

The Expression Pattern of Adhesion G Protein-Coupled Receptor F5 Is Related to Cell Adhesion and Metastatic Pathways in Colorectal Cancer—Comprehensive Study Based on In Silico Analysis



Abstract
1. Introduction
2. Materials and Methods
2.1. Datasets and Patient Characteristics
2.2. Correlation Analysis
2.3. Pathway Enrichment Analysis
2.4. Gene Expression Profiling
2.5. Survival Analysis
2.6. Immune Cell Infiltration Analysis
2.7. Statistical Analysis
3. Results
3.1. ADGRF5 Is Overexpressed in the Colon of Patients with Colorectal Cancer
3.2. ADGRF5 Expression Is Related to Clinical Characterization of Patients with Colorectal Cancer
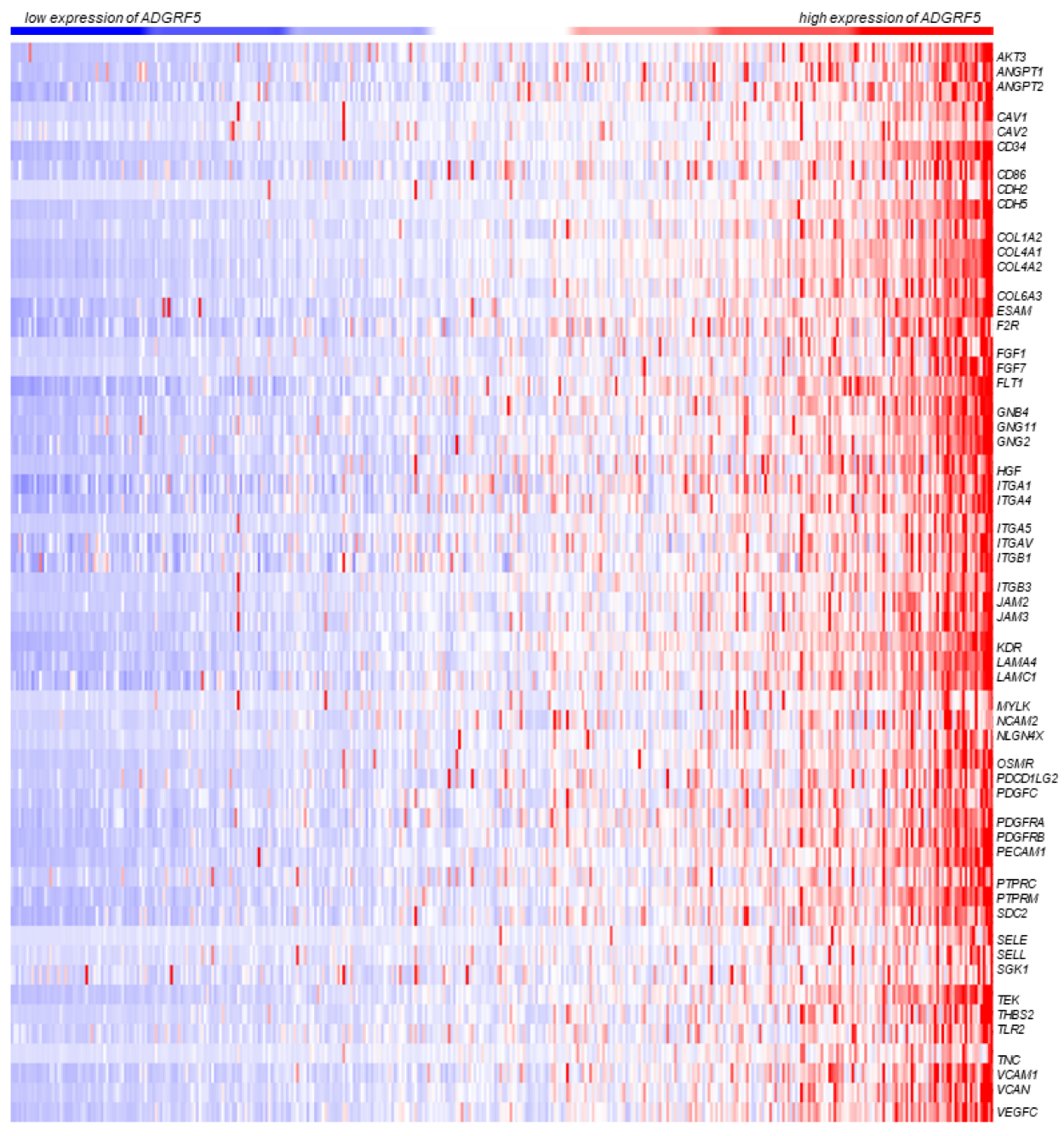
3.3. ADGRF5 Expression Is Positively Correlated with Genes Participating in Cell Adhesion and Metastatic Pathways in Colorectal Cancer
3.4. ADGRF5 Expression Is Negatively Correlated with Genes Participate in Ribosome Pathway in Colorectal Cancer
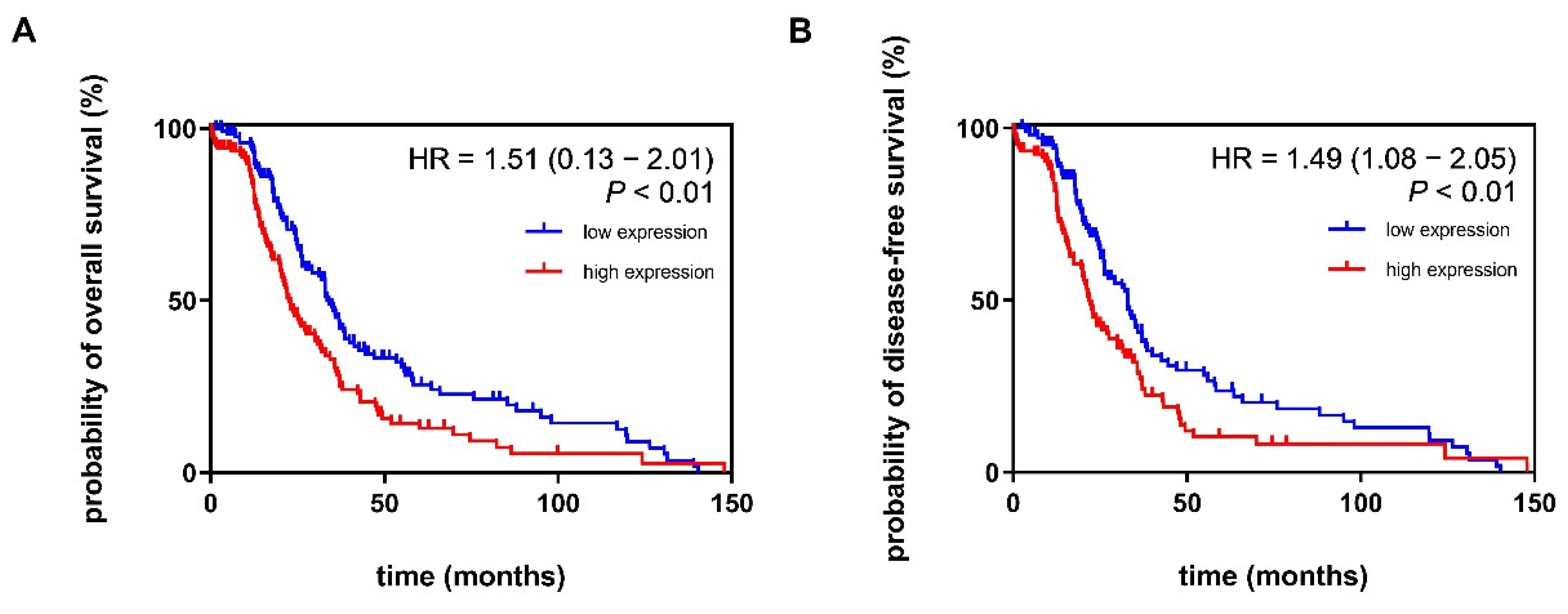
3.5. Increased Expression Pattern of ADGRF5 in the Colon of Patients with CRC Is Associated with Poor Probability of Overall Survival and Disease-Free Survival
3.6. Expression Pattern of ADGRF5 Is Correlated with the Levels of Tumor-Infiltrating Immune Cells in the Colon of Patients with CRC
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lao, V.V.; Grady, W.M. Epigenetics and colorectal cancer. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 686–700. [Google Scholar] [CrossRef]
- Dekker, E.; Tanis, P.J.; Vleugels, J.L.A.; Kasi, P.M.; Wallace, M.B. Colorectal cancer. Lancet 2019, 394, 1467–1480. [Google Scholar] [CrossRef]
- Nishihara, H.; Hwang, M.; Kizaka-Kondoh, S.; Eckmann, L.; Insel, P.A. Cyclic AMP promotes cAMP-responsive element-binding protein-dependent induction of cellular inhibitor of apoptosis protein-2 and suppresses apoptosis of colon cancer cells through ERK1/2 and p38 MAPK. J. Biol. Chem. 2004, 279, 26176–26183. [Google Scholar] [CrossRef]
- Agarwal, A.; Das, K.; Lerner, N.; Sathe, S.; Cicek, M.; Casey, G.; Sizemore, N. The AKT/I kappa B kinase pathway promotes angiogenic/metastatic gene expression in colorectal cancer by activating nuclear factor-kappa B and beta-catenin. Oncogene 2005, 24, 1021–1031. [Google Scholar] [CrossRef]
- Silva, A.L.; Dawson, S.N.; Arends, M.J.; Guttula, K.; Hall, N.; Cameron, E.A.; Huang, T.H.; Brenton, J.D.; Tavaré, S.; Bienz, M.; et al. Boosting Wnt activity during colorectal cancer progression through selective hypermethylation of Wnt signaling antagonists. BMC Cancer 2014, 14, 891. [Google Scholar] [CrossRef] [PubMed]
- Voorneveld, P.W.; Kodach, L.L.; Jacobs, R.J.; Liv, N.; Zonnevylle, A.C.; Hoogenboom, J.P.; Biemond, I.; Verspaget, H.W.; Hommes, D.W.; de Rooij, K.; et al. Loss of SMAD4 alters BMP signaling to promote colorectal cancer cell metastasis via activation of Rho and ROCK. Gastroenterology 2014, 147, 196–208.e13. [Google Scholar] [CrossRef] [PubMed]
