
Ectopic Tumor VCAM-1 Expression in Cancer Metastasis and Therapy Resistance


{kind=link}
{kind=link}
Abstract
1. Introduction
2. Targeting VCAM-1/VLA-4 Interaction in Osteosarcoma
3. Expression and Biological Function of VCAM-1
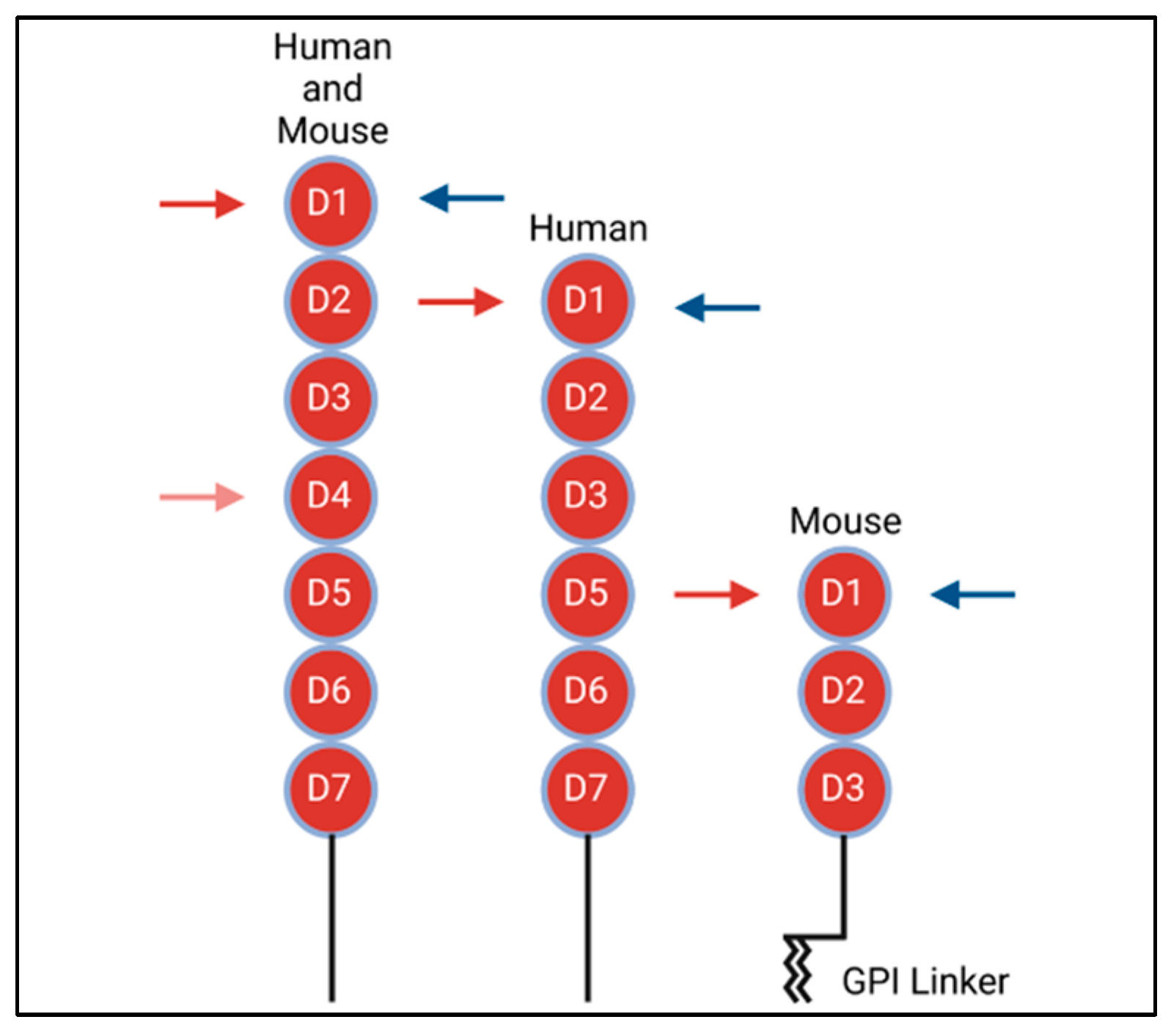
4. VCAM-1 Structure and Binding to Integrin
5. VCAM-1 in Tumor Metastatic Potential of Adult and Pediatric Cancers
6. sVCAM-1 in Tumor Metastatic Potential in Adult-Onset Cancers
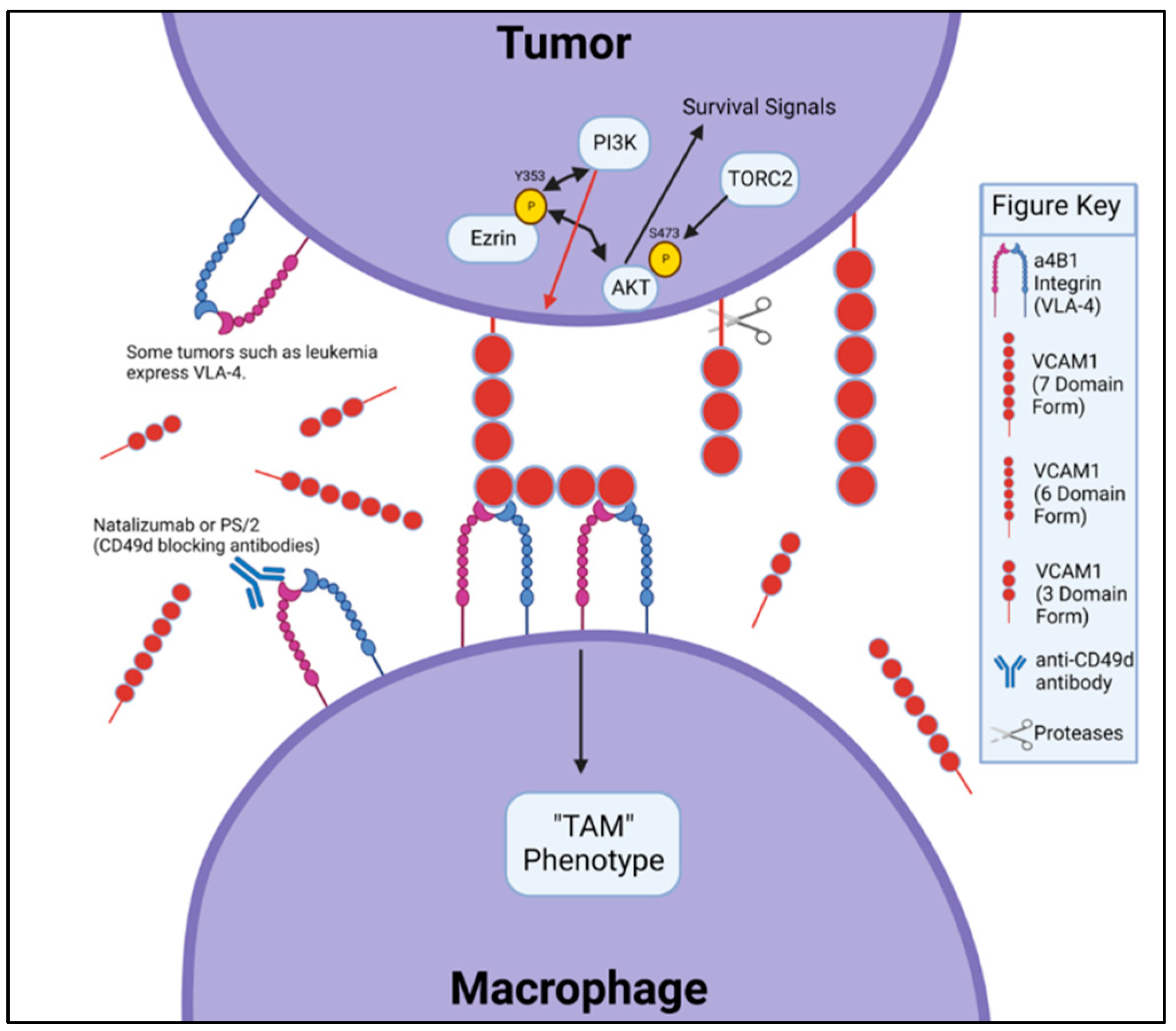
7. Role of α4β1 (VLA-4) in Metastatic Potential of Adult and Pediatric Cancers
8. Disruption of VCAM-1/VLA-4 Interaction as a Therapeutic Target in the Pediatric, Adolescent and Young Adult (AYA) Population
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bacci, G.; Briccoli, A.; Longhi, A.; Ferrari, S.; Mercuri, M.; Faggioli, F.; Versari, M.; Picci, P. Treatment and outcome of recurrent osteosarcoma: Experience at Rizzoli in 235 patients initially treated with neoadjuvant chemotherapy. Acta Oncol. 2005, 44, 748–755. [Google Scholar] [CrossRef] [PubMed]
- Rice, G.E.; Bevilacqua, M.P. An inducible endothelial cell surface glycoprotein mediates melanoma adhesion. Science 1989, 246, 1303–1306. [Google Scholar] [CrossRef] [PubMed]
- Osborn, L.; Hession, C.; Tizard, R.; Vassallo, C.; Luhowskyj, S.; Chi-Rosso, G.; Lobb, R. Direct expression cloning of vascular cell adhesion molecule 1, a cytokine-induced endothelial protein that binds to lymphocytes. Cell 1989, 59, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Alon, R.; Kassner, P.D.; Carr, M.W.; Finger, E.B.; Hemler, M.E.; Springer, T.A. The integrin VLA-4 supports tethering and rolling in flow on VCAM-1. J. Cell Biol. 1995, 128, 1243–1253. [Google Scholar] [CrossRef]
- Cerutti, C.; Ridley, A.J. Endothelial cell-cell adhesion and signaling. Exp. Cell Res. 2017, 358, 31–38. [Google Scholar] [CrossRef]
- Kong, D.H.; Kim, Y.K.; Kim, M.R.; Jang, J.H.; Lee, S. Emerging Roles of Vascular Cell Adhesion Molecule-1 (VCAM-1) in Immunological Disorders and Cancer. Int. J. Mol. Sci. 2018, 19, 1057. [Google Scholar] [CrossRef]
- Freedman, A.S.; Munro, J.M.; Rice, G.E.; Bevilacqua, M.P.; Morimoto, C.; McIntyre, B.W.; Rhynhart, K.; Pober, J.S.; Nadler, L.M. Adhesion of human B cells to germinal centers in vitro involves VLA-4 and INCAM-110. Science 1990, 249, 1030–1033. [Google Scholar] [CrossRef]
