammasomes: ROS as Trigger or Effector? Antioxid. Redox Signal 2015, 22, 1111–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, R.; Yazdi, A.S.; Menu, P.; Tschopp, J. A role for mitochondria in NLRP3 inflammasome activation. Nature 2010, 469, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, T.S.; de Sá, K.S.; Ishimoto, A.Y.; Becerra, A.; Oliveira, S.; Almeida, L.; Gonçalves, A.V.; Perucello, D.B.; Andrade, W.A.; Castro, R.; et al. Inflammasomes are activated in response to SARS-CoV-2 infection and are associated with COVID-19 severity in patients. J. Exp. Med. 2021, 218, e20201707. [Google Scholar] [CrossRef]
- Pan, P.; Shen, M.; Yu, Z.; Ge, W.; Chen, K.; Tian, M.; Xiao, F.; Wang, Z.; Wang, J.; Jia, Y.; et al. SARS-CoV-2 N protein promotes NLRP3 inflammasome activation to induce hyperinflammation. Nat. Commun. 2021, 12, 4664. [Google Scholar] [CrossRef]
- Chen, R.; Lan, Z.; Ye, J.; Pang, L.; Liu, Y.; Wu, W.; Qin, X.; Guo, Y.; Zhang, P. Cytokine Storm: The Primary Determinant for the Pathophysiological Evolution of COVID-19 Deterioration. Front. Immunol. 2021, 12, 589095. [Google Scholar] [CrossRef]
- Žarković, N.; Orehovec, B.; Milković, L.; Baršić, B.; Tatzber, F.; Wonisch, W.; Tarle, M.; Kmet, M.; Mataić, A.; Jakovčević, A.; et al. Preliminary Findings on the Association of the Lipid Peroxidation Product 4-Hydroxynonenal with the Lethal Outcome of Aggressive COVID-19. Antioxidants 2021, 10, 1341. [Google Scholar] [CrossRef] [PubMed]
- Jaganjac, M.; Cindrić, M.; Jakovčević, A.; Žarković, K.; Žarković, N. Lipid peroxidation in brain tumors. Neurochem. Int. 2021, 149, 105118. [Google Scholar] [CrossRef] [PubMed]
- Jaganjac, M.; Milkovic, L.; Gęgotek, A.; Cindric, M.; Zarkovic, K.; Skrzydlewska, E.; Zarkovic, N. The relevance of pathophysiological alterations in redox signaling of 4-hydroxynonenal for pharmacological therapies of major stress-associated diseases. Free Radic. Biol. Med. 2020, 157, 128–153. [Google Scholar] [CrossRef]
- Kauppinen, A.; Niskanen, H.; Suuronen, T.; Kinnunen, K.; Salminen, A.; Kaarniranta, K. Oxidative stress activates NLRP3 inflammasomes in ARPE-19 cells—Implications for age-related macular degeneration (AMD). Immunol. Lett. 2012, 147, 29–33. [Google Scholar] [CrossRef]
- Jacobs, W.; Lammens, M.; Kerckhofs, A.; Voets, E.; Van San, E.; Van Coillie, S.; Peleman, C.; Mergeay, M.; Sirimsi, S.; Matheeussen, V.; et al. Fatal lymphocytic cardiac damage in coronavirus disease 2019 (COVID-19): Autopsy reveals a ferroptosis signature. ESC Heart Fail. 2020, 7, 3772–3781. [Google Scholar] [CrossRef]
- Iles, K.E.; Dickinson, D.A.; Wigley, A.F.; Welty, N.E.; Blank, V.; Forman, H.J. HNE increases HO-1 through activation of the ERK pathway in pulmonary epithelial cells. Free Radic. Biol. Med. 2005, 39, 355–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elrayess, M.; Almuraikhy, S.; Kafienah, W.; Al-Menhali, A.; Al-Khelaifi, F.; Bashah, M.; Zarkovic, K.; Zarkovic, N.; Waeg, G.; Alsayrafi, M.; et al. 4-hydroxynonenal causes impairment of human subcutaneous adipogenesis and induction of adipocyte insulin resistance. Free Radic. Biol. Med. 2017, 104, 129–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarkovic, K.; Jakovcevic, A.; Zarkovic, N. Contribution of the HNE-immunohistochemistry to modern pathological concepts of major human diseases. Free. Radic. Biol. Med. 2017, 111, 110–126. [Google Scholar] [CrossRef] [PubMed]
- Pomara, C.; Salerno, M.; Sessa, F.; Esposito, M.; Barchitta, M.; Ledda, C.; Grassi, P.; Liberto, A.; Mattaliano, A.; Rapisarda, V.; et al. Safe Management Strategies in Clinical Forensic Autopsies of Confirmed COVID-19 Cases. Diagnostics 2021, 11, 457. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Collection and Submission of Postmortem Specimens from Deceased Persons with Confirmed or Suspected COVID-19. Available online: https://www.cdc.gov/coronavirus/2019-ncov/hcp/guidance-postmortem-specimens.html (accessed on 20 December 2020).
