
TFEB Regulates ATP7B Expression to Promote Platinum Chemoresistance in Human Ovarian Cancer Cells

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Transfection
2.2. MTT Cell Viability Assay
2.3. Immunofluorescence
2.4. DNA Adduct Evaluation by Dot Blot
2.5. Quantitative Real-Time PCR (RT-qPCR)
- β-ACTIN forward: (5′-AAGAGCTACGAGCTGCCTGA-3′);
- β-ACTIN reverse: (5′-GACTCCATGCCCAGGAAGG-3′);
- ATP7B forward: (5′-TCTCTGGTCATCCTGGTGGTT-3′);
- ATP7B reverse: (5′-GGGCTTCTGAGGTTTTGCTCT-3′);
- TFEB forward: (5′-CAAGGCCAATGACCTGGAC-3′);
- TFEB reverse: (5′-AGCTCCCTGGACTTTTGCAG-3′)
2.6. Western Blotting
2.7. Chromatin Immunoprecipitation Assays (ChIP)
- ATP6V1H Forward: (5′-TCGGGAATCCAGTTGTCCG-3′);
- ATP6V1H Reverse: (5′-GCCGCACAGGTAGAAGGAA-3′);
- ChIP3 Forward: (5′-GCGTGAGATCCCAGTACAGTGT-3′);
- ChIP3 Reverse: (5′-ACAATGTCCTCTGCCGTGCC-3′);
- ChIP4 Forward: (5′-GCGCAACTTTGAATCATCCGT-3′);
- ChIP4 Reverse: (5′-TAAAGCAAACAGGGGTCCGG-3′);
- ChIP5/6 Forward: (5′-CAGGCGCAGAGTCCGAGGAG-3′);
- ChIP6 Reverse: (5′-CTGTACTGGGATCTCACGCG-3′);
- ChIP8 Forward: (5′-CCAAATGAAGGGGCGGTTCC-3′);
- ChIP8 Reverse: (5′-GAGGAGGCGCAGAGTGTGAG-3′);
- ChIP10 Forward: (5′-CCAGGTGACCTTTTGCTCTGA-3′);
- ChIP10 Reverse: (5′-TGGCTGTGATCTGTCTCTCCT-3′);
- ChIP11 Forward: (5′-ATGTCTTGGCGTGGGAGAAAA-3′);
- ChiP11 Reverse: (5′-ATGAATTTTGAGGTGCGGGGT-3′);
- ChIP12 Forward: (5′-CAGCTCCAGGGATGTCTTGG-3′);
- ChIP12 Reverse: (5′-TCTCCAGCATCAGACCCCTT-3′).
2.8. Bioinformatic Prediction of TFEB Binding Sites
2.9. Vector Constructs and Reporter Assays
- ATP7B full (XhoI)Forward: (5′CTAGCCCGGGCTCGAGCGTGGCCTGTGATTGACAGC-3′);
- ATP7B full (HINDIII) Reverse: (5′-CCGGAATGCCAAGCTTTTAGGGCTCTAGCACAGCAACTCG-3′).
- ATP7B (CLEAR 1 XhoI) Forward: (5′-CTCGAGGCGATCGATTTTCCAGGTGCGGA-3′);
- ATP7B (CLEAR1 HINDIII) Reverse: (5′-TTCGAA CAGTGCCACAATGTCCTCTG-3′);
- ATP7B (CLEAR2 XhoI) Forward: (5′-CTCGAGC GCGCGCAACTTTGAATCATC-3′);
- ATP7B (CLEAR2 HINDIII) Reverse: (5′-TTCGAATGGCTGTGATCTGTCTCTCC-3′);
- ATP7B (CLEAR3 XhoI) Forward: (5′-CTCGAGGC GCCAGTCGGAAAGTGAGTTT-3′);
- ATP7B (CLEAR3 HINDIII) Reverse: (5′-TTCGAATTAGGGCTCTAGCACAGCAAC-3′).
