
lncRNA2919 Suppresses Rabbit Dermal Papilla Cell Proliferation via trans-Regulatory Actions


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Cell Culture
2.3. Rapid Amplification of cDNA Ends and Bioinformatics Analysis
2.4. Plasmid Preparation, Small Interfering RNA, and Short Hairpin RNA
2.5. RNA Isolation and Quantitative RT-PCR Analysis
2.6. Western Blotting System (Protein Simple Wes)
2.7. Cell Proliferation and Apoptosis Assays
2.8. Dual-Luciferase Assay Verification of the Promoter Region
2.9. Electrophoretic Mobility Shift Assay
2.10. RNA Pull-Down and RNA-Binding Protein Immunoprecipitation Assays
2.11. Statistical Analysis
3. Results
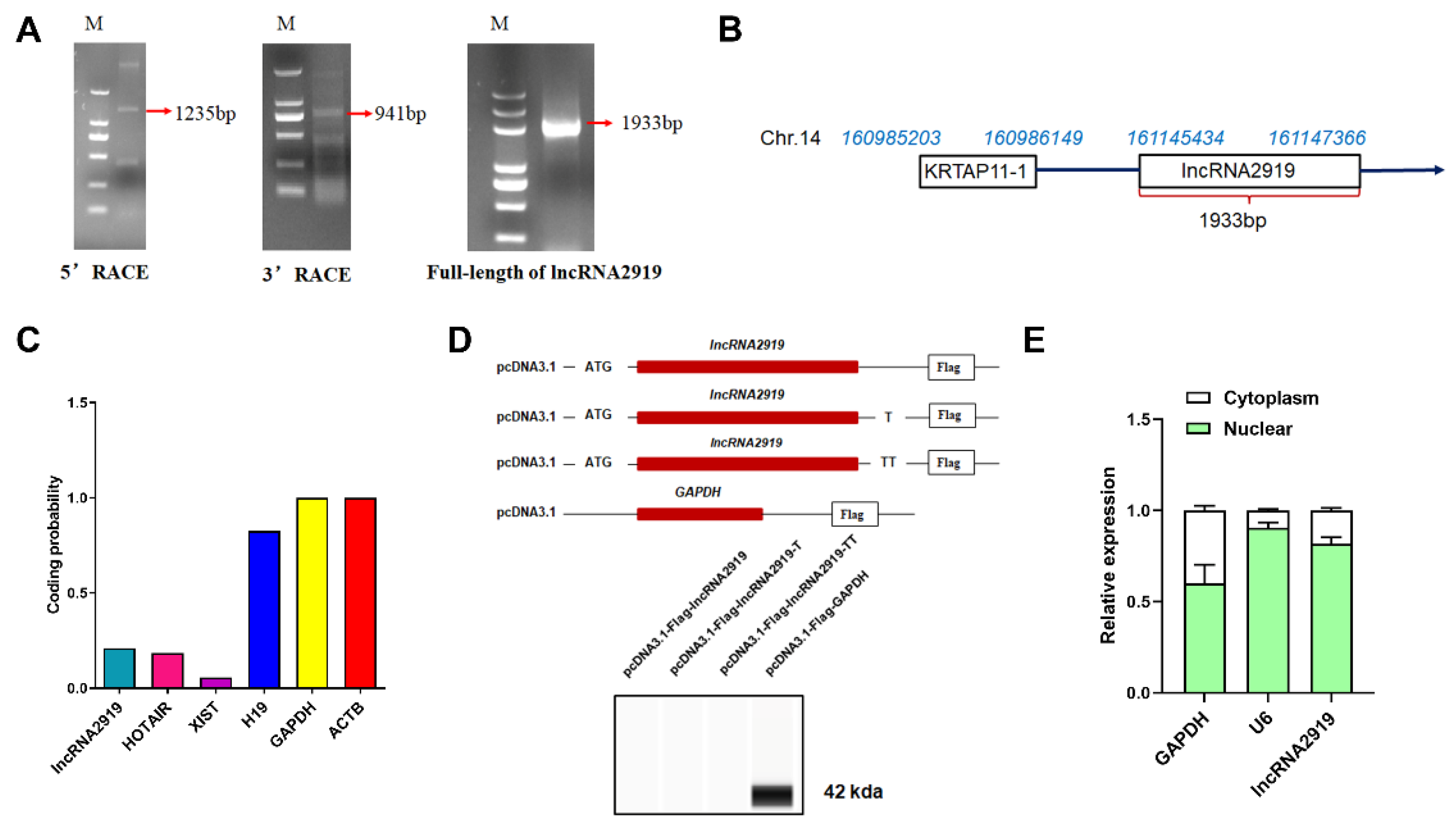
3.1. Identification and Characterization of lncRNA2919
3.2. lncRNA2919 Plays a Negative Role in DPC Proliferation
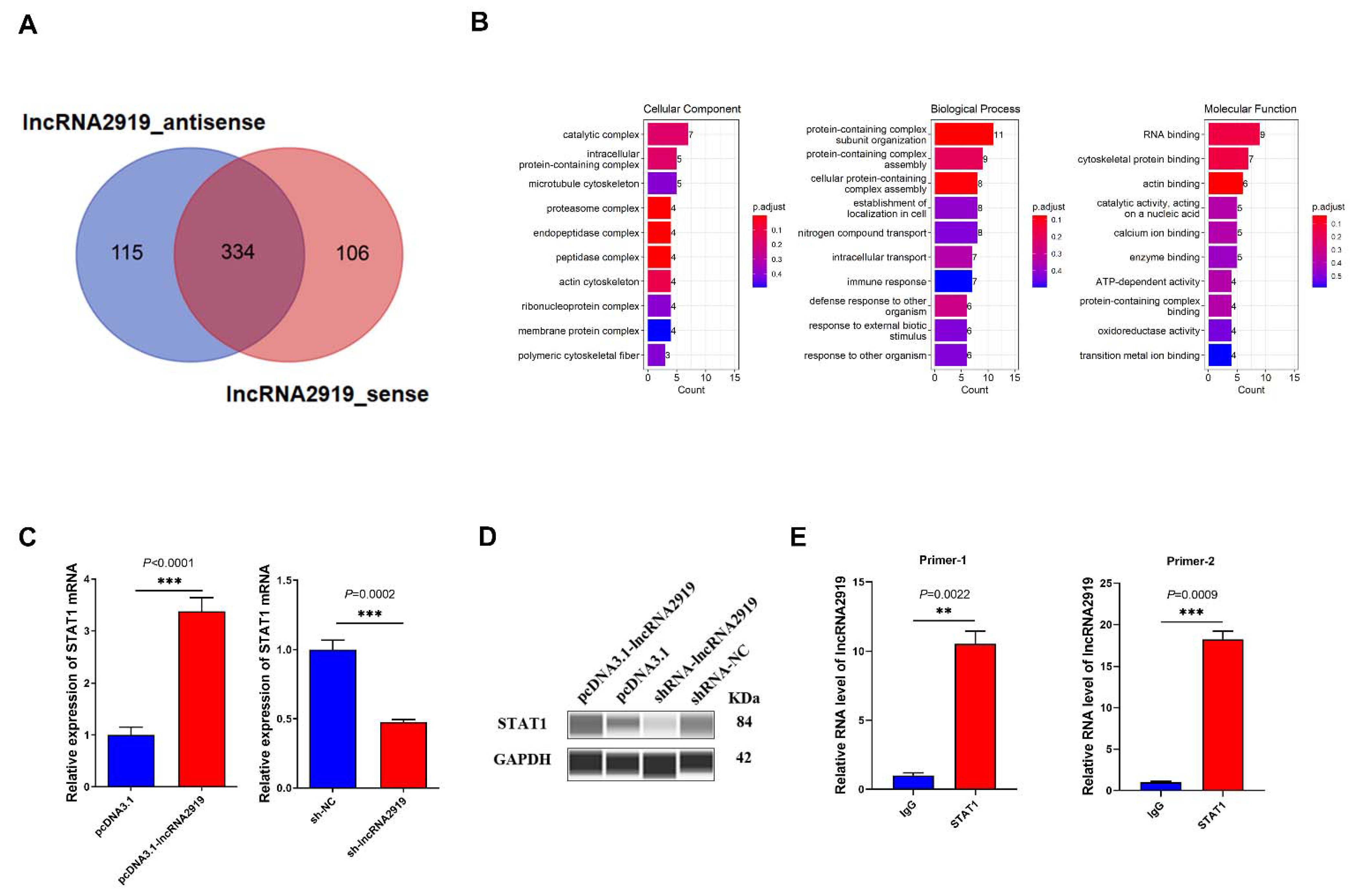
3.3. lncRNA2919 Recruits STAT1 Protein
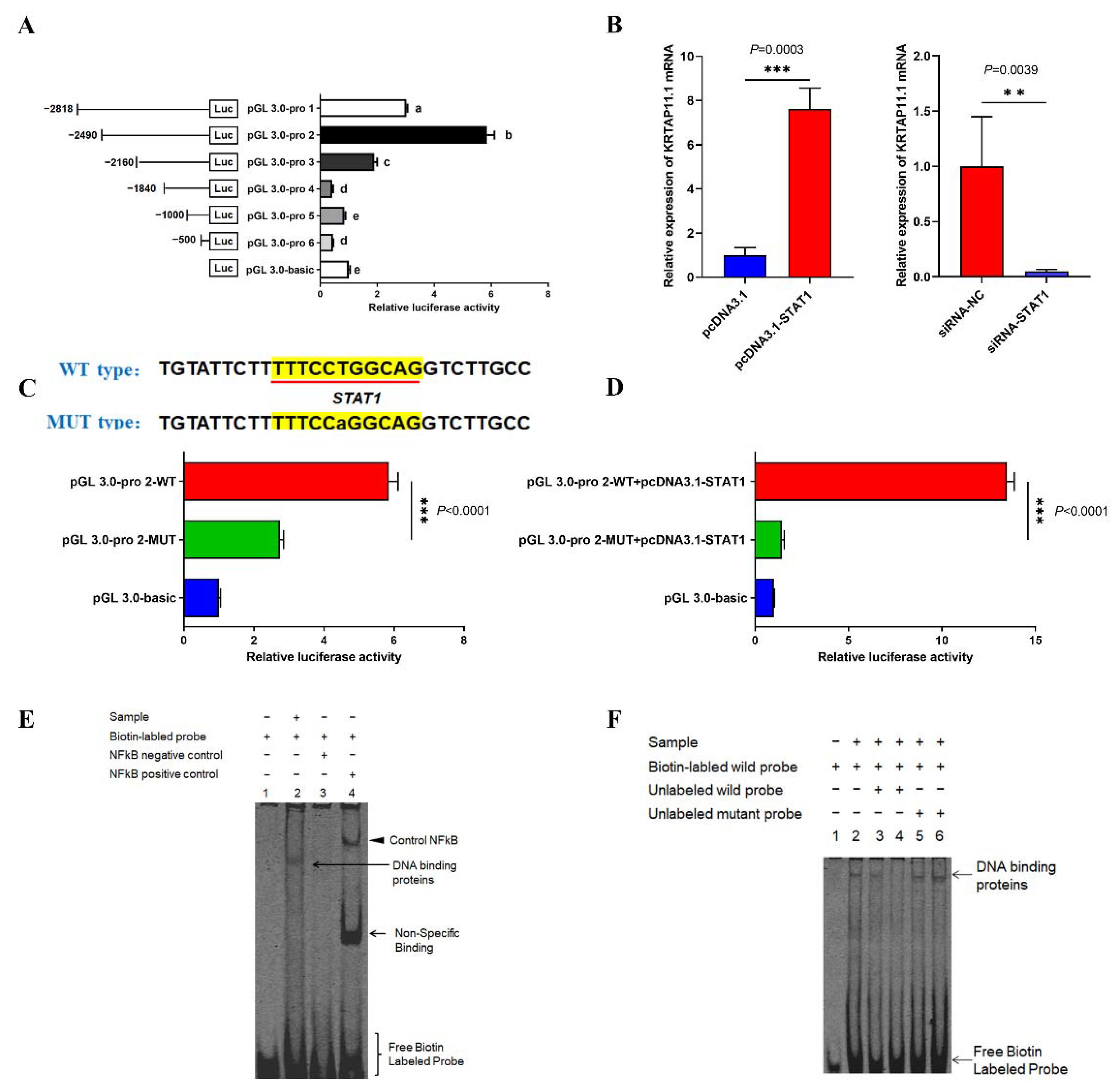
3.4. lncRNA2919 Regulates HF Cyclic Regeneration by the trans-Regulatory Axis of lncRNA2919–STAT1–KRTAP11-1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alonso, L.; Fuchs, E. The hair cycle. J. Cell Sci. 2006, 119, 391–393. [Google Scholar] [CrossRef] [PubMed]
- Hardy, M.H. The secret life of the hair follicle. Trends Genet. 1992, 8, 55–61. [Google Scholar] [CrossRef]
- Oro, A.E.; Scott, M.P. Splitting hairs: Dissecting roles of signaling systems in epidermal development. Cell 1998, 95, 575–578. [Google Scholar] [CrossRef]
- Oshima, H.; Rochat, A.; Kedzia, C.; Kobayashi, K.; Barrandon, Y. Morphogenesis and renewal of hair follicles from adult multipotent stem cells. Cell 2001, 104, 233–245. [Google Scholar] [CrossRef]
- Paus, R.; Cotsarelis, G. The biology of hair follicles. N. Engl. J. Med. 1999, 341, 491–497. [Google Scholar] [CrossRef]
- Stenn, K.; Paus, R. Controls of hair follicle cycling. Physiol. Rev. 2001, 81, 449–494. [Google Scholar] [CrossRef]
- Guttman, M.; Rinn, J.L. Modular regulatory principles of large non-coding rnas. Nature 2012, 482, 339–346. [Google Scholar] [CrossRef]
