

Type XXII Collagen Complements Fibrillar Collagens in the Serological Assessment of Tumor Fibrosis and the Outcome in Pancreatic Cancer

 , , , , , , ,
, , , , , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Generation of Monoclonal Antibodies Targeting the C-Terminal of Type XXII Collagen
2.2. PRO-C22 ELISA Protocol
2.3. Technical Validation of the PRO-C22 ELISA
2.4. Type I and III Collagen Assessment
2.5. Patient Samples
2.6. Data and Statistical Analysis
3. Results
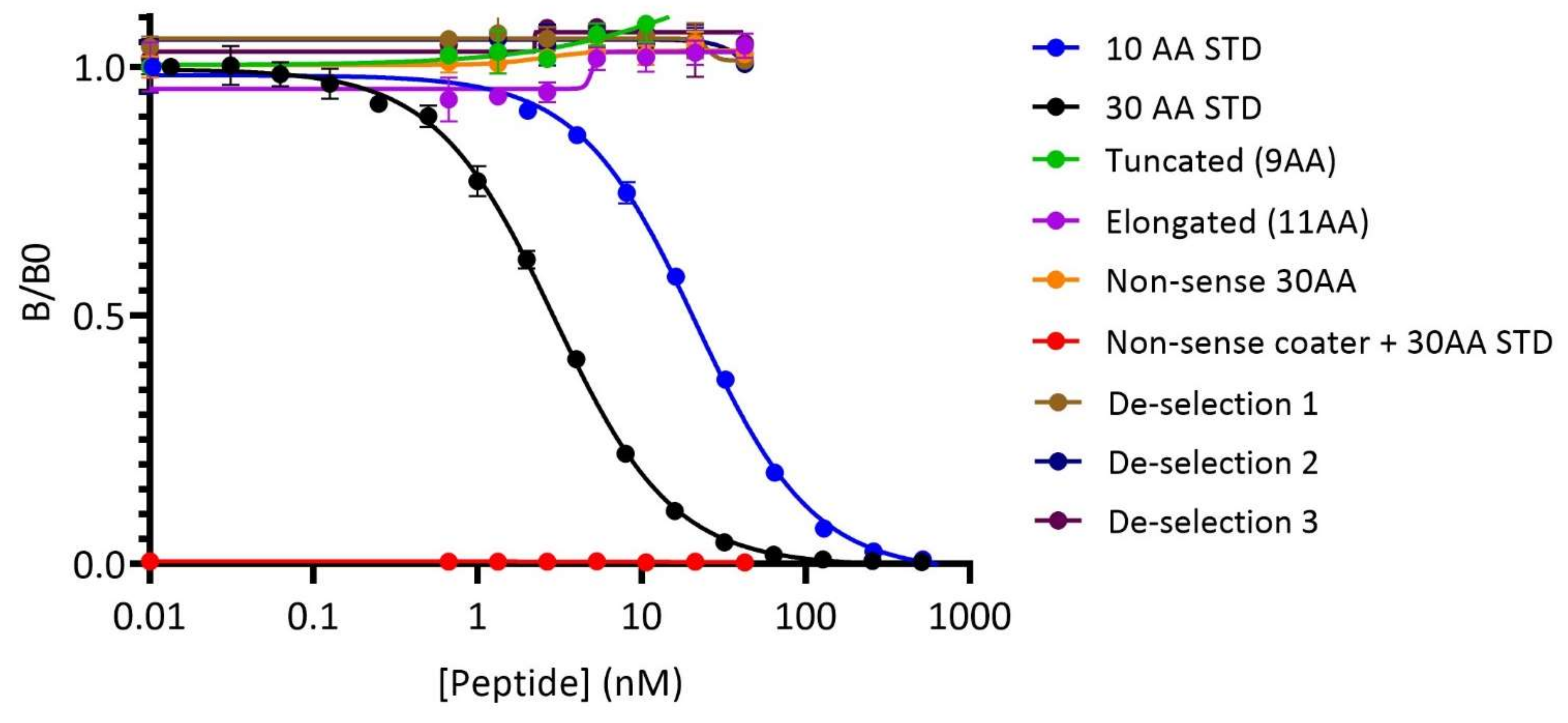
3.1. The PRO-C22 ELISA Assay Was Validated as a Specific and Technically Robust Biomarker
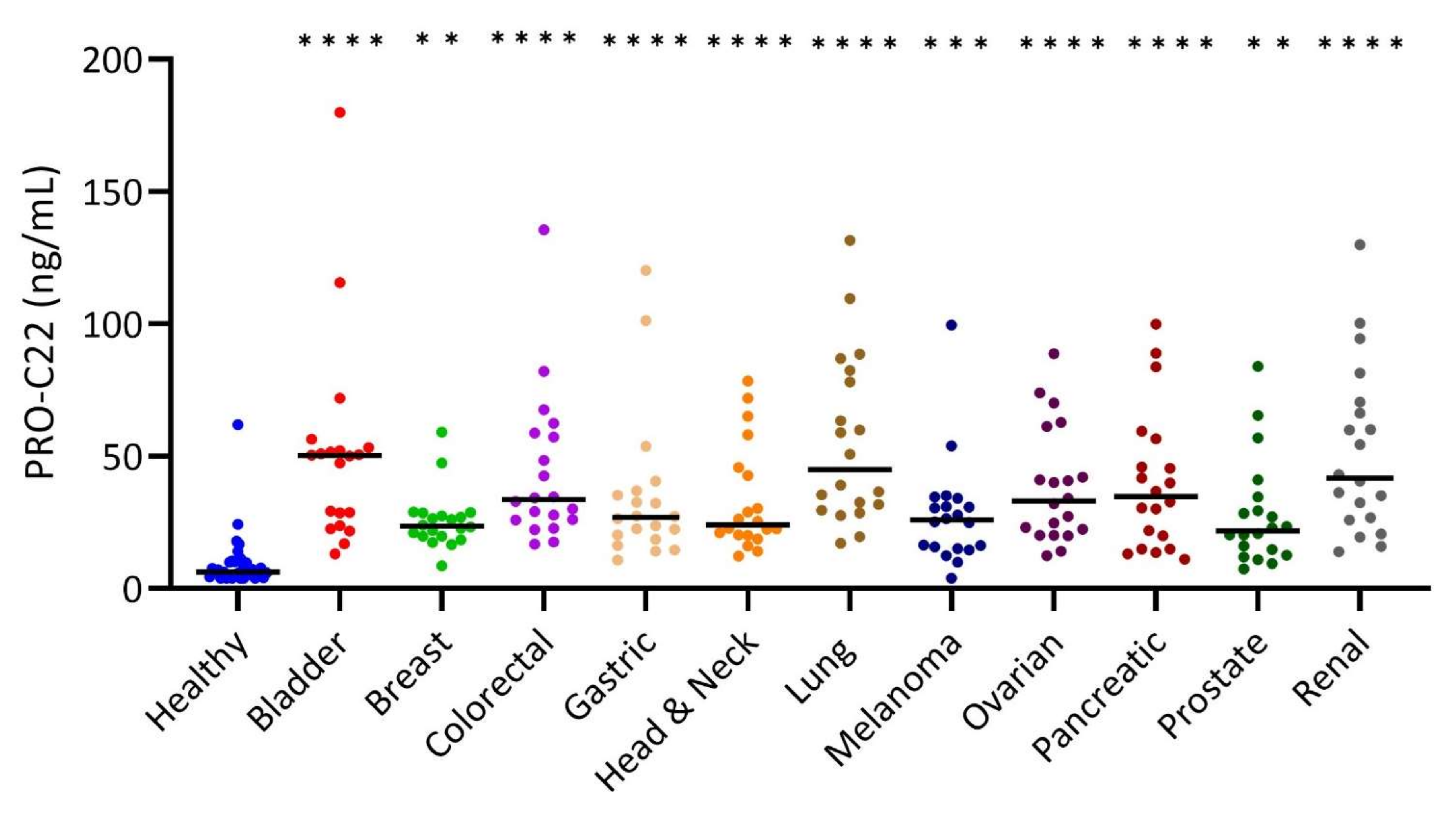
3.2. PRO-C22 Was Significantly Elevated in the Serum from Patients with Solid Cancers
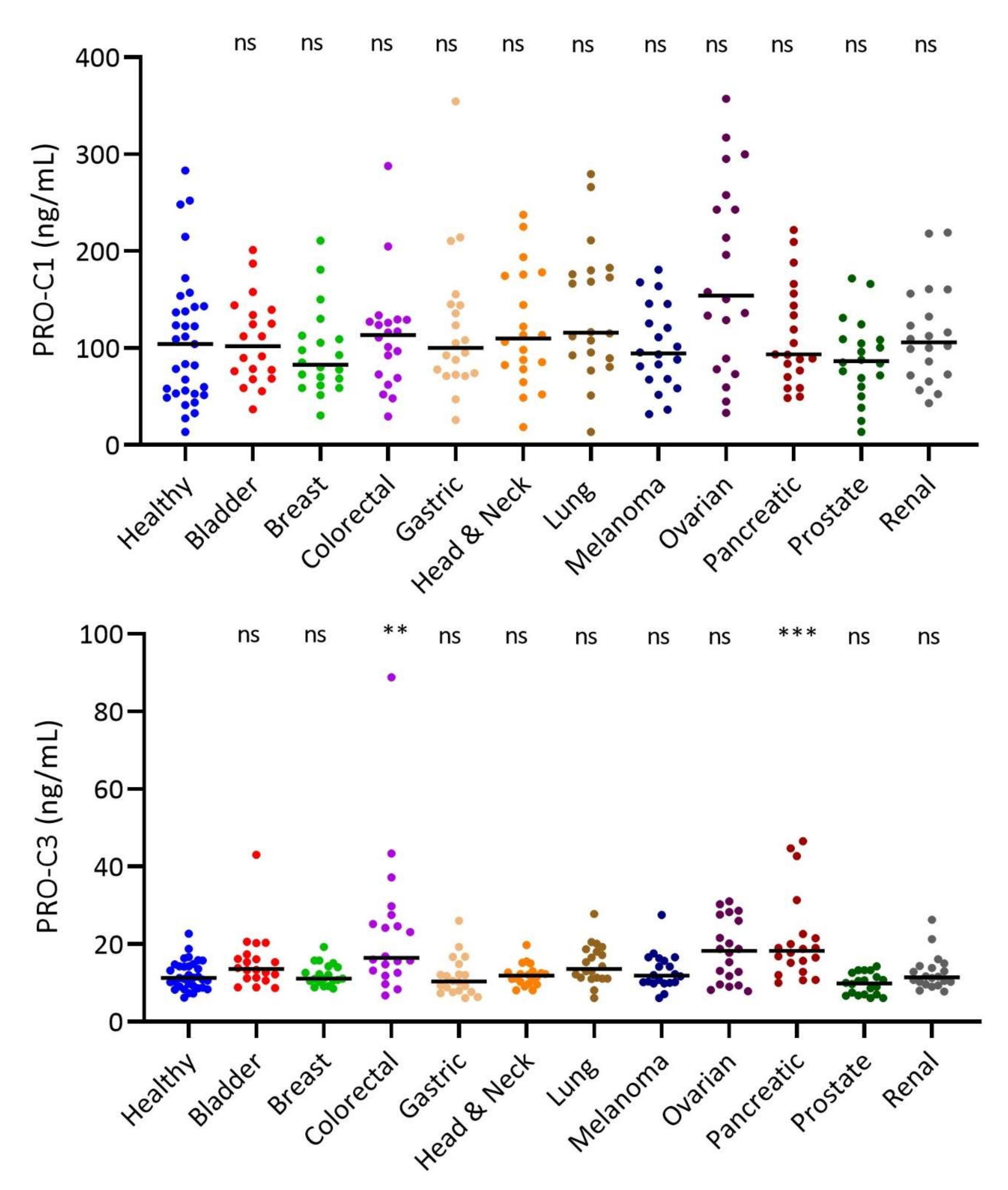
3.3. Interstitial Matrix Biomarkers PRO-C1 and PRO-C3 in Solid Tumor Types
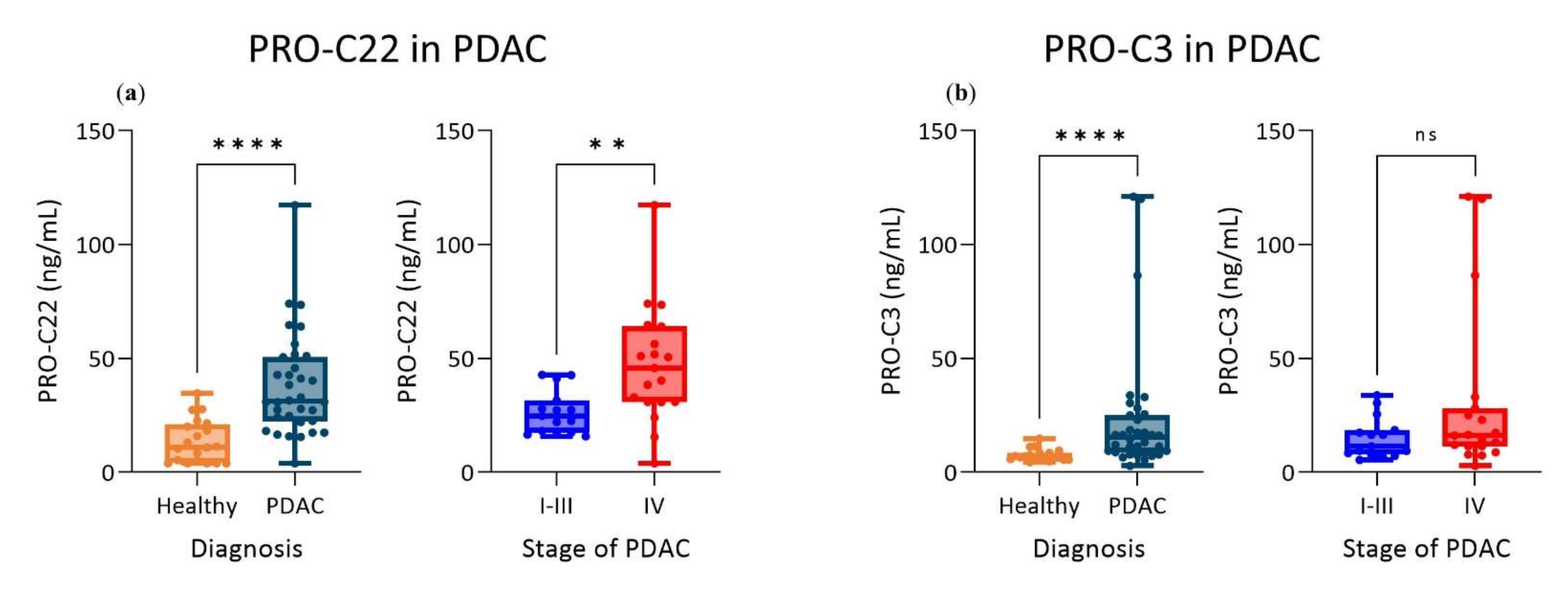
3.4. PRO-C22 and PRO-C3 Were Significantly Elevated in Patients with PDAC (Cohort 2)
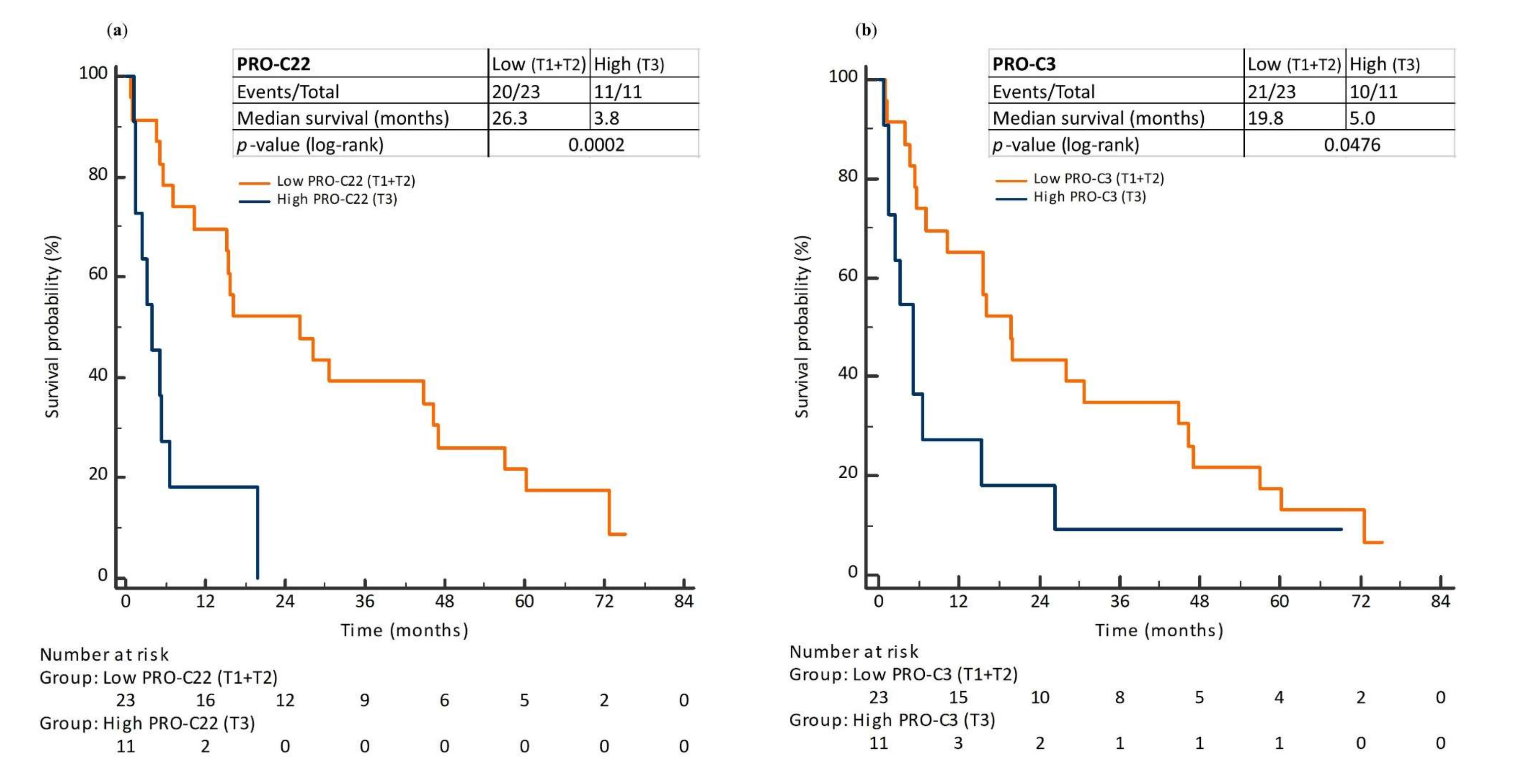
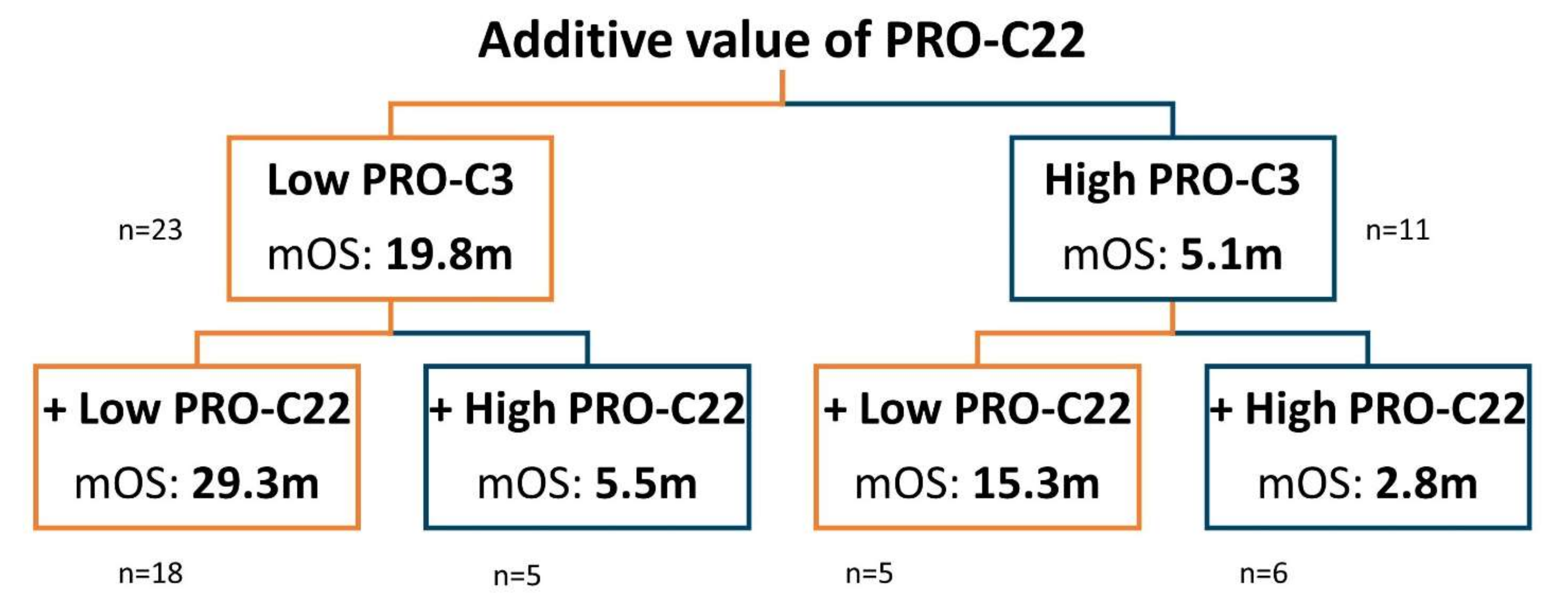
3.5. Prognostic Perspective of PRO-C22 and PRO-C3 in Patients with PDAC (Cohort 2)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- WHO. WHO|Cancer Factsheet. Available online: http://www.who.int/mediacentre/factsheets/fs297/en/ (accessed on 23 July 2021).
- World Health Organization. WHO Report on Cancer: Setting Priorities, Investing Wisely and Providing Care for All; World Health Organization: Geneva, Switzerland, 2020.
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Rahib, L.; Wehner, M.R.; Matrisian, L.M.; Nead, K.T. Estimated Projection of US Cancer Incidence and Death to 2040. JAMA Netw. Open 2021, 4, e214708. [Google Scholar] [CrossRef]
- Murakami, T.; Hiroshima, Y.; Matsuyama, R.; Homma, Y.; Hoffman, R.M.; Endo, I. Role of the Tumor Microenvironment in Pancreatic Cancer. Ann. Gastroenterol. Surg. 2019, 3, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Pickup, M.W.; Mouw, J.K.; Weaver, V.M. The Extracellular Matrix Modulates the Hallmarks of Cancer. EMBO Rep. 2014, 15, 1243–1253. [Google Scholar] [CrossRef]
- Gialeli, C.; Theocharis, A.D.; Karamanos, N.K. Roles of Matrix Metalloproteinases in Cancer Progression and Their Pharmacological Targeting. FEBS J. 2011, 278, 16–27. [Google Scholar] [CrossRef]
- Cox, T.R. The Matrix in Cancer. Nat. Rev. Cancer 2021, 21, 217–238. [Google Scholar] [CrossRef]
- Vasan, N.; Baselga, J.; Hyman, D.M. A View on Drug Resistance in Cancer. Nature 2019, 575, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Jensen, C.; Sinkeviciute, D.; Madsen, D.H.; Önnerfjord, P.; Hansen, M.; Schmidt, H.; Karsdal, M.A.; Svane, I.M.; Willumsen, N. Granzyme B Degraded Type IV Collagen Products in Serum Identify Melanoma Patients Responding to Immune Checkpoint Blockade. Cancers 2020, 12, 2786. [Google Scholar] [CrossRef]
- Balkwill, F.R.; Capasso, M.; Hagemann, T. The Tumor Microenvironment at a Glance. J. Cell Sci. 2012, 125, 5591–5596. [Google Scholar] [CrossRef]