2.10. Statistical Analysis
3. Results
3.1. TFEB Regulates ATP7B Expression in Pt-Resistant Ovarian Cancer Cells
3.2. ATP7B Gene Is a Direct Target of TFEB
3.3. TFEB Stimulates ATP7B Expression in a Pt-Dependent Manner
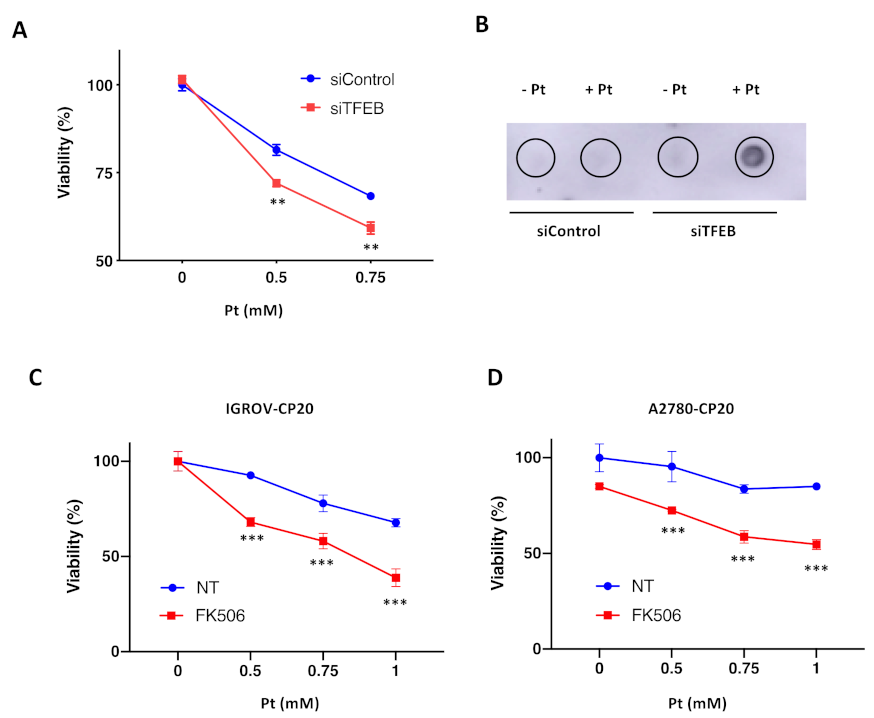
3.4. TFEB Suppression Promotes Cisplatin Toxicity in Resistant Ovarian Cancer Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Polishchuk, R.; Lutsenko, S. Golgi in copper homeostasis: A view from the membrane trafficking field. Histochem. Cell Biol. 2013, 140, 285–295. [Google Scholar] [CrossRef]
- Nevitt, T.; Öhrvik, H.; Thiele, D.J. Charting the travels of copper in eukaryotes from yeast to mammals. Biochim. Biophys. Acta 2012, 1823, 1580–1593. [Google Scholar] [CrossRef]
- Brady, D.C.; Crowe, M.; Turski, M.L.; Hobbs, A.; Yao, X.; Chaikuad, A.; Knapp, S.; Xiao, K.; Campbell, S.; Thiele, D.J.; et al. Copper is required for oncogenic BRAF signalling and tumorigenesis. Nature 2014, 509, 492–496. [Google Scholar] [CrossRef]
- Krishnamoorthy, L.; Cotruvo, J.A., Jr.; Chan, J.; Kaluarachchi, H.; Muchenditsi, A.; Pendyala, V.S.; Jia, S.; Aron, A.T.; Ackerman, C.M.; Vander Wal, M.N.; et al. Copper regulates cyclic-AMP-dependent lipolysis. Nat. Chem. Biol. 2016, 12, 586–592. [Google Scholar] [CrossRef]
- Tsang, T.; Posimo, J.M.; Gudiel, A.A.; Cicchini, M.; Feldser, D.M.; Brady, D.C. Copper is an essential regulator of the autophagic kinases ULK1/2 to drive lung adenocarcinoma. Nat. Cell Biol. 2020, 22, 412–424. [Google Scholar] [CrossRef] [PubMed]
- Gitlin, J.D. Wilson disease. Gastroenterology 2003, 125, 1868–1877. [Google Scholar] [CrossRef]
- Miyashita, H.; Nitta, Y.; Mori, S.; Kanzaki, A.; Nakayama, K.; Terada, K.; Sugiyama, T.; Kawamura, H.; Sato, A.; Morikawa, H.; et al. Expression of copper-transporting P-type adenosine triphosphatase (ATP7B) as a chemoresistance marker in human oral squamous cell carcinoma treated with cisplatin. Oral Oncol. 2003, 39, 157–162. [Google Scholar] [CrossRef]
- Nakayama, K.; Kanzaki, A.; Terada, K.; Mutoh, M.; Ogawa, K.; Sugiyama, T.; Takenoshita, S.; Itoh, K.; Yaegashi, N.; Miyazaki, K.; et al. Prognostic Value of the Cu-Transporting ATPase in Ovarian Carcinoma Patients Receiving Cisplatin-Based Chemotherapy. Clin. Cancer Res. 2004, 10, 2804–2811. [Google Scholar] [CrossRef]