- Engreitz, J.M.; Haines, J.E.; Perez, E.M.; Munson, G.; Chen, J.; Kane, M.; McDonel, P.E.; Guttman, M.; Lander, E.S. Local regulation of gene expression by lncrna promoters, transcription and splicing. Nature 2016, 539, 452–455. [Google Scholar] [CrossRef]
- Ferre, F.; Colantoni, A.; Helmer-Citterich, M. Revealing protein–lncrna interaction. Brief. Bioinform. 2016, 17, 106–116. [Google Scholar] [CrossRef]
- Kopp, F.; Mendell, J.T. Functional classification and experimental dissection of long noncoding rnas. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef]
- Mathieu, E.-L.; Belhocine, M.; Dao, L.; Puthier, D.; Spicuglia, S. Functions of lncrna in development and diseases. Med. Sci. M/S 2014, 30, 790–796. [Google Scholar]
- Quan, J.; Pan, X.; Zhao, L.; Li, Z.; Dai, K.; Yan, F.; Liu, S.; Ma, H.; Lai, Y. Lncrna as a diagnostic and prognostic biomarker in bladder cancer: A systematic review and meta-analysis. OncoTargets Ther. 2018, 11, 6415. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Liu, Y.; Huang, K.; Chen, X.; Cai, B.; Li, H.; Yuan, Y.; Zhang, H.; Li, Y. Long noncoding rna expression in dermal papilla cells contributes to hairy gene regulation. Biochem. Biophys. Res. Commun. 2014, 453, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Si, Y.; Bai, J.; Wu, J.; Li, Q.; Mo, Y.; Fang, R.; Lai, W. Lncrna plncrna-1 regulates proliferation and differentiation of hair follicle stem cells through tgf-β1-mediated wnt/β-catenin signal pathway. Mol. Med. Rep. 2018, 17, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Zheng, Y.; Ma, S.; Xing, Q.; Wang, X.; Yang, B.; Yin, G.; Guan, F. Long non-coding rna regulates hair follicle stem cell proliferation and differentiation through pi3k/akt signal pathway. Mol. Med. Rep. 2018, 17, 5477–5483. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Q.; Yin, R.H.; Zhao, S.J.; Wang, Z.Y.; Zhu, Y.B.; Wang, W.; Zheng, Y.Y.; Yin, X.B.; Guo, D.; Wang, S.Q. Identification and molecular analysis of a lncrna-hotair transcript from secondary hair follicle of cashmere goat reveal integrated regulatory network with the expression regulated potentially by its promoter methylation. Gene 2019, 688, 182–192. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, Z.; Zhu, Y.; Wang, W.; Bai, M.; Jiao, Q.; Wang, Y.; Zhao, S.; Yin, X.; Guo, D. Lncrna-000133 from secondary hair follicle of cashmere goat: Identification, regulatory network and its effects on inductive property of dermal papilla cells. Anim. Biotechnol. 2020, 31, 122–134. [Google Scholar] [CrossRef]
- Wang, S.; Ge, W.; Luo, Z.; Guo, Y.; Jiao, B.; Qu, L.; Zhang, Z.; Wang, X. Integrated analysis of coding genes and non-coding rnas during hair follicle cycle of cashmere goat (Capra hircus). BMC Genom. 2017, 18, 767. [Google Scholar] [CrossRef]
- Zhou, G.; Kang, D.; Ma, S.; Wang, X.; Gao, Y.; Yang, Y.; Wang, X.; Chen, Y. Integrative analysis reveals ncrna-mediated molecular regulatory network driving secondary hair follicle regression in cashmere goats. BMC Genom. 2018, 19, 222. [Google Scholar] [CrossRef]
- Zhao, B.; Chen, Y.; Hu, S.; Yang, N.; Wang, M.; Liu, M.; Li, J.; Xiao, Y.; Wu, X. Systematic analysis of non-coding rnas involved in the angora rabbit (oryctolagus cuniculus) hair follicle cycle by rna sequencing. Front. Genet. 2019, 10, 407. [Google Scholar] [CrossRef]
- Huang, M.; Zhong, Z.; Lv, M.; Shu, J.; Tian, Q.; Chen, J. Comprehensive analysis of differentially expressed profiles of lncrnas and circrnas with associated co-expression and cerna networks in bladder carcinoma. Oncotarget 2016, 7, 47186. [Google Scholar] [CrossRef] [PubMed]
- Luo, R.; Song, J.; Zhang, W.; Ran, L. Identification of mfi2-as1, a novel pivotal lncrna for prognosis of stage iii/iv colorectal cancer. Dig. Dis. Sci. 2020, 65, 3538–3550. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, T.; Tang, H.; Sha, Z.; Chen, R.; Chen, L.; Yu, Y.; Rowe, G.C.; Das, S.; Xiao, J. Inhibition of lncrna maat controls multiple types of muscle atrophy by cis- and trans-regulatory actions. Mol. Ther. 2021, 29, 1102–1119. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, S.; Takase, T.; Kadono, N.; Maekubo, K.; Hirai, Y. Krtap11-1, a hair keratin-associated protein, as a possible crucial element for the physical properties of hair shafts. J. Dermatol. Sci. 2014, 74, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Bao, Z.; Zhao, B.; Zhou, T.; Li, J.; Liu, M.; Hu, S.; Yang, N.; Chen, Y.; Wu, X. Characterization and functional analysis of krtap11-1 during hair follicle development in angora rabbits (Oryctolagus cuniculus). Genes Genom. 2020, 42, 1281–1290. [Google Scholar] [CrossRef] [PubMed]
- Rombel, I.T.; Sykes, K.F.; Rayner, S.; Johnston, S.A. Orf-finder: A vector for high-throughput gene identification. Gene 2002, 282, 33–41. [Google Scholar] [CrossRef]
- Kang, Y.-J.; Yang, D.-C.; Kong, L.; Hou, M.; Meng, Y.-Q.; Wei, L.; Gao, G. Cpc2: A fast and accurate coding potential calculator based on sequence intrinsic features. Nucleic Acids Res. 2017, 45, W12–W16. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time pcr data by the comparative c t method. Nat. Protoc. 2008, 3, 1101. [Google Scholar] [CrossRef]
- Harris, V.M. Protein detection by simple western™ analysis. In Western Blotting; Springer: Berlin/Heidelberg, Germany, 2015; pp. 465–468. [Google Scholar]
- Millar, S.E. Molecular mechanisms regulating hair follicle development. J. Investig. Dermatol. 2002, 118, 216–225. [Google Scholar] [CrossRef]
- Schneider, M.R.; Schmidt-Ullrich, R.; Paus, R. The hair follicle as a dynamic miniorgan. Curr. Biol. 2009, 19, R132–R142. [Google Scholar] [CrossRef]
- Amoh, Y.; Li, L.; Yang, M.; Moossa, A.; Katsuoka, K.; Penman, S.; Hoffman, R.M. Nascent blood vessels in the skin arise from nestin-expressing hair-follicle cells. Proc. Natl. Acad. Sci. USA 2004, 101, 13291–13295. [Google Scholar] [CrossRef] [PubMed]