- Fazio, C.; Piazzi, G.; Vitaglione, P.; Fogliano, V.; Munarini, A.; Prossomariti, A.; Milazzo, M.; D’Angelo, L.; Napolitano, M.; Chieco, P.; et al. Inflammation increases NOTCH1 activity via MMP9 and is counteracted by Eicosapentaenoic Acid-free fatty acid in colon cancer cells. Sci. Rep. 2016, 6, 20670. [Google Scholar] [CrossRef] [PubMed]
- Dozois, E.J.; Boardman, L.A.; Suwanthanma, W.; Limburg, P.J.; Cima, R.R.; Bakken, J.L.; Vierkant, R.A.; Aakre, J.A.; Larson, D.W. Young-onset colorectal cancer in patients with no known genetic predisposition: Can we increase early recognition and improve outcome? Medicine 2008, 87, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Hamann, J.; Aust, G.; Araç, D.; Engel, F.B.; Formstone, C.; Fredriksson, R.; Hall, R.A.; Harty, B.L.; Kirchhoff, C.; Knapp, B.; et al. International Union of Basic and Clinical Pharmacology. XCIV. Adhesion G protein-coupled receptors. Pharmacol. Rev. 2015, 67, 338–367. [Google Scholar] [CrossRef] [PubMed]
- Vizurraga, A.; Adhikari, R.; Yeung, J.; Yu, M.; Tall, G.G. Mechanisms of adhesion G protein-coupled receptor activation. J. Biol. Chem. 2020, 295, 14065–14083. [Google Scholar] [CrossRef]
- Bridges, J.P.; Ludwig, M.G.; Mueller, M.; Kinzel, B.; Sato, A.; Xu, Y.; Whitsett, J.A.; Ikegami, M. Orphan G protein-coupled receptor GPR116 regulates pulmonary surfactant pool size. Am. J. Respir. Cell Mol. Biol. 2013, 49, 348–357. [Google Scholar] [CrossRef]
- Fukuzawa, T.; Ishida, J.; Kato, A.; Ichinose, T.; Ariestanti, D.M.; Takahashi, T.; Ito, K.; Abe, J.; Suzuki, T.; Wakana, S.; et al. Lung surfactant levels are regulated by Ig-Hepta/GPR116 by monitoring surfactant protein D. PLoS ONE. 2013, 8, e69451. [Google Scholar] [CrossRef]
- Yang, M.Y.; Hilton, M.B.; Seaman, S.; Haines, D.C.; Nagashima, K.; Burks, C.M.; Tessarollo, L.; Ivanova, P.T.; Brown, H.A.; Umstead, T.M.; et al. Essential regulation of lung surfactant homeostasis by the orphan G protein-coupled receptor GPR116. Cell Rep. 2013, 3, 1457–1464. [Google Scholar] [CrossRef]
- Nie, T.; Hui, X.; Gao, X.; Li, K.; Lin, W.; Xiang, X.; Ding, M.; Kuang, Y.; Xu, A.; Fei, J.; et al. Adipose tissue deletion of Gpr116 impairs insulin sensitivity through modulation of adipose function. FEBS Lett. 2012, 586, 3618–3625. [Google Scholar] [CrossRef]
- Niaudet, C.; Hofmann, J.J.; Mäe, M.A.; Jung, B.; Gaengel, K.; Vanlandewijck, M.; Ekvärn, E.; Salvado, M.D.; Mehlem, A.; Al Sayegh, S.; et al. Gpr116 receptor regulates distinctive functions in pneumocytes and vascular endothelium. PLoS ONE 2015, 10, e0137949. [Google Scholar] [CrossRef]
- Ariestanti, D.M.; Ando, H.; Hirose, S.; Nakamura, N. Targeted disruption of Ig-Hepta/Gpr116 causes emphysema-like symptoms that are associated with alveolar macrophage activation. J. Biol. Chem. 2015, 290, 11032–11040. [Google Scholar] [CrossRef]
