
Clinical and Translational Significance of Basophils in Patients with Cancer


 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction: Characteristics and Functions of Basophils in Protective Immunity, Th2 and Inflammatory Diseases
2. Basophil Infiltration and Functions in Solid Tumors
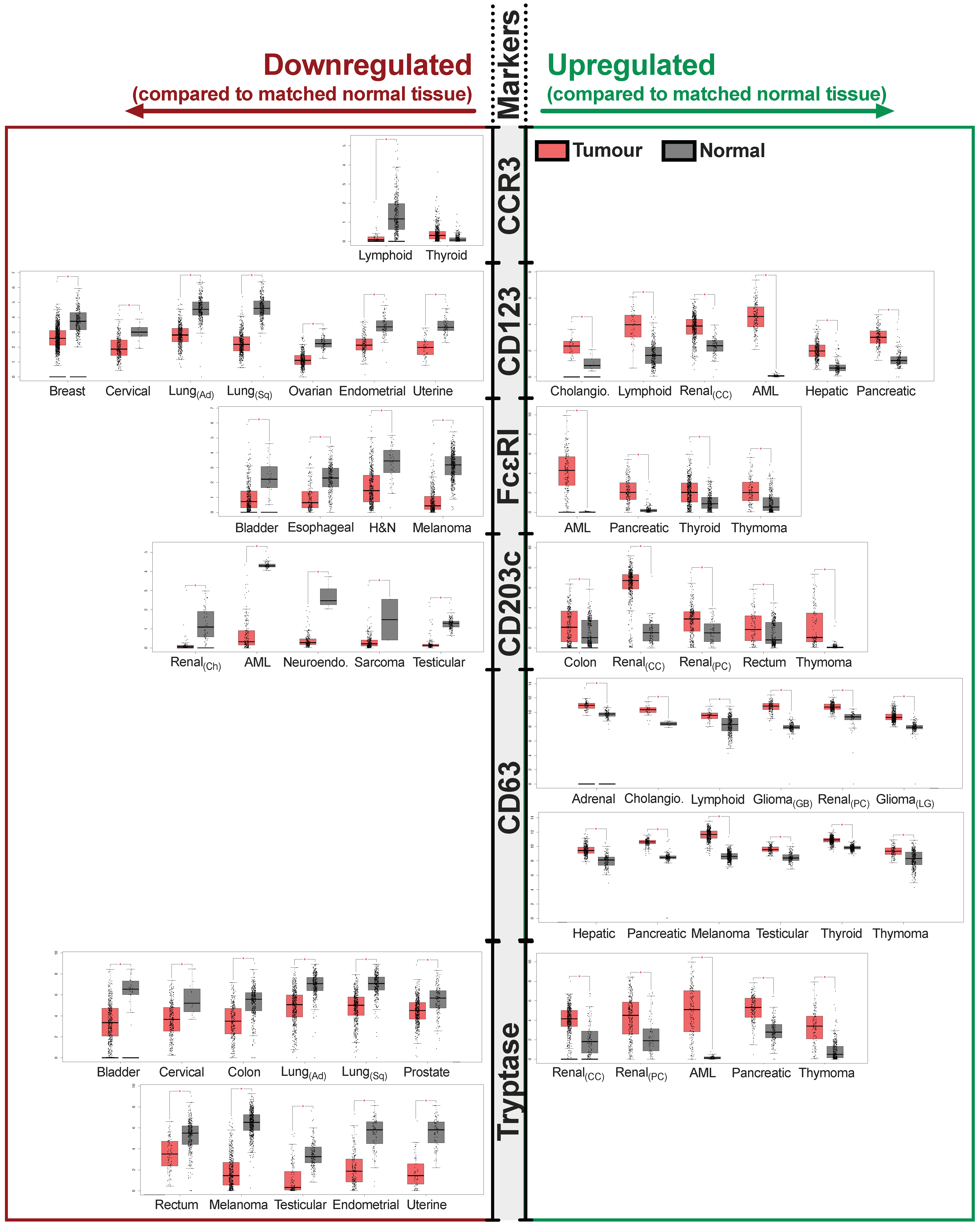
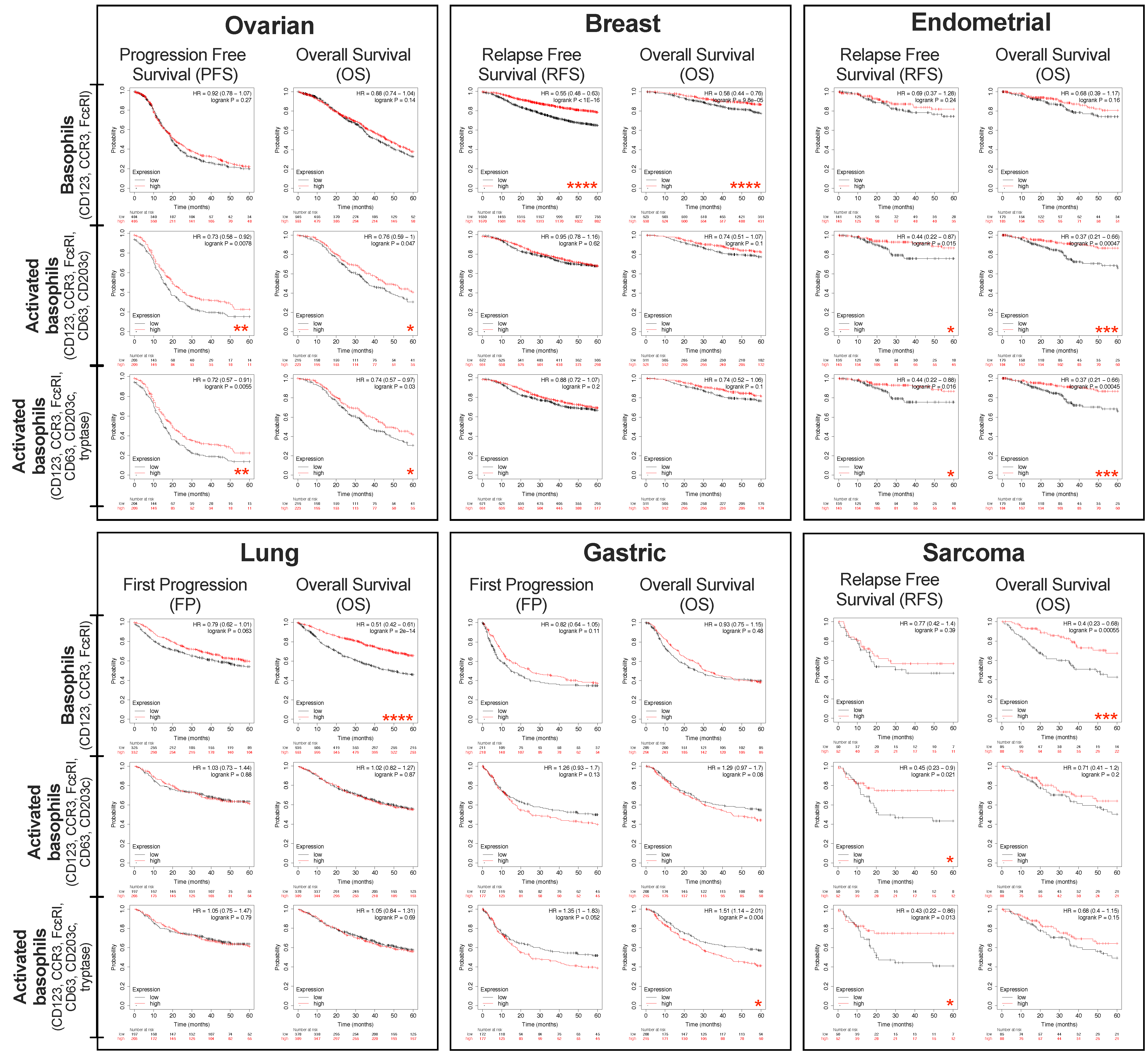
3. Evaluating the Presence and Activation States of Basophils and Associations with Clinical Outcomes in Different Solid Tumors
4. Basophil Activation Studied Ex Vivo to Detect Allergic Reactions to Therapeutic Agents and to Monitor Desensitization to Oncology Drugs
4.1. Basophil Activation Testing for the Detection of Allergic Reactions
4.2. Basophil Activation Testing in AllergoOncology
5. Basophil Activation Evaluated in the BAT as a Tool to Predict Propensity for Type 1 Hypersensitivity to Emerging IgE Immunotherapy Agents
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marone, G.; Schroeder, J.T.; Mattei, F.; Loffredo, S.; Gambardella, A.R.; Poto, R.; De Paulis, A.; Schiavoni, G.; Varricchi, G. Is There a Role for Basophils in Cancer? Front. Immunol. 2020, 11, 2103. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, A.M.; Monahan-Earley, R.A.; Estrella, P.; Kissell, S.; Donahue, R.E. Ultrastructure of monkey peripheral blood basophils stimulated to develop in vivo by recombinant human interleukin 3. Lab. Investig. 1989, 61, 677–690. [Google Scholar] [PubMed]
- Hardin, J.H.; Spicer, S.S. Ultrastructural Localization of Dialyzed Iron-Reactive Mucosubstance in Rabbit Heterophils, Basophils, and Eosinophils. J. Cell Biol. 1971, 48, 368–386. [Google Scholar] [CrossRef] [PubMed]
- Seder, R.A.; Paul, W.E.; Dvorak, A.M.; Sharkis, S.J.; Kagey-Sobotka, A.; Niv, Y.; Finkelman, F.D.; Barbieri, S.A.; Galli, S.J.; Plaut, M. Mouse splenic and bone marrow cell populations that express high-affinity Fc epsilon receptors and produce interleukin 4 are highly enriched in basophils. Proc. Natl. Acad. Sci. USA 1991, 88, 2835–2839. [Google Scholar] [CrossRef]
- Cohen, M.; Giladi, A.; Gorki, A.-D.; Solodkin, D.G.; Zada, M.; Hladik, A.; Miklosi, A.; Salame, T.-M.; Halpern, K.B.; David, E.; et al. Lung Single-Cell Signaling Interaction Map Reveals Basophil Role in Macrophage Imprinting. Cell 2018, 175, 1031–1044.e18. [Google Scholar] [CrossRef] [PubMed]
- Daëron, M. Innate myeloid cells under the control of adaptive immunity: The example of mast cells and basophils. Curr. Opin. Immunol. 2016, 38, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Kämpfer, S.S.; Odermatt, A.; Dahinden, C.A.; Fux, M. Late IL-3–induced phenotypic and functional alterations in human basophils require continuous IL-3 receptor signaling. J. Leukoc. Biol. 2016, 101, 227–238. [Google Scholar] [CrossRef]
- Lantz, C.S.; Boesiger, J.; Song, C.H.; Mach, N.; Kobayashi, T.; Mulligan, R.C.; Nawa, Y.; Dranoff, G.; Galli, S.J. Role for interleukin-3 in mast-cell and basophil development and in immunity to parasites. Nature 1998, 392, 90–93. [Google Scholar] [CrossRef]
- MacGlashan, D. Expression of CD203c and CD63 in human basophils: Relationship to differential regulation of piecemeal and anaphylactic degranulation processes. Clin. Exp. Allergy 2010, 40, 1365–1377. [Google Scholar] [CrossRef]
- Metcalfe, D.D.; Pawankar, R.; Ackerman, S.J.; Akin, C.; Clayton, F.; Falcone, F.H.; Gleich, G.J.; Irani, A.-M.; Johansson, M.W.; Klion, A.D.; et al. Biomarkers of the involvement of mast cells, basophils and eosinophils in asthma and allergic diseases. World Allergy Organ. J. 2016, 9, 7. [Google Scholar] [CrossRef]
- Kojima, T.; Obata-Ninomiya, K.; Mukai, K.; Sato, S.; Takai, T.; Minegishi, Y.; Karasuyama, H. Mast Cells and Basophils Are Selectively Activated In Vitro and In Vivo through CD200R3 in an IgE-Independent Manner. J. Immunol. 2007, 179, 7093–7100. [Google Scholar] [CrossRef] [PubMed]
- Obata-Ninomiya, K.; Mukai, K.; Tsujimura, Y.; Ishiwata, K.; Kawano, Y.; Minegishi, Y.; Watanabe, N.; Karasuyama, H. Basophils are essential initiators of a novel type of chronic allergic inflammation. Blood 2007, 110, 913–920. [Google Scholar] [CrossRef] [PubMed]