- Paus, R.; Arck, P.; Tiede, S. (neuro-) endocrinology of epithelial hair follicle stem cells. Mol. Cell. Endocrinol. 2008, 288, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Moran, V.A.; Perera, R.J.; Khalil, A.M. Emerging functional and mechanistic paradigms of mammalian long non-coding rnas. Nucleic Acids Res. 2012, 40, 6391–6400. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.B.; Wang, Z.Y.; Yin, R.H.; Jiao, Q.; Zhao, S.J.; Cong, Y.Y.; Xue, H.L.; Guo, D.; Wang, S.Q.; Zhu, Y.X. A lncrna-h19 transcript from secondary hair follicle of liaoning cashmere goat: Identification, regulatory network and expression regulated potentially by its promoter methylation. Gene 2018, 641, 78–85. [Google Scholar] [CrossRef]
- Ding, H.; Zhao, H.; Zhao, X.; Qi, Y.; Wang, X.; Huang, D. Analysis of histology and long noncoding rnas involved in the rabbit hair follicle density using rna sequencing. BMC Genom. 2021, 22, 1–10. [Google Scholar] [CrossRef]
- Zheng, Y.Y.; Sheng, S.D.; Hui, T.Y.; Yue, C.; Sun, J.M.; Guo, D.; Guo, S.L.; Li, B.J.; Xue, H.L.; Wang, Z.Y. An integrated analysis of cashmere fineness lncrnas in cashmere goats. Genes 2019, 10, 266. [Google Scholar] [CrossRef]
- Lin, B.-J.; Lin, G.-Y.; Zhu, J.-Y.; Yin, G.-Q.; Huang, D.; Yan, Y.-Y. Lncrna-pcat1 maintains characteristics of dermal papilla cells and promotes hair follicle regeneration by regulating mir-329/wnt10b axis. Exp. Cell Res. 2020, 394, 112031. [Google Scholar] [CrossRef]
- Wu, M.; Liu, H. Hair follicle development from human mesenchymal cell with asymmetric distribution of polarity molecules. Cell Res. 2008, 18, S61. [Google Scholar] [CrossRef][Green Version]
- Gong, H.; Zhou, H.; Dyer, J.M.; Hickford, J.G. Identification of the ovine kap11-1 gene (krtap11-1) and genetic variation in its coding sequence. Mol. Biol. Rep. 2011, 38, 5429–5433. [Google Scholar] [CrossRef]
- Rion, N.; Rüegg, M.A. Lncrna-encoded peptides: More than translational noise? Cell Res. 2017, 27, 604–605. [Google Scholar] [CrossRef]