- Oleszowsky, M.; Seidel, M.F. Serum Soluble Vascular Cell Adhesion Molecule-1 Overexpression Is a Disease Marker in Patients with First-Time Diagnosed Antinuclear Antibodies: A Prospective, Observational Pilot Study. BioMed Res. Int. 2018, 2018, 8286067. [Google Scholar] [CrossRef]
- Banks, R.E.; Gearing, A.J.; Hemingway, I.K.; Norfolk, D.R.; Perren, T.J.; Selby, P.J. Circulating intercellular adhesion molecule-1 (ICAM-1), E-selectin and vascular cell adhesion molecule-1 (VCAM-1) in human malignancies. Br. J. Cancer 1993, 68, 122–124. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, X.H.; Massague, J. Macrophage binding to receptor VCAM-1 transmits survival signals in breast cancer cells that invade the lungs. Cancer Cell 2011, 20, 538–549. [Google Scholar] [CrossRef]
- Ding, Y.B.; Chen, G.Y.; Xia, J.G.; Zang, X.W.; Yang, H.Y.; Yang, L. Association of VCAM-1 overexpression with oncogenesis, tumor angiogenesis and metastasis of gastric carcinoma. World J. Gastroenterol. 2003, 9, 1409–1414. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.Y.; Lu, D.; Hung, C.F.; Peng, S.; Huang, L.; Jie, C.; Murillo, F.; Rowley, J.; Tsai, Y.C.; He, L.; et al. Ectopic expression of vascular cell adhesion molecule-1 as a new mechanism for tumor immune evasion. Cancer Res. 2007, 67, 1832–1841. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Mu, E.; Wei, Y.; Riethdorf, S.; Yang, Q.; Yuan, M.; Yan, J.; Hua, Y.; Tiede, B.J.; Lu, X.; et al. VCAM-1 promotes osteolytic expansion of indolent bone micrometastasis of breast cancer by engaging alpha4beta1-positive osteoclast progenitors. Cancer Cell 2011, 20, 701–714. [Google Scholar] [CrossRef] [PubMed]
- Slack-Davis, J.K.; Atkins, K.A.; Harrer, C.; Hershey, E.D.; Conaway, M. Vascular cell adhesion molecule-1 is a regulator of ovarian cancer peritoneal metastasis. Cancer Res. 2009, 69, 1469–1476. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.G.; Dai, X.Y.; Kozak, C.A.; Mims, M.P.; Gotto, A.M.; Ballantyne, C.M. Murine VCAM-1. Molecular cloning, mapping, and analysis of a truncated form. J. Immunol. 1994, 153, 4088–4098. [Google Scholar] [PubMed]
- Cybulsky, M.I.; Fries, J.W.; Williams, A.J.; Sultan, P.; Davis, V.M.; Gimbrone, M.A., Jr.; Collins, T. Alternative splicing of human VCAM-1 in activated vascular endothelium. Am. J. Pathol. 1991, 138, 815–820. [Google Scholar]