- World Health Organization. Infection Prevention and Control for the Safe Management of a Dead Body in the Context of COVID-19: Interim Guidance. Available online: https://www.who.int/publications/i/item/infection-prevention-and-control-for-the-safe-management-of-a-dead-body-in-the-context-of-covid-19-interim-guidance (accessed on 18 January 2022).
- Musso, N.; Falzone, L.; Stracquadanio, S.; Bongiorno, D.; Salerno, M.; Esposito, M.; Sessa, F.; Libra, M.; Stefani, S.; Pomara, C. Post-Mortem Detection of SARS-CoV-2 RNA in Long-Buried Lung Samples. Diagnostics 2021, 11, 1158. [Google Scholar] [CrossRef]
- Pomara, C.; Sessa, F.; Galante, D.; Pace, L.; Fasanella, A.; Di Nunno, N.; Esposito, M.; Salerno, M. Do We Really Need Hazard Prevention at the Expense of Safeguarding Death Dignity in COVID-19? Diagnostics 2021, 11, 1913. [Google Scholar] [CrossRef]
- Zarkovic, K.; Juric, G.; Waeg, G.; Kolenc, D.; Zarkovic, N. Immunohistochemical appearance of HNE-protein conjugates in human astrocytomas. BioFactors 2005, 24, 33–40. [Google Scholar] [CrossRef]
- Cherkas, A.; Zarkovic, K.; Gasparovic, A.C.; Jaganjac, M.; Milkovic, L.; Abrahamovych, O.; Yatskevych, O.; Waeg, G.; Yelisyeyeva, O.; Zarkovic, N. Amaranth oil reduces accumulation of 4-hydroxynonenal-histidine adducts in gastric mucosa and improves heart rate variability in duodenal peptic ulcer patients undergoing Helicobacter pylori eradication. Free Radic. Res. 2018, 52, 135–149. [Google Scholar] [CrossRef]
- Jakovčević, A.; Žarković, K.; Jakovčević, D.; Rakušić, Z.; Prgomet, D.; Waeg, G.; Šunjić, S.B.; Žarković, N. The Appearance of 4-Hydroxy-2-Nonenal (HNE) in Squamous Cell Carcinoma of the Oropharynx. Molecules 2020, 25, 868. [Google Scholar] [CrossRef] [Green Version]
- Henkel, M.; Weikert, T.; Marston, K.; Schwab, N.; Sommer, G.; Haslbauer, J.; Franzeck, F.; Anastasopoulos, C.; Stieltjes, B.; Michel, A.; et al. Lethal COVID-19: Radiologic-Pathologic Correlation of the Lungs. Radiol. Cardiothorac. Imaging 2020, 2, e200406. [Google Scholar] [CrossRef]
- Libby, P.; Lüscher, T. COVID-19 is, in the end, an endothelial disease. Eur. Heart J. 2020, 41, 3038–3044. [Google Scholar] [CrossRef] [PubMed]