- Nelson, B.R.; Makarewich, C.A.; Anderson, D.M.; Winders, B.R.; Troupes, C.D.; Wu, F.; Reese, A.L.; McAnally, J.R.; Chen, X.; Kavalali, E.T. A peptide encoded by a transcript annotated as long noncoding rna enhances serca activity in muscle. Science 2016, 351, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Engreitz, J.M.; Ollikainen, N.; Guttman, M. Long non-coding rnas: Spatial amplifiers that control nuclear structure and gene expression. Nat. Rev. Mol. Cell Biol. 2016, 17, 756. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Chang, C. Long non-coding rna and chromatin remodeling. RNA Biol. 2015, 12, 1094–1098. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; He, L.; Du, Y.; Zhu, P.; Huang, G.; Luo, J.; Yan, X.; Ye, B.; Li, C.; Xia, P. The long noncoding rna lnctcf7 promotes self-renewal of human liver cancer stem cells through activation of wnt signaling. Cell Stem Cell 2015, 16, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Song, L.L.; Cui, Y.; Yu, S.J.; Liu, P.G.; Zhang, Q. Expression characteristics of bmp2, bmpr-ia and noggin in different stages of hair follicle in yak skin. Gen. Comp. Endocrinol. 2017, 260, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Lyle, S.; Liu, Y.; Solky, B.; Cotsarelis, G. Differential expression of cyclin d1 in the human hair follicle. Am. J. Pathol. 2003, 163, 969–978. [Google Scholar] [CrossRef]
- Liu, X.; Driskell, R.R.; Luo, M.; Abbott, D.; Filali, M.; Cheng, N.; Sigmund, C.D.; Engelhardt, J.F. Characterization of lef-1 promoter segments that facilitate inductive developmental expression in skin. J. Investig. Dermatol. 2004, 123, 264–274. [Google Scholar] [CrossRef]
- Bayle, J.; Fitch, J.; Jacobsen, K.; Kumar, R.; Lafyatis, R.; Lemaire, R. Increased expression of wnt2 and sfrp4 in tsk mouse skin: Role of wnt signaling in altered dermal fibrillin deposition and systemic sclerosis. J. Investig. Dermatol. 2008, 128, 871–881. [Google Scholar] [CrossRef]
- Shu, X.; Shu, S.; Cheng, H. A novel lncrna-mediated trans-regulatory mechanism in the development of cleft palate in mouse. Mol. Genet. Genom. Med. 2019, 7, e00522. [Google Scholar] [CrossRef]
- Koster, I.M. Building models for keratin disorders. J. Investig. Dermatol. 2012, 132, 1324–1326. [Google Scholar] [CrossRef]