- Davidson, B.; Stavnes, H.T.; Risberg, B.; Nesland, J.M.; Wohlschlaeger, J.; Yang, Y.; Shih, I.M.; Wang, T.L. Gene expression signatures differentiate adenocarcinoma of lung and breast origin in effusions. Hum. Pathol. 2012, 43, 684–694. [Google Scholar] [CrossRef]
- Tang, X.; Jin, R.; Qu, G.; Wang, X.; Li, Z.; Yuan, Z.; Zhao, C.; Siwko, S.; Shi, T.; Wang, P.; et al. GPR116, an adhesion G-protein-coupled receptor, promotes breast cancer metastasis via the Gαq-p63RhoGEF-Rho GTPase pathway. Cancer Res. 2013, 73, 6206–6218. [Google Scholar] [CrossRef]
- Yang, L.; Lin, X.L.; Liang, W.; Fu, S.W.; Lin, W.F.; Tian, X.Q.; Gao, Y.J.; Chen, H.Y.; Dai, J.; Ge, Z.Z. High expression of GPR116 indicates poor survival outcome and promotes tumor progression in colorectal carcinoma. Oncotarget 2017, 8, 47943–47956. [Google Scholar] [CrossRef]
- Wuensch, T.; Wizenty, J.; Quint, J.; Spitz, W.; Bosma, M.; Becker, O.; Adler, A.; Veltzke-Schlieker, W.; Stockmann, M.; Weiss, S.; et al. Expression analysis of Fibronectin Type III Domain-Containing (FNDC) genes in inflammatory bowel disease and colorectal cancer. Gastroenterol. Res. Pract. 2019, 2019, 3784172. [Google Scholar] [CrossRef]
- Wang, C.; Fan, H.Q.; Zhang, Y.W. MiR-511-5p functions as a tumor suppressor and a predictive of prognosis in colorectal cancer by directly targeting GPR116. Eur Rev. Med. Pharmacol. Sci. 2019, 23, 6119–6130. [Google Scholar] [CrossRef] [PubMed]
- Muthiah, I.; Rajendran, K.; Dhanaraj, P.; Vallinayagam, S. In silico structure prediction, molecular docking and dynamic simulation studies on G Protein-Coupled Receptor 116: A novel insight into breast cancer therapy. J. Biomol. Struct. Dyn. 2020, 1, 9. [Google Scholar] [CrossRef] [PubMed]
- Muthiah, I.; Rajendran, K.; Dhanaraj, P. In silico molecular docking and physicochemical property studies on effective phytochemicals targeting GPR116 for breast cancer treatment. Mol. Cell Biochem. 2021, 476, 883–896. [Google Scholar] [CrossRef] [PubMed]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef]
- Li, B.; Seversion, E.; Pignon, J.-C.; Zhao, H.; Li, T.; Novak, J.; Jiang, P.; Shen, H.; Aster, J.C.; Rodig, S.; et al. Comprehensive analyses of tumor immunity: Implications for cancer immunotherapy. Genome Biol. 2016, 17, 174. [Google Scholar] [CrossRef]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. TIMER: A web server for comprehensive analysis of tumor-infiltrating immune cells. Cancer Res. 2017, 77, 108–110. [Google Scholar] [CrossRef]
- Li, T.; Fu, J.; Zeng, Z.; Cohen, D.; Li, J.; Chen, Q.; Li, B.; Liu, X.S. TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic Acids Res. 2020, 48, 509–514. [Google Scholar] [CrossRef]
- Bosman, F.T.; Carneiro, F.; Hruban, R.H.; Theise, N.D. WHO Classification of Tumours of the Digestive System, 4th ed.; International Agency for Research on Cancer: Lyon, France, 2010; Volume 3.
