
Potential Therapeutic Role of Mesenchymal-Derived Stem Cells as an Alternative Therapy to Combat COVID-19 through Cytokines Storm

 , ,
, ,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Cytokines
3. COVID-19 and the Cytokine Storm
4. Stem Cells and Stem Cell Therapy
5. Stem Cell Therapy for COVID-19 and Cytokine Storm
6. Challenges in Stem Cell Therapy
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ARDS | Acute respiratory distress syndrome |
| MSCs | Mesenchymal stem cells |
| IL | Interleukin |
| (LT) | Leukotriene |
| CRPs | C-reactive proteins |
| ESCs | Embryonic stem cells |
| VEGF | Vascular endothelial growth factor |
| FGF | Fibroblast growth factor |
| TNFs | Tumor Necrosis Factors |
| HGF | Hepatocyte growth factor |
| HSCs | Hematopoietic Stem Cells |
| ESCs | Epithelial Stem Cells |
| NSCs | Neural Stem Cells |
| NKA | Numb-associated kinases |
| ACE-2 | Angiotensin-converting enzyme-2 |
| ASCs | Adult Stem Cells |
| GSCF | Granulocyte Colony-Stimulating Factor |
| iPSCs | Induced Pluripotent Stem Cells |
| MIP1A | Macrophage Inflammatory Protein 1 α |
References
- World Health Organization (WHO). Novel Coronavirus (2019-nCoV) Situation Report 22. 2020. Available online: https://www.who.int/docs/default-source/coronaviruse/situationreports/20200211-sitrep-22-ncov.pdf?sfvrsn$=$fb6d49b1_2 (accessed on 1 April 2020).
- Goel, H.; Goyal, K.; Baranwal, P.; Dixit, A.; Upadhyay, T.K.; Upadhye, V.J. The diagnostics technologies and control of COVID-19. Lett. Appl. NanoBioSci. 2021, 11, 3120–3133. [Google Scholar] [CrossRef]
- Chen, N.; Zhou, M.; Dong, X.; Qu, J.; Gong, F.; Han, Y.; Qiu, Y.; Wang, J.; Liu, Y.; Wei, Y.; et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: A descriptive study. Lancet 2020, 395, 507–513. [Google Scholar] [CrossRef]
- Goyal, K.; Goel, H.; Baranwal, P.; Tewary, A.; Dixit, A.; Pandey, A.K.; Benjamin, M.; Tanwar, P.; Dey, A.; Khan, F.; et al. Immunological mechanisms of vaccine-induced protection against SARS-CoV-2 in humans. Immuno 2021, 1, 442–456. [Google Scholar] [CrossRef]
- Zhang, J.; Xie, B.; Hashimoto, K. Current status of potential therapeutic candidates for the COVID-19 crisis. Brain Behav. Immun. 2020, 87, 59–73. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, H.; Ann Peer, W.; Richter, G.; Blakeslee, J.; Bandyopadhyay, A.; Titapiwantakun, B.; Undurraga, S.; Khodakovskaya, M.; Richards, E.L.; et al. Arabidopsis H+-PPase AVP1 regulates auxin-mediated organ development. Science 2005, 310, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Europe PMC. Available online: https://europepmc.org/article/PMC/PMC7839510 (accessed on 22 April 2021).
- Volz, E.; Mishra, S.; Chand, M.; Barrett, J.C.; Johnson, R.; Geidelberg, L.; Hinsley, W.R.; Laydon, D.J.; Dabrera, G.; O’Toole, Á.; et al. Transmission of SARS-CoV-2 lineage B. 1.1. 7 in England: Insights from linking epidemiological and genetic data. medRxiv 2021, 593. [Google Scholar] [CrossRef]
- Tegally, H.; Wilkinson, E.; Giovanetti, M.; Iranzadeh, A.; Fonseca, V.; Giandhari, J.; Doolabh, D.; Pillay, S.; San, E.J.; Msomi, N.; et al. Emergence and rapid spread of a new severe acute respiratory syndrome-related coronavirus 2 (SARS-CoV-2) lineage with multiple spike mutations in South Africa. medRxiv 2020. [Google Scholar] [CrossRef]
- Wibmer, C.K.; Ayres, F.; Hermanus, T.; Madzivhandila, M.; Kgagudi, P.; Oosthuysen, B.; Lambson, B.E.; De Oliveira, T.; Vermeulen, M.; Van der Berg, K.; et al. SARS-CoV-2 501Y. V2 escapes neutralization by South African COVID-19 donor plasma. Nat. Med. 2021, 27, 622–625. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Davis, B.D.; Chen, S.S.; Martinez, J.M.S.; Plummer, J.T.; Vail, E. Emergence of a novel SARS-CoV-2 strain in Southern California, USA. medRxiv 2021. [Google Scholar] [CrossRef]
- Yang, X.; Yu, Y.; Xu, J.; Shu, H.; Liu, H.; Wu, Y.; Zhang, L.; Yu, Z.; Fang, M.; Yu, T.; et al. Clinical course and outcomes of critically ill patients with SARS-CoV-2 pneumonia in Wuhan, China: A single-centered, retrospective, observational study. Lancet Respir. Med. 2020, 8, 475–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Reghunathan, R.; Jayapal, M.; Hsu, L.Y.; Chng, H.H.; Tai, D.; Leung, B.P.; Melendez, A.J. Expression profile of immune response genes in patients with severe acute respiratory syndrome. BMC Immunol. 2005, 6, 1–11. [Google Scholar] [CrossRef]
- Ding, Y.; He, L.I.; Zhang, Q.; Huang, Z.; Che, X.; Hou, J.; Wang, H.; Shen, H.; Qiu, L.; Li, Z.; et al. Organ distribution of severe acute respiratory syndrome (SARS) associated coronavirus (SARS-CoV) in SARS patients: Implications for pathogenesis and virus transmission pathways. J. Pathol. J. Pathol. Soc. Great Br. Irel. 2004, 203, 622–630. [Google Scholar] [CrossRef]
- Soy, M.; Keser, G.; Atagündüz, P.; Tabak, F.; Atagündüz, I.; Kayhan, S. Cytokine storm in COVID-19: Pathogenesis and overview of anti-inflammatory agents used in treatment. Clin. Rheumatol. 2020, 39, 2085–2094. [Google Scholar] [CrossRef]
- Leng, Z.; Zhu, R.; Hou, W.; Feng, Y.; Yang, Y.; Han, Q.; Shan, G.; Meng, F.; Du, D.; Wang, S.; et al. Transplantation of ACE2-mesenchymal stem cells improves the outcome of patients with COVID-19 pneumonia. Aging Dis. 2020, 11, 216. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef]
- Prockop, D.J.; Oh, J.Y. Mesenchymal stem/stromal cells (MSCs): Role as guardians of inflammation. Mol. Ther. 2012, 20, 14–20. [Google Scholar] [CrossRef]
- Lanzoni, G.; Linetsky, E.; Correa, D.; Messinger Cayetano, S.; Alvarez, R.A.; Kouroupis, D.; Alvarez Gil, A.; Poggioli, R.; Ruiz, P.; Marttos, A.C.; et al. Umbilical cord mesenchymal stem cells for COVID-19 acute respiratory distress syndrome: A double-blind, phase 1/2a, randomized controlled trial. Stem Cells Transl. Med. 2021, 10, 660–673. [Google Scholar] [CrossRef]
- Yilmaz, R.; Adas, G.; Cukurova, Z.; Yasar, K.K.; Isiksacan, N.; Oztel, O.N.; Karaoz, E. Mesenchymal stem cells treatment in COVID-19 patient with multi-organ involvement. Bratisl. Lek. Listy 2020, 121, 847–852. [Google Scholar] [CrossRef]