- Perrigoue, J.G.; Saenz, S.A.; Siracusa, M.C.; Allenspach, E.; Taylor, B.C.; Giacomin, P.; Nair, M.; Du, Y.; Zaph, C.; Van Rooijen, N.; et al. MHC class II–dependent basophil–CD4+ T cell interactions promote TH2 cytokine–dependent immunity. Nat. Immunol. 2009, 10, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Sokol, C.L.; Chu, N.-Q.; Yu, S.; Nish, S.A.; Laufer, T.M.; Medzhitov, R. Basophils function as antigen-presenting cells for an allergen-induced T helper type 2 response. Nat. Immunol. 2009, 10, 713–720. [Google Scholar] [CrossRef]
- Yoshimoto, T.; Yasuda, K.; Tanaka, H.; Nakahira, M.; Imai, Y.; Fujimori, Y.; Nakanishi, K. Basophils contribute to TH2-IgE responses in vivo via IL-4 production and presentation of peptide–MHC class II complexes to CD4+ T cells. Nat. Immunol. 2009, 10, 706–712. [Google Scholar] [CrossRef]
- El Hachem, C.; Hener, P.; Kirstetter, P.; Li, J.; Chan, S.; Li, M. Treatment of MCPT8DTR mice with high- or low-dose diphtheria toxin leads to differential depletion of basophils and granulocyte-macrophage progenitors. Eur. J. Immunol. 2018, 48, 861–873. [Google Scholar] [CrossRef]
- Poorafshar, M.; Helmby, H.; Troye-Blomberg, M.; Hellman, L. MMCP-8, the first lineage-specific differentiation marker for mouse basophils. Elevated numbers of potent IL-4-producing and MMCP-8-positive cells in spleens of malaria-infected mice. Eur. J. Immunol. 2000, 30, 2660–2668. [Google Scholar] [CrossRef]
- Sawaguchi, M.; Tanaka, S.; Nakatani, Y.; Harada, Y.; Mukai, K.; Matsunaga, Y.; Ishiwata, K.; Oboki, K.; Kambayashi, T.; Watanabe, N.; et al. Role of Mast Cells and Basophils in IgE Responses and in Allergic Airway Hyperresponsiveness. J. Immunol. 2012, 188, 1809–1818. [Google Scholar] [CrossRef]
- Ugajin, T.; Kojima, T.; Mukai, K.; Obata-Ninomiya, K.; Kawano, Y.; Minegishi, Y.; Eishi, Y.; Yokozeki, H.; Karasuyama, H. Basophils preferentially express mouse mast cell protease 11 among the mast cell tryptase family in contrast to mast cells. J. Leukoc. Biol. 2009, 86, 1417–1425. [Google Scholar] [CrossRef]
- Wada, T.; Ishiwata, K.; Koseki, H.; Ishikura, T.; Ugajin, T.; Ohnuma, N.; Obata-Ninomiya, K.; Ishikawa, R.; Yoshikawa, S.; Mukai, K.; et al. Selective ablation of basophils in mice reveals their nonredundant role in acquired immunity against ticks. J. Clin. Investig. 2010, 120, 2867–2875. [Google Scholar] [CrossRef]
- Mukai, K.; BenBarak, M.J.; Tachibana, M.; Nishida, K.; Karasuyama, H.; Taniuchi, I.; Galli, S.J. Critical role of P1-Runx1 in mouse basophil development. Blood 2012, 120, 76–85. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ohnmacht, C.; Schwartz, C.; Panzer, M.; Schiedewitz, I.; Naumann, R.; Voehringer, D. Basophils Orchestrate Chronic Allergic Dermatitis and Protective Immunity against Helminths. Immunity 2010, 33, 364–374. [Google Scholar] [CrossRef]
- Sullivan, B.M.; Liang, H.-E.; Bando, J.; Wu, D.; Cheng, L.E.; McKerrow, J.K.; Allen, C.D.C.; Locksley, R.M. Genetic analysis of basophil function in vivo. Nat. Immunol. 2011, 12, 527–535. [Google Scholar] [CrossRef]
- Kirshenbaum, A.S.; Goff, J.P.; Dreskin, S.C.; Irani, A.M.; Schwartz, L.B.; Metcalfe, D.D. IL-3-dependent growth of basophil-like cells and mastlike cells from human bone marrow. J. Immunol. 1989, 142, 2424–2429. [Google Scholar] [PubMed]
- Ohnmacht, C.; Voehringer, D. Basophil effector function and homeostasis during helminth infection. Blood 2009, 113, 2816–2825. [Google Scholar] [CrossRef] [PubMed]
- Lavin, Y.; Kobayashi, S.; Leader, A.; Amir, E.-A.D.; Elefant, N.; Bigenwald, C.; Remark, R.; Sweeney, R.; Becker, C.D.; Levine, J.H.; et al. Innate Immune Landscape in Early Lung Adenocarcinoma by Paired Single-Cell Analyses. Cell 2017, 169, 750–765.e17. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Satoh, T.; Takayama, K.; Miyagishi, C.; Walls, A.; Yokozeki, H. Basophil recruitment and activation in inflammatory skin diseases. Allergy 2011, 66, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Bax, H.J.; Chauhan, J.; Stavraka, C.; Khiabany, A.; Nakamura, M.; Pellizzari, G.; Ilieva, K.M.; Lombardi, S.; Gould, H.J.; Corrigan, C.J.; et al. Basophils from Cancer Patients Respond to Immune Stimuli and Predict Clinical Outcome. Cells 2020, 9, 1631. [Google Scholar] [CrossRef]