- Cox, T.R.; Erler, J.T. Remodeling and Homeostasis of the Extracellular Matrix: Implications for Fibrotic Diseases and Cancer. Dis. Model. Mech. 2011, 4, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Cox, T.R.; Erler, J.T. Fibrosis and Cancer: Partners in Crime or Opposing Forces? Trends Cancer 2016, 2, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Willumsen, N.; Jensen, C.; Green, G.; Nissen, N.I.; Neely, J.; Nelson, D.M.; Pedersen, R.S.; Frederiksen, P.; Chen, I.M.; Boisen, M.K.; et al. Fibrotic Activity Quantified in Serum by Measurements of Type III Collagen Pro-Peptides Can Be Used for Prognosis across Different Solid Tumor Types. Cell. Mol. Life Sci. 2022, 79, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jensen, C.; Madsen, D.H.; Hansen, M.; Schmidt, H.; Svane, I.M.; Karsdal, M.A.; Willumsen, N. Non-Invasive Biomarkers Derived from the Extracellular Matrix Associate with Response to Immune Checkpoint Blockade (Anti-CTLA-4) in Metastatic Melanoma Patients. J. Immunother. Cancer 2018, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hurkmans, D.P.; Jensen, C.; Koolen, S.L.W.; Aerts, J.; Karsdal, M.A.; Mathijssen, R.H.J.; Willumsen, N. Blood-Based Extracellular Matrix Biomarkers Are Correlated with Clinical Outcome after PD-1 Inhibition in Patients with Metastatic Melanoma. J. Immunother. Cancer 2020, 8, e001193. [Google Scholar] [CrossRef] [PubMed]
- Nissen, N.I.; Kehlet, S.; Boisen, M.K.; Liljefors, M.; Jensen, C.; Johansen, A.Z.; Johansen, J.S.; Erler, J.T.; Karsdal, M.; Mortensen, J.H.; et al. Prognostic Value of Blood-Based Fibrosis Biomarkers in Patients with Metastatic Colorectal Cancer Receiving Chemotherapy and Bevacizumab. Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Willumsen, N.; Ali, S.M.; Leitzel, K.; Drabick, J.J.; Yee, N.; Polimera, H.V.; Nagabhairu, V.; Krecko, L.; Ali, A.; Maddukuri, A.; et al. Collagen Fragments Quantified in Serum as Measures of Desmoplasia Associate with Survival Outcome in Patients with Advanced Pancreatic Cancer. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef]
- Lipton, A.; Leitzel, K.; Ali, S.M.; Polimera, H.V.; Nagabhairu, V.; Marks, E.; Richardson, A.E.; Krecko, L.; Ali, A.; Koestler, W.; et al. High Turnover of Extracellular Matrix Reflected by Specific Protein Fragments Measured in Serum Is Associated with Poor Outcomes in Two Metastatic Breast Cancer Cohorts. Int. J. Cancer 2018, 143, 3027–3034. [Google Scholar] [CrossRef]
- Hanash, S.M.; Baik, C.S.; Kallioniemi, O. Emerging Molecular Biomarkers-Blood-Based Strategies to Detect and Monitor Cancer. Nat. Rev. Clin. Oncol. 2011, 8, 142–150. [Google Scholar] [CrossRef]
- Sund, M.; Kalluri, R. Tumor Stroma Derived Biomarkers in Cancer. Cancer Metastasis Rev. 2009, 28, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Mayeux, R. Biomarkers: Potential Uses and Limitations. NeuroRX 2004, 1, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Karsdal, M.A. Biochemistry of Collagens, Laminins and Elastin: Structure, Function and Biomarkers; Elsevier: Amsterdam, The Netherlands, 2019; ISBN 9780128170687. [Google Scholar]