- Charo, I.F.; Ransohoff, R.M. The Many Roles of Chemokines and Chemokine Receptors in Inflammation. N. Engl. J. Med. 2006, 354, 610–621. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Immunological and Inflammatory Functions of the Interleukin-1 Family. Annu. Rev. Immunol. 2009, 27, 519–550. [Google Scholar] [CrossRef] [PubMed]
- Behrens, E.M.; Canna, S.W.; Slade, K.; Rao, S.; Kreiger, P.A.; Paessler, M.; Kambayashi, T.; Koretzky, G.A. Repeated TLR9 stimulation results in macrophage activation syndrome–like disease in mice. J. Clin. Investig. 2011, 121, 2264–2277. [Google Scholar] [CrossRef] [PubMed]
- Metcalfe, S.M. Mesenchymal stem cells and management of COVID-19 pneumonia. Drug Discov. Today 2020, 5, 100019. [Google Scholar] [CrossRef]
- Kato, T. Granulocyte colony-stimulating factor. In Handbook of Hormones; Academic Press: Cambridge, MA, USA, 2021; pp. 467–470. [Google Scholar] [CrossRef]
- Xie, M.; Zhang, S.; Dong, F.; Zhang, Q.; Wang, J.; Wang, C.; Zhu, C.; Zhang, S.; Luo, B.; Wu, P.; et al. Granulocyte colony-stimulating factor directly acts on mouse lymphoid-biased but not myeloid-biased hematopoietic stem cells. Haematologica 2021, 106, 1647–1658. [Google Scholar] [CrossRef] [PubMed]
- Modi, J.; Menzie-Suderam, J.; Xu, H.; Trujillo, P.; Medley, K.; Marshall, M.L.; Tao, R.; Prentice, H.; Wu, J.Y. Mode of action of granulocyte-colony stimulating factor (G-CSF) as a novel therapy for stroke in a mouse model. J. Biomed. Sci. 2020, 27, 1–19. [Google Scholar] [CrossRef]
- Taub, D.D.; Lloyd, A.R.; Conlon, K.; Wang, J.M.; Ortaldo, J.R.; Harada, A.; Matsushima, K.; Kelvin, D.J.; Oppenheim, J.J. Recombinant human interferon-inducible protein 10 is a chemoattractant for human monocytes and T lymphocytes and promotes T cell adhesion to endothelial cells. J. Exp. Med. 1993, 177, 1809–1814. [Google Scholar] [CrossRef]
- Taub, D.D.; Longo, D.L.; Murphy, W.J. Human interferon-inducible protein-10 induces mononuclear cell infiltration in mice and promotes the migration of human T lymphocytes into the peripheral tissues of human peripheral blood lymphocytes-SCID mice. Blood 1996, 87, 1423–1431. [Google Scholar] [CrossRef]
- Lacotte, S.; Brun, S.; Muller, S.; Dumortier, H. CXCR3, inflammation, and autoimmune diseases. Ann. N. Y. Acad. Sci. 2009, 1173, 310–317. [Google Scholar] [CrossRef]
- Cushing, S.D.; Berliner, J.A.; Valente, A.J.; Territo, M.C.; Navab, M.; Parhami, F.; Gerrity, R.; Schwartz, C.J.; Fogelman, A.M. Minimally modified low density lipoprotein induces monocyte chemotactic protein 1 in human endothelial cells and smooth muscle cells. Proc. Natl. Acad. Sci. USA 1990, 87, 5134–5138. [Google Scholar] [CrossRef]
- Widera, D.; Holtkamp, W.; Entschladen, F.; Niggemann, B.; Zänker, K.; Kaltschmidt, B.; Kaltschmidt, C. MCP-1 induces migration of adult neural stem cells. Eur. J. Cell Biol. 2004, 83, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Anshita, D.; Ravichandiran, V. MCP-1: Function, regulation, and involvement in disease. Int. Immunopharmacol. 2021, 101, 107598. [Google Scholar] [CrossRef] [PubMed]
- Menten, P.; Wuyts, A.; Van Damme, J. Macrophage inflammatory protein-1. Cytokine Growth Factor Rev. 2002, 13, 455–481. [Google Scholar] [CrossRef]
- Bhavsar, I.; Miller, C.S.; Al-Sabbagh, M. Macrophage inflammatory protein-1 alpha (MIP-1 alpha)/CCL3: As a biomarker. Gen. Methods Biomark. Res. Appl. 2015, 223–249. [Google Scholar] [CrossRef]
- Arenas-Ramirez, N.; Woytschak, J.; Boyman, O. Interleukin-2: Biology, design and application. Trends Immunol. 2015, 36, 763–777. [Google Scholar] [CrossRef]
- Liao, W.; Lin, J.X.; Leonard, W.J. IL-2 family cytokines: New insights into the complex roles of IL-2 as a broad regulator of T helper cell differentiation. Curr. Opin. Immunol. 2011, 23, 598–604. [Google Scholar] [CrossRef]
- Spolski, R.; Li, P.; Leonard, W.J. Biology and regulation of IL-2: From molecular mechanisms to human therapy. Nat. Rev. Immunol. 2018, 18, 648–659. [Google Scholar] [CrossRef]
- Srirangan, S.; Choy, E.H. The role of interleukin 6 in the pathophysiology of rheumatoid arthritis. Ther. Adv. Musculoskelet. Dis. 2010, 2, 247–256. [Google Scholar] [CrossRef]
- Dienz, O.; Rincon, M. The effects of IL-6 on CD4 T cell responses. Clin. Immunol. 2009, 130, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Green, A.M.; DiFazio, R.; Flynn, J.L. IFN-γ from CD4 T cells is essential for host survival and enhances CD8 T cell function during Mycobacterium tuberculosis infection. J. Immunol. 2013, 190, 270–277. [Google Scholar] [CrossRef]
- Zamisch, M.; Moore-Scott, B.; Su, D.M.; Lucas, P.J.; Manley, N.; Richie, E.R. Ontogeny and regulation of IL-7-expressing thymic epithelial cells. J. Immunol. 2005, 174, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Offner, F.; Plum, J. The role of interleukin-7 in early T-cell development. Leuk. Lymphoma 1998, 30, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Idriss, H.T.; Naismith, J.H. TNF-α and the TNF receptor superfamily: Structure-function relationship (s). Microsc. Res. Tech. 2000, 50, 184–195. [Google Scholar] [CrossRef]
- Kale, V.P.; Gilhooley, P.J.; Phadtare, S.; Nabavizadeh, A.; Pandey, M.K. Role of gambogic acid in chemosensitization of cancer. In Role of Nutraceuticals in Cancer Chemosensitization; Academic Press: Cambridge, MA, USA, 2018; pp. 151–167. [Google Scholar] [CrossRef]
- Mehta, A.K.; Gracias, D.T.; Croft, M. TNF activity and T cells. Cytokine 2018, 101, 14–18. [Google Scholar] [CrossRef]
- Remick, D.G. Interleukin-8. Crit. Care Med. 2005, 33, S466–S467. [Google Scholar] [CrossRef]