- Luo, B.H.; Carman, C.V.; Springer, T.A. Structural basis of integrin regulation and signaling. Annu. Rev. Immunol. 2007, 25, 619–647. [Google Scholar] [CrossRef]
- Xiao, T.; Takagi, J.; Coller, B.S.; Wang, J.H.; Springer, T.A. Structural basis for allostery in integrins and binding to fibrinogen-mimetic therapeutics. Nature 2004, 432, 59–67. [Google Scholar] [CrossRef]
- Takada, Y.; Elices, M.J.; Crouse, C.; Hemler, M.E. The primary structure of the alpha 4 subunit of VLA-4: Homology to other integrins and a possible cell-cell adhesion function. EMBO J. 1989, 8, 1361–1368. [Google Scholar] [CrossRef]
- Kilger, G.; Needham, L.A.; Nielsen, P.J.; Clements, J.; Vestweber, D.; Holzmann, B. Differential regulation of alpha 4 integrin-dependent binding to domains 1 and 4 of vascular cell adhesion molecule-1. J. Biol. Chem. 1995, 270, 5979–5984. [Google Scholar] [CrossRef]
- Sheppard, A.M.; Onken, M.D.; Rosen, G.D.; Noakes, P.G.; Dean, D.C. Expanding roles for alpha 4 integrin and its ligands in development. Cell Adhes. Commun. 1994, 2, 27–43. [Google Scholar] [CrossRef]
- Kamata, T.; Puzon, W.; Takada, Y. Identification of putative ligand-binding sites of the integrin alpha 4 beta 1 (VLA-4, CD49d/CD29). Biochem. J. 1995, 305 Pt 3, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Schurpf, T.; Springer, T.A. How natalizumab binds and antagonizes alpha4 integrins. J. Biol. Chem. 2013, 288, 32314–32325. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.S.; Lin, H.Y.; Lai, S.W.; Huang, C.Y.; Huang, B.R.; Chen, P.Y.; Wei, K.C.; Lu, D.Y. MiR-181b modulates EGFR-dependent VCAM-1 expression and monocyte adhesion in glioblastoma. Oncogene 2017, 36, 5006–5022. [Google Scholar] [CrossRef] [PubMed]
- Minn, A.J.; Gupta, G.P.; Siegel, P.M.; Bos, P.D.; Shu, W.; Giri, D.D.; Viale, A.; Olshen, A.B.; Gerald, W.L.; Massague, J. Genes that mediate breast cancer metastasis to lung. Nature 2005, 436, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Scalici, J.M.; Arapovic, S.; Saks, E.J.; Atkins, K.A.; Petroni, G.; Duska, L.R.; Slack-Davis, J.K. Mesothelium expression of vascular cell adhesion molecule-1 (VCAM-1) is associated with an unfavorable prognosis in epithelial ovarian cancer (EOC). Cancer 2017, 123, 977–984. [Google Scholar] [CrossRef]