- Varga, Z.; Flammer, A.J.; Steiger, P.; Haberecker, M.; Andermatt, R.; Zinkernagel, A.S.; Mehra, M.R.; Schuepbach, R.; Ruschitzka, F.; Moch, H. Endothelial cell infection and endotheliitis in COVID-19. Lancet 2020, 395, 1417–1418. [Google Scholar] [CrossRef]
- Thacker, V.V.; Sharma, K.; Dhar, N.; Mancini, G.; Sordet-Dessimoz, J.; McKinney, J.D. Rapid endotheliitis and vascular damage characterize SARS-CoV-2 infection in a human lung-on-chip model. EMBO Rep. 2021, 22, e52744. [Google Scholar] [CrossRef]
- Cui, X.; Chen, W.; Zhou, H.; Gong, Y.; Zhu, B.; Lv, X.; Guo, H.; Duan, J.; Zhou, J.; Marcon, E.; et al. Pulmonary Edema in COVID-19 Patients: Mechanisms and Treatment Potential. Front. Pharmacol. 2021, 12, 664349. [Google Scholar] [CrossRef] [PubMed]
- Zwaveling, S.; van Wijk, R.G.; Karim, F. Pulmonary edema in COVID-19: Explained by bradykinin? J. Allergy Clin. Immunol. 2020, 146, 1454–1455. [Google Scholar] [CrossRef]
- Zarkovic, K.; Larroque-Cardoso, P.; Pucelle, M.; Salvayre, R.; Waeg, G.; Negre-Salvayre, A.; Zarkovic, N. Elastin aging and lipid oxidation products in human aorta. Redox Biol. 2015, 4, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Jaganjac, M.; Almuraikhy, S.; Al-Khelaifi, F.; Al-Jaber, M.; Bashah, M.; Mazloum, N.A.; Zarkovic, K.; Zarkovic, N.; Waeg, G.; Kafienah, W.; et al. Combined metformin and insulin treatment reverses metabolically impaired omental adipogenesis and accumulation of 4-hydroxynonenal in obese diabetic patients. Redox Biol. 2017, 12, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Riahi, Y.; Kaiser, N.; Cohen, G.; Abd-Elrahman, I.; Blum, G.; Shapira, O.M.; Koler, T.; Simionescu, M.; Sima, A.V.; Zarkovic, N.; et al. Foam cell-derived 4-hydroxynonenal induces endothelial cell senescence in a TXNIP-dependent manner. J. Cell. Mol. Med. 2015, 19, 1887–1899. [Google Scholar] [CrossRef] [Green Version]
- Abu-Farha, M.; Thanaraj, T.A.; Qaddoumi, M.G.; Hashem, A.; Abubaker, J.; Al-Mulla, F. The Role of Lipid Metabolism in COVID-19 Virus Infection and as a Drug Target. Int. J. Mol. Sci. 2020, 21, 3544. [Google Scholar] [CrossRef]