- Hedges, J.C.; Singer, C.A.; Gerthoffer, W.T. Mitogen-activated protein kinases regulate cytokine gene expression in human airway myocytes. Am. J. Respir. 2000, 23, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.G.; Sanders, A.J.; Ruge, F.; Harding, K.G. Influence of interleukin-8 (IL-8) and IL-8 receptors on the migration of human keratinocytes, the role of PLC-γ and potential clinical implications. Exp. Ther. Med. 2012, 3, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Loick, H.M.; Theissen, J.L. Die Eicosanoide als Mediatoren beim Ards. AINS-Anästhesiol. Intensivmed. Notf. Schmerzther. 1994, 29, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Berger, A. What are leukotrienes and how do they work in asthma? BMJ 1999, 319, 90. [Google Scholar] [CrossRef] [PubMed]
- Cuzzo, B.; Lappin, S.L. Physiology, Leukotrienes; StatPearls Publishing: Tampa, FL, USA, 2021. [Google Scholar]
- Jo-Watanabe, A.; Okuno, T.; Yokomizo, T. The role of leukotrienes as potential therapeutic targets in allergic disorders. Int. J. Mol. Sci. 2019, 20, 3580. [Google Scholar] [CrossRef]
- Tang, Y.; Liu, J.; Zhang, D.; Xu, Z.; Ji, J.; Wen, C. Cytokine storm in COVID-19: The current evidence and treatment strategies. Front. Immunol. 2020, 11, 1708. [Google Scholar] [CrossRef] [PubMed]
- Ren, K.; Torres, R. Role of interleukin-1β during pain and inflammation. Brain Res. Rev. 2009, 60, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Anforth, H.R.; Bluthe, R.M.; Bristow, A.; Hopkins, S.; Lenczowski, M.J.; Luheshi, G.; Lundkvist, J.; Michaud, B.; Mistry, Y.; Van Dam, A.M.; et al. Biological activity and brain actions of recombinant rat interleukin-1alpha and interleukin-1beta. Eur. Cytokine Netw. 1998, 9, 279–288. [Google Scholar] [PubMed]
- Heufler, C.; Koch, F.; Stanzl, U.; Topar, G.; Wysocka, M.; Trinchieri, G.; Enk, A.; Steinman, R.M.; Romani, N.; Schuler, G. Interleukin-12 is produced by dendritic cells and mediates T helper 1 development as well as interferon-γ production by T helper 1 cells. Eur. J. Immunol. 1996, 26, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Truyen, E.; Coteur, L.; Dilissen, E.; Overbergh, L.; Dupont, L.J.; Ceuppens, J.L.; Bullens, D.M. Evaluation of airway inflammation by quantitative Th1/Th2 cytokine mRNA measurement in sputum of asthma patients. Thorax 2006, 61, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Trinchieri, G. Interleukin-12 and the regulation of innate resistance and adaptive immunity. Nat. Rev. Immunol. 2003, 3, 133–146. [Google Scholar] [CrossRef]
- Drake, L.Y.; Kita, H. IL-33: Biological properties, functions, and roles in airway disease. Immunol. Rev. 2017, 278, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Halim, T.Y.; Krauß, R.H.; Sun, A.C.; Takei, F. Lung natural helper cells are a critical source of Th2 cell-type cytokines in protease allergen-induced airway inflammation. Immunity 2012, 36, 451–463. [Google Scholar] [CrossRef] [Green Version]
- Lefrançais, E.; Duval, A.; Mirey, E.; Roga, S.; Espinosa, E.; Cayrol, C.; Girard, J.P. Central domain of IL-33 is cleaved by mast cell proteases for potent activation of group-2 innate lymphoid cells. Proc. Natl. Acad. Sci. USA 2014, 111, 15502–15507. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, J.J.; Gadina, M.; Siegel, R.M. Cytokines and cytokine receptors. In Clinical immunology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 127–155.e1. [Google Scholar] [CrossRef]
- Branton, M.H.; Kopp, J.B. TGF-β and fibrosis. Microbes Infect. 1999, 1, 1349–1365. [Google Scholar] [CrossRef]
- Hassan, S.; Shah, H.; Shawana, S. Dysregulated epidermal growth factor and tumor growth factor-beta receptor signaling through GFAP-ACTA2 protein interaction in liver fibrosis. Pak. J. Med. Sci. 2020, 36, 782. [Google Scholar] [CrossRef] [PubMed]
- Chevigné, A.; Janji, B.; Meyrath, M.; Reynders, N.; D’uonnolo, G.; Uchański, T.; Xiao, M.; Berchem, G.; Ollert, M.; Kwon, Y.J.; et al. CXCL10 is an agonist of the CC family chemokine scavenger receptor ACKR2/D6. Cancers 2021, 13, 1054. [Google Scholar] [CrossRef]
- Tannenbaum, C.S.; Tubbs, R.; Armstrong, D.; Finke, J.H.; Bukowski, R.M.; Hamilton, T.A. The CXC chemokines IP-10 and Mig are necessary for IL-12-mediated regression of the mouse RENCA tumor. J. Immunol. 1998, 161, 927–932. [Google Scholar] [PubMed]
- Booth, V.; Keizer, D.W.; Kamphuis, M.B.; Clark-Lewis, I.; Sykes, B.D. The CXCR3 binding chemokine IP-10/CXCL10: Structure and receptor interactions. Biochemistry 2002, 41, 10418–10425. [Google Scholar] [CrossRef] [PubMed]
- Tau, G.; Rothman, P. Biologic functions of the IFN-γ receptors. Allergy 1999, 54, 1233. [Google Scholar] [CrossRef] [PubMed]
- Dafny, N.; Lincoln, J. The role of interferons on the central nervous system in health and disease. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar] [CrossRef]
- Baron, S.; Tyring, S.K.; Fleischmann, W.R.; Coppenhaver, D.H.; Niesel, D.W.; Klimpel, G.R.; Stanton, G.J.; Hughes, T.K. The interferons: Mechanisms of action and clinical applications. JAMA 1991, 266, 1375–1383. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A.; Novick, D.; Kim, S.; Kaplanski, G. Interleukin-18 and IL-18 binding protein. Front. Immunol. 2013, 4, 289. [Google Scholar] [CrossRef]
- O’Shea, J.J.; Gadina, M.; Richard; Siegel, M.; Farber, J. 13–Cytokines. In Rheumatology, 6th ed.; Hochberg, M.C., Silman, A.J., Smolen, J.S., Weinblatt, M.E., Weisman, M.H., Eds.; Mosby: Maryland Heights, MO, USA, 2015; pp. 99–112. [Google Scholar] [CrossRef]
- Cron, R.Q.; Behrens, E.M. Cytokine Storm Syndrome; Springer Nature: Cham, Switzerland, 2019. [Google Scholar]
- Tisoncik, J.R.; Korth, M.J.; Simmons, C.P.; Farrar, J.; Martin, T.R.; Katze, M.G. Into the eye of the cytokine storm. Microbiol. Mol. Biol. Rev. 2012, 76, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Hussman, J.P. Cellular and molecular pathways of COVID-19 and potential points of therapeutic intervention. Front. Pharmacol. 2020, 11, 1169. [Google Scholar] [CrossRef] [PubMed]