- Alexiou, D.; Karayiannakis, A.J.; Syrigos, K.N.; Zbar, A.; Kremmyda, A.; Bramis, I.; Tsigris, C. Serum levels of E-selectin, ICAM-1 and VCAM-1 in colorectal cancer patients: Correlations with clinicopathological features, patient survival and tumour surgery. Eur. J. Cancer 2001, 37, 2392–2397. [Google Scholar] [CrossRef]
- Zhang, D.; Bi, J.; Liang, Q.; Wang, S.; Zhang, L.; Han, F.; Li, S.; Qiu, B.; Fan, X.; Chen, W.; et al. VCAM1 Promotes Tumor Cell Invasion and Metastasis by Inducing EMT and Transendothelial Migration in Colorectal Cancer. Front. Oncol. 2020, 10, 1066. [Google Scholar] [CrossRef]
- Milstone, D.S.; Ilyama, M.; Chen, M.; O’Donnell, P.; Davis, V.M.; Plutzky, J.; Brown, J.D.; Haldar, S.M.; Siu, A.; Lau, A.C.; et al. Differential role of an NF-kappaB transcriptional response element in endothelial versus intimal cell VCAM-1 expression. Circ. Res. 2015, 117, 166–177. [Google Scholar] [CrossRef]
- Xia, Y.F.; Liu, L.P.; Zhong, C.P.; Geng, J.G. NF-kappaB activation for constitutive expression of VCAM-1 and ICAM-1 on B lymphocytes and plasma cells. Biochem. Biophys. Res. Commun. 2001, 289, 851–856. [Google Scholar] [CrossRef]
- Li, P.; Sanz, I.; O’Keefe, R.J.; Schwarz, E.M. NF-kappa B regulates VCAM-1 expression on fibroblast-like synoviocytes. J. Immunol. 2000, 164, 5990–5997. [Google Scholar] [CrossRef] [PubMed]
- Condeelis, J.; Pollard, J.W. Macrophages: Obligate partners for tumor cell migration, invasion, and metastasis. Cell 2006, 124, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Linde, N.; Casanova-Acebes, M.; Sosa, M.S.; Mortha, A.; Rahman, A.; Farias, E.; Harper, K.; Tardio, E.; Reyes Torres, I.; Jones, J.; et al. Macrophages orchestrate breast cancer early dissemination and metastasis. Nat. Commun. 2018, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Su, X.; Jiang, X.; Zhang, T.; Min, I.; Ding, Y.; Wang, X.; Mao, Z.; Cao, J.; Teng, X.; et al. VCAM-1 Upregulation Contributes to Insensitivity of Vemurafenib in BRAF-Mutant Thyroid Cancer. Transl. Oncol. 2020, 13, 441–451. [Google Scholar] [CrossRef]
- Pinho, S.; Wei, Q.; Maryanovich, M.; Zhang, D.; Balandran, J.C.; Pierce, H.; Nakahara, F.; Di Staulo, A.; Bartholdy, B.A.; Xu, J.; et al. VCAM1 confers innate immune tolerance on haematopoietic and leukaemic stem cells. Nat. Cell Biol. 2022, 24, 290–298. [Google Scholar] [CrossRef]