- Vlasov, I.; Panteleeva, A.; Usenko, T.; Nikolaev, M.; Izumchenko, A.; Gavrilova, E.; Shlyk, I.; Miroshnikova, V.; Shadrina, M.; Polushin, Y.; et al. Transcriptomic Profiles Reveal Downregulation of Low-Density Lipoprotein Particle Receptor Pathway Activity in Patients Surviving Severe COVID-19. Cells 2021, 10, 3495. [Google Scholar] [CrossRef]
- Archambault, A.-S.; Zaid, Y.; Rakotoarivelo, V.; Doré, É.; Dubuc, I.; Martin, C.; Amar, Y.; Cheikh, A.; Fares, H.; El Hassani, A.; et al. Lipid storm within the lungs of severe COVID-19 patients: Extensive levels of cyclooxygenase and lipoxygenase-derived inflammatory metabolites. medRxiv 2020. [Google Scholar] [CrossRef]
- Wakita, C.; Honda, K.; Shibata, T.; Akagawa, M.; Uchida, K. A method for detection of 4-hydroxy-2-nonenal adducts in proteins. Free. Radic. Biol. Med. 2011, 51, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.; Milkovic, L.; Bennett, S.J.; Griffiths, H.R.; Zarkovic, N.; Grune, T. Measurement of HNE-protein adducts in human plasma and serum by ELISA—Comparison of two primary antibodies. Redox Biol. 2013, 1, 226–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binder, C.J.; Papac-Milicevic, N.; Witztum, J.L. Innate sensing of oxidation-specific epitopes in health and disease. Nat. Rev. Immunol. 2016, 16, 485–497. [Google Scholar] [CrossRef]
- Leibundgut, G.; Witztum, J.L.; Tsimikas, S. Oxidation-specific epitopes and immunological responses: Translational biotheranostic implications for atherosclerosis. Curr. Opin. Pharmacol. 2013, 13, 168–179. [Google Scholar] [CrossRef] [Green Version]
- Miller, Y.I.; Choi, S.-H.; Wiesner, P.; Fang, L.; Harkewicz, R.; Hartvigsen, K.; Boullier, A.; Gonen, A.; Diehl, C.; Que, X.; et al. Oxidation-Specific Epitopes Are Danger-Associated Molecular Patterns Recognized by Pattern Recognition Receptors of Innate Immunity. Circ. Res. 2011, 108, 235–248. [Google Scholar] [CrossRef] [Green Version]
- Zhivaki, D.; Kagan, J.C. Innate immune detection of lipid oxidation as a threat assessment strategy. Nat. Rev. Immunol. 2021, 2021, 1–9. [Google Scholar] [CrossRef]
- Macdonald, J.; Galley, H.; Webster, N. Oxidative stress and gene expression in sepsis. Br. J. Anaesth. 2003, 90, 221–232. [Google Scholar] [CrossRef] [Green Version]