- Fajgenbaum, D.C.; June, C.H. Cytokine storm. N. Engl. J. Med. 2020, 383, 2255–2273. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.W.; Santomasso, B.D.; Locke, F.L.; Ghobadi, A.; Turtle, C.J.; Brudno, J.N.; Maus, M.V.; Park, J.H.; Mead, E.; Pavletic, S.; et al. ASTCT consensus grading for cytokine release syndrome and neurologic toxicity associated with immune effector cells. Biol. Blood Marrow Trans. 2019, 25, 625–638. [Google Scholar] [CrossRef]
- Lee, D.W.; Gardner, R.; Porter, D.L.; Louis, C.U.; Ahmed, N.; Jensen, M.; Grupp, S.A.; Mackall, C.L. Current concepts in the diagnosis and management of cytokine release syndrome. Blood Am. J. Hematol. 2014, 124, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.; Huang, Y.; Guo, Y.; Yin, M.; Chen, X.; Xiao, L.; Deng, G. Association of inflammatory markers with the severity of COVID-19: A meta-analysis. Int. J. Infect. Dis. 2020, 96, 467–474. [Google Scholar] [CrossRef]
- Potempa, L.A.; Rajab, I.M.; Hart, P.C.; Bordon, J.; Fernandez-Botran, R. Insights into the use of C-reactive protein as a diagnostic index of disease severity in COVID-19 infections. Am. J. Trop. Med. Hyg. 2020, 103, 561. [Google Scholar] [CrossRef] [PubMed]
- McElvaney, O.J.; McEvoy, N.L.; McElvaney, O.F.; Carroll, T.P.; Murphy, M.P.; Dunlea, D.M.; Ní Choileáin, O.; Clarke, J.; O’Connor, E.; Hogan, G.; et al. Characterization of the inflammatory response to severe COVID-19 illness. Am. J. Respir. Crit. Care Med. 2020, 202, 812–821. [Google Scholar] [CrossRef] [PubMed]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef]
- Mitra, P.; Misra, S.; Sharma, P. COVID-19 pandemic in India: What lies ahead? J. Clin. Biochem. 2020, 35, 257–259. [Google Scholar] [CrossRef]
- Li, X.; Geng, M.; Peng, Y.; Meng, L.; Lu, S. Molecular immune pathogenesis and diagnosis of COVID-19. J. Pharm. Anal. 2020, 10, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M. Clinical features of cytokine storm syndrome. In Cytokine Storm Syndrome; Cron, R., Behrens, E., Eds.; Springer: Cham, Switzerland, 2019; pp. 31–42. [Google Scholar] [CrossRef]
- Shimabukuro-Vornhagen, A.; Gödel, P.; Subklewe, M.; Stemmler, H.J.; Schlößer, H.A.; Schlaak, M.; Kochanek, M.; Böll, B.; von Bergwelt-Baildon, M.S. Cytokine release syndrome. J. Immunother. Cancer 2018, 6, 1–14. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. Immunotherapeutic implications of IL-6 blockade for cytokine storm. Immunotherapy 2016, 8, 959–970. [Google Scholar] [CrossRef] [PubMed]
- Hunter, C.A.; Jones, S.A. IL-6 as a keystone cytokine in health and disease. Nat. Immunol. 2015, 16, 448–457. [Google Scholar] [CrossRef]
- Sinha, P.; Matthay, M.A.; Calfee, C.S. Is a “cytokine storm” relevant to COVID-19? JAMA Intern. Med. 2020, 180, 1152–1154. [Google Scholar] [CrossRef]
- Jensen, S.; Thomsen, A.R. Sensing of RNA viruses: A review of innate immune receptors involved in recognizing RNA virus invasion. J. Virol. 2012, 86, 2900–2910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, A.; Iwasaki, A. Type I and type III interferons—Induction, signaling, evasion, and application to combat COVID-19. Cell Host Microbe 2020, 27, 870–878. [Google Scholar] [CrossRef] [PubMed]
- Isakson, M.; De Blacam, C.; Whelan, D.; McArdle, A.; Clover, A.J. Mesenchymal stem cells and cutaneous wound healing: Current evidence and future potential. Stem Cells Int. 2015, 2015, 831095. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Gang, X.; Sun, C.; Wang, G. Mesenchymal stem cells improve healing of diabetic foot ulcer. J. Diabetes Res. 2017, 2017, 9328347. [Google Scholar] [CrossRef] [PubMed]
- Mizukami, H.; Yagihashi, S. Exploring a new therapy for diabetic polyneuropathy–the application of stem cell transplantation. Front. Endocrinol. 2014, 5, 45. [Google Scholar] [CrossRef]
- Sasaki, M.; Abe, R.; Fujita, Y.; Ando, S.; Inokuma, D.; Shimizu, H. Mesenchymal stem cells are recruited into wounded skin and contribute to wound repair by transdifferentiation into multiple skin cell type. J. Immunol. 2008, 180, 2581–2587. [Google Scholar] [CrossRef] [PubMed]
- Golchin, A.; Seyedjafari, E.; Ardeshirylajimi, A. Mesenchymal stem cell therapy for COVID-19: Present or future. Stem Cell Rev. Rep. 2020, 16, 427–433. [Google Scholar] [CrossRef]
- Golchin, A.; Farahany, T.Z. Biological products: Cellular therapy and FDA approved products. Stem Cell Rev. Rep. 2019, 15, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Li, H.; Guo, Z. Mesenchymal stem cell-like properties in fibroblasts. Cell. Physiol. Biochem. 2014, 34, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Ankrum, J.A.; Ong, J.F.; Karp, J.M. Mesenchymal stem cells: Immune evasive, not immune privileged. Nat. Biotechnol. 2014, 32, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Katsha, A.M.; Ohkouchi, S.; Xin, H.; Kanehira, M.; Sun, R.; Nukiwa, T.; Saijo, Y. Paracrine factors of multipotent stromal cells ameliorate lung injury in an elastase-induced emphysema model. Mol. Ther. 2011, 19, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I. Mesenchymal stem cells. J. Orthop. Surg. Res. 1991, 9, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I.; Correa, D. The MSC: An injury drugstore. Cell Stem Cell 2011, 9, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Djouad, F.; Bouffi, C.; Ghannam, S.; Noël, D.; Jorgensen, C. Mesenchymal stem cells: Innovative therapeutic tools for rheumatic diseases. Nat. Rev. Rheumatol. 2009, 5, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Nakagami, H.; Morishita, R.; Maeda, K.; Kikuchi, Y.; Ogihara, T.; Kaneda, Y. Adipose tissue-derived stromal cells as a novel option for regenerative cell therapy. J. Atheroscler. Thromb. 2006, 13, 77–81. [Google Scholar] [CrossRef]
- Dominici, M.L.B.K.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The international society for cellular therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Kamihata, H.; Matsubara, H.; Nishiue, T.; Fujiyama, S.; Tsutsumi, Y.; Ozono, R.; Masaki, H.; Mori, Y.; Iba, O.; Tateishi, E.; et al. Implantation of bone marrow mononuclear cells into ischemic myocardium enhances collateral perfusion and regional function via side supply of angioblasts, angiogenic ligands, and cytokines. Circulation 2001, 104, 1046–1052. [Google Scholar] [CrossRef] [PubMed]