- Garton, K.J.; Gough, P.J.; Philalay, J.; Wille, P.T.; Blobel, C.P.; Whitehead, R.H.; Dempsey, P.J.; Raines, E.W. Stimulated shedding of vascular cell adhesion molecule 1 (VCAM-1) is mediated by tumor necrosis factor-alpha-converting enzyme (ADAM 17). J. Biol. Chem. 2003, 278, 37459–37464. [Google Scholar] [CrossRef]
- Guaiquil, V.; Swendeman, S.; Yoshida, T.; Chavala, S.; Campochiaro, P.A.; Blobel, C.P. ADAM9 is involved in pathological retinal neovascularization. Mol. Cell Biol. 2009, 29, 2694–2703. [Google Scholar] [CrossRef]
- Matsuno, O.; Miyazaki, E.; Nureki, S.; Ueno, T.; Kumamoto, T.; Higuchi, Y. Role of ADAM8 in experimental asthma. Immunol. Lett. 2006, 102, 67–73. [Google Scholar] [CrossRef]
- Matsuno, O.; Miyazaki, E.; Nureki, S.; Ueno, T.; Ando, M.; Ito, K.; Kumamoto, T.; Higuchi, Y. Elevated soluble ADAM8 in bronchoalveolar lavage fluid in patients with eosinophilic pneumonia. Int. Arch. Allergy Immunol. 2007, 142, 285–290. [Google Scholar] [CrossRef]
- Peduto, L. ADAM9 as a potential target molecule in cancer. Curr. Pharm. Des. 2009, 15, 2282–2287. [Google Scholar] [CrossRef]
- Silva, H.C.; Garcao, F.; Coutinho, E.C.; De Oliveira, C.F.; Regateiro, F.J. Soluble VCAM-1 and E-selectin in breast cancer: Relationship with staging and with the detection of circulating cancer cells. Neoplasma 2006, 53, 538–543. [Google Scholar]
- Okugawa, Y.; Miki, C.; Toiyama, Y.; Koike, Y.; Inoue, Y.; Kusunoki, M. Serum level of soluble vascular cell adhesion molecule 1 is a valuable prognostic marker in colorectal carcinoma. Dis. Colon Rectum 2009, 52, 1330–1336. [Google Scholar] [CrossRef] [PubMed]
- Tempia-Caliera, A.A.; Horvath, L.Z.; Zimmermann, A.; Tihanyi, T.T.; Korc, M.; Friess, H.; Buchler, M.W. Adhesion molecules in human pancreatic cancer. J. Surg. Oncol. 2002, 79, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Zhou, Q.; Zheng, S.; Li, G.; Lin, Q.; Wei, L.; Fu, Z.; Zhang, B.; Liu, Y.; Li, Z.; et al. Tumor-associated macrophages promote progression and the Warburg effect via CCL18/NF-kB/VCAM-1 pathway in pancreatic ductal adenocarcinoma. Cell Death Dis. 2018, 9, 453. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, R.; Ijichi, H.; Sano, M.; Miyabayashi, K.; Mohri, D.; Kim, J.; Kimura, G.; Nakatsuka, T.; Fujiwara, H.; Yamamoto, K.; et al. Soluble VCAM-1 promotes gemcitabine resistance via macrophage infiltration and predicts therapeutic response in pancreatic cancer. Sci. Rep. 2020, 10, 21194. [Google Scholar] [CrossRef]