- Numata, M.; Shiozawa, M.; Watanabe, T.; Tamagawa, H.; Yamamoto, N.; Morinaga, S.; Watanabe, K.; Godai, T.; Oshima, T.; Fujii, S.; et al. The clinicopathological features of colorectal mucinous adenocarcinoma and a therapeutic strategy for the disease. World J. Surg. Oncol. 2012, 10, 109. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Huh, J.W.; Park, Y.A.; Cho, Y.B.; Yun, S.H.; Kim, H.C.; Lee, W.Y.; Chun, H.K. Prognostic comparison between mucinous and nonmucinous adenocarcinoma in colorectal cancer. Medicine 2015, 94, e658. [Google Scholar] [CrossRef]
- Curtale, G.; Renzi, T.A.; Drufuca, L.; Rubino, M.; Locati, M. Glucocorticoids downregulate TLR4 signaling activity via its direct targeting by miR-511-5p. Eur. J. Immunol. 2017, 47, 2080–2089. [Google Scholar] [CrossRef]
- Chen, J.; Liu, A.; Wang, Z.; Wang, B.; Chai, X.; Lu, W.; Cao, T.; Li, R.; Wu, M.; Lu, Z.; et al. LINC00173.v1 promotes angiogenesis and progression of lung squamous cell carcinoma by sponging miR-511-5p to regulate VEGFA expression. Mol. Cancer 2020, 19, 98. [Google Scholar] [CrossRef]
- Gonzalez, D.M.; Medici, D. Signaling mechanisms of the epithelial-mesenchymal transition. Sci. Signal. 2014, 7, re8. [Google Scholar] [CrossRef]
- Johnson, S.M.; Gulhati, P.; Rampy, B.A.; Han, Y.; Rychahou, P.G.; Doan, H.Q.; Weiss, H.L.; Evers, B.M. Novel expression patterns of PI3K/Akt/mTOR signaling pathway components in colorectal cancer. J. Am. Coll. Surg. 2010, 210, 776–778. [Google Scholar] [CrossRef]
- Suman, S.; Kurisetty, V.; Das, T.P.; Vadodkar, A.; Ramos, G.; Lakshmanaswamy, R.; Damodaran, C. Activation of AKT signaling promotes epithelial-mesenchymal transition and tumor growth in colorectal cancer cells. Mol. Carcinog. 2014, 53 (Suppl. S1), E151–E160. [Google Scholar] [CrossRef]
- Grille, S.J.; Bellacosa, A.; Upson, J.; Klein-Szanto, A.J.; van Roy, F.; Lee-Kwon, W.; Donowitz, M.; Tsichlis, P.N.; Larue, L. The protein kinase Akt induces epithelial mesenchymal transition and promotes enhanced motility and invasiveness of squamous cell carcinoma lines. Cancer Res. 2003, 63, 2172–2178. [Google Scholar]
- Petrova, Y.I.; Schecterson, L.; Gumbiner, B.M. Roles for E-cadherin cell surface regulation in cancer. Mol. Biol. Cell 2016, 27, 3233–3244. [Google Scholar] [CrossRef]
- Fischer, H.; Stenling, R.; Rubio, C.; Lindblom, A. Colorectal carcinogenesis is associated with stromal expression of COL11A1 and COL5A2. Carcinogenesis 2001, 22, 875–878. [Google Scholar] [CrossRef]
- Fischer, H.; Salahshor, S.; Stenling, R.; Björk, J.; Lindmark, G.; Iselius, L.; Rubio, C.; Lindblom, A. COL11A1 in FAP polyps and in sporadic colorectal tumors. BMC Cancer 2001, 1, 17. [Google Scholar] [CrossRef] [PubMed]
- Boudjadi, S.; Carrier, J.C.; Beaulieu, J.F. Integrin α1 subunit is up-regulated in colorectal cancer. Biomark. Res. 2013, 1, 16. [Google Scholar] [CrossRef] [PubMed]
- Oh, B.Y.; Kim, K.H.; Chung, S.S.; Hong, K.S.; Lee, R.A. Role of β1-integrin in colorectal cancer: Case-control study. Ann. Coloproctol. 2014, 30, 61–70. [Google Scholar] [CrossRef]
- Zhao, Z.; Han, F.H.; Yang, S.B.; Hua, L.X.; Wu, J.H.; Zhan, W.H. Loss of stromal caveolin-1 expression in colorectal cancer predicts poor survival. World J. Gastroenterol. 2015, 21, 1140–1147. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Q.; Tang, Z.X.; Yu, D.; Cui, S.J.; Jiang, Y.H.; Zhang, Q.; Wang, J.; Yang, P.Y.; Liu, F. Epithelial but not stromal expression of collagen alpha-1(III) is a diagnostic and prognostic indicator of colorectal carcinoma. Oncotarget 2016, 7, 8823–8838. [Google Scholar] [CrossRef]
- Zhao, H.; Yu, H.; Martin, T.A.; Zhang, Y.; Chen, G.; Jiang, W.G. Effect of junctional adhesion molecule-2 expression on cell growth, invasion and migration in human colorectal cancer. Int. J. Oncol. 2016, 48, 929–936. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, D.; Liu, Z.; Li, S.; Ge, Y.; Sun, W.; Liu, B. The inhibitory effects of COL1A2 on colorectal cancer cell proliferation, migration, and invasion. J. Cancer 2018, 9, 2953–2962. [Google Scholar] [CrossRef]