- Sekiguchi, R.; Yamada, K.M. Basement Membranes in Development and Disease. In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 130, pp. 143–191. ISBN 9780128098028. [Google Scholar]
- Choi, Y.S.; Holle, A.W.; Engler, A.J. Extracellular Matrix in Development; DeSimone, D.W., Mecham, R.P., Eds.; Biology of Extracellular Matrix; Springer: Berlin/Heidelberg, Germany, 2013; ISBN 978-3-642-35934-7. [Google Scholar]
- Karsdal, M.A.; Detlefsen, S.; Daniels, S.J.; Nielsen, M.J.; Krag, A.; Schuppan, D. Is the Total Amount as Important as Localization and Type of Collagen in Liver Fibrosis Attributable to Steatohepatitis? Hepatology 2020, 71, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Olsen, B.R.; Winterhalter, K.H.; Gordon, M.K. FACIT Collagens and Their Biological Roles. Trends Glycosci. Glycotechnol. 1995, 7, 115–127. [Google Scholar] [CrossRef]
- Bourgot, I.; Primac, I.; Louis, T.; Noël, A.; Maquoi, E. Reciprocal Interplay between Fibrillar Collagens and Collagen-Binding Integrins: Implications in Cancer Progression and Metastasis. Front. Oncol. 2020, 10, 1488. [Google Scholar] [CrossRef]
- Sumiyoshi, H.; Inoguchi, K.; Khaleduzzaman, M.; Ninomiya, Y.; Yoshioka, H. Ubiquitous Expression of the A1(XIX) Collagen Gene (Col19a1) during Mouse Embryogenesis Becomes Restricted to a Few Tissues in the Adult Organism. J. Biol. Chem. 1997, 272, 17104–17111. [Google Scholar] [CrossRef]
- Koch, M.; Foley, J.E.; Hahn, R.; Zhou, P.; Burgeson, R.E.; Gerecke, D.R.; Gordon, M.K. A1(XX) Collagen, a New Member of the Collagen Subfamily, Fibril-Associated Collagens with Interrupted Triple Helices. J. Biol. Chem. 2001, 276, 23120–23126. [Google Scholar] [CrossRef]
- Koch, M.; Schulze, J.; Hansen, U.; Ashwodt, T.; Keene, D.R.; Brunken, W.J.; Burgeson, R.E.; Bruckner, P.; Bruckner-Tuderman, L. A Novel Marker of Tissue Junctions, Collagen XXII. J. Biol. Chem. 2004, 279, 22514–22521. [Google Scholar] [CrossRef] [PubMed]
- Charvet, B.; Guiraud, A.; Malbouyres, M.; Zwolanek, D.; Guillon, E.; Bretaud, S.; Monnot, C.; Schulze, J.; Bader, H.L.; Allard, B.; et al. Knockdown of Col22a1 Gene in Zebrafish Induces a Muscular Dystrophy by Disruption of the Myotendinous Junction. Development 2013, 140, 4602–4613. [Google Scholar] [CrossRef] [PubMed]
- Calvo, A.; Moreno, L.; Moreno, L.; Toivonen, J.; Manzano, R.; Molina, N.; de la Torre, M.; López, T.; Miana-Mena, F.; Muñoz, M.; et al. Type XIX Collagen: A Promising Biomarker from the Basement Membranes. Neural Regen. Res. 2020, 15, 988. [Google Scholar] [CrossRef]
- Vaughan, L.; Mendler, M.; Huber, S.; Bruckner, P.; Winterhalter, K.H.; Irwin, M.I.; Mayne, R. D-Periodic Distribution of Collagen Type IX along Cartilage Fibrils. J. Cell Biol. 1988, 106, 991–997. [Google Scholar] [CrossRef]
- Blumbach, K.; Bastiaansen-Jenniskens, Y.M.; DeGroot, J.; Paulsson, M.; van Osch, G.J.V.M.; Zaucke, F. Combined Role of Type IX Collagen and Cartilage Oligomeric Matrix Protein in Cartilage Matrix Assembly: Cartilage Oligomeric Matrix Protein Counteracts Type IX Collagen–Induced Limitation of Cartilage Collagen Fibril Growth in Mouse Chondrocyt. Arthritis Rheum. 2009, 60, 3676–3685. [Google Scholar] [CrossRef]