- Safaei, R. Role of copper transporters in the uptake and efflux of platinum containing drugs. Cancer Lett. 2006, 234, 34–39. [Google Scholar] [CrossRef]
- Gupta, A.; Lutsenko, S. Human copper transporters: Mechanism, role in human diseases and therapeutic potential. Futur. Med. Chem. 2009, 1, 1125–1142. [Google Scholar] [CrossRef] [PubMed]
- Petruzzelli, R.; Polishchuk, R.S. Activity and Trafficking of Copper-Transporting ATPases in Tumor Development and Defense against Platinum-Based Drugs. Cells 2019, 8, 1080. [Google Scholar] [CrossRef] [PubMed]
- Mariniello, M.; Petruzzelli, R.; Wanderlingh, L.G.; La Montagna, R.; Carissimo, A.; Pane, F.; Amoresano, A.; Ilyechova, E.Y.; Galagudza, M.M.; Catalano, F.; et al. Synthetic Lethality Screening Identifies FDA-Approved Drugs that Overcome ATP7B-Mediated Tolerance of Tumor Cells to Cisplatin. Cancers 2020, 12, 608. [Google Scholar] [CrossRef] [PubMed]
- Amable, L. Cisplatin resistance and opportunities for precision medicine. Pharmacol. Res. 2016, 106, 27–36. [Google Scholar] [CrossRef]
- Galluzzi, L.; Senovilla, L.; Vitale, I.; Michels, J.; Martins, I.; Kepp, O.; Castedo, M.; Kroemer, G. Molecular mechanisms of cisplatin resistance. Oncogene 2012, 31, 1869–1883. [Google Scholar] [CrossRef] [PubMed]
- Kuo, M.T. Roles of Multidrug Resistance Genes in Breast Cancer Chemoresistance. Adv. Exp. Med. Biol. 2007, 608, 23–30. [Google Scholar] [CrossRef]
- Komatsu, M.; Sumizawa, T.; Mutoh, M.; Chen, Z.S.; Terada, K.; Furukawa, T.; Yang, X.L.; Gao, H.; Miura, N.; Sugiyama, T.; et al. Copper-transporting P-type adenosine triphosphatase (ATP7B) is associated with cisplatin resistance. Cancer Res. 2000, 60, 1312–1316. [Google Scholar]
- Polishchuk, E.V.; Concilli, M.; Iacobacci, S.; Chesi, G.; Pastore, N.; Piccolo, P.; Paladino, S.; Baldantoni, D.; Van Ijzendoorn, S.C.D.; Chan, J.; et al. Wilson Disease Protein ATP7B Utilizes Lysosomal Exocytosis to Maintain Copper Homeostasis. Dev. Cell 2014, 29, 686–700. [Google Scholar] [CrossRef]
- Safaei, R.; Larson, B.J.; Cheng, T.C.; Gibson, M.A.; Otani, S.; Naerdemann, W.; Howell, S.B. Abnormal lysosomal trafficking and enhanced exosomal export of cisplatin in drug-resistant human ovarian carcinoma cells. Mol. Cancer Ther. 2005, 4, 1595–1604. [Google Scholar] [CrossRef]