- Squillaro, T.; Peluso, G.; Galderisi, U. Clinical trials with mesenchymal stem cells: An update. Cell Transplant. 2016, 25, 829–848. [Google Scholar] [CrossRef]
- Crivelli, B.; Chlapanidas, T.; Perteghella, S.; Lucarelli, E.; Pascucci, L.; Brini, A.T.; Ferrero, I.; Marazzi, M.; Pessina, A.; Torre, M.L.; et al. Mesenchymal stem/stromal cell extracellular vesicles: From active principle to next generation drug delivery system. J. Control. Release 2017, 262, 104–117. [Google Scholar] [CrossRef]
- Park, W.S.; Ahn, S.Y.; Sung, S.I.; Ahn, J.Y.; Chang, Y.S. Strategies to enhance paracrine potency of transplanted mesenchymal stem cells in intractable neonatal disorders. Pediatr. Res. 2018, 83, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Lalu, M.M.; McIntyre, L.; Pugliese, C.; Stewart, D.J. Safety of cell therapy with mesenchymal stromal cells (MSCs): A systematic review. D49 Clin. Trials Crit. Care 2010, A6043. [Google Scholar]
- Marks, P.W.; Witten, C.M.; Califf, R.M. Clarifying stem-cell therapy’s benefits and risks. N. Engl. J. Med. 2017, 376, 1007–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cyranoski, D. Korean deaths spark inquiry: Cases highlight the challenge of policing multinational trade in stem-cell treatments. Nature 2010, 468, 485–486. [Google Scholar] [CrossRef]
- Jung, J.W.; Kwon, M.; Choi, J.C.; Shin, J.W.; Park, I.W.; Choi, B.W.; Kim, J.Y. Familial occurrence of pulmonary embolism after intravenous, adipose tissue-derived stem cell therapy. Yonsei Med. J. 2013, 54, 1293–1296. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, S.; Zhou, L.; Cai, J.; Tan, J.; Gao, X.; Zeng, Z.; Li, D. Thromboembolism induced by umbilical cord mesenchymal stem cell infusion: A report of two cases and literature review. Transplant. Proc. 2017, 49, 1656–1658. [Google Scholar] [CrossRef] [PubMed]
- George, M.J.; Prabhakara, K.; Toledano-Furman, N.E.; Wang, Y.W.; Gill, B.S.; Wade, C.E.; Olson, S.D.; Cox Jr, C.S. Clinical cellular therapeutics accelerate clot formation. Stem Cells Transl. Med. 2018, 7, 731–739. [Google Scholar] [CrossRef]
- Park, J.S.; Suryaprakash, S.; Lao, Y.H.; Leong, K.W. Engineering mesenchymal stem cells for regenerative medicine and drug delivery. Methods 2015, 84, 3–16. [Google Scholar] [CrossRef]
- Glennie, S.; Soeiro, I.; Dyson, P.J.; Lam, E.W.F.; Dazzi, F. Bone marrow mesenchymal stem cells induce division arrest anergy of activated T cells. Blood 2005, 105, 2821–2827. [Google Scholar] [CrossRef] [PubMed]
- Golchin, A.; Farahany, T.Z.; Khojasteh, A.; Soleimanifar, F.; Ardeshirylajimi, A. The clinical trials of mesenchymal stem cell therapy in skin diseases: An update and concise review. Curr. Stem Cell Res. Ther. 2019, 14, 22–33. [Google Scholar] [CrossRef]
- Ogawa, M.; LaRue, A.C.; Mehrotra, M. Hematopoietic stem cells are pluripotent and not just “hematopoietic”. Blood Cells Mol. Dis. 2013, 51, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.A. Hematopoietic stem cells. In Principles of Tissue Engineering; Academic Press: Cambridge, MA, USA, 2014; pp. 989–1040. [Google Scholar] [CrossRef]
- Delaney, C.; Gutman, J.A.; Appelbaum, F.R. Cord blood transplantation for haematological malignancies: Conditioning regimens, double cord transplant and infectious complications. Br. J. Haematol. 2009, 147, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Munoz, J.; Shah, N.; Rezvani, K.; Hosing, C.; Bollard, C.M.; Oran, B.; Olson, A.; Popat, U.; Molldrem, J.; McNiece, I.K.; et al. Concise review: Umbilical cord blood transplantation: Past, present, and future. Stem Cells Transl. Med. 2014, 3, 1435–1443. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ema, H. Mechanisms of self-renewal in hematopoietic stem cells. Int. J. Hematol. 2016, 103, 498–509. [Google Scholar] [CrossRef]
- Blanpain, C.; Horsley, V.; Fuchs, E. Epithelial stem cells: Turning over new leaves. Cell 2007, 128, 445–458. [Google Scholar] [CrossRef]
- Das, D.; Fletcher, R.B.; Ngai, J. Cellular mechanisms of epithelial stem cell self-renewal and differentiation during homeostasis and repair. WIREs Dev. Biol. 2020, 9, e361. [Google Scholar] [CrossRef]
- Beattie, R.; Hippenmeyer, S. Mechanisms of radial glia progenitor cell lineage progression. FEBS Lett. 2017, 591, 3993–4008. [Google Scholar] [CrossRef] [PubMed]
- Arvidsson, A.; Collin, T.; Kirik, D.; Kokaia, Z.; Lindvall, O. Neuronal replacement from endogenous precursors in the adult brain after stroke. Nat. Med. 2002, 8, 963–970. [Google Scholar] [CrossRef]
- Ilic, D.; Ogilvie, C. Concise review: Human embryonic stem cells—what have we done? What are we doing? Where are we going? Stem Cells 2017, 35, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Amit, M.; Itskovitz-Eldor, J. Embryonic stem cells: Isolation, characterization and culture. In Engineering of Stem Cells; Springer: Berlin/Heidelberg, Germany, 2009; pp. 173–184. [Google Scholar] [CrossRef]
- Tseng, A.M.; Mahnke, A.H.; Salem, N.A.; Miranda, R.C. Noncoding RNA regulatory networks, epigenetics, and programming stem cell renewal and differentiation: Implications for stem cell therapy. In Epigenetics in Human Disease; Academic Press: Cambridge, MA, USA, 2018; pp. 903–933. [Google Scholar] [CrossRef]
- Prochazkova, M.; Chavez, M.G.; Prochazka, J.; Felfy, H.; Mushegyan, V.; Klein, O.D. Embryonic versus adult stem cells. In Stem Cell Biology and Tissue Engineering in Dental Sciences; Academic Press: Cambridge, MA, USA, 2015; pp. 249–262. [Google Scholar] [CrossRef]
- Kmiecik, G.; Niklińska, W.; Kuć, P.; Pancewicz-Wojtkiewicz, J.; Fil, D.; Karwowska, A.; Karczewski, J.; Mackiewicz, Z. Fetal membranes as a source of stem cells. Adv. Med. Sci. 2013, 58, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Rippon, H.J.; Bishop, A.E. Embryonic stem cells. Cell Prolif. 2004, 37, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Gnecchi, M.; Zhang, Z.; Ni, A.; Dzau, V.J. Paracrine mechanisms in adult stem cell signaling and therapy. Circ. Res. 2008, 103, 1204–1219. [Google Scholar] [CrossRef] [PubMed]