- Schlesinger, M.; Bendas, G. Contribution of very late antigen-4 (VLA-4) integrin to cancer progression and metastasis. Cancer Metastasis Rev. 2015, 34, 575–591. [Google Scholar] [CrossRef]
- Kil, S.H.; Krull, C.E.; Cann, G.; Clegg, D.; Bronner-Fraser, M. The alpha4 subunit of integrin is important for neural crest cell migration. Dev. Biol. 1998, 202, 29–42. [Google Scholar] [CrossRef]
- Testaz, S.; Duband, J.L. Central role of the alpha4beta1 integrin in the coordination of avian truncal neural crest cell adhesion, migration, and survival. Dev. Dyn. 2001, 222, 127–140. [Google Scholar] [CrossRef]
- Young, S.A.; McCabe, K.E.; Bartakova, A.; Delaney, J.; Pizzo, D.P.; Newbury, R.O.; Varner, J.A.; Schlaepfer, D.D.; Stupack, D.G. Integrin alpha4 Enhances Metastasis and May Be Associated with Poor Prognosis in MYCN-low Neuroblastoma. PLoS ONE 2015, 10, e0120815. [Google Scholar] [CrossRef]
- Paavonen, T.; Tiisala, S.; Majuri, M.L.; Bohling, T.; Renkonen, R. In vivo evidence of the role of alpha 4 beta 1-VCAM-1 interaction in sarcoma, but not in carcinoma extravasation. Int. J. Cancer 1994, 58, 298–302. [Google Scholar] [CrossRef]
- Prifti, S.; Zourab, Y.; Koumouridis, A.; Bohlmann, M.; Strowitzki, T.; Rabe, T. Role of integrins in invasion of endometrial cancer cell lines. Gynecol. Oncol. 2002, 84, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Redondo-Munoz, J.; Escobar-Diaz, E.; Samaniego, R.; Terol, M.J.; Garcia-Marco, J.A.; Garcia-Pardo, A. MMP-9 in B-cell chronic lymphocytic leukemia is up-regulated by alpha4beta1 integrin or CXCR4 engagement via distinct signaling pathways, localizes to podosomes, and is involved in cell invasion and migration. Blood 2006, 108, 3143–3151. [Google Scholar] [CrossRef] [PubMed]
- Schadendorf, D.; Gawlik, C.; Haney, U.; Ostmeier, H.; Suter, L.; Czarnetzki, B.M. Tumour progression and metastatic behaviour in vivo correlates with integrin expression on melanocytic tumours. J. Pathol. 1993, 170, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Bernard-Trifilo, J.A.; Lim, Y.; Lim, S.T.; Mitra, S.K.; Uryu, S.; Chen, M.; Pallen, C.J.; Cheung, N.K.; Mikolon, D.; et al. Distinct FAK-Src activation events promote alpha5beta1 and alpha4beta1 integrin-stimulated neuroblastoma cell motility. Oncogene 2008, 27, 1439–1448. [Google Scholar] [CrossRef]