- Karagiannis, G.S.; Petraki, C.; Prassas, I.; Saraon, P.; Musrap, N.; Dimitromanolakis, A.; Diamandis, E.P. Proteomic Signatures of the Desmoplastic Invasion Front Reveal Collagen Type XII as a Marker of Myofibroblastic Differentiation during Colorectal Cancer Metastasis. Oncotarget 2012, 3, 267–285. [Google Scholar] [CrossRef]
- Papanicolaou, M.; Parker, A.L.; Yam, M.; Filipe, E.C.; Wu, S.Z.; Chitty, J.L.; Wyllie, K.; Tran, E.; Mok, E.; Nadalini, A.; et al. Temporal Profiling of the Breast Tumour Microenvironment Reveals Collagen XII as a Driver of Metastasis. Nat. Commun. 2022, 13, 1–21. [Google Scholar] [CrossRef]
- Jensen, C.; Nielsen, S.H.; Mortensen, J.H.; Kjeldsen, J.; Klinge, L.G.; Krag, A.; Harling, H.; Jørgensen, L.N.; Karsdal, M.A.; Willumsen, N. Serum Type XVI Collagen Is Associated with Colorectal Cancer and Ulcerative Colitis Indicating a Pathological Role in Gastrointestinal Disorders. Cancer Med. 2018, 7, 4619–4626. [Google Scholar] [CrossRef] [PubMed]
- Thorlacius-Ussing, J.; Manon-Jensen, T.; Sun, S.; Leeming, D.J.; Sand, J.M.; Karsdal, M.; Willumsen, N. Serum Type XIX Collagen Is Significantly Elevated in Non-Small Cell Lung Cancer: A Preliminary Study on Biomarker Potential. Cancers 2020, 12, 1510. [Google Scholar] [CrossRef] [PubMed]
- Thorlacius-Ussing, J.; Jensen, C.; Madsen, E.A.; Nissen, N.I.; Manon-Jensen, T.; Chen, I.M.; Johansen, J.S.; Diab, H.M.H.; Jørgensen, L.N.; Karsdal, M.A.; et al. Type XX Collagen Is Elevated in Circulation of Patients with Solid Tumors. Int. J. Mol. Sci. 2022, 23, 4144. [Google Scholar] [CrossRef]
- Zhao, W.; Wiedemann, P.; Wölfel, E.M.; Neven, M.; Peters, S.; Imhof, T.; Koch, M.; Busse, B.; Amling, M.; Schinke, T.; et al. Decreased Trabecular Bone Mass in Col22a1-Deficient Mice. Cells 2021, 10, 3020. [Google Scholar] [CrossRef] [PubMed]
- Balemans, M.C.M.; Ansar, M.; Oudakker, A.R.; van Caam, A.P.M.; Bakker, B.; Vitters, E.L.; van der Kraan, P.M.; de Bruijn, D.R.H.; Janssen, S.M.; Kuipers, A.J.; et al. Reduced Euchromatin Histone Methyltransferase 1 Causes Developmental Delay, Hypotonia, and Cranial Abnormalities Associated with Increased Bone Gene Expression in Kleefstra Syndrome Mice. Dev. Biol. 2014, 386, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Chan, W.C.W.; Lam, Y.; Wang, X.; Chen, P.; Niu, B.; Ng, V.C.W.; Yeo, J.C.; Stricker, S.; Cheah, K.S.E.; et al. Lgr5 and Col22a1 Mark Progenitor Cells in the Lineage toward Juvenile Articular Chondrocytes. Stem Cell Rep. 2019, 13, 713–729. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Berndt, A.; Sundberg, B.A.; Silva, K.A.; Kennedy, V.E.; Cario, C.L.; Richardson, M.A.; Chase, T.H.; Schofield, P.N.; Uitto, J.; et al. Mouse Genome-Wide Association Study Identifies Polymorphisms on Chromosomes 4, 11, and 15 for Age-Related Cardiac Fibrosis. Mamm. Genome 2016, 27, 179–190. [Google Scholar] [CrossRef]