- Galatenko, V.V.; Maltseva, D.V.; Galatenko, A.V.; Rodin, S.; Tonevitsky, A.G. Cumulative prognostic power of laminin genes in colorectal cancer. BMC Med. Genom. 2018, 11 (Suppl. S1), 9. [Google Scholar] [CrossRef]
- Liu, W.; Li, L.; Ye, H.; Tao, H.; He, H. Role of COL6A3 in colorectal cancer. Oncol. Rep. 2018, 39, 2527–2536. [Google Scholar] [CrossRef]
- Lu, L.; Xie, R.; Wei, R.; Cai, C.; Bi, D.; Yin, D.; Liu, H.; Zheng, J.; Zhang, Y.; Song, F.; et al. Integrin α5 subunit is required for the tumor supportive role of fibroblasts in colorectal adenocarcinoma and serves as a potential stroma prognostic marker. Mol. Oncol. 2019, 13, 2697–2714. [Google Scholar] [CrossRef]
- Zhou, D.; Tang, W.; Zhang, Y.; An, H.X. JAM3 functions as a novel tumor suppressor and is inactivated by DNA methylation in colorectal cancer. Cancer Manag. Res. 2019, 11, 2457–2470. [Google Scholar] [CrossRef]
- Zhang, D.; Bi, J.; Liang, Q.; Wang, S.; Zhang, L.; Han, F.; Li, S.; Qiu, B.; Fan, X.; Chen, W.; et al. VCAM1 promotes tumor cell invasion and metastasis by inducing EMT and transendothelial migration in colorectal cancer. Front. Oncol. 2020, 10, 1066. [Google Scholar] [CrossRef]
- Wang, Y.; Sui, J.; Li, X.; Cao, F.; He, J.; Yang, B.; Zhu, X.; Sun, Y.; Pu, Y.D. RPS24 knockdown inhibits colorectal cancer cell migration and proliferation in vitro. Gene 2015, 571, 286–291. [Google Scholar] [CrossRef]
- Yu, C.; Hong, H.; Zhang, S.; Zong, Y.; Ma, J.; Lu, A.; Sun, J.; Zheng, M. Identification of key genes and pathways involved in microsatellite instability in colorectal cancer. Mol. Med. Rep. 2019, 19, 2065–2076. [Google Scholar] [CrossRef]
- Dong, Z.; Jiang, H.; Liang, S.; Wang, Y.; Jiang, W.; Zhu, C. Ribosomal Protein L15 is involved in Colon Carcinogenesis. Int. J. Med. Sci. 2019, 16, 1132–1141. [Google Scholar] [CrossRef]
- Zou, D.; Zhang, H.; Ke, J.; Li, J.; Zhu, Y.; Gong, Y.; Yang, Y.; Tian, J.; Zhang, Y.; Peng, X.; et al. Three functional variants were identified to affect RPS24 expression and significantly associated with risk of colorectal cancer. Arch. Toxicol. 2020, 94, 295–303. [Google Scholar] [CrossRef]
- Wang, Y.; Dong, J.; Quan, Q.; Liu, S.; Chen, X.; Cai, X.; Qiu, H.; Zhang, B.; Guo, G. Immune cell infiltration of the primary tumor microenvironment predicted the treatment outcome of chemotherapy with or without bevacizumab in metastatic colorectal cancer patients. Front. Oncol. 2021, 10, 581051. [Google Scholar] [CrossRef]
- Christoffersson, G.; Vågesjö, E.; Vandooren, J.; Lidén, M.; Massena, S.; Reinert, R.B.; Brissova, M.; Powers, A.C.; Opdenakker, G.; Phillipson, M. VEGF-A recruits a proangiogenic MMP-9-delivering neutrophil subset that induces angiogenesis in transplanted hypoxic tissue. Blood 2012, 120, 4653–4662. [Google Scholar] [CrossRef]
- Winkler, J.; Abisoye-Ogunniyan, A.; Metcalf, K.J.; Werb, Z. Concepts of extracellular matrix remodelling in tumour progression and metastasis. Nat. Commun. 2020, 11, 5120. [Google Scholar] [CrossRef]
- Vinnakota, K.; Zhang, Y.; Selvanesan, B.C.; Topi, G.; Salim, T.; Sand-Dejmek, J.; Jönsson, G.; Sjölander, A. M2-like macrophages induce colon cancer cell invasion via matrix metalloproteinases. J. Cell. Physiol. 2017, 232, 3468–3480. [Google Scholar] [CrossRef]
- Sandel, M.H.; Dadabayev, A.R.; Menon, A.G.; Morreau, H.; Melief, C.J.; Offringa, R.; van der Burg, S.H.; Janssen-van Rhijn, C.M.; Ensink, N.G.; Tollenaar, R.A.; et al. Prognostic value of tumor-infiltrating dendritic cells in colorectal cancer: Role of maturation status and intratumoral localization. Clin. Cancer Res. 2005, 11, 2576–2582. [Google Scholar] [CrossRef] [PubMed]
- Kubo, F.; Ariestanti, D.M.; Oki, S.; Fukuzawa, T.; Demizu, R.; Sato, T.; Sabirin, R.M.; Hirose, S.; Nakamura, N. Loss of the adhesion G-protein coupled receptor ADGRF5 in mice induces airway inflammation and the expression of CCL2 in lung endothelial cells. Respir. Res. 2019, 20, 11. [Google Scholar] [CrossRef] [PubMed]