- Taichman, D.B.; Cybulsky, M.I.; Djaffar, I.; Longenecker, B.M.; Teixido, J.; Rice, G.E.; Aruffo, A.; Bevilacqua, M.P. Tumor cell surface alpha 4 beta 1 integrin mediates adhesion to vascular endothelium: Demonstration of an interaction with the N-terminal domains of INCAM-110/VCAM-1. Cell Regul. 1991, 2, 347–355. [Google Scholar] [CrossRef]
- Jacamo, R.; Chen, Y.; Wang, Z.; Ma, W.; Zhang, M.; Spaeth, E.L.; Wang, Y.; Battula, V.L.; Mak, P.Y.; Schallmoser, K.; et al. Reciprocal leukemia-stroma VCAM-1/VLA-4-dependent activation of NF-kappaB mediates chemoresistance. Blood 2014, 123, 2691–2702. [Google Scholar] [CrossRef]
- Hsieh, Y.T.; Gang, E.J.; Geng, H.; Park, E.; Huantes, S.; Chudziak, D.; Dauber, K.; Schaefer, P.; Scharman, C.; Shimada, H.; et al. Integrin alpha4 blockade sensitizes drug resistant pre-B acute lymphoblastic leukemia to chemotherapy. Blood 2013, 121, 1814–1818. [Google Scholar] [CrossRef]
- Matsunaga, T.; Fukai, F.; Miura, S.; Nakane, Y.; Owaki, T.; Kodama, H.; Tanaka, M.; Nagaya, T.; Takimoto, R.; Takayama, T.; et al. Combination therapy of an anticancer drug with the FNIII14 peptide of fibronectin effectively overcomes cell adhesion-mediated drug resistance of acute myelogenous leukemia. Leukemia 2008, 22, 353–360. [Google Scholar] [CrossRef]
- Mudry, R.E.; Fortney, J.E.; York, T.; Hall, B.M.; Gibson, L.F. Stromal cells regulate survival of B-lineage leukemic cells during chemotherapy. Blood 2000, 96, 1926–1932. [Google Scholar] [CrossRef]
- Chen, Q.; Massague, J. Molecular pathways: VCAM-1 as a potential therapeutic target in metastasis. Clin. Cancer Res. 2012, 18, 5520–5525. [Google Scholar] [CrossRef]
- Schlesinger, M.; Bendas, G. Vascular cell adhesion molecule-1 (VCAM-1)--an increasing insight into its role in tumorigenicity and metastasis. Int. J. Cancer 2015, 136, 2504–2514. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger, M.; Schmitz, P.; Zeisig, R.; Naggi, A.; Torri, G.; Casu, B.; Bendas, G. The inhibition of the integrin VLA-4 in MV3 melanoma cell binding by non-anticoagulant heparin derivatives. Thromb. Res. 2012, 129, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Vanderslice, P.; Biediger, R.J.; Woodside, D.G.; Berens, K.L.; Holland, G.W.; Dixon, R.A. Development of cell adhesion molecule antagonists as therapeutics for asthma and COPD. Pulm. Pharmacol. Ther. 2004, 17, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yeh, E.A.; Weinstock-Guttman, B. Natalizumab in pediatric multiple sclerosis patients. Ther. Adv. Neurol. Disord. 