- Misawa, K.; Kanazawa, T.; Imai, A.; Endo, S.; Mochizuki, D.; Fukushima, H.; Misawa, Y.; Mineta, H. Prognostic Value of Type XXII and XXIV Collagen MRNA Expression in Head and Neck Cancer Patients. Mol. Clin. Oncol. 2014, 2, 285–291. [Google Scholar] [CrossRef]
- Gefter, M.L.; Margulies, D.H.; Scharff, M.D. A Simple Method for Polyethylene Glycol-Promoted Hybridization of Mouse Myeloma Cells. Somat. Cell Genet. 1977, 3, 231–236. [Google Scholar] [CrossRef]
- Leeming, D.J.; Larsen, D.V.; Zhang, C.; Hi, Y.; Veidal, S.S.; Nielsen, R.H.; Henriksen, K.; Zheng, Q.; Barkholt, V.; Riis, B.J.; et al. Enzyme-Linked Immunosorbent Serum Assays (ELISAs) for Rat and Human N-Terminal pro-Peptide of Collagen Type I (PINP)—Assessment of Corresponding Epitopes. Clin. Biochem. 2010, 43, 1249–1256. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.J.; Nedergaard, A.F.; Sun, S.; Veidal, S.S.; Larsen, L.; Zheng, Q.; Suetta, C.; Henriksen, K.; Christiansen, C.; Karsdal, M.A.; et al. The Neo-Epitope Specific PRO-C3 ELISA Measures True Formation of Type III Collagen Associated with Liver and Muscle Parameters. Am. J. Transl. Res. 2013, 5, 303–315. [Google Scholar] [PubMed]
- Chen, I.; Vittrup Jensen, B.; Bojesen, S.E.; Johansen, A.Z.; Schultz, N.A.; Hansen, C.P.; Hasselby, J.P.; Holländer, N.H.; Nissen, M.H.B.; Bjerregaard, J.K.; et al. Identification of New Biomarkers in Patients with Pancreatic Cancer (BIOPAC): A Study Protocol of an Open Cohort Study. J. Cancer Sci. Ther. 2019, 11, 232–239. [Google Scholar]
- Mukaka, M.M. Statistics Corner: A Guide to Appropriate Use of Correlation Coefficient in Medical Research. Malawi Med. J. 2012, 24, 69–71. [Google Scholar]
- Willumsen, N.; Thomsen, L.B.; Bager, C.L.; Jensen, C.; Karsdal, M.A. Quantification of Altered Tissue Turnover in a Liquid Biopsy: A Proposed Precision Medicine Tool to Assess Chronic Inflammation and Desmoplasia Associated with a pro-Cancerous Niche and Response to Immuno-Therapeutic Anti-Tumor Modalities. Cancer Immunol. Immunother. 2018, 67, 1–12. [Google Scholar] [CrossRef]
- Zhou, Z.-H.; Ji, C.-D.; Xiao, H.-L.; Zhao, H.-B.; Cui, Y.-H.; Bian, X.-W. Reorganized Collagen in the Tumor Microenvironment of Gastric Cancer and Its Association with Prognosis. J. Cancer 2017, 8, 1466–1476. [Google Scholar] [CrossRef]
- Rittié, L. Method for picrosirius red-polarization detection of collagen fibers in tissue sections. In Fibrosis; Humana Press: New York, NY, USA, 2017; pp. 395–407. [Google Scholar]
- Lattouf, R.; Younes, R.; Lutomski, D.; Naaman, N.; Godeau, G.; Senni, K.; Changotade, S. Picrosirius Red Staining. J. Histochem. Cytochem. 2014, 62, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Di Lullo, G.A.; Sweeney, S.M.; Körkkö, J.; Ala-Kokko, L.; San Antonio, J.D. Mapping the Ligand-Binding Sites and Disease-Associated Mutations on the Most Abundant Protein in the Human, Type I Collagen. J. Biol. Chem. 2002, 277, 4223–4231. [Google Scholar] [CrossRef] [PubMed]