- Oliveira-Nascimento, L.; Massari, P.; Wetzler, L.M. The role of TLR2 in infection and immunity. Front. Immunol. 2012, 3, 79. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic of Patients with Colorectal Cancer | ADGRF5 Expression | p-Value | Number of Subjects |
|---|---|---|---|
| Sex | |||
| Men | −0.0406 ± 0.779 | ns | 207 |
| Women | 0.0470 ± 1.226 | 169 | |
| Age | |||
| Below the age of 50 | 0.1844 ± 1.140 | ns | 57 |
| Above the age of 50 | −0.0343 ± 0.976 | 319 | |
| Race | |||
| Black or African American | 0.1247 ± 1.804 | ns | 60 |
| White | −0.0109 ± 0.770 | 270 | |
| Site of tumor | |||
| Colon | −0.0428 ± 1.033 | * | 283 |
| Rectum | 0.1219 ± 0.913 | 91 | |
| Histological diagnosis—colon | |||
| Colon adenocarcinoma | −0.1569 ± 0.612 | ** | 242 |
| Colon mucinous adenocarcinoma | 0.7217 ± 2.233 | 38 | |
| Histological diagnosis—rectum | |||
| Rectal adenocarcinoma | 0.0855 ± 0.736 | ns | 86 |
| Rectal mucinous adenocarcinoma | 0.748 ± 2.596 | 5 | |
| Stage | |||
| I/II | −0.0861 ± 1.041 | * | 191 |
| III/IV | 0.0762 ± 0.905 | 166 | |
| TNM—tumor | |||
| 1/2 | −0.2936 ± 0.525 | ** | 67 |
| 3/4 | 0.0644 ± 1.074 | 307 | |
| TNM—node | |||
| 0 | −0.0999 ± 1.011 | * | 205 |
| 1/2 | 0.1131 ± 0.994 | 168 | |
| TNM—metastasis | |||
| 0 | −0.0513 ± 0.788 | ns | 256 |
| 1 | 0.1005 ± 0.753 | 50 | |
| Gene Symbol | Gene ID | Gene Description | Spearman Correlation |
|---|---|---|---|
| AKT3 | 10000 | Serine/threonine kinase 3 | 0.80 |
| ANGPT1 | 284 | Angiopoietin 1 | 0.66 |
| ANGPT2 | 285 | Angiopoietin 2 | 0.74 |
| COL1A2 | 1278 | Collagen type I α2 chain | 0.71 |
| COL4A1 | 1282 | Collagen type IV α1 chain | 0.92 |
| COL4A2 | 1284 | Collagen type IV α2 chain | 0.89 |
| COL6A3 | 1293 | Collagen type VI α3 chain | 0.78 |
| F2R | 2149 | Coagulation factor II thrombin receptor | 0.72 |
| FGF1 | 2246 | Fibroblast growth factor 1 | 0.70 |
| FGF7 | 2252 | Fibroblast growth factor 7 | 0.71 |
| FLT1 | 2321 | FMS-related receptor tyrosine kinase 1 | 0.81 |
| GNB4 | 59345 | G protein subunit β4 | 0.78 |
| GNG2 | 54331 | G protein subunit γ2 | 0.75 |
| GNG11 | 2791 | G protein subunit γ11 | 0.76 |
| HGF | 3082 | Hepatocyte growth factor | 0.72 |
| ITGA1 | 3672 | Integrin subunit α1 | 0.71 |
| ITGA4 | 3676 | Integrin subunit α4 | 0.74 |
| ITGA5 | 3678 | Integrin subunit α5 | 0.79 |
| ITGAV | 3685 | Integrin subunit αV | 0.63 |
| ITGB1 | 3688 | Integrin subunit β1 | 0.61 |
| ITGB3 | 3690 | Integrin subunit β3 | 0.83 |
| KDR | 3791 | Kinase insert domain receptor | 0.88 |
| LAMA4 | 3910 | Laminin subunit α4 | 0.83 |
| LAMC1 | 3915 | Laminin subunit γ1 | 0.74 |
| OSMR | 9180 | Oncostatin M receptor | 0.81 |
| PDGFC | 56034 | Platelet-derived growth factor C | 0.65 |
| PDGFRA | 5156 | Platelet-derived growth factor receptor α | 0.69 |
| PDGFRB | 5159 | Platelet-derived growth factor receptor β | 0.80 |
| SGK1 | 6446 | Serum/glucocorticoid regulated kinase 1 | 0.50 |
| TEK | 7010 | TEK receptor tyrosine kinase | 0.85 |
| THBS2 | 7058 | Thrombospondin 2 | 0.68 |
| TLR2 | 7097 | Toll-like receptor 2 | 0.59 |
| TNC | 3371 | Tenascin C | 0.71 |
| VEGFC | 7424 | Vascular endothelial growth factor C | 0.78 |
| Gene Symbol | Gene ID | Gene Description | Spearman Correlation |
|---|---|---|---|
| AKT3 | 10000 | Serine/threonine kinase 3 | 0.80 |
| CAV1 | 857 | Caveolin 1 | 0.78 |
| CAV2 | 858 | Caveolin 2 | 0.58 |
| COL1A2 | 1278 | Collagen type I α2 chain | 0.71 |
| COL4A1 | 1282 | Collagen type IV α1 chain | 0.92 |
| COL4A2 | 1284 | Collagen type IV α2 chain | 0.89 |
| COL6A3 | 1293 | Collagen type VI α3 chain | 0.78 |
| FLT1 | 2321 | FMS-related receptor tyrosine kinase 1 | 0.81 |
| HGF | 3082 | Hepatocyte growth factor | 0.72 |
| ITGA1 | 3672 | Integrin subunit α1 | 0.71 |
| ITGA4 | 3676 | Integrin subunit α4 | 0.74 |
| ITGA5 | 3678 | Integrin subunit α5 | 0.79 |