- Choi, H.I.; Choi, G.; Kim, E.; Choi, Y.; Sohn, K.; Lee, Y.; Kim, C.; Yoon, T.; Sohn, H.; Han, S. Hair greying is associated with active hair growth. Br. J. Dermatol. 2011, 165, 1183–1189. [Google Scholar] [CrossRef] [PubMed]
- Calò, V.; Migliavacca, M.; Bazan, V.; Macaluso, M.; Buscemi, M.; Gebbia, N.; Russo, A. Stat proteins: From normal control of cellular events to tumorigenesis. J. Cell. Physiol. 2003, 197, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Coulombe, P.A. Keratin 17 modulates hair follicle cycling in a tnfα-dependent fashion. Genes Dev. 2006, 20, 1353–1364. [Google Scholar] [CrossRef]
- McGowan, M.K. Keratin 17 null mice exhibit age- and strain-dependent alopecia. Genes Dev 2002, 16, 1412–1422. [Google Scholar] [CrossRef] [PubMed]
- Müller-Röver, S.; Rossiter, H.; Lindner, G.; Peters, E.M.; Kupper, T.S.; Paus, R. Hair follicle apoptosis and bcl-2. J. Investig. Dermatol. Symp. Proc. 1999, 4, 272–277. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, B.; Li, J.; Liu, M.; Hu, S.; Yang, N.; Liang, S.; Zhang, X.; Dai, Y.; Bao, Z.; Chen, Y.; et al. lncRNA2919 Suppresses Rabbit Dermal Papilla Cell Proliferation via trans-Regulatory Actions. Cells 2022, 11, 2443. https://doi.org/10.3390/cells11152443
Zhao B, Li J, Liu M, Hu S, Yang N, Liang S, Zhang X, Dai Y, Bao Z, Chen Y, et al. lncRNA2919 Suppresses Rabbit Dermal Papilla Cell Proliferation via trans-Regulatory Actions. Cells. 2022; 11(15):2443. https://doi.org/10.3390/cells11152443
Chicago/Turabian StyleZhao, Bohao, Jiali Li, Ming Liu, Shuaishuai Hu, Naisu Yang, Shuang Liang, Xiyu Zhang, Yingying Dai, Zhiyuan Bao, Yang Chen, and et al. 2022. "lncRNA2919 Suppresses Rabbit Dermal Papilla Cell Proliferation via trans-Regulatory Actions" Cells 11, no. 15: 2443. https://doi.org/10.3390/cells11152443
APA StyleZhao, B., Li, J., Liu, M., Hu, S., Yang, N., Liang, S., Zhang, X., Dai, Y., Bao, Z., Chen, Y., & Wu, X. (2022). lncRNA2919 Suppresses Rabbit Dermal Papilla Cell Proliferation via trans-Regulatory Actions. Cells, 11(15), 2443. https://doi.org/10.3390/cells11152443