- Mantzarlis, K.; Tsolaki, V.; Zakynthinos, E. Role of Oxidative Stress and Mitochondrial Dysfunction in Sepsis and Potential Therapies. Oxidative Med. Cell. Longev. 2017, 2017, 1–10. [Google Scholar] [CrossRef]
- Colombo, D.; Falasca, L.; Marchioni, L.; Tammaro, A.; Adebanjo, G.A.R.; Ippolito, G.; Zumla, A.; Piacentini, M.; Nardacci, R.; Del Nonno, F. Neuropathology and Inflammatory Cell Characterization in 10 Autoptic COVID-19 Brains. Cells 2021, 10, 2262. [Google Scholar] [CrossRef]
- Hafezi, B.; Chan, L.; Knapp, J.; Karimi, N.; Alizadeh, K.; Mehrani, Y.; Bridle, B.; Karimi, K. Cytokine Storm Syndrome in SARS-CoV-2 Infections: A Functional Role of Mast Cells. Cells 2021, 10, 1761. [Google Scholar] [CrossRef] [PubMed]
- Wong, D.; Klinkhammer, B.; Djudjaj, S.; Villwock, S.; Timm, M.; Buhl, E.; Wucherpfennig, S.; Cacchi, C.; Braunschweig, T.; Knüchel-Clarke, R.; et al. Multisystemic Cellular Tropism of SARS-CoV-2 in Autopsies of COVID-19 Patients. Cells 2021, 10, 1900. [Google Scholar] [CrossRef] [PubMed]
- Forcados, G.E.; Muhammad, A.; Oladipo, O.O.; Makama, S.; Meseko, C.A. Metabolic Implications of Oxidative Stress and Inflammatory Process in SARS-CoV-2 Pathogenesis: Therapeutic Potential of Natural Antioxidants. Front. Cell. Infect. Microbiol. 2021, 11, 654813. [Google Scholar] [CrossRef] [PubMed]
- Suhail, S.; Zajac, J.; Fossum, C.; Lowater, H.; McCracken, C.; Severson, N.; Laatsch, B.; Narkiewicz-Jodko, A.; Johnson, B.; Liebau, J.; et al. Role of Oxidative Stress on SARS-CoV (SARS) and SARS-CoV-2 (COVID-19) Infection: A Review. J. Protein Chem. 2020, 39, 644–656. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | Sex | Comorbidity | Diagnoses | Organ | Tissue Findings | Inflammatory Cells * | Bleeding | Thrombosis | Edema |
|---|---|---|---|---|---|---|---|---|---|
| 58 | M | DM2 1 | ARDS 2 | lungs | DAD 3, interstitial pneumonia | Ly, Pl, Ma, PMN 4 | no | yes | yes |
| Hypertension | pulmonary emboly pneumonia interstitialis | heart | myocardial fibrosis atherosclerosis | PMN | no | no | yes | ||
| Steatosis hepatis | atherosclerosis | ||||||||