- Wakahara, K.; Baba, N.; Van, V.Q.; Bégin, P.; Rubio, M.; Ferraro, P.; Panzini, B.; Wassef, R.; Lahaie, R.; Caussignac, Y.; et al. Human basophils interact with memory T cells to augment Th17 responses. Blood 2012, 120, 4761–4771. [Google Scholar] [CrossRef]
- Piliponsky, A.M.; Shubin, N.J.; Lahiri, A.K.; Truong, P.; Clauson, M.; Niino, K.; Tsuha, A.L.; Nedospasov, S.A.; Karasuyama, H.; Reber, L.; et al. Basophil-derived tumor necrosis factor can enhance survival in a sepsis model in mice. Nat. Immunol. 2019, 20, 129–140. [Google Scholar] [CrossRef]
- Sektioglu, I.M.; Carretero, R.; Bulbuc, N.; Bald, T.; Tüting, T.; Rudensky, A.Y.; Hämmerling, G.J. Basophils Promote Tumor Rejection via Chemotaxis and Infiltration of CD8+ T Cells. Cancer Res. 2017, 77, 291–302. [Google Scholar] [CrossRef]
- De Monte, L.; Woermann, S.; Brunetto, E.; Heltai, S.; Magliacane, G.; Reni, M.; Paganoni, A.M.; Recalde, H.; Mondino, A.; Falconi, M.; et al. Basophil Recruitment into Tumor-Draining Lymph Nodes Correlates with Th2 Inflammation and Reduced Survival in Pancreatic Cancer Patients. Cancer Res. 2016, 76, 1792–1803. [Google Scholar] [CrossRef]
- Liu, Q.; Luo, D.; Cai, S.; Li, Q.; Li, X. Circulating basophil count as a prognostic marker of tumor aggressiveness and survival outcomes in colorectal cancer. Clin. Transl. Med. 2020, 9, 6. [Google Scholar] [CrossRef]
- Rigoni, A.; Colombo, M.; Pucillo, C. Mast cells, basophils and eosinophils: From allergy to cancer. Semin. Immunol. 2018, 35, 29–34. [Google Scholar] [CrossRef]
- Ciprandi, G.; Cirillo, I.; Vizzaccaro, A.; Milanese, M.; Tosca, M.A. Nasal Obstruction in Patients with Seasonal Allergic Rhinitis: Relationships between Allergic Inflammation and Nasal Airflow. Int. Arch. Allergy Immunol. 2004, 134, 34–40. [Google Scholar] [CrossRef]
- Ciprandi, G.; Murdaca, G.; Colombo, B.M.; DE Amici, M.; Marseglia, G.L. Serum vascular endothelial growth factor in allergic rhinitis and systemic lupus erythematosus. Hum. Immunol. 2008, 69, 510–512. [Google Scholar] [CrossRef]
- Murdaca, G.; Spanò, F.; Miglino, M.; Puppo, F. Effects of TNF-α inhibitors upon the mechanisms of action of VEGF. Immunother. 2013, 5, 113–115. [Google Scholar] [CrossRef]
- Castells, M.C.; Irani, A.M.; Schwartz, L.B. Evaluation of human peripheral blood leukocytes for mast cell tryptase. J. Immunol. 1987, 138, 2184–2189. [Google Scholar]
- Falkencrone, S.; Poulsen, L.K.; Bindslev-Jensen, C.; Woetmann, A.; Odum, N.; Poulsen, B.C.; Blom, L.; Jensen, B.M.; Gibbs, B.F.; Yasinska, I.M.; et al. IgE-mediated basophil tumour necrosis factor alpha induces matrix metalloproteinase-9 from monocytes. Allergy 2013, 68, 614–620. [Google Scholar] [CrossRef]
- Foster, B.; Schwartz, L.B.; Devouassoux, G.; Metcalfe, D.D.; Prussin, C. Characterization of mast-cell tryptase-expressing peripheral blood cells as basophils. J. Allergy Clin. Immunol. 2002, 109, 287–293. [Google Scholar] [CrossRef]
- Jogie-Brahim, S.; Min, H.-K.; Fukuoka, Y.; Xia, H.-Z.; Schwartz, L.B. Expression of α-tryptase and β-tryptase by human basophils. J. Allergy Clin. Immunol. 2004, 113, 1086–1092. [Google Scholar] [CrossRef]
- Chavez, J.; Young, H.W.J.; Corry, D.B.; Lieberman, M.W. Interactions between Leukotriene C4 and Interleukin 13 Signaling Pathways in a Mouse Model of Airway Disease. Arch. Pathol. Lab. Med. 2006, 130, 440–446. [Google Scholar] [CrossRef]
- Duah, E.; Adapala, R.K.; Al-Azzam, N.; Kondeti, V.; Gombedza, F.; Thodeti, C.K.; Paruchuri, S. Cysteinyl leukotrienes regulate endothelial cell inflammatory and proliferative signals through CysLT2 and CysLT1 receptors. Sci. Rep. 2013, 3, 3274. [Google Scholar] [CrossRef]
- Ni, N.C.; Yan, D.; Ballantyne, L.L.; Barajas-Espinosa, A.; Amand, T.S.; Pratt, D.A.; Funk, C.D. A Selective Cysteinyl Leukotriene Receptor 2 Antagonist Blocks Myocardial Ischemia/Reperfusion Injury and Vascular Permeability in Mice. J. Pharmacol. Exp. Ther. 2011, 339, 768–778. [Google Scholar] [CrossRef]