| ITGAV | 3685 | Integrin subunit αV | 0.63 |
| ITGB1 | 3688 | Integrin subunit β1 | 0.61 |
| ITGB3 | 3690 | Integrin subunit β3 | 0.83 |
| KDR | 3791 | Kinase insert domain receptor | 0.88 |
| LAMA4 | 3910 | Laminin subunit α4 | 0.83 |
| LAMC1 | 3915 | Laminin subunit γ1 | 0.74 |
| MYLK | 4638 | Myosin light chain kinase | 0.68 |
| PDGFC | 56034 | Platelet-derived growth factor C | 0.65 |
| PDGFRA | 5156 | Platelet-derived growth factor receptor α | 0.69 |
| PDGFRB | 5159 | Platelet-derived growth factor receptor β | 0.80 |
| THBS2 | 7058 | Thrombospondin 2 | 0.68 |
| TNC | 3371 | Tenascin C | 0.71 |
| VEGFC | 7424 | Vascular endothelial growth factor C | 0.78 |
| Gene Symbol | Gene ID | Gene Description | Spearman Correlation |
|---|---|---|---|
| CD34 | 947 | CD34 molecule | 0.90 |
| CD86 | 942 | CD86 molecule | 0.65 |
| CDH2 | 1000 | Cadherin 2 | 0.63 |
| CDH5 | 1003 | Cadherin 5 | 0.92 |
| ESAM | 90952 | Endothelial cell adhesion molecule | 0.81 |
| ITGA4 | 3676 | Integrin subunit α4 | 0.74 |
| ITGAV | 3685 | Integrin subunit αV | 0.63 |
| ITGB1 | 3688 | Integrin subunit β1 | 0.61 |
| JAM2 | 58494 | Junctional adhesion molecule 2 | 0.74 |
| JAM3 | 83700 | Junctional adhesion molecule 3 | 0.79 |
| NCAM2 | 4685 | Neural cell adhesion molecule 2 | 0.57 |
| NLGN4X | 57502 | Neuroligin 4 X-linked | 0.70 |
| PDCD1LG2 | 80380 | Programmed cell death 1 ligand 2 | 0.67 |
| PECAM1 | 5175 | Platelet and endothelial cell adhesion molecule 1 | 0.83 |
| PTPRC | 5788 | Protein tyrosine phosphatase receptor type C | 0.59 |
| PTPRM | 5797 | Protein tyrosine phosphatase receptor type M | 0.83 |
| SDC2 | 6383 | Syndecan 2 | 0.72 |
| SELE | 6401 | Selectin E | 0.70 |
| SELL | 6402 | Selectin L | 0.59 |
| VCAM1 | 7412 | Vascular cell adhesion molecule 1 | 0.74 |
| VCAN | 1462 | Versican | 0.77 |
| Gene Symbol | Gene ID | Gene Description | Spearman Correlation |
|---|---|---|---|
| FAU | 2197 | FAU ubiquitin-like and ribosomal protein S30 fusion | −0.36 |
| MRPL1 | 65008 | Mitochondrial ribosomal protein L1 | −0.37 |
| MRPL2 | 51069 | Mitochondrial ribosomal protein L2 | −0.30 |
| MRPL3 | 11222 | Mitochondrial ribosomal protein L3 | −0.30 |
| MRPL4 | 51073 | Mitochondrial ribosomal protein L4 | −0.37 |
| MRPL11 | 65003 | Mitochondrial ribosomal protein L11 | −0.39 |
| MRPL12 | 6182 | Mitochondrial ribosomal protein L12 | −0.44 |
| MRPL14 | 64928 | Mitochondrial ribosomal protein L14 | −0.38 |
| MRPL15 | 29088 | Mitochondrial ribosomal protein L15 | −0.29 |
| MRPL16 | 54948 | Mitochondrial ribosomal protein L16 | −0.35 |
| MRPL17 | 63875 | Mitochondrial ribosomal protein L17 | −0.37 |
| MRPL20 | 55052 | Mitochondrial ribosomal protein L20 | −0.36 |
| MRPL21 | 219927 | Mitochondrial ribosomal protein L21 | −0.38 |
| MRPL22 | 29093 | Mitochondrial ribosomal protein L22 | −0.39 |
| MRPL23 | 6150 | Mitochondrial ribosomal protein L23 | −0.38 |
| MRPL24 | 29093 | Mitochondrial ribosomal protein L24 | −0.41 |
| MRPL27 | 51264 | Mitochondrial ribosomal protein L27 | −0.47 |
| MRPL32 | 64983 | Mitochondrial ribosomal protein L32 | −0.34 |
| MRPL34 | 64981 | Mitochondrial ribosomal protein L34 | −0.36 |
| MRPL36 | 64979 | Mitochondrial ribosomal protein L36 | −0.38 |
| MRPS2 | 51116 | Mitochondrial ribosomal protein S2 | −0.37 |
| MRPS5 | 64969 | Mitochondrial ribosomal protein S5 | −0.33 |
| MRPS9 | 64965 | Mitochondrial ribosomal protein S9 | −0.35 |
| MRPS12 | 6183 | Mitochondrial ribosomal protein S12 | −0.41 |
| MRPS15 | 64960 | Mitochondrial ribosomal protein S15 | −0.37 |
| MRPS16 | 51021 | Mitochondrial ribosomal protein S16 | −0.40 |
| MRPS17 | 51373 | Mitochondrial ribosomal protein S17 | −0.36 |
| MRPS18A | 55168 | Mitochondrial ribosomal protein S18A | −0.29 |
| RPL3 | 6122 | Ribosomal protein L3 | −0.29 |
| RPL5 | 6125 | Ribosomal protein L5 | −0.31 |
| RPL6 | 6128 | Ribosomal protein L6 | −0.33 |
| RPL7A | 6130 | Ribosomal protein L7a | −0.35 |
| RPL8 | 6132 | Ribosomal protein L8 | −0.30 |