| dilatatio cordis | brain | brain edema | PMN, Mo | yes | no | yes | |||
| 54 | M | Obesity Hypertension | ARDS (DAD) | lungs | DAD, inerstitial pneumonia, DIC | Mo, PMN | yes | no | yes |
| pneumonia | |||||||||
| DIC 5 | brain | edema, DIC | PMN, Mo | yes | yes | yes | |||
| myocardial fibrosis hypertrophia and dilatatio cordis | heart | myocardial fibrosis | Mo | no | no | no | |||
| steatosis hepatis renal ATN 7 | liver | steatosis hepatis | Mo | no | no | no | |||
| 55 | M | Hypertension | (sepsis staphylococcica) ARDS (DAD) | lungs | DAD, pneumonia abscedens | PMN | no | no | yes |
| pneumonia abscedens | brain | brain edema | Ly. PMN, Mo | no | no | yes | |||
| steatosis hepatis | liver | steatosis hepatis | Ly, PMN | no | no | no | |||
| atherosclerosis heart dilatation | heart | myocardial fibrosis | PMN | no | no | no | |||
| 57 6 | M | (sepsis A. baumanii), ARDS (DAD) | lungs | pneumonia chronica and suppurativa, DAD | Ma, Mo, Ly, Pl, PMN | yes | yes | yes | |
| pneumonia | kidney | renal ATN | PMN | no | no | no | |||
| renal ATN | heart | myocardial fibrosis and lipomatosis | Ly | no | no | no | |||
| myocardial fibrosis | liver | centrilobular cyanosis | Ly | no | no | no | |||
| brain | brain edema | Ly, Mo | no | no | no | ||||
| 65 | M | DM2 | bilateral pneumonia | heart | myocardial fibrosis | Ly, Mo, PMN | no | no | no |
| Hypertension Hyperlipidemia | pulmonary microthrombosis | lungs | lung fibrosis, Chronic DAD, acute hemorrhage | Ma, PMN | yes | yes | yes | ||
| Atherosclerosis | atherosclerosis | kidney | renal ATN | PMN | no | no | no | ||
| IBD 8 | heart hypertrophia | liver | steatosis hepatis | Ly, PMN | no | no | no | ||
| myocardial fibrosis | heart | myocardial fibrosis | PMN | no | no | no | |||
| brain | brain edema | Ly | no | no | no | ||||
| 11 | M | MIS 9, ARDS (DAD) | lungs | ARDS (DAD) | Mo, Ly | yes | no | yes | |
| myocarditis encephalitis | heart | myocarditis | |||||||
| edema cerebri | brain | encephalitis, edema | PMN, Ly, Ma | no | no | no | |||
| MOF 10, renal ATN | kidney | renal ATN | PMN | no | no | no | |||
| centrilobular liver necrosis | liver | autolysis | PMN, Mo | no | no | yes | |||