- Riccioni, G.; Bäck, M. Leukotrienes as Modifiers of Preclinical Atherosclerosis? Sci. World J. 2012, 2012, 1–6. [Google Scholar] [CrossRef]
- Barajas-Espinosa, A.; Ni, N.C.; Yan, D.; Zarini, S.; Murphy, R.C.; Funk, C.D. The cysteinyl leukotriene 2 receptor mediates retinal edema and pathological neovascularization in a murine model of oxygen-induced retinopathy. FASEB J. 2011, 26, 1100–1109. [Google Scholar] [CrossRef]
- Marone, G.; Casolaro, V.; Paganelli, R.; Quinti, I. IgG Anti-IgE from Atopic Dermatitis Induces Mediator Release from Basophils and Mast Cells. J. Investig. Dermatol. 1989, 93, 246–252. [Google Scholar] [CrossRef]
- MacGlashan, D.W.; Peters, S.P.; Warner, J.; Lichtenstein, L.M. Characteristics of human basophil sulfidopeptide leukotriene release: Releasability defined as the ability of the basophil to respond to dimeric cross-links. J. Immunol. 1986, 136, 2231–2239. [Google Scholar]
- Duah, E.; Teegala, L.R.; Kondeti, V.; Adapala, R.K.; Keshamouni, V.G.; Kanaoka, Y.; Austen, K.F.; Thodeti, C.K.; Paruchuri, S. Cysteinyl leukotriene 2 receptor promotes endothelial permeability, tumor angiogenesis, and metastasis. Proc. Natl. Acad. Sci. 2019, 116, 199–204. [Google Scholar] [CrossRef]
- Morshed, M.; Hlushchuk, R.; Simon, D.; Walls, A.F.; Obata-Ninomiya, K.; Karasuyama, H.; Djonov, V.; Eggel, A.; Kaufmann, T.; Simon, H.-U.; et al. NADPH Oxidase–Independent Formation of Extracellular DNA Traps by Basophils. J. Immunol. 2014, 192, 5314–5323. [Google Scholar] [CrossRef]
- Schorn, C.; Janko, C.; Latzko, M.; Chaurio, R.; Schett, G.; Herrmann, M. Monosodium urate crystals induce extracellular DNA traps in neutrophils, eosinophils, and basophils but not in mononuclear cells. Front. Immunol. 2012, 3, 277. [Google Scholar] [CrossRef]
- Yousefi, S.; Morshed, M.; Amini, P.; Stojkov, D.; Simon, D.; von Gunten, S.; Kaufmann, T.; Simon, H.-U. Basophils exhibit antibacterial activity through extracellular trap formation. Allergy 2015, 70, 1184–1188. [Google Scholar] [CrossRef]
- Park, J.; Wysocki, R.W.; Amoozgar, Z.; Maiorino, L.; Fein, M.R.; Jorns, J.; Schott, A.F.; Kinugasa-Katayama, Y.; Lee, Y.; Won, N.H.; et al. Cancer cells induce metastasis-supporting neutrophil extracellular DNA traps. Sci. Transl. Med. 2016, 8, 361ra138. [Google Scholar] [CrossRef]
- Cools-Lartigue, J.J.; Spicer, J.D.; McDonald, B.; Chow, S.; Kubes, P.; Ferri, L.E. Abstract 2972: Neutrophil extracellular traps sequester circulating tumor cellsin vitroand in a murine model of metastasis. Tumor Biology 2012, 72, 2972. [Google Scholar] [CrossRef]
- Teijeira, Á.; Garasa, S.; Gato, M.; Alfaro, C.; Migueliz, I.; Cirella, A.; de Andrea, C.; Ochoa, M.C.; Otano, I.; Etxeberria, I.; et al. CXCR1 and CXCR2 Chemokine Receptor Agonists Produced by Tumors Induce Neutrophil Extracellular Traps that Interfere with Immune Cytotoxicity. Immunity 2020, 52, 856–871.e8. [Google Scholar] [CrossRef]
- Yang, L.; Liu, Q.; Zhang, X.; Liu, X.; Zhou, B.; Chen, J.; Huang, D.; Li, J.; Li, H.; Chen, F.; et al. DNA of neutrophil extracellular traps promotes cancer metastasis via CCDC25. Nature 2020, 583, 133–138. [Google Scholar] [CrossRef]
- Crawford, G.; Hayes, M.D.; Seoane, R.C.; Ward, S.; Dalessandri, T.; Lai, C.; Healy, E.; Kipling, D.; Proby, C.; Moyes, C.; et al. Epithelial damage and tissue γδ T cells promote a unique tumor-protective IgE response. Nat. Immunol. 2018, 19, 859–870. [Google Scholar] [CrossRef]
- Andreone, S.; Spadaro, F.; Buccione, C.; Mancini, J.; Tinari, A.; Sestili, P.; Gambardella, A.R.; Lucarini, V.; Ziccheddu, G.; Parolini, I.; et al. IL-33 Promotes CD11b/CD18-Mediated Adhesion of Eosinophils to Cancer Cells and Synapse-Polarized Degranulation Leading to Tumor Cell Killing. Cancers 2019, 11, 1664. [Google Scholar] [CrossRef]
- Lucarini, V.; Ziccheddu, G.; Macchia, I.; La Sorsa, V.; Peschiaroli, F.; Buccione, C.; Sistigu, A.; Sanchez, M.; Andreone, S.; D’Urso, M.T.; et al. IL-33 restricts tumor growth and inhibits pulmonary metastasis in melanoma-bearing mice through eosinophils. OncoImmunology 2017, 6, e1317420. [Google Scholar] [CrossRef]