- Hirschi, K.K.; Li, S.; Roy, K. Induced pluripotent stem cells for regenerative medicine. Annu. Rev. Biomed. Eng. 2014, 16, 277. [Google Scholar] [CrossRef] [PubMed]
- Hosoya, M.; Czysz, K. Translational prospects and challenges in human induced pluripotent stem cell research in drug discovery. Cells 2016, 5, 46. [Google Scholar] [CrossRef] [PubMed]
- Taura, D.; Sone, M.; Homma, K.; Oyamada, N.; Takahashi, K.; Tamura, N.; Yamanaka, S.; Nakao, K. Induction and isolation of vascular cells from human induced pluripotent stem cells—Brief report. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1100–1103. [Google Scholar] [CrossRef]
- Kumar, D.; Talluri, T.R.; Anand, T.; Kues, W.A. Induced pluripotent stem cells: Mechanisms, achievements and perspectives in farm animals. World J. Stem Cells 2015, 7, 315. [Google Scholar] [CrossRef] [PubMed]
- Gnecchi, M.; Melo, L.G. Bone marrow-derived mesenchymal stem cells: Isolation, expansion, characterization, viral transduction, and production of conditioned medium. Methods Mol. Biol. 2009, 482, 281–294. [Google Scholar] [CrossRef] [PubMed]
- WHO. 2022. Available online: https://www.who.int/news/item/22-02-2022-statement-on-omicron-sublineage-ba.2 (accessed on 11 April 2022).
- Lucas, C.; Wong, P.; Klein, J.; Castro, T.B.; Silva, J.; Sundaram, M.; Ellingson, M.K.; Mao, T.; Oh, J.E.; Israelow, B.; et al. Longitudinal analyses reveal immunological misfiring in severe COVID-19. Nature 2020, 584, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Maleki, M.; Ghanbarvand, F.; Behvarz, M.R.; Ejtemaei, M.; Ghadirkhomi, E. Comparison of mesenchymal stem cell markers in multiple human adult stem cells. Int. J. Stem Cells 2014, 7, 118–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, F.J.; Tuan, R.S.; Cheung, K.M.; Leung, V.Y. Concise review: The surface markers and identity of human mesenchymal stem cells. Stem Cells 2014, 32, 1408–1419. [Google Scholar] [CrossRef] [PubMed]
- Weiss, A.R.R.; Dahlke, M.H. Immunomodulation by mesenchymal stem cells (MSCs): Mechanisms of action of living, apoptotic, and dead MSCs. Front. Immunol. 2019, 10, 1191. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Xia, Z.F. Paracrine activity of stem cells in therapy for acute lung injury and adult respiratory distress syndrome. J. Trauma Acute Care Surg. 2013, 74, 1351–1356. [Google Scholar] [CrossRef] [PubMed]
- Ottaviano, G.; Chiesa, R.; Feuchtinger, T.; Vickers, M.A.; Dickinson, A.; Gennery, A.R.; Veys, P.; Todryk, S. Adoptive T cell therapy strategies for viral infections in patients receiving haematopoietic stem cell transplantation. Cells 2019, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.X.; Sun, Y.Q.; Shi, J.; Li, C.L.; Fang, S.B.; Wang, D.; Deng, X.Q.; Wen, W.; Fu, Q.L. Effects of mesenchymal stem cells from human induced pluripotent stem cells on differentiation, maturation, and function of dendritic cells. Stem Cell Res. Ther. 2017, 8, 48. [Google Scholar] [CrossRef] [PubMed]
- Romano, B.; Elangovan, S.; Erreni, M.; Sala, E.; Petti, L.; Kunderfranco, P.; Massimino, L.; Restelli, S.; Sinha, S.; Lucchetti, D.; et al. TNF-stimulated Gene-6 is a key regulator in switching stemness and biological properties of mesenchymal stem cells. Stem Cells 2019, 37, 973–987. [Google Scholar] [CrossRef] [PubMed]
- Kebria, M.M.; Milan, P.B.; Peyravian, N.; Kiani, J.; Khatibi, S.; Mozafari, M. Stem cell therapy for COVID-19 pneumonia. Mol. Biomed. 2022, 3, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Cogle, C.R.; Guthrie, S.M.; Sanders, R.C.; Allen, W.L.; Scott, E.W.; Petersen, B.E. An overview of stem cell research and regulatory issues. Mayo Clin. Proc. 2003, 78, 993–1003. [Google Scholar] [CrossRef]
- Wu, X.; Thi, V.L.D.; Huang, Y.; Billerbeck, E.; Saha, D.; Hoffmann, H.H.; Wang, Y.; Silva, L.A.V.; Sarbanes, S.; Sun, T.; et al. Intrinsic immunity shapes viral resistance of stem cells. Cell 2018, 172, 423–438. [Google Scholar] [CrossRef] [PubMed]
- Van Den Akker, F.; Deddens, J.C.; Doevendans, P.A.; Sluijter, J.P.G. Cardiac stem cell therapy to modulate inflammation upon myocardial infarction. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2013, 1830, 2449–2458. [Google Scholar] [CrossRef]
- Liang, B.; Chen, J.; Li, T.; Wu, H.; Yang, W.; Li, Y.; Li, J.; Yu, C.; Nie, F.; Ma, Z.; et al. Clinical remission of a critically ill COVID-19 patient treated by human umbilical cord mesenchymal stem cells: A case report. Medicine 2020, 99, e21429. [Google Scholar] [CrossRef]
- Song, N.; Wakimoto, H.; Rossignoli, F.; Bhere, D.; Ciccocioppo, R.; Chen, K.S.; Khalsa, J.K.; Mastrolia, I.; Samarelli, A.V.; Dominici, M.; et al. Mesenchymal stem cell immunomodulation: In pursuit of controlling COVID-19 related cytokine storm. Stem Cells 2021, 39, 707–722. [Google Scholar] [CrossRef]
- Tropea, K.A.; Leder, E.; Aslam, M.; Lau, A.N.; Raiser, D.M.; Lee, J.H.; Balasubramaniam, V.; Fredenburgh, L.E.; Alex Mitsialis, S.; Kourembanas, S.; et al. Bronchioalveolar stem cells increase after mesenchymal stromal cell treatment in a mouse model of bronchopulmonary dysplasia. Am. J. Physiol. -Lung Cell. Mol. 2012, 302, L829–L837. [Google Scholar] [CrossRef] [PubMed]
- Bari, E.; Ferrarotti, I.; Saracino, L.; Perteghella, S.; Torre, M.L.; Corsico, A.G. Mesenchymal stromal cell secretome for severe COVID-19 infections: Premises for the therapeutic use. Cells 2020, 9, 924. [Google Scholar] [CrossRef]
- Yagi, H.; Soto-Gutierrez, A.; Parekkadan, B.; Kitagawa, Y.; Tompkins, R.G.; Kobayashi, N.; Yarmush, M.L. Mesenchymal stem cells: Mechanisms of immunomodulation and homing. Cell Transplant. 2010, 19, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Pittenger, M.F. Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood 2005, 105, 1815–1822. [Google Scholar] [CrossRef] [PubMed]