| 74 | F | DM2 | ARDS | ||||||
| Hypertension | steatosis hepatis myocardial fibrosis | liver | hepatic centrilobular necrosis | Ly | no | no | no | ||
| heart dilatation | |||||||||
| pneumonia | lungs | DAD, interstitial pneumonia | PMN, Ma | yes | no | yes | |||
| renal ATN | kidney | renal ATN | PMN | yes | no | yes | |||
| myocardial fibrosis | heart | myocarditis | PMN, Ly | no | no | no | |||
| 65 6 | M | Atherosclerosis | ARDS (DAD) pneumonia | lungs | DAD, pneumonia suppurativa, | no | yes | yes | |
| Hypertension | hematoma intracerebrale | brain | acute bleeding and brain edema | PMN, Ly | yes | no | yes | ||
| Obesity | hematocephalus | ||||||||
| myocardial fibrosis | heart | myocardial fibrosis | PMN | no | no | no | |||
| 75 | M | Atherosclerosis | ARDS (DAD) | ||||||
| interstitial pneumonia | lungs | pneumonia, DAD | PMN, Mo, Ly | no | no | yes | |||
| myocardial fibrosis, heart dilatation and hypertrophy | heart | myocardial fibrosis | PMN, Ly | no | no | no | |||
| renal ATN | kidney | renal ATN | PMN | no | no | no |
| Organ Structure/Tissue Appearance | Inflammatory Cells | Immunohistochemistry for 4-HNE Protein Adducts | SOD2 Immunohistochemistry | |
|---|---|---|---|---|
| Lungs | Edematous Diffuse alveolar damage Acute and chronic interstitial pneumonia | All types of leukocytes—mostly PMNs within blood vessels, otherwise mostly pulmonary macrophages and T lymphocytes | Very strong positivity of the blood vessels, pneumocytes, hyaline membranes, and edematous liquid Some inflammatory cells positive, mostly PMNs, alveolar macrophages positive | Observed in the smooth muscle cells of the blood vessel walls and less common in the endothelium and vascular contents Prominent in alveolar macrophages in interstitial pneumonia and DAD PMNs negative |
| Heart | Often fibrotic Occasional myocarditis | T lymphocytes around blood vessels and in edematous endomysia PMNs in the blood vessels | Strong positivity of blood vessels Positive cardiac muscle fibers Positive inflammatory cells | Positive muscle fibers |
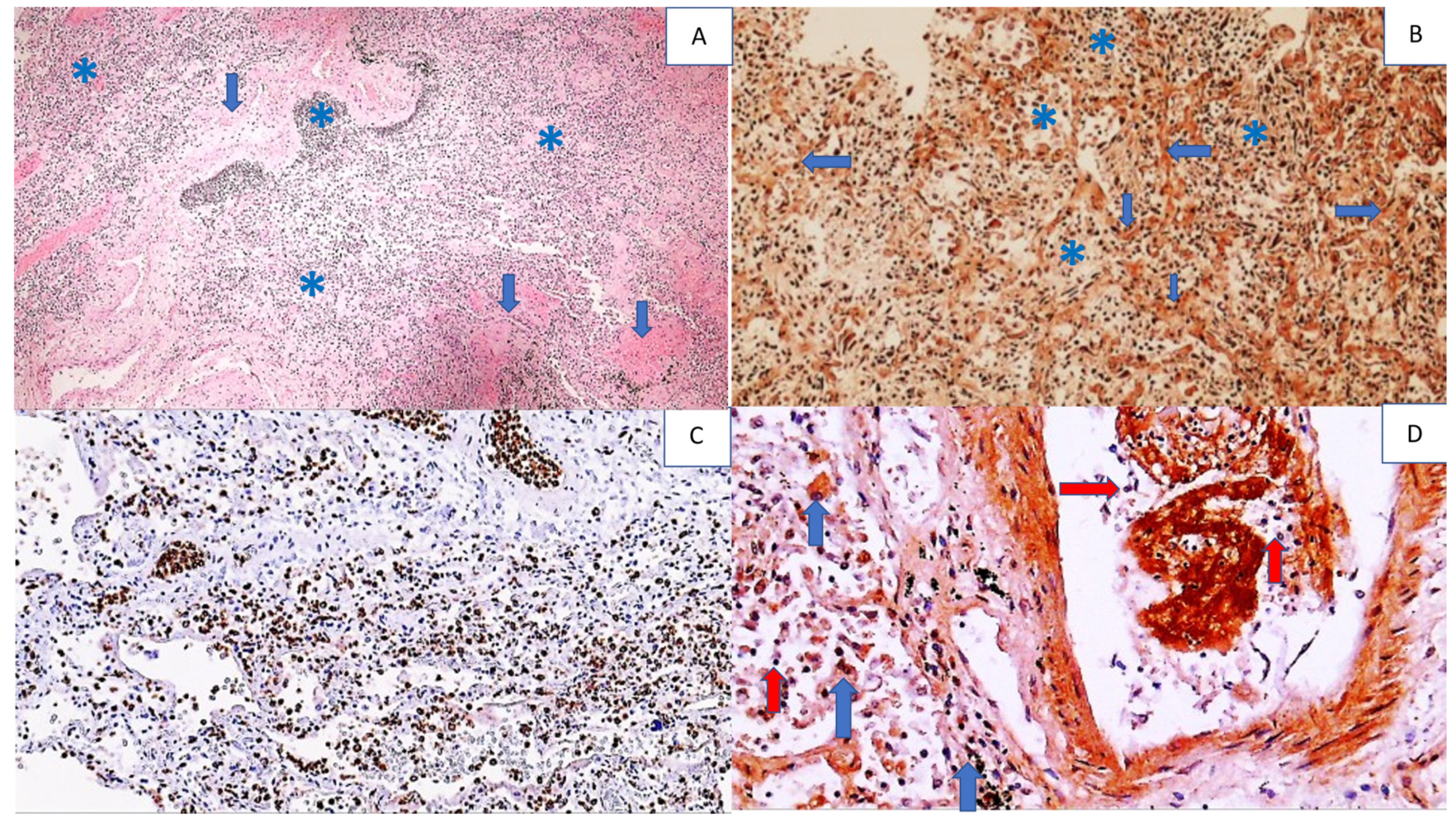