- Tschopp, C.M.; Spiegl, N.; Didichenko, S.; Lutmann, W.; Julius, P.; Virchow, J.C.; Hack, C.E.; Dahinden, C.A. Granzyme B, a novel mediator of allergic inflammation: Its induction and release in blood basophils and human asthma. Blood 2006, 108, 2290–2299. [Google Scholar] [CrossRef]
- Schroeder, J.T.; Adeosun, A.A.; Do, D.; Bieneman, A.P. Galectin-3 is essential for IgE-dependent activation of human basophils by A549 lung epithelial cells. J. Allergy Clin. Immunol. 2019, 144, 312–315.e1. [Google Scholar] [CrossRef]
- Sciacchitano, S.; Lavra, L.; Morgante, A.; Ulivieri, A.; Magi, F.; De Francesco, G.P.; Bellotti, C.; Salehi, L.B.; Ricci, A. Galectin-3: One Molecule for an Alphabet of Diseases, from A to Z. Int. J. Mol. Sci. 2018, 19, 379. [Google Scholar] [CrossRef]
- Schroeder, J.T.; Bieneman, A.P. Activation of Human Basophils by A549 Lung Epithelial Cells Reveals a Novel IgE-Dependent Response Independent of Allergen. J. Immunol. 2017, 199, 855–865. [Google Scholar] [CrossRef]
- Sharma, M.; Das, M.; Stephen-Victor, E.; Galeotti, C.; Karnam, A.; Maddur, M.S.; Bruneval, P.; Kaveri, S.V.; Bayry, J. Regulatory T cells induce activation rather than suppression of human basophils. Sci. Immunol. 2018, 3, eaan0829. [Google Scholar] [CrossRef]
- Messaoudene, M.; Périer, A.; Fregni, G.; Neves, E.; Zitvogel, L.; Cremer, I.; Chanal, J.; Sastre-Garau, X.; Deschamps, L.; Marinho, E.; et al. Characterization of the Microenvironment in Positive and Negative Sentinel Lymph Nodes from Melanoma Patients. PLoS ONE 2015, 10, e0133363. [Google Scholar] [CrossRef]
- Cochran, A.J.; Roberts, A.; Wen, D.-R.; Huang, R.-R.; Itakura, E.; Luo, F.; Binder, S.W. Optimized assessment of sentinel lymph nodes for metastatic melanoma: Implications for regional surgery and overall treatment planning. Ann. Surg. Oncol. 2004, 11, 156–161. [Google Scholar] [CrossRef]
- Mansfield, A.S.; Holtan, S.G.; Grotz, T.E.; Allred, J.B.; Jakub, J.W.; Erickson, L.A.; Markovic, S.N. Regional immunity in melanoma: Immunosuppressive changes precede nodal metastasis. Mod. Pathol. 2010, 24, 487–494. [Google Scholar] [CrossRef]
- Mansfield, A.S.; Heikkila, P.; Von Smitten, K.; Vakkila, J.; Leidenius, M. Metastasis to sentinel lymph nodes in breast cancer is associated with maturation arrest of dendritic cells and poor co-localization of dendritic cells and CD8+ T cells. Virchows Arch. 2011, 459, 391–398. [Google Scholar] [CrossRef]
- Merluzzi, S.; Betto, E.; Ceccaroni, A.A.; Magris, R.; Giunta, M.; Mion, F. Mast cells, basophils and B cell connection network. Mol. Immunol. 2015, 63, 94–103. [Google Scholar] [CrossRef]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef]
- Győrffy, B. Survival analysis across the entire transcriptome identifies biomarkers with the highest prognostic power in breast cancer. Comput. Struct. Biotechnol. J. 2021, 19, 4101–4109. [Google Scholar] [CrossRef]
- Cheung, A.; Opzoomer, J.; Ilieva, K.M.; Gazinska, P.; Hoffmann, R.M.; Mirza, H.; Marlow, R.; Francesch-Domenech, E.; Fittall, M.; Rodriguez, D.D.; et al. Anti-Folate Receptor Alpha–Directed Antibody Therapies Restrict the Growth of Triple-negative Breast Cancer. Clin. Cancer Res. 2018, 24, 5098–5111. [Google Scholar] [CrossRef]
- Harris, R.J.; Cheung, A.; Ng, J.C.F.; Laddach, R.; Chenoweth, A.M.; Crescioli, S.; Fittall, M.; Dominguez-Rodriguez, D.; Roberts, J.; Levi, D.; et al. Tumor-Infiltrating B Lymphocyte Profiling Identifies IgG-Biased, Clonally Expanded Prognostic Phenotypes in Triple-Negative Breast Cancer. Cancer Res. 2021, 81, 4290–4304. [Google Scholar] [CrossRef]
- Moneret-Vautrin, D.A.; Sainte-Laudy, J.; Kanny, G.; Frémont, S. Human basophil activation measured by CD63 expression and LTC4 release in IgE-mediated food allergy. Ann. Allergy Asthma Immunol. 1999, 82, 33–40. [Google Scholar] [CrossRef]
- Ocmant, A.; Mulier, S.; Hanssens, L.; Goldman, M.; Casimir, G.; Mascart, F.; Schandené, L. Basophil activation tests for the diagnosis of food allergy in children. Clin. Exp. Allergy 2009, 39, 1234–1245. [Google Scholar] [CrossRef]
- Santos, A.; Douiri, A.; Bécares, N.; Wu, S.-Y.; Stephens, A.; Radulovic, S.; Chan, S.M.; Fox, A.; Du Toit, G.; Turcanu, V.; et al. Basophil activation test discriminates between allergy and tolerance in peanut-sensitized children. J. Allergy Clin. Immunol. 2014, 134, 645–652. [Google Scholar] [CrossRef]