- Dufrane, D. Impact of age on human adipose stem cells for bone tissue engineering. Cell Transplant. 2017, 26, 1496–1504. [Google Scholar] [CrossRef]
- Liu, M.; Lei, H.; Dong, P.; Fu, X.; Yang, Z.; Yang, Y.; Ma, J.; Liu, X.; Cao, Y.; Xiao, R. Adipose-derived mesenchymal stem cells from the elderly exhibit decreased migration and differentiation abilities with senescent properties. Cell Transplant. 2017, 26, 1505–1519. [Google Scholar] [CrossRef] [PubMed]
- Kokai, L.E.; Traktuev, D.O.; Zhang, L.; Merfeld-Clauss, S.; DiBernardo, G.; Lu, H.; Marra, K.G.; Donnenberg, A.; Donnenberg, V.; Meyer, E.M.; et al. Adipose stem cell function maintained with age: An intra-subject study of long-term cryopreserved cells. Aesthetic Surg. J. 2017, 37, 454–463. [Google Scholar] [CrossRef]
- Pachón-Peña, G.; Serena, C.; Ejarque, M.; Petriz, J.; Duran, X.; Oliva-Olivera, W.; Simó, R.; Tinahones, F.J.; Fernández-Veledo, S.; Vendrell, J. Obesity determines the immunophenotypic profile and functional characteristics of human mesenchymal stem cells from adipose tissue. Stem Cells Transl. Med. 2016, 5, 464–475. [Google Scholar] [CrossRef] [PubMed]
- Price, M.J.; Chou, C.C.; Frantzen, M.; Miyamoto, T.; Kar, S.; Lee, S.; Shah, P.K.; Martin, B.J.; Lill, M.; Forrester, J.S.; et al. Intravenous mesenchymal stem cell therapy early after reperfused acute myocardial infarction improves left ventricular function and alters electrophysiologic properties. Int. J. Cardiol. 2006, 111, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Rubenfeld, G.D.; Caldwell, E.; Peabody, E.; Weaver, J.; Martin, D.P.; Neff, M.; Stern, E.J.; Hudson, L.D. Incidence and outcomes of acute lung injury. N. Engl. J. Med. 2005, 353, 1685–1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
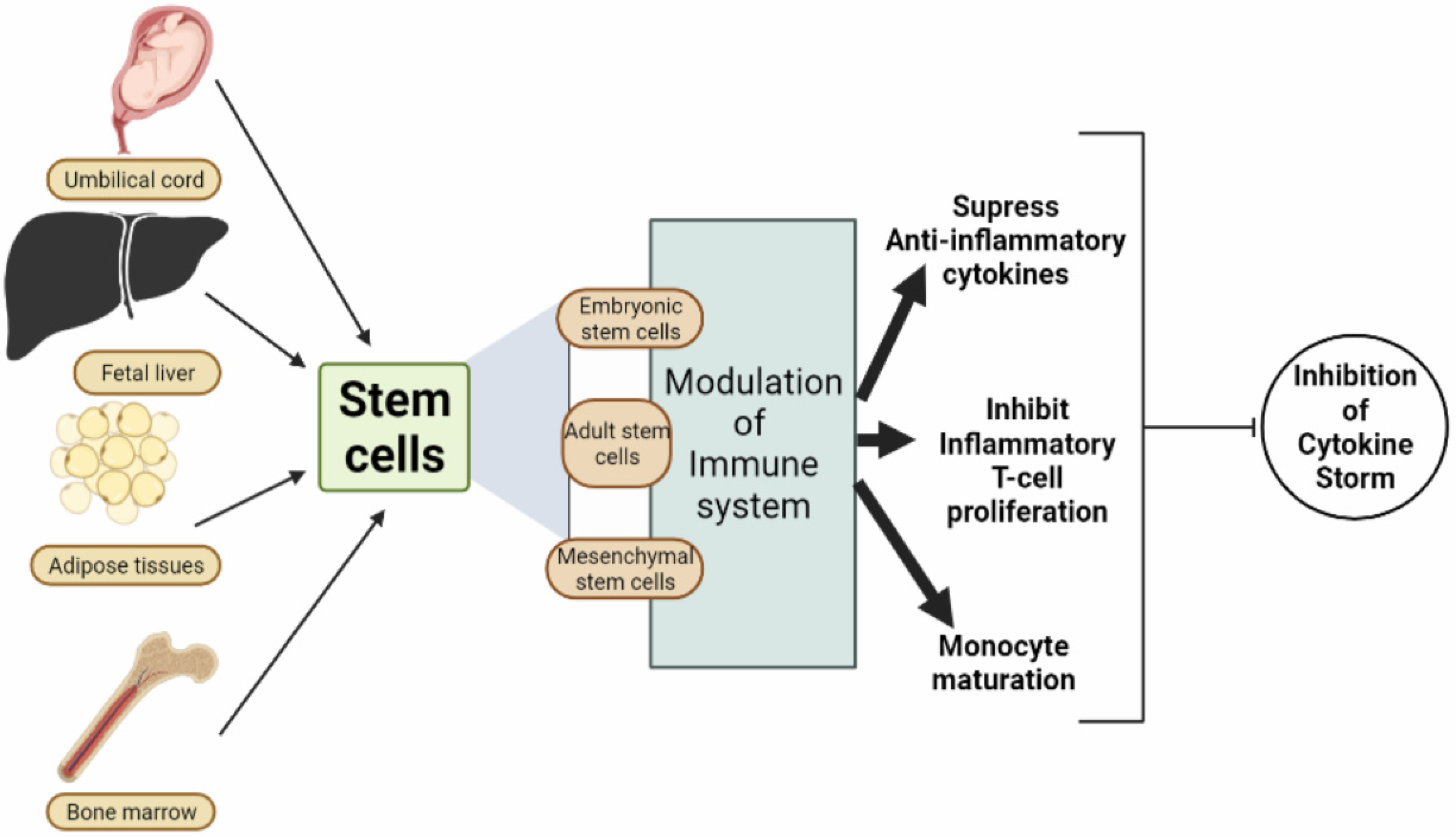
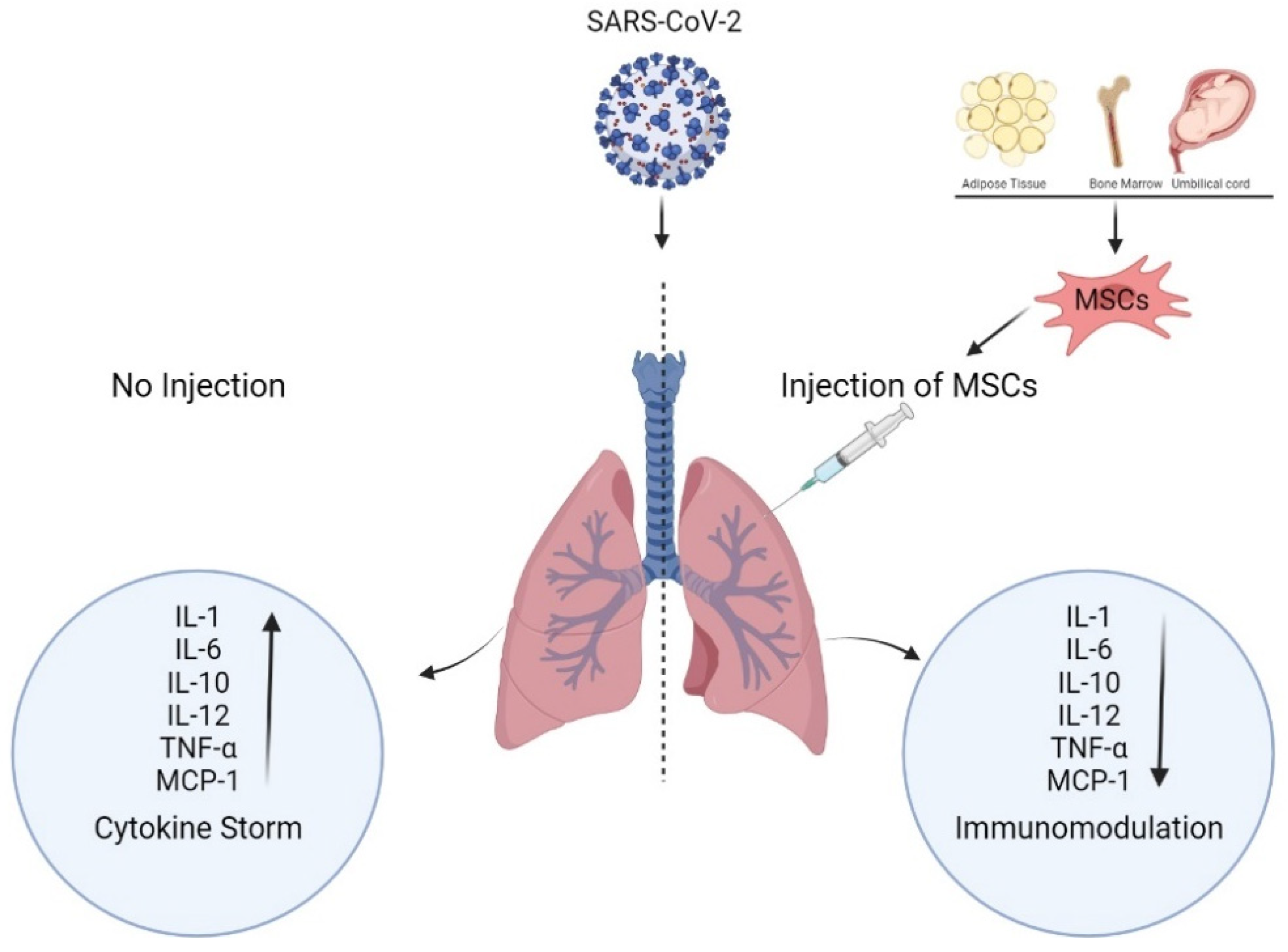

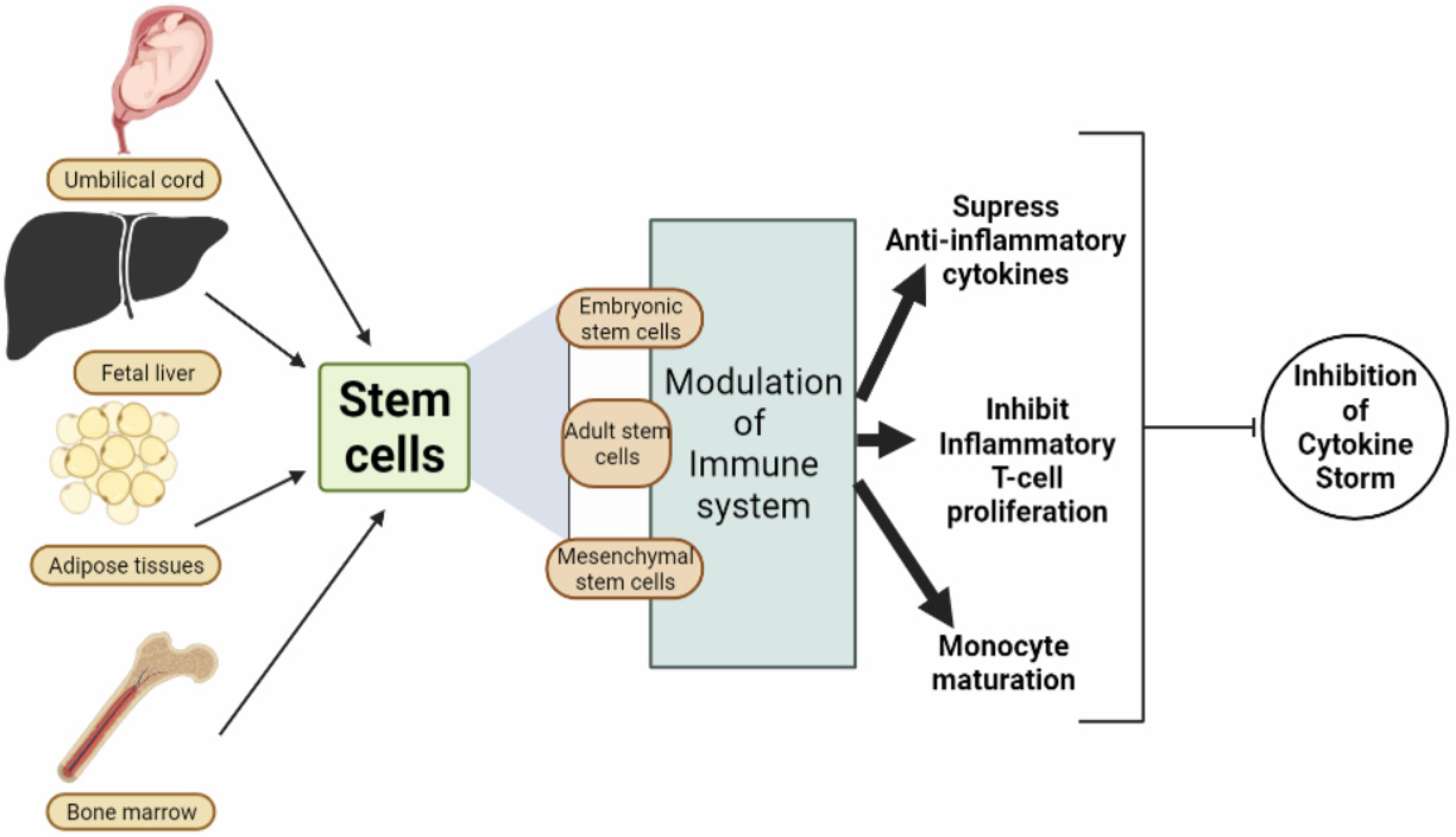{kind=link}
{kind=link}
| Family | Cytokine, Pro-Inflammatory Factor | Secreted by | Types of Cells on Which It Acts/Function | Mode of Action/Mechanism | References |
|---|---|---|---|---|---|
| Cytokine | GSCF (Granulocyte Colony-Stimulating Factor) | Endothelium, macrophages | Mouse lymphoid-biased | Anti-apoptotic, angiogenic, neurogenesis and functions. | [26,27,28,29] |
| Cytokine | IP10 | Monocytes, T-cells, endothelial cells, and keratinocytes | It recruits immune cells to fight at inflammatory sites | To stimulate apoptosis, chemotaxis, cell growth, and angiostasis | [26,30,31,32] |
| Chemokines | MCP1 (Monocyte Chemoattractant Protein 1) | Microglial cells, mesangial, epithelial, smooth muscle, astrocytic, monocytic, and endothelial | Attracts T- lymphocytes, monocytes, and natural killer cells | It infiltrates, facilitates the migration of inflammatory cells and other cytokines towards the site of Inflammation. | [26,33,34,35] |
| Chemokines | MIP1A (Macrophage Inflammatory Protein 1 α) | Monocytes and macrophages | Act upon inflammatory cells and maintain impulsive immune response. | Healing wounded cells and halting stem cells. | [36,37] |
| Cytokine | IL-2 | CD4+ T cells | Act against microbial infection as a natural impedance. It also promotes T cells differentiation into an effector T cell and then into memory T cell as the incident with antigen. | Ameliorate AICD (Activation Induced Cell Death) and increase the killing activity of Tc (Cytotoxic T) cells and NK cells. | [38,39,40] |
| Cytokine | IL-6 | Dendritic cell and macrophages | Inflamed acute-phase protein synthesis, neutrophile in bone marrow, and help in the growth of B-cells. | IFN-γ secretion is affected by IL-6 through CD4 T cells, i.e., curial interferon that uplifts, IL-6 triggers CD4 cells to release IL-4 and directly affects Th2. | [41,42,43] |
| Cytokine | IL-7 | Stromal cells in thymus and bone marrow | It affects mature T-cells and immature B-cells and leads to secondary cytokine release. | It involves mechanically on TCR-gamma and TCR-gamma delta thymocyte maturation. | [44,45] |
| Cytokine, superfamilyTNF | TNF-α (Tumor necrosis factor α) | Macrophages/monocytes | Perform miscellaneous functions within the cells during acute inflammation, and it activates and proliferates naïve and effector T cells. | Diverse signaling pathways lead to necrosis or apoptosis. | [46,47,48] |
| Chemokine (CXC Family) | IL-8 | Mainly by macrophages /monocytes and some other cell types like epithelial cells, endothelial cells, smooth muscle cells, and airways | It has a direct effect on immune cells and polymorphonuclear cells. | IL-8 is considered a prognostic and therapeutic factor for wound healing. | [49,50,51] |
| Eicosanoid inflammatory mediators | Leukotriene (LT) | Mast cells | Create inflammatory cascade, effect on leukocytes, and stenosis of smooth muscles. | Their mode of action depends on the effective binding with G-protein-coupled receptors, and every LT receptor has an abnormal expression pattern and function. | [52,53,54,55] |
| Cytokine | IL-1β | Dendritic cell, activated macrophages | Pro-inflammatory cytokine and held in inflammation, autoimmune conditions, and pain. | IL-1β binds to the IL-1 type 1 receptor (IL-1R1), leads to the illustration of inflammation, and has the potency to induce fever when delivered exogenously. | [56,57,58] |
| Cytokine | IL-12 | Dendritic cells | IL-12 receptors are present on T cells and NK cells, stimulating TH1 and NK cell growth while inhibiting TH2 cell responses. | This molecule produces interferon (IFN-γ), encourages the differentiation of T helper 1 (TH1) cells, and provides a link between innate defenses and adaptive defenses. | [59,60,61] |
| Cytokine | IL-33 | Cellular damage area of bronchial epithelial cells, airway, endothelial cells of high endothelial venules | Generally, mast cells become degranulated when exposed to IL-33, and the effect also occurs in basophils and granulocytes. | It enhances Th2 responses. | [62,63,64,65] |