- Ueno, H.; Kajiwara, Y.; Ajioka, Y.; Sugai, T.; Sekine, S.; Ishiguro, M.; Takashima, A.; Kanemitsu, Y. Histopathological Atlas of Desmoplastic Reaction Characterization in Colorectal Cancer. Jpn. J. Clin. Oncol. 2021, 51, 1004–1012. [Google Scholar] [CrossRef]
- Wang, E.; Shibutani, M.; Nagahara, H.; Fukuoka, T.; Iseki, Y.; Okazaki, Y.; Kashiwagi, S.; Tanaka, H.; Maeda, K.; Hirakawa, K.; et al. Abundant Intratumoral Fibrosis Prevents Lymphocyte Infiltration into Peritoneal Metastases of Colorectal Cancer. PLoS ONE 2021, 16, e0255049. [Google Scholar] [CrossRef]
- Jiang, H.; Hegde, S.; DeNardo, D.G. Tumor-Associated Fibrosis as a Regulator of Tumor Immunity and Response to Immunotherapy. Cancer Immunol. Immunother. 2017, 66, 1037–1048. [Google Scholar] [CrossRef]
- Nissen, N.I.; Johansen, A.Z.; Chen, I.; Johansen, J.S.; Pedersen, R.S.; Hansen, C.P.; Karsdal, M.A.; Willumsen, N. Collagen Biomarkers Quantify Fibroblast Activity In Vitro and Predict Survival in Patients with Pancreatic Ductal Adenocarcinoma. Cancers 2022, 14, 819. [Google Scholar] [CrossRef]
- Chen, I.M.; Willumsen, N.; Dehlendorff, C.; Johansen, A.Z.; Jensen, B.V.; Hansen, C.P.; Hasselby, J.P.; Bojesen, S.E.; Pfeiffer, P.; Nielsen, S.E.; et al. Clinical Value of Serum Hyaluronan and Propeptide of Type III Collagen in Patients with Pancreatic Cancer. Int. J. Cancer 2020, 146, 2913–2922. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Frost, D.B.; Mlakar, L.; Heywood, J.; da Silveira, W.A.; Hardiman, G.; Feghali-Bostwick, C. A Human Skin Model Recapitulates Systemic Sclerosis Dermal Fibrosis and Identifies COL22A1 as a TGFβ Early Response Gene That Mediates Fibroblast to Myofibroblast Transition. Genes 2019, 10, 75. [Google Scholar] [CrossRef]
- Baker Frost, D.; Savchenko, A.; Ogunleye, A.; Armstrong, M.; Feghali-Bostwick, C. Elucidating the Cellular Mechanism for E2-Induced Dermal Fibrosis. Arthritis Res. Ther. 2021, 23, 1–10. [Google Scholar] [CrossRef]
- Amenta, P.S.; Hadad, S.; Lee, M.T.; Barnard, N.; Li, D.; Myers, J.C. Loss of Types XV and XIX Collagen Precedes Basement Membrane Invasion in Ductal Carcinoma of the Female Breast. J. Pathol. 2003, 199, 298–308. [Google Scholar] [CrossRef]
- Myers, J.C.; Li, D.; Bageris, A.; Abraham, V.; Dion, A.S.; Amenta, P.S. Biochemical and Immunohistochemical Characterization of Human Type XIX Defines a Novel Class of Basement Membrane Zone Collagens. Am. J. Pathol. 1997, 151, 1729–1740. [Google Scholar] [PubMed]
- Ton, Q.V.; Leino, D.; Mowery, S.A.; Bredemeier, N.O.; Lafontant, P.J.; Lubert, A.; Gurung, S.; Farlow, J.L.; Foroud, T.M.; Broderick, J.; et al. Collagen COL22A1 Maintains Vascular Stability and Mutations in COL22A1 Are Potentially Associated with Intracranial Aneurysms. Dis. Model. Mech. 2018, 11, dmm033654. [Google Scholar] [CrossRef]
- Oudart, J.-B.; Monboisse, J.-C.; Maquart, F.-X.; Brassart, B.; Brassart-Pasco, S.; Ramont, L. Type XIX Collagen: A New Partner in the Interactions between Tumor Cells and Their Microenvironment. Matrix Biol. 2017, 57, 169–177. [Google Scholar] [CrossRef]