- Santos, A.F.; Shreffler, W.G. Road map for the clinical application of the basophil activation test in food allergy. Clin. Exp. Allergy 2017, 47, 1115–1124. [Google Scholar] [CrossRef]
- Sato, S.; Tachimoto, H.; Shukuya, A.; Kurosaka, N.; Yanagida, N.; Utsunomiya, T.; Iguchi, M.; Komata, T.; Imai, T.; Tomikawa, M.; et al. Basophil Activation Marker CD203c Is Useful in the Diagnosis of Hen’s Egg and Cow’s Milk Allergies in Children. Int. Arch. Allergy Immunol. 2010, 152, 54–61. [Google Scholar] [CrossRef]
- Ford, L.S.; Bloom, K.A.; Nowak-Wegrzyn, A.; Shreffler, W.; Masilamani, M.; Sampson, H.A. Basophil reactivity, wheal size, and immunoglobulin levels distinguish degrees of cow’s milk tolerance. J. Allergy Clin. Immunol. 2013, 131, 180–186.e3. [Google Scholar] [CrossRef]
- Aranda, A.; Mayorga, C.; Ariza, A.; Doña, I.; Rosado, A.; Blanca-Lopez, N.; Andreu, I.; Torres, M.J. In vitro evaluation of IgE-mediated hypersensitivity reactions to quinolones. Allergy 2010, 66, 247–254. [Google Scholar] [CrossRef]
- Dewachter, P.; Chollet-Martin, S.; Mouton-Faivre, C.; de Chaisemartin, L.; Nicaise-Roland, P. Comparison of Basophil Activation Test and Skin Testing Performances in NMBA Allergy. J. Allergy Clin. Immunol. Pr. 2018, 6, 1681–1689. [Google Scholar] [CrossRef] [PubMed]
- Fernández, T.D.; Ariza, A.; Palomares, F.; Montañez, M.I.; Salas, M.; Martín-Serrano, A.; Fernández, R.; Ruiz, A.; Blanca, M.; Mayorga, C.; et al. Hypersensitivity to fluoroquinolones. Medicine 2016, 95, e3679. [Google Scholar] [CrossRef] [PubMed]
- Gómez, E.; Blanca-Lopez, N.; Torres, M.J.; Requena, G.; Rondon, C.; Canto, G.; Blanca, M.; Mayorga, C. Immunogloblin E-mediated immediate allergic reactions to dipyrone: Value of basophil activation test in the identification of patients. Clin. Exp. Allergy 2009, 39, 1217–1224. [Google Scholar] [CrossRef]
- Laguna, J.J.; Bogas, G.; Salas, M.; Mayorga, C.; Dionicio, J.; Gonzalez-Mendiola, R.; Ariza, A.; Fernández-Santamaría, R.; Olazabal, I.; Doña, I.; et al. The Basophil Activation Test Can Be of Value for Diagnosing Immediate Allergic Reactions to Omeprazole. J. Allergy Clin. Immunol. Pr. 2018, 6, 1628–1636.e2. [Google Scholar] [CrossRef] [PubMed]
- Salas, M.; Fernández-Santamaría, R.; Mayorga, C.; Barrionuevo, E.; Ariza, A.; Posadas, T.; Laguna, J.J.; Montañez, M.I.; Molina, N.; Fernández, T.D.; et al. Use of the Basophil Activation Test May Reduce the Need for Drug Provocation in Amoxicillin-Clavulanic Allergy. J. Allergy Clin. Immunol. Pr. 2018, 6, 1010–1018.e2. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.J.; Padial, A.; Mayorga, C.; Fernandez, T.D.; Sanchez-Sabate, E.; Cornejo-García, J.A.; Antunez, C.; Blanca, M. The diagnostic interpretation of basophil activation test in immediate allergic reactions to betalactams. Clin. Exp. Allergy 2004, 34, 1768–1775. [Google Scholar] [CrossRef]
- Bokanovic, D.; Arzt-Gradwohl, L.; Schwarz, I.; Schrautzer, C.; Laipold, K.; Aberer, W.; Binder, B.; Sturm, G.J. Possible utility of basophil activation test in dual honeybee and vespid sensitization. J. Allergy Clin. Immunol. Pr. 2020, 8, 392–394.e5. [Google Scholar] [CrossRef]
- Erdmann, S.M.; Sachs, B.; Kwiecien, R.; Moll-Slodowy, S.; Sauer, I.; Merk, H.F. The basophil activation test in wasp venom allergy: Sensitivity, specificity and monitoring specific immunotherapy. Allergy 2004, 59, 1102–1109. [Google Scholar] [CrossRef]
- Sturm, E.; Bohm, E.; Trummer, M.; Weiglhofer, I.; Heinemann, A.; Aberer, W. The CD63 basophil activation test in Hymenoptera venom allergy: A prospective study. Allergy 2004, 59, 1110–1117. [Google Scholar] [CrossRef]
- Hemmings, O.; Kwok, M.; McKendry, R.; Santos, A.F. Basophil Activation Test: Old and New Applications in Allergy. Curr. Allergy Asthma Rep. 2018, 18, 1–12. [Google Scholar] [CrossRef]
- Jensen-Jarolim, E.; Bax, H.J.; Bianchini, R.; Capron, M.; Corrigan, C.; Castells, M.; Dombrowicz, D.; Daniels-Wells, T.R.; Fazekas, J.; Fiebiger, E.; et al. AllergoOncology—the impact of allergy in oncology: EAACI position paper. Allergy 2017, 72, 866–887. [Google Scholar] [CrossRef] [PubMed]