- Tadini-Buoninsegni, F.; Bartolommei, G.; Moncelli, M.R.; Inesi, G.; Galliani, A.; Sinisi, M.; Losacco, M.; Natile, G.; Arnesano, F. Translocation of Platinum Anticancer Drugs by Human Copper ATPases ATP7A and ATP7B. Angew. Chem. Int. Ed. 2014, 53, 1297–1301. [Google Scholar] [CrossRef]
- Ohbu, M.; Ogawa, K.; Konno, S.; Kanzaki, A.; Terada, K.; Sugiyama, T.; Takebayashi, Y. Copper-transporting P-type adenosine triphosphatase (ATP7B) is expressed in human gastric carcinoma. Cancer Lett. 2003, 189, 33–38. [Google Scholar] [CrossRef]
- Polishchuk, E.V.; Polishchuk, R.S. The emerging role of lysosomes in copper homeostasis. Metallomics 2016, 8, 853–862. [Google Scholar] [CrossRef]
- Polishchuk, R.S.; Polishchuk, E.V. From and to the Golgi—Defining the Wilson disease protein road map. FEBS Lett. 2019, 593, 2341–2350. [Google Scholar] [CrossRef] [PubMed]
- Safaei, R.; Otani, S.; Larson, B.J.; Rasmussen, M.L.; Howell, S.B. Transport of Cisplatin by the Copper Efflux Transporter ATP7B. Mol. Pharmacol. 2008, 73, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Zhitomirsky, B.; Assaraf, Y.G. Lysosomes as mediators of drug resistance in cancer. Drug Resist. Updat. 2016, 24, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, G.; Ballabio, A. TFEB at a glance. J. Cell Sci. 2016, 129, 2475–2481. [Google Scholar] [CrossRef]
- Chu, H.-Y.; Wang, W.; Chen, X.; Jiang, Y.-E.; Cheng, R.; Qi, X.; Zhong, Z.-M.; Zeng, M.-S.; Zhu, X.-F.; Sun, C.-Z. Bafilomycin A1 increases the sensitivity of tongue squamous cell carcinoma cells to cisplatin by inhibiting the lysosomal uptake of platinum ions but not autophagy. Cancer Lett. 2018, 423, 105–112. [Google Scholar] [CrossRef]
- Polishchuk, E.V.; Merolla, A.; Lichtmannegger, J.; Romano, A.; Indrieri, A.; Ilyechova, E.Y.; Concilli, M.; De Cegli, R.; Crispino, R.; Mariniello, M.; et al. Activation of autophagy, observed in liver tissues from patients with wilson disease and from ATP7B-deficient animals, protects hepatocytes from copper-induced apoptosis. Gastroenterology 2019, 156, 1173–1189.e1175. [Google Scholar] [CrossRef]
- Petruzzelli, R.; Christensen, D.R.; Parry, K.L.; Sanchez-Elsner, T.; Houghton, F.D. HIF-2α Regulates NANOG Expression in Human Embryonic Stem Cells following Hypoxia and Reoxygenation through the Interaction with an Oct-Sox Cis Regulatory Element. PLoS ONE 2014, 9, e108309. [Google Scholar] [CrossRef]
- TFEB Database. Available online: http://tfeb.tigem.it (accessed on 10 November 2021).