- Oudart, J.-B.; Doué, M.; Vautrin, A.; Brassart, B.; Sellier, C.; Dupont-Deshorgue, A.; Monboisse, J.-C.; Maquart, F.-X.; Brassart-Pasco, S.; Ramont, L. The Anti-Tumor NC1 Domain of Collagen XIX Inhibits the FAK/PI3K/Akt/MTOR Signaling Pathway through Avβ3 Integrin Interaction. Oncotarget 2016, 7, 1516–1528. [Google Scholar] [CrossRef]
- Zwolanek, D.; Veit, G.; Eble, J.A.; Gullberg, D.; Ruggiero, F.; Heino, J.; Meier, M.; Stetefeld, J.; Koch, M. Collagen XXII Binds to Collagen-Binding Integrins via the Novel Motifs GLQGER and GFKGER. Biochem. J. 2014, 459, 217–227. [Google Scholar] [CrossRef]
- Ricard-Blum, S. The Collagen Family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef]
- Shaw, L.M.; Olsen, B.R. FACIT Collagens: Diverse Molecular Bridges in Extracellular Matrices. Trends Biochem. Sci. 1991, 16, 191–194. [Google Scholar] [CrossRef]
- Holländer, N.; Sauerbrei, W.; Schumacher, M. Confidence Intervals for the Effect of a Prognostic Factor after Selection of an ‘Optimal’ Cutpoint. Stat. Med. 2004, 23, 1701–1713. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy Subjects (n = 33) | Patients with Cancer (n = 220) | |
|---|---|---|
| Healthy | 33 (100%) | |
| Diagnosis | ||
| Bladder cancer | 20 (9%) | |
| Breast cancer | 20 (9%) | |
| Colorectal cancer | 20 (9%) | |
| Gastric cancer | 20 (9%) | |
| Head and neck cancer | 20 (9%) | |
| Lung cancer | 20 (9%) | |
| Melanoma | 20 (9%) | |
| Ovarian cancer | 20 (9%) | |
| Pancreatic cancer | 20 (9%) | |
| Prostate cancer | 20 (9%) | |
| Renal cancer | 20 (9%) | |
| Stage | ||
| I | 7 (3%) | |
| II | 46 (21%) | |
| III | 93 (42%) | |
| IV | 74 (34%) | |
| Age (years) | ||
| Mean (SD) | 58 (6) | 59 (11) |
| Median (Min; Max) | 57 (49; 69) | 61 (30; 87) |
| Missing | 1 (0.5%) | |
| Sex | ||
| Male | 21 (64%) | 119 (54%) |
| Female | 12 (36%) | 101 (46%) |
| Healthy Subjects (n = 20) | Patients with PDAC (n = 34) | |
|---|---|---|
| Stage | ||
| I | 1 (3%) | |
| II | 7 (20%) | |
| III | 7 (20%) | |
| IV | 19 (56%) | |
| Missing | 20 (100%) | |
| Age (years) | ||
| Median (Min; Max) | 58 (45; 72) | 69 (52; 79) |
| Sex | ||
| Male | 10 (50%) | 18 (53%) |
| Female | 10 (50%) | 16 (47%) |
| Performance status | ||
| 0 | 14 (41%) | |
| 1 | 12 (35%) | |
| 2 | 3 (9%) | |
| Unknown | 5 (15%) |
| Test | Result |
|---|---|
| Measurement range | 1–53 ng/mL |
| Lower limit of quantification (LLOQ) in serum | 2 ng/mL |
| Dilution recovery of serum (1:2 to 1:4) | 104% |
| Spiking recovery of peptide in serum | 105% |
| Spiking recovery of serum in serum | 97% |
| Biotin interference recovery, low/high concentration | 99%/85% |
| Lipid interference recovery, low/high concentration | 105%/102% |
| Hemoglobin recovery, low/high concentration | 101%/113% |
| Inter-assay variation | 4% |
| Intra-assay variation | 4% |
| Analyte stability (48 h 4 °C/48 h 20 °C) | 107%/106% |
| Freeze-thaw stability up to five cycles | 103% |
| Kit stability (24 h 20 °C) | 94% |
|
Solid Tumor Type (n = 20) |
AUROC (95% CI) |
Sensitivity% (95% CI) |
Specificity% (95% CI) |
| Bladder | 0.96 (0.87–1.00) | 95 (75.1–99.9) | 91 (75.7–98.1) |