- Jensen-Jarolim, E.; Bax, H.J.; Bianchini, R.; Crescioli, S.; Daniels-Wells, T.R.; Dombrowicz, D.; Fiebiger, E.; Gould, H.J.; Irshad, S.; Janda, J.; et al. AllergoOncology: Opposite outcomes of immune tolerance in allergy and cancer. Allergy 2018, 73, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Jean-Luc, H. Use of Basophil Activation Test in a Case of Oxaliplatin ypersensitivity. J. Allergy Ther. 2011, 02, 1–3. [Google Scholar] [CrossRef]
- Iwamoto, T.; Hirai, H.; Yamaguchi, N.; Kobayashi, N.; Sugimoto, H.; Tabata, T.; Okuda, M. Carboplatin-induced severe hypersensitivity reaction: Role of IgE-dependent basophil activation and FcεRI. Cancer Sci. 2014, 105, 1472–1479. [Google Scholar] [CrossRef]
- Wilson, J.M.; Platts-Mills, T.A. IgE to galactose-α-1,3-galactose and the α-Gal syndrome: Insights from basophil activation testing. J. Allergy Clin. Immunol. 2019, 143, 101–103. [Google Scholar] [CrossRef]
- Ilieva, K.M.; Fazekas-Singer, J.; Bax, H.J.; Crescioli, S.; Montero-Morales, L.; Mele, S.; Sow, H.S.; Stavraka, C.; Josephs, D.H.; Spicer, J.; et al. AllergoOncology: Expression platform development and functional profiling of an anti-HER2 IgE antibody. Allergy 2019, 74, 1985–1989. [Google Scholar] [CrossRef]
- Bax, H.J.; Khiabany, A.; Stavraka, C.; Pellizzari, G.; Hak, C.C.W.; Robinson, A.; Ilieva, K.M.; Woodman, N.; Naceur-Lombardelli, C.; Gillett, C.; et al. Basophil activation test in cancer patient blood evaluating potential hypersensitivity to an anti-tumor IgE therapeutic candidate. Allergy 2020, 75, 2069–2073. [Google Scholar] [CrossRef]
- Sánchez, L.D.L.V.; Alenazy, L.A.; Garcia-Neuer, M.; Castells, M.C. Drug Hypersensitivity and Desensitizations: Mechanisms and New Approaches. Int. J. Mol. Sci. 2017, 18, 1316. [Google Scholar] [CrossRef]
- Giavina-Bianchi, P.; Galvão, V.R.; Picard, M.; Caiado, J.; Castells, M.C. Basophil Activation Test is a Relevant Biomarker of the Outcome of Rapid Desensitization in Platinum Compounds-Allergy. J. Allergy Clin. Immunol. Pr. 2017, 5, 728–736. [Google Scholar] [CrossRef]
- Iwamoto, T.; Sugimoto, H.; Tabata, T.; Okuda, M. Clinical Utility of Basophil CD203c as a Biomarker for Predicting the Timing of Hypersensitivity Reaction in Carboplatin Rechallenge: Three Case Reports. Clin. Ther. 2016, 38, 1537–1541. [Google Scholar] [CrossRef]
- Iwamoto, T.; Yuta, A.; Tabata, T.; Sugimoto, H.; Gabazza, E.C.; Hirai, H.; Kojima, S.; Okuda, M. Evaluation of basophil CD203c as a predictor of carboplatin-related hypersensitivity reaction in patients with gynecologic cancer. Biol. Pharm. Bull. 2012, 35, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Ornelas, C.; Caiado, J.; Melo, A.C.; Barbosa, M.P.; Castells, M.C.; Dos Santos, M.C.P. The Contribution of the Basophil Activation Test to the Diagnosis of Hypersensitivity Reactions to Oxaliplatin. Int. Arch. Allergy Immunol. 2018, 177, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Caiado, J.; Castells, M.C. Drug Desensitizations for Chemotherapy: Safety and Efficacy in Preventing Anaphylaxis. Curr. Allergy Asthma Rep. 2021, 21, 1–13. [Google Scholar] [CrossRef]
- González-De-Olano, D.; Morgado, J.M.; Juárez-Guerrero, R.; Sanchez-Muñoz, L.; Letellez-Fernández, J.; Malón-Giménez, D.; Castells, M.C. Positive basophil activation test following anaphylaxis to pertuzumab and successful treatment with rapid desensitization. J. Allergy Clin. Immunol. Pr. 2016, 4, 338–340. [Google Scholar] [CrossRef] [PubMed]
- Markman, M.; Kennedy, A.; Webster, K.; Elson, P.; Peterson, G.; Kulp, B.; Belinson, J. Clinical Features of Hypersensitivity Reactions to Carboplatin. J. Clin. Oncol. 1999, 17, 1141. [Google Scholar] [CrossRef] [PubMed]
- Makrilia, N.; Syrigou, E.; Kaklamanos, I.; Manolopoulos, L.; Saif, M.W. Hypersensitivity Reactions Associated with Platinum Antineoplastic Agents: A Systematic Review. Met. Drugs 2010, 2010, 1–11. [Google Scholar] [CrossRef]
- Gadducci, A.; Tana, R.; Teti, G.; Zanca, G.; Fanucchi, A.; Genazzani, A. Analysis of the pattern of hypersensitivity reactions in patients receiving carboplatin retreatment for recurrent ovarian cancer. Int. J. Gynecol. Cancer 2008, 18, 615–620. [Google Scholar] [CrossRef]