- Lee, C.M.; Barber, G.P.; Casper, J.; Clawson, H.; Diekhans, M.; Gonzalez, J.N.; Hinrichs, A.; Lee, B.T.; Nassar, L.R.; Powell, C.C.; et al. UCSC Genome Browser enters 20th year. Nucleic Acids Res. 2020, 48, D756–D761. [Google Scholar] [CrossRef]
- Gambardella, G.; Staiano, L.; Moretti, M.N.; De Cegli, R.; Fagnocchi, L.; Di Tullio, G.; Polletti, S.; Braccia, C.; Armirotti, A.; Zippo, A.; et al. GADD34 is a modulator of autophagy during starvation. Sci. Adv. 2020, 6, eabb0205. [Google Scholar] [CrossRef]
- Sardiello, M.; Palmieri, M.; Di Ronza, A.; Medina, D.L.; Valenza, M.; Gennarino, V.A.; Di Malta, C.; Donaudy, F.; Embrione, V.; Polishchuk, R.S.; et al. A Gene Network Regulating Lysosomal Biogenesis and Function. Science 2009, 325, 473–477. [Google Scholar] [CrossRef]
- Palmieri, M.; Impey, S.; Kang, H.; di Ronza, A.; Pelz, C.; Sardiello, M.; Ballabio, A. Characterization of the CLEAR network reveals an integrated control of cellular clearance pathways. Hum. Mol. Genet. 2011, 20, 3852–3866. [Google Scholar] [CrossRef]
- Stewart, D.J.; Short, K.K.; Maniaci, B.; Burkhead, J.L. COMMD1 and PtdIns(4,5)P2 interaction maintain ATP7B copper transporter trafficking fidelity in HepG2 cells. J. Cell Sci. 2019, 132, jcs.231753. [Google Scholar] [CrossRef]
- Das, S.; Maji, S.; Ruturaj, R.; Bhattacharya, I.; Saha, T.; Naskar, N.; Gupta, A. Retromer retrieves the Wilson Disease protein ATP7B from endolysosomes in a copper-dependent mode. J. Cell Sci. 2020, 133, jcs246819. [Google Scholar] [CrossRef] [PubMed]
- Ezer, D.; Zabet, N.R.; Adryan, B. Homotypic clusters of transcription factor binding sites: A model system for understanding the physical mechanics of gene expression. Comput. Struct. Biotechnol. J. 2014, 10, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Höflich, C.; Brieger, A.; Zeuzem, S.; Plotz, G. Investigation of the Wilson gene ATP7B transcriptional start site and the effect of core promoter alterations. Sci. Rep. 2021, 11, 76741. [Google Scholar] [CrossRef] [PubMed]
- Medina, D.L.; Di Paola, S.; Peluso, I.; Armani, A.; De Stefani, D.; Venditti, R.; Montefusco, S.; Rosato, A.S.; Prezioso, C.; Forrester, A.; et al. Lysosomal calcium signalling regulates autophagy through calcineurin and TFEB. Nat. Cell Biol. 2015, 17, 288–299. [Google Scholar] [CrossRef]
- Mangala, L.S.; Zuzel, V.; Schmandt, R.; LeShane, E.S.; Halder, J.B.; Armaiz-Pena, G.N.; Spannuth, W.A.; Tanaka, T.; Shahzad, M.M.; Lin, Y.G.; et al. Therapeutic Targeting of ATP7B in Ovarian Carcinoma. Clin. Cancer Res. 2009, 15, 3770–3780. [Google Scholar] [CrossRef]
- Puchkova, L.V.; Broggini, M.; Polishchuk, E.V.; Ilyechova, E.Y.; Polishchuk, R.S. Silver Ions as a Tool for Understanding Different Aspects of Copper Metabolism. Nutrients 2019, 11, 1364. [Google Scholar] [CrossRef]
- Zhitomirsky, B.; Assaraf, Y.G. Lysosomal sequestration of hydrophobic weak base chemotherapeutics triggers lysosomal biogenesis and lysosome-dependent cancer multidrug resistance. Oncotarget 2014, 6, 1143–1156. [Google Scholar] [CrossRef]
- Rodrigo, M.A.M.; Buchtelova, H.; Rios, V.D.L.; Casal, J.I.; Eckschlager, T.; Hrabeta, J.; Belhajova, M.; Heger, Z.; Adam, V. Proteomic Signature of Neuroblastoma Cells UKF-NB-4 Reveals Key Role of Lysosomal Sequestration and the Proteasome Complex in Acquiring Chemoresistance to Cisplatin. J. Proteome Res. 2019, 18, 1255–1263. [Google Scholar] [CrossRef]
- Kauffman, E.C.; Ricketts, C.J.; Rais-Bahrami, S.; Yang, Y.; Merino, M.J.; Bottaro, D.P.; Srinivasan, R.; Linehan, W.M. Molecular genetics and cellular features of TFE3 and TFEB fusion kidney cancers. Nat. Rev. Urol. 2014, 11, 465–475. [Google Scholar] [CrossRef]