| RPL11 | 6135 | Ribosomal protein L11 | −0.33 |
| RPL12 | 6136 | Ribosomal protein L12 | −0.37 |
| RPL13 | 6137 | Ribosomal protein L13 | −0.40 |
| RPL14 | 9045 | Ribosomal protein L14 | −0.41 |
| RPL15 | 6138 | Ribosomal protein L15 | −0.34 |
| RPL18 | 6141 | Ribosomal protein L18 | −0.42 |
| RPL18A | 6142 | Ribosomal protein L18a | −0.29 |
| RPL19 | 6143 | Ribosomal protein L19 | −0.38 |
| RPL23 | 9349 | Ribosomal protein L23 | −0.31 |
| RPL23A | 6147 | Ribosomal protein L23a | −0.33 |
| RPL24 | 6152 | Ribosomal protein L24 | −0.43 |
| RPL27 | 6155 | Ribosomal protein L27 | −0.37 |
| RPL27A | 6157 | Ribosomal protein L27A | −0.37 |
| RPL28 | 6158 | Ribosomal protein L28 | −0.35 |
| RPL29 | 6159 | Ribosomal protein L29 | −0.42 |
| RPL31 | 6160 | Ribosomal protein L31 | −0.33 |
| RPL32 | 6161 | Ribosomal protein L32 | −0.42 |
| RPL34 | 6164 | Ribosomal protein L34 | −0.34 |
| RPL35 | 11224 | Ribosomal protein L35 | −0.39 |
| RPL35A | 6165 | Ribosomal protein L35A | −0.38 |
| RPL36 | 25873 | Ribosomal protein L36 | −0.43 |
| RPL37 | 6167 | Ribosomal protein L37 | −0.31 |
| RPL37A | 6173 | Ribosomal protein L37a | −0.37 |
| RPL38 | 6169 | Ribosomal protein L38 | −0.39 |
| RPL39 | 6170 | Ribosomal protein L39 | −0.36 |
| RPLP0 | 6175 | Ribosomal protein lateral stalk subunit P0 | −0.39 |
| RPLP1 | 6176 | Ribosomal protein lateral stalk subunit P1 | −0.36 |
| RPLP2 | 6181 | Ribosomal protein lateral stalk subunit P2 | −0.44 |
| RPS2 | 6187 | Ribosomal protein S2 | −0.42 |
| RPS3 | 6188 | Ribosomal protein S3 | −0.34 |
| RPS4X | 6191 | Ribosomal protein S4X | −0.31 |
| RPS6 | 6194 | Ribosomal protein S6 | −0.29 |
| RPS7 | 6201 | Ribosomal protein S7 | −0.35 |
| RPS8 | 6202 | Ribosomal protein S8 | −0.38 |
| RPS9 | 6203 | Ribosomal protein S9 | −0.35 |
| RPS10 | 6204 | Ribosomal protein S10 | −0.34 |
| RPS11 | 6205 | Ribosomal protein S11 | −0.35 |
| RPS13 | 6207 | Ribosomal protein S13 | −0.38 |
| RPS14 | 6208 | Ribosomal protein S14 | −0.40 |
| RPS15 | 6209 | Ribosomal protein S15 | −0.39 |
| RPS15A | 6210 | Ribosomal protein S15A | −0.31 |
| RPS16 | 6217 | Ribosomal protein S16 | −0.39 |
| RPS17 | 6218 | Ribosomal protein S17 | −0.30 |
| RPS18 | 6222 | Ribosomal protein S18 | −0.34 |
| RPS19 | 6223 | Ribosomal protein S19 | −0.34 |
| RPS21 | 6227 | Ribosomal protein S21 | −0.29 |
| RPS23 | 6228 | Ribosomal protein S23 | −0.30 |
| RPS24 | 6229 | Ribosomal protein S24 | −0.31 |
| RPS25 | 6230 | Ribosomal protein S25 | −0.33 |
| RPS27A | 6233 | Ribosomal protein S27A | −0.33 |
| RPS29 | 6235 | Ribosomal protein S29 | −0.31 |
| RPSA | 3921 | Ribosomal protein SA | −0.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, H.; Fichna, J.; Matlawska-Wasowska, K.; Jacenik, D. The Expression Pattern of Adhesion G Protein-Coupled Receptor F5 Is Related to Cell Adhesion and Metastatic Pathways in Colorectal Cancer—Comprehensive Study Based on In Silico Analysis. Cells 2022, 11, 3876. https://doi.org/10.3390/cells11233876
Kang H, Fichna J, Matlawska-Wasowska K, Jacenik D. The Expression Pattern of Adhesion G Protein-Coupled Receptor F5 Is Related to Cell Adhesion and Metastatic Pathways in Colorectal Cancer—Comprehensive Study Based on In Silico Analysis. Cells. 2022; 11(23):3876. https://doi.org/10.3390/cells11233876
Chicago/Turabian StyleKang, Huining, Jakub Fichna, Ksenia Matlawska-Wasowska, and Damian Jacenik. 2022. "The Expression Pattern of Adhesion G Protein-Coupled Receptor F5 Is Related to Cell Adhesion and Metastatic Pathways in Colorectal Cancer—Comprehensive Study Based on In Silico Analysis" Cells 11, no. 23: 3876. https://doi.org/10.3390/cells11233876
APA StyleKang, H., Fichna, J., Matlawska-Wasowska, K., & Jacenik, D. (2022). The Expression Pattern of Adhesion G Protein-Coupled Receptor F5 Is Related to Cell Adhesion and Metastatic Pathways in Colorectal Cancer—Comprehensive Study Based on In Silico Analysis. Cells, 11(23), 3876. https://doi.org/10.3390/cells11233876