| Brain | Edematous | Perivascular CD3-positive T cells PMNs and T lymphocytes within the blood vessels | Strong positivity of blood vessels and vascular content Very strong and diffuse immunopositivity in neurons Positive astroglia | Positive neurons and reactive astrocytes |
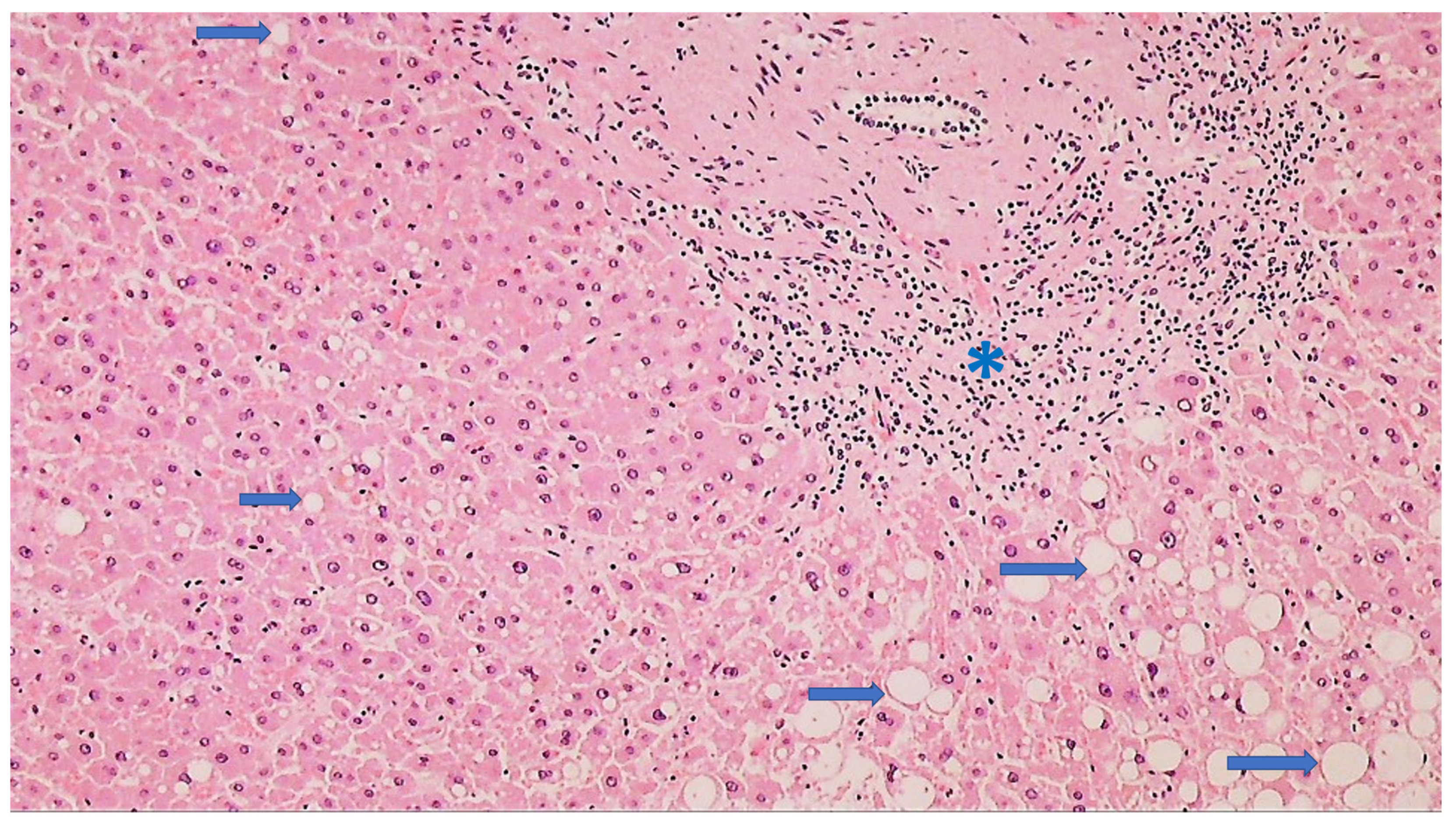
| Liver | Often steatosis Rarely centrilobular cyanosis | Not prominent | Positivity of the blood vessels Positive hepatocytes | Positive hepatocytes and epithelium of the bile ducts |
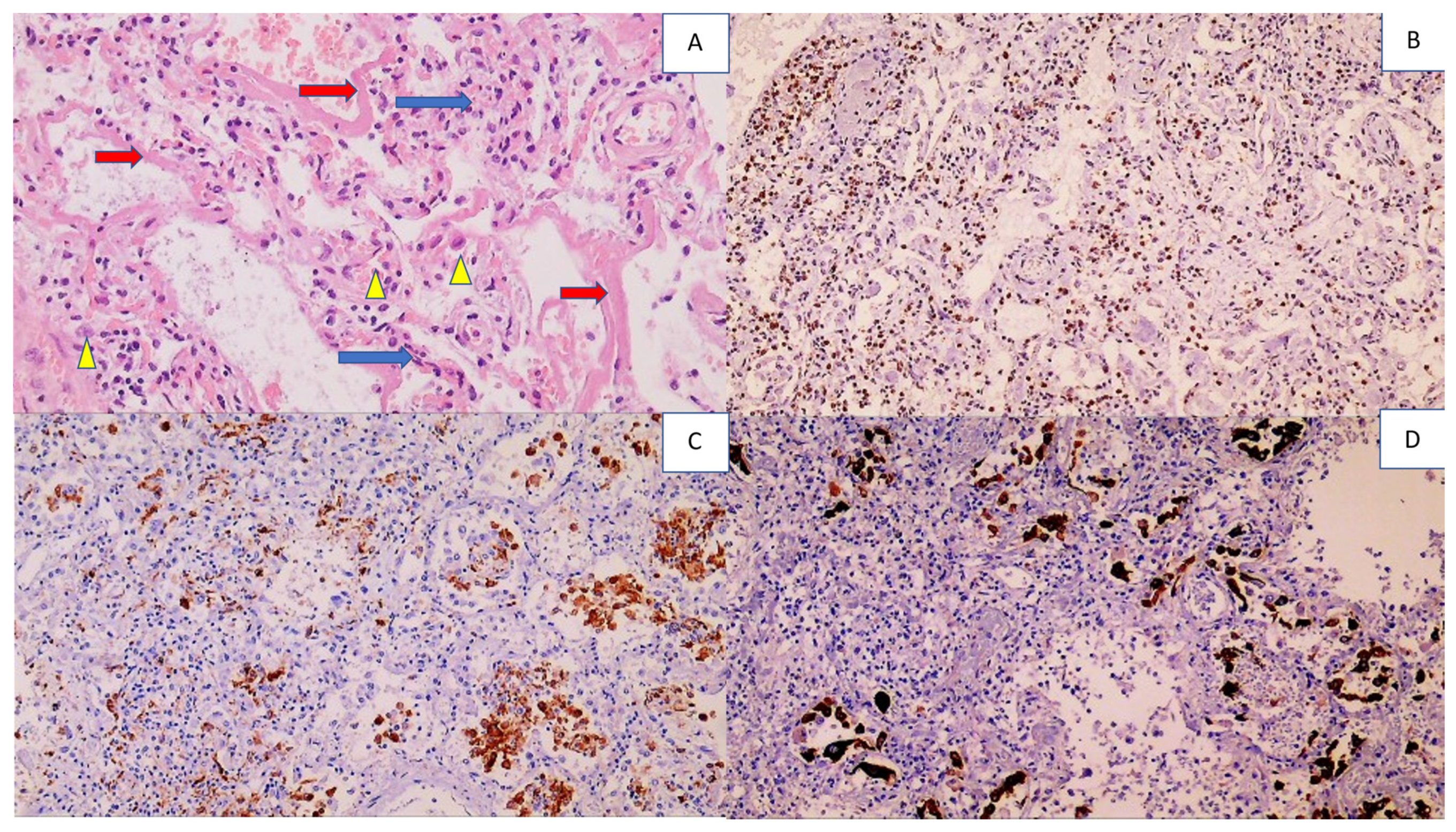
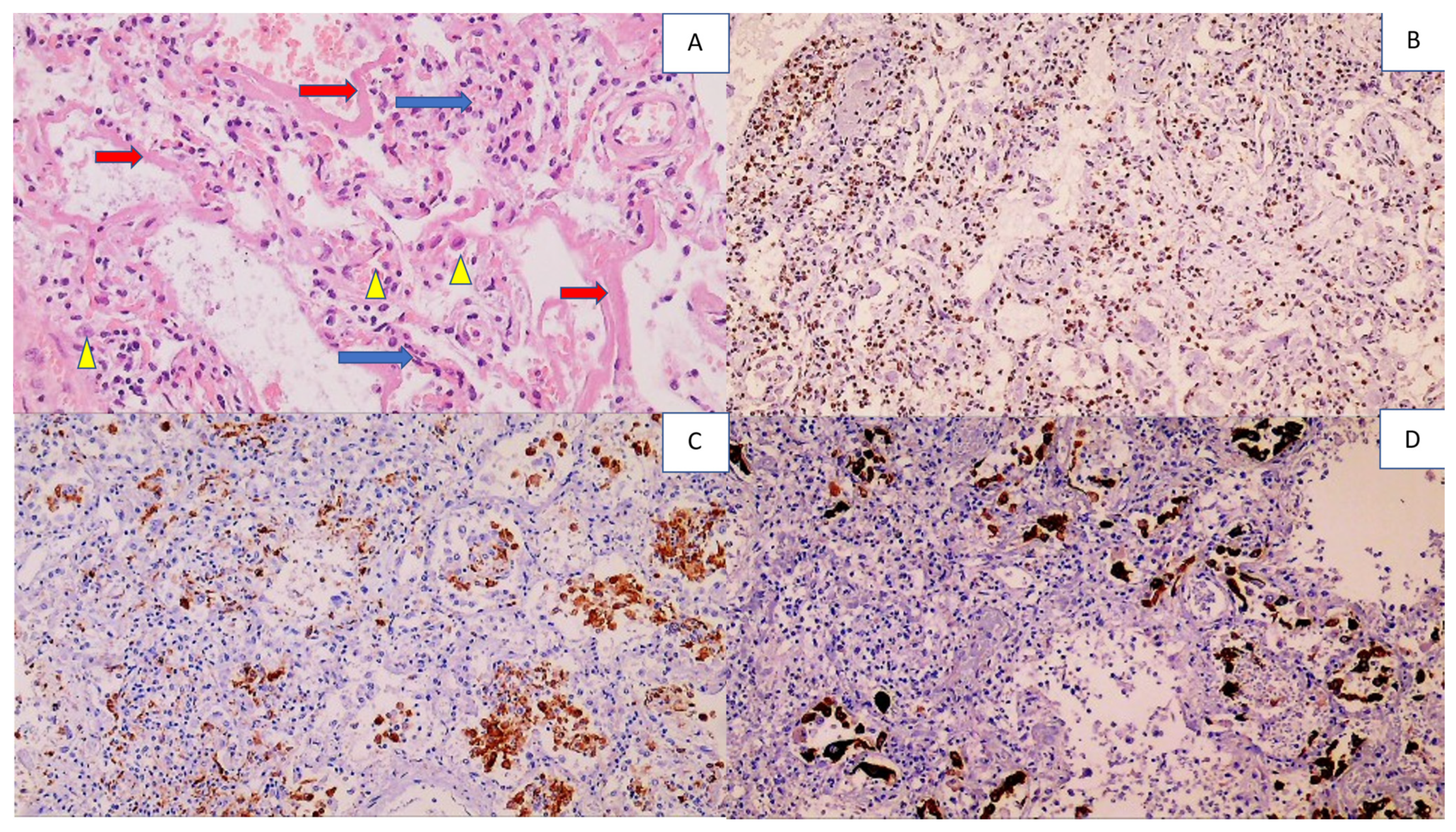
| Kidneys | Acute tubular necrosis, with necrotization of the epithelium of the proximal and distal canals Structure of the glomeruli and collecting ducts preserved | Inflammatory cells were not found | Strong positivity, but only in collective tubules | Positive glomerular cells and collecting ducts |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zarkovic, N.; Jakovcevic, A.; Mataic, A.; Jaganjac, M.; Vukovic, T.; Waeg, G.; Zarkovic, K. Post-mortem Findings of Inflammatory Cells and the Association of 4-Hydroxynonenal with Systemic Vascular and Oxidative Stress in Lethal COVID-19. Cells 2022, 11, 444. https://doi.org/10.3390/cells11030444
Zarkovic N, Jakovcevic A, Mataic A, Jaganjac M, Vukovic T, Waeg G, Zarkovic K. Post-mortem Findings of Inflammatory Cells and the Association of 4-Hydroxynonenal with Systemic Vascular and Oxidative Stress in Lethal COVID-19. Cells. 2022; 11(3):444. https://doi.org/10.3390/cells11030444
Chicago/Turabian StyleZarkovic, Neven, Antonia Jakovcevic, Ana Mataic, Morana Jaganjac, Tea Vukovic, Georg Waeg, and Kamelija Zarkovic. 2022. "Post-mortem Findings of Inflammatory Cells and the Association of 4-Hydroxynonenal with Systemic Vascular and Oxidative Stress in Lethal COVID-19" Cells 11, no. 3: 444. https://doi.org/10.3390/cells11030444
APA StyleZarkovic, N., Jakovcevic, A., Mataic, A., Jaganjac, M., Vukovic, T., Waeg, G., & Zarkovic, K. (2022). Post-mortem Findings of Inflammatory Cells and the Association of 4-Hydroxynonenal with Systemic Vascular and Oxidative Stress in Lethal COVID-19. Cells, 11(3), 444. https://doi.org/10.3390/cells11030444