- Giavina-Bianchi, P.; Aun, M.V.; Galvão, V.R.; Castells, M. Rapid Desensitization in Immediate Hypersensitivity Reaction to Drugs. Curr. Treat. Options Allergy 2015, 2, 268–285. [Google Scholar] [CrossRef]
- Josephs, D.H.; Bax, H.; Dodev, T.; Georgouli, M.; Nakamura, M.; Pellizzari, G.; Saul, L.; Karagiannis, P.; Cheung, A.; Herraiz, C.; et al. Anti-Folate Receptor-α IgE but not IgG Recruits Macrophages to Attack Tumors via TNFα/MCP-1 Signaling. Cancer Res. 2017, 77, 1127–1141. [Google Scholar] [CrossRef]
- Josephs, D.H.; Nakamura, M.; Bax, H.; Dodev, T.S.; Muirhead, G.; Saul, L.; Karagiannis, P.; Ilieva, K.M.; Crescioli, S.; Gazinska, P.; et al. An immunologically relevant rodent model demonstrates safety of therapy using a tumour-specific IgE. Allergy 2018, 73, 2328–2341. [Google Scholar] [CrossRef]
- Nakamura, M.; Souri, E.; Osborn, G.; Laddach, R.; Chauhan, J.; Stavraka, C.; Lombardi, S.; Black, A.; Khiabany, A.; Khair, D.; et al. IgE Activates Monocytes from Cancer Patients to Acquire a Pro-Inflammatory Phenotype. Cancers 2020, 12, 3376. [Google Scholar] [CrossRef] [PubMed]
- Pellizzari, G.; Hoskin, C.; Crescioli, S.; Mele, S.; Gotovina, J.; Chiaruttini, G.; Bianchini, R.; Ilieva, K.; Bax, H.J.; Papa, S.; et al. IgE re-programs alternatively-activated human macrophages towards pro-inflammatory anti-tumoural states. EBioMedicine 2019, 43, 67–81. [Google Scholar] [CrossRef]
- Pellizzari, G.; Martinez, O.; Crescioli, S.; Page, R.; Di Meo, A.; Mele, S.; Chiaruttini, G.; Hoinka, J.; Batruch, I.; Prassas, I.; et al. Immunotherapy using IgE or CAR T cells for cancers expressing the tumor antigen SLC3A. J. Immunother. Cancer 2021, 9, e002140. [Google Scholar] [CrossRef] [PubMed]
- Spicer, J.; Basu, B.; Montes, A.; Banerji, U.; Kristeleit, R.; Veal, G.J.; Corrigan, C.; Till, S.; Nintos, G.; Brier, T.; et al. Abstract CT141: Phase 1 trial of MOv18, a first-in-class IgE antibody therapy for cancer. Tumor Biol. 2020, 80, CT141. [Google Scholar] [CrossRef]
- Chauhan, J.; McCraw, A.; Nakamura, M.; Osborn, G.; Sow, H.; Cox, V.; Stavraka, C.; Josephs, D.; Spicer, J.; Karagiannis, S.; et al. IgE Antibodies against Cancer: Efficacy and Safety. Antibodies 2020, 9, 55. [Google Scholar] [CrossRef]
- Hayes, M.D.; Ward, S.; Crawford, G.; Seoane, R.C.; Jackson, W.; Kipling, D.; Voehringer, D.; Dunn-Walters, D.; Strid, J. Inflammation-induced IgE promotes epithelial hyperplasia and tumour growth. eLife 2020, 9, 9. [Google Scholar] [CrossRef]
- Marone, G.; Gambardella, A.R.; Mattei, F.; Mancini, J.; Schiavoni, G.; Varricchi, G. Basophils in Tumor Microenvironment and Surroundings. Adv. Exp. Med. Biol. 2020, 1224, 21–34. [Google Scholar] [CrossRef]
- Gooch, J.L.; Lee, A.V.; Yee, D. Interleukin 4 inhibits growth and induces apoptosis in human breast cancer cells. Cancer Res. 1998, 58, 4199–4205. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chauhan, J.; Stavraka, C.; Grandits, M.; Palhares, L.C.G.F.; Josephs, D.H.; Lacy, K.E.; Spicer, J.; Bax, H.J.; Karagiannis, S.N. Clinical and Translational Significance of Basophils in Patients with Cancer. Cells 2022, 11, 438. https://doi.org/10.3390/cells11030438
Chauhan J, Stavraka C, Grandits M, Palhares LCGF, Josephs DH, Lacy KE, Spicer J, Bax HJ, Karagiannis SN. Clinical and Translational Significance of Basophils in Patients with Cancer. Cells. 2022; 11(3):438. https://doi.org/10.3390/cells11030438
Chicago/Turabian StyleChauhan, Jitesh, Chara Stavraka, Melanie Grandits, Lais C. G. F. Palhares, Debra H. Josephs, Katie E. Lacy, James Spicer, Heather J. Bax, and Sophia N. Karagiannis. 2022. "Clinical and Translational Significance of Basophils in Patients with Cancer" Cells 11, no. 3: 438. https://doi.org/10.3390/cells11030438
APA StyleChauhan, J., Stavraka, C., Grandits, M., Palhares, L. C. G. F., Josephs, D. H., Lacy, K. E., Spicer, J., Bax, H. J., & Karagiannis, S. N. (2022). Clinical and Translational Significance of Basophils in Patients with Cancer. Cells, 11(3), 438. https://doi.org/10.3390/cells11030438