| Breast | 0.94 (0.83–0.98) | 95 (75.1–99.9) | 88 (71.8–96.6) |
| Colorectal | 0.98 (0.88–1.00) | 100 (83.2–100.0) | 91 (75.7–98.1) |
| Head and Neck | 0.95 (0.85–0.99) | 100 (83.2–100.0) | 85 (68.1–94.9) |
| Gastric | 0.94 (0.84–0.99) | 95 (75.1–99.9) | 88 (71.8–96.6) |
| Lung | 0.98 (0.89–1.00) | 100 (83.2–100.0) | 91 (75.7–98.1) |
| Melanoma | 0.89 (0.77–0.96) | 90 (68.3–98.8) | 85 (68.1–94.9) |
| Ovarian | 0.96 (0.86–0.99) | 100 (83.2–100.0) | 85 (68.1–94.9) |
| Pancreatic | 0.94 (0.84–0.99) | 100 (83.2–100.0) | 82 (64.5–93.0) |
| Prostate | 0.90 (0.79–0.96) | 90 (68.3–98.8) | 82 (64.5–93.0) |
| Renal | 0.97 (0.88–1.00) | 100 (83.2–100.0) | 85 (68.1–94.9) |
| Variable | Cut-Point | HR | 95% CI | p |
|---|---|---|---|---|
| PRO-C22 | T3 vs. T1 + T2 | 4.52 | 1.90 to 10.7 | 0.0006 |
| PRO-C3 | T3 vs. T1 + T2 | 2.15 | 0.99 to 4.68 | 0.0527 |
| Stage | Stage IV vs. Stage I–III | 3.26 | 1.53 to 6.96 | 0.0022 |
| Age | Continuous | 1.02 | 0.97 to 1.06 | 0.4805 |
| Sex | Female vs. Male | 0.67 | 0.32 to 1.41 | 0.2957 |
| PRO-C22 | Cut-Point | HR | 95% CI | p |
|---|---|---|---|---|
| Co-variate | ||||
| Stage (IV vs. I–III) | T3 vs. T1 + T2 | 3.12 | 1.24 to 7.85 | 0.0159 |
| PRO-C3 | Cut-Point | HR | 95% CI | p |
|---|---|---|---|---|
| Co-variate | ||||
| Stage (IV vs. I–III) | T3 vs. T1 + T2 | 2.91 | 1.26 to 6.71 | 0.0121 |
| PRO-C22 | Cut-Point | HR | 95% CI | p |
|---|---|---|---|---|
| Co-variate | ||||
| PRO-C3 (T3 vs. T1 + T2) | T3 vs. T1 + T2 | 4.27 | 1.76 to 10.4 | 0.0013 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madsen, E.A.; Thorlacius-Ussing, J.; Nissen, N.I.; Jensen, C.; Chen, I.M.; Johansen, J.S.; Diab, H.M.H.; Jørgensen, L.N.; Hansen, C.P.; Karsdal, M.A.; et al. Type XXII Collagen Complements Fibrillar Collagens in the Serological Assessment of Tumor Fibrosis and the Outcome in Pancreatic Cancer. Cells 2022, 11, 3763. https://doi.org/10.3390/cells11233763
Madsen EA, Thorlacius-Ussing J, Nissen NI, Jensen C, Chen IM, Johansen JS, Diab HMH, Jørgensen LN, Hansen CP, Karsdal MA, et al. Type XXII Collagen Complements Fibrillar Collagens in the Serological Assessment of Tumor Fibrosis and the Outcome in Pancreatic Cancer. Cells. 2022; 11(23):3763. https://doi.org/10.3390/cells11233763
Chicago/Turabian StyleMadsen, Emilie A., Jeppe Thorlacius-Ussing, Neel I. Nissen, Christina Jensen, Inna M. Chen, Julia S. Johansen, Hadi M. H. Diab, Lars N. Jørgensen, Carsten P. Hansen, Morten A. Karsdal, and et al. 2022. "Type XXII Collagen Complements Fibrillar Collagens in the Serological Assessment of Tumor Fibrosis and the Outcome in Pancreatic Cancer" Cells 11, no. 23: 3763. https://doi.org/10.3390/cells11233763
APA StyleMadsen, E. A., Thorlacius-Ussing, J., Nissen, N. I., Jensen, C., Chen, I. M., Johansen, J. S., Diab, H. M. H., Jørgensen, L. N., Hansen, C. P., Karsdal, M. A., & Willumsen, N. (2022). Type XXII Collagen Complements Fibrillar Collagens in the Serological Assessment of Tumor Fibrosis and the Outcome in Pancreatic Cancer. Cells, 11(23), 3763. https://doi.org/10.3390/cells11233763