| (TGF-β) family | TGF-β | Monocytes/macrophages, lymphocytes and platelets | In addition to interacting with the surrounding cells, this TGF-β acts on smooth muscle cells, immune cells, and endothelial cells. | The condition causes angiogenesis and immunosuppression, which makes cancer more aggressive. | [66,67] |
| CC Family Chemokine Scavenger Receptor | CXCL-10 | Dendritic cell and macrophages | This protein controls the differentiation of naive T cells into T helper 1 (Th1) cells and mediates immune cell migration to the foci. | This CXCL-10 chemokine binds to the CXCR-3 receptor to produce its effects in the cell. | [56,68,69,70] |
| Signaling proteins | IF | Natural killer (NK) cells, activated T cells, dendritic cells and macrophages. | Several cells, including monocytes, macrophages, T-lymphocytes, glia, and neurons, have IFN receptors. | When IFN-γ is produced, its effects are antiviral, antimicrobial, antitumor, and immunomodulatory. IFN proteins beta, alpha, and gamma are what produce those effects. | [56,71,72,73] |
| Cytokine | IL-18 | Monocyte/macrophage | IL-18 activates th1 cells, and CD8+ T and natural killer (NK) cells are enhanced by it. | It increases the cytotoxic activity of CD8+ T cells and NK cells by upregulation of FasL. | [74,75] |
| S. No. | Stem Cell | Type of Cell | Isolated from Which Portion | Mode of Action | References |
|---|---|---|---|---|---|
| 1. | Mesenchymal Stem Cell (MSC) | Multipotent stem cells | Fetal liver, bone marrow, umbilical cord, menstrual blood, dental pulp, adipose tissues, etc. | They perform an endogenous repair of stem cells and prevent the excessive release of cytokines from the immune system. | [122] |
| 2. | Hematopoietic Stem Cells (HSCs) | HSCs are pluripotent and have ambient self-renewal efficiency. | HSCs are predominantly found in the bone marrow region, sternum, femur portion, umbilical cord, and even in a few segments of peripheral blood. | Regulated in two forms of mechanism. The first mechanism says they control the G0 phase, and in another mechanism it is fate determination, i.e., either differentiation or self-renew) | [123,124,125,126,127] |
| 3. | Epithelial Stem Cells (ESCs) | ESCs are multipotent stem cells due to self-renewal capability throughout the life and/or unipotent progenitor cells. | They were isolated from the different layers of skin, i.e., from ectoderm, mesoderm, and endoderm. | In its action, various cellular-signaling mechanisms take parts, such as bone morphogenetic protein, WNT, and Sonic Hedgehog, which play a prominent part. These signaling pathways govern the conserved mechanisms behind the self-renewal capability of adult epithelial structures. | [128,129] |
| 4. | Neural Stem Cells (NSCs) | They are self-renewal and multipotent stem cells, | In the adult mammalian brain, the sub-granular zone and subventricular zone have the reservoir of NSCs. | The formation of new hippocampal NSCs and its cellular mechanism taking part in it, along with a decrease in neurogenic potential is still unclear and therapeutic cargoes exchange in horizontal to host cell through extracellular vesicles is also not fully understood. | [130,131] |
| 5. | Embryonic Stem Cells (ESCs) | The ESCs or human embryonic stem cells (hESC) possess tremendous pluripotent property and an extraordinary proliferative and growth capacity. | These ESCs are isolated from the mammalian blastocyst. | The ESCs mechanism of action depends on transcription factors associated with four genes viz., Sox2, Oct4, Tcf3, and Nanog that maintain pluripotency. | [132,133,134] |
| 6. | Adult Stem Cells (ASCs) | These are multipotent, undifferentiated cells that renew themselves and preclude them into specialized cell types. | ASCs can be isolated from blood, bone marrow, skin, adipose tissue, and liver. | Due to environmental stimuli, ASCs release biologically active compounds that lead to exerting paracrine action on different neighboring cells and hence leading to repair, tissue protection, regeneration, self-renewal, and proliferation taking place. | [135,136,137,138] |
| 7. | Induced Pluripotent Stem Cells (iPSCs) | These are (iPSCs) genetically engineered from somatic cells and pluripotent. | These are isolated from human adult somatic cells. | The remarkable feature of iPSCs to differentiate it into required specialized cell types and this property provides a source for innovative cell therapies with unlimited cell sources. | [139,140,141,142] |
| 8. | Umbilical cord-derived MSCs | They are multipotent stem cells. | Isolated from the human embryo. | The mechanism of action (MOA) is still unknown | [143] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Upadhyay, T.K.; Trivedi, R.; Khan, F.; Pandey, P.; Sharangi, A.B.; Goel, H.; Saeed, M.; Park, M.N.; Kim, B. Potential Therapeutic Role of Mesenchymal-Derived Stem Cells as an Alternative Therapy to Combat COVID-19 through Cytokines Storm. Cells 2022, 11, 2686. https://doi.org/10.3390/cells11172686
Upadhyay TK, Trivedi R, Khan F, Pandey P, Sharangi AB, Goel H, Saeed M, Park MN, Kim B. Potential Therapeutic Role of Mesenchymal-Derived Stem Cells as an Alternative Therapy to Combat COVID-19 through Cytokines Storm. Cells. 2022; 11(17):2686. https://doi.org/10.3390/cells11172686
Chicago/Turabian StyleUpadhyay, Tarun Kumar, Rashmi Trivedi, Fahad Khan, Pratibha Pandey, Amit Baran Sharangi, Harsh Goel, Mohd Saeed, Moon Nyeo Park, and Bonglee Kim. 2022. "Potential Therapeutic Role of Mesenchymal-Derived Stem Cells as an Alternative Therapy to Combat COVID-19 through Cytokines Storm" Cells 11, no. 17: 2686. https://doi.org/10.3390/cells11172686
APA StyleUpadhyay, T. K., Trivedi, R., Khan, F., Pandey, P., Sharangi, A. B., Goel, H., Saeed, M., Park, M. N., & Kim, B. (2022). Potential Therapeutic Role of Mesenchymal-Derived Stem Cells as an Alternative Therapy to Combat COVID-19 through Cytokines Storm. Cells, 11(17), 2686. https://doi.org/10.3390/cells11172686