2010, 3, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Alroughani, R.; Ahmed, S.F.; Behbehani, R.; Al-Hashel, J. The Use of Natalizumab in Pediatric Patients With Active Relapsing Multiple Sclerosis: A Prospective Study. Pediatr. Neurol. 2017, 70, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Arnal-Garcia, C.; Garcia-Montero, M.R.; Malaga, I.; Millan-Pascual, J.; Oliva-Nacarino, P.; Ramio-Torrenta, L.; Oreja-Guevara, C. Natalizumab use in pediatric patients with relapsing-remitting multiple sclerosis. Eur. J. Paediatr. Neurol. 2013, 17, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Ghezzi, A.; Pozzilli, C.; Grimaldi, L.M.; Brescia Morra, V.; Bortolon, F.; Capra, R.; Filippi, M.; Moiola, L.; Rocca, M.A.; Rottoli, M.; et al. Safety and efficacy of natalizumab in children with multiple sclerosis. Neurology 2010, 75, 912–917. [Google Scholar] [CrossRef]
- Mattila, P.; Majuri, M.L.; Renkonen, R. VLA-4 integrin on sarcoma cell lines recognizes endothelial VCAM-1. Differential regulation of the VLA-4 avidity on various sarcoma cell lines. Int. J. Cancer 1992, 52, 918–923. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
VanHeyst, K.A.; Choi, S.H.; Kingsley, D.T.; Huang, A.Y. Ectopic Tumor VCAM-1 Expression in Cancer Metastasis and Therapy Resistance. Cells 2022, 11, 3922. https://doi.org/10.3390/cells11233922
VanHeyst KA, Choi SH, Kingsley DT, Huang AY. Ectopic Tumor VCAM-1 Expression in Cancer Metastasis and Therapy Resistance. Cells. 2022; 11(23):3922. https://doi.org/10.3390/cells11233922
Chicago/Turabian StyleVanHeyst, Kristen A., Sung Hee Choi, Daniel T. Kingsley, and Alex Y. Huang. 2022. "Ectopic Tumor VCAM-1 Expression in Cancer Metastasis and Therapy Resistance" Cells 11, no. 23: 3922. https://doi.org/10.3390/cells11233922
APA StyleVanHeyst, K. A., Choi, S. H., Kingsley, D. T., & Huang, A. Y. (2022). Ectopic Tumor VCAM-1 Expression in Cancer Metastasis and Therapy Resistance. Cells, 11(23), 3922. https://doi.org/10.3390/cells11233922