- Sakamoto, H.; Yamashita, K.; Okamoto, K.; Kadowaki, T.; Sakai, E.; Umeda, M.; Tsukuba, T. Transcription factor EB influences invasion and migration in oral squamous cell carcinomas. Oral Dis. 2018, 24, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, A.; Bianconi, V.; Pirro, M.; Orafai, H.M.; Sahebkar, A. The role of TFEB in tumor cell autophagy: Diagnostic and therapeutic opportunities. Life Sci. 2020, 244, 117341. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, X.; Wang, S.; Chen, Y.; Liu, H. Regulation of TFEB activity and its potential as a therapeutic target against kidney diseases. Cell Death Discov. 2020, 6, 32. [Google Scholar] [CrossRef]
- Giatromanolaki, A.; Kalamida, D.; Sivridis, E.; Karagounis, I.V.; Gatter, K.C.; Harris, A.L.; Koukourakis, M.I. Increased expression of transcription factor EB (TFEB) is associated with autophagy, migratory phenotype and poor prognosis in non-small cell lung cancer. Lung Cancer 2015, 90, 98–105. [Google Scholar] [CrossRef]
- Perera, R.M.; Stoykova, S.; Nicolay, B.N.; Ross, K.N.; Fitamant, J.; Boukhali, M.; Lengrand, J.; Deshpande, V.; Selig, M.K.; Ferrone, C.R.; et al. Transcriptional control of autophagy–lysosome function drives pancreatic cancer metabolism. Nature 2015, 524, 361–365. [Google Scholar] [CrossRef]
- Klein, K.; Werner, K.; Teske, C.; Schenk, M.; Giese, T.; Weitz, J.; Welsch, T. Role of TFEB-driven autophagy regulation in pancreatic cancer treatment. Int. J. Oncol. 2016, 49, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Giatromanolaki, A.; Sivridis, E.; Kalamida, D.; Koukourakis, M.I. Transcription Factor EB Expression in Early Breast Cancer Relates to Lysosomal/Autophagosomal Markers and Prognosis. Clin. Breast Cancer 2017, 17, e119–e125. [Google Scholar] [CrossRef]
- Gallon, J.; Loomis, E.; Curry, E.; Martin, N.; Brody, L.; Garner, I.; Brown, R.; Flanagan, J.M. Chromatin accessibility changes at intergenic regions are associated with ovarian cancer drug resistance. Clin. Epigenetics 2021, 13, 122. [Google Scholar] [CrossRef]
- Poisson, L.M.; Munkarah, A.; Madi, H.; Datta, I.; Hensley-Alford, S.; Tebbe, C.; Buekers, T.; Giri, S.; Rattan, R. A metabolomic approach to identifying platinum resistance in ovarian cancer. J. Ovarian Res. 2015, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Bergamini, A.; Bocciolone, L.; Fodor, A.; Candiani, M.; Mangili, G. Management of recurrent ovarian cancer: When platinum-based regimens are not a therapeutic option. Int. J. Gynecol. Cancer 2019, 29, 1431–1436. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petruzzelli, R.; Mariniello, M.; De Cegli, R.; Catalano, F.; Guida, F.; Di Schiavi, E.; Polishchuk, R.S. TFEB Regulates ATP7B Expression to Promote Platinum Chemoresistance in Human Ovarian Cancer Cells. Cells 2022, 11, 219. https://doi.org/10.3390/cells11020219
Petruzzelli R, Mariniello M, De Cegli R, Catalano F, Guida F, Di Schiavi E, Polishchuk RS. TFEB Regulates ATP7B Expression to Promote Platinum Chemoresistance in Human Ovarian Cancer Cells. Cells. 2022; 11(2):219. https://doi.org/10.3390/cells11020219
Chicago/Turabian StylePetruzzelli, Raffaella, Marta Mariniello, Rossella De Cegli, Federico Catalano, Floriana Guida, Elia Di Schiavi, and Roman S. Polishchuk. 2022. "TFEB Regulates ATP7B Expression to Promote Platinum Chemoresistance in Human Ovarian Cancer Cells" Cells 11, no. 2: 219. https://doi.org/10.3390/cells11020219
APA StylePetruzzelli, R., Mariniello, M., De Cegli, R., Catalano, F., Guida, F., Di Schiavi, E., & Polishchuk, R. S. (2022). TFEB Regulates ATP7B Expression to Promote Platinum Chemoresistance in Human Ovarian Cancer Cells. Cells, 11(2), 219. https://doi.org/10.3390/cells11020219