
Cross Talk between Hydrogen Peroxide and Nitric Oxide in the Unicellular Green Algae Cell Cycle: How Does It Work?



Abstract
:1. Introduction
2. ROS and RNS in Plant and Algal Cells
3. H2O2 and NO Cross-Talk
4. The Cell Cycle of Unicellular Green Algae
5. Redox-Dependent Components of Cell Cycle Machinery
6. ROS and RNS Interactions with Photosynthesis
7. ROS and RNS Mediated Control of Cell Cycle-Evidence in Higher Plants and Algae
7.1. Case 1: Medicago Sativa (Alfalfa)
7.2. Case 2: Arabidopsis
7.3. Case 3 and 4: Chlamydomonas
8. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Mullineaux, P.M.; Exposito-Rodriguez, M.; Laissue, P.P.; Smirnoff, N. ROS-dependent signalling pathways in plants and algae exposed to high light: Comparisons with other eukaryotes. Free Radic. Biol. Med. 2018, 122, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Oxidative damage and antioxidative system in algae. Toxicol. Rep. 2019, 6, 1309–1313. [Google Scholar] [CrossRef] [PubMed]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic stress and reactive oxygen species: Generation, signaling, and defense mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Michelet, L.; Zaffagnini, M.; Morisse, S.; Sparla, F.; Pérez-Pérez, M.E.; Francia, F.; Lemaire, S.D. Redox regulation of the Calvin–Benson cycle: Something old, something new. Front. Plant Sci. 2013, 4, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pokora, W.; Aksmann, A.; Baścik-Remisiewicz, A.; Dettlaff-Pokora, A.; Tukaj, Z. Exogenously applied hydrogen peroxide modifies the course of the Chlamydomonas reinhardtii cell cycle. J. Plant Physiol. 2018, 230, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Gough, D.R.; Cotter, T.G. Hydrogen peroxide: A Jekyll and Hyde signalling molecule. Cell Death Dis. 2011, 2, e213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foyer, C.H.; Noctor, G. Stress-triggered redox signalling: What’s in pROSpect? Plant Cell Environ. 2016, 39, 951–964. [Google Scholar] [CrossRef]
- Chaux, F.; Burlacot, A.; Mekhalfi, M.; Auroy, P.; Blangy, S.; Richaud, P.; Peltier, G. Flavodiiron proteins promote fast and transient O2 photoreduction in Chlamydomonas. Plant Physiol. 2017, 174, 1825–1836. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Guo, Y.; Jia, L.; Chu, H.; Zhou, S.; Chen, K.; Zhao, L. Hydrogen peroxide acts upstream of nitric oxide in the heat shock pathway in Arabidopsis seedlings. Plant Physiol. 2014, 164, 2184–2196. [Google Scholar] [CrossRef] [Green Version]
- Miyagishima, S.Y.; Nakanishi, H.; Kabeya, Y. Structure, regulation, and evolution of the plastid division machinery. Int. Rev. Cell Mol. Biol. 2011, 291, 115–153. [Google Scholar]
- Burlacot, A.; Peltier, G.; Li-Beisson, Y. Subcellular energetics and carbon storage in Chlamydomonas. Cells 2019, 8, 1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waszczak, C.; Carmody, M.; Kangasjärvi, J. Reactive oxygen species in plant signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neil, S.; Hancock, J.T.; Wilson, I.D.; Yoshioka, K.; Shinozaki, K. Reactive oxygen species, nitric oxide and signal crosstalk. In Signal. Crosstalk Plant Stress Responses; Wiley-Blackwell: Hoboken, NJ, USA, 2009; pp. 136–160. [Google Scholar]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant. Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Rentel, M.C.; Knight, M.R. Oxidative stress-induced calcium signaling in Arabidopsis. Plant Physiol. 2004, 135, 1471–1479. [Google Scholar] [CrossRef] [Green Version]
- Ugya, A.Y.; Imam, T.S.; Li, A.; Ma, J.; Hua, X. Antioxidant response mechanism of freshwater microalgae species to reactive oxygen species production: A mini review. Chem. Ecol. 2020, 36, 174–193. [Google Scholar] [CrossRef]
- Conte, M.L.; Carroll, K.S. The redox biochemistry of protein sulfenylation and sulfinylation. J. Biol. Chem. 2013, 288, 26480–26488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besson-Bard, A.; Pugin, A.; Wendehenne, D. New insights into nitric oxide signaling in plants. Annu. Rev. Plant. Biol. 2008, 59, 21–39. [Google Scholar] [CrossRef] [PubMed]
- Palavan-Unsal, N.; Arisan, D. Nitric oxide signalling in plants. Bot. Rev. 2009, 75, 203–229. [Google Scholar] [CrossRef]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, A.; Galvan, A.; Fernandez, E. Nitrate reductase regulates plant nitric oxide homeostasis. Trends Plant Sci. 2017, 22, 163–174. [Google Scholar] [CrossRef]
- Sharma, A.; Soares, C.; Sousa, B.; Martins, M.; Kumar, V.; Shahzad, B.; Zheng, B. Nitric oxide-mediated regulation of oxidative stress in plants under metal stress: A review on molecular and biochemical aspects. Physiol. Plant. 2020, 168, 318–344. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Responses of nitric oxide and hydrogen sulfide in regulating oxidative defence system in wheat plants grown under cadmium stress. Physiol. Plant. 2020, 168, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Astier, J.; Rossi, J.; Chatelain, P.; Klinguer, A.; Besson-Bard, A.; Rosnoblet, C.; Wendehenne, D. Nitric oxide production and signalling in algae. J. Exp. Bot. 2021, 72, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Gupta, A.K. Transcriptional regulation by nitric oxide in plant disease resistance. In Nitric Oxide in Plant Biology; Academic Press: Cambridge, MA, USA, 2022; pp. 597–614. [Google Scholar]
- Ötvös, K.; Pasternak, T.P.; Miskolczi, P.; Domoki, M.; Dorjgotov, D.; Szucs, A.; Fehér, A. Nitric oxide is required for, and promotes auxin-mediated activation of, cell division and embryogenic cell formation but does not influence cell cycle progression in alfalfa cell cultures. Plant J. 2005, 43, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Pokora, W.; Aksmann, A.; Baścik-Remisiewicz, A.; Dettlaff-Pokora, A.; Rykaczewski, M.; Gappa, M.; Tukaj, Z. Changes in nitric oxide/hydrogen peroxide content and cell cycle progression: Study with synchronized cultures of green alga Chlamydomonas reinhardtii. J. Plant Physiol. 2017, 208, 84–93. [Google Scholar] [CrossRef]
- Estevez, M.S.; Puntarulo, S. Nitric oxide generation upon growth of Antarctic Chlorella sp. cells. Physiol. Plant. 2005, 125, 192–201. [Google Scholar] [CrossRef]
- Zhang, Z.B.; Liu, C.Y.; Wu, Z.Z.; Xing, L.; Li, P.F. Detection of nitric oxide in culture media and studies on nitric oxide formation by marine microalgae. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2006, 12, BR75-85. [Google Scholar]
- Kim, D.; Kang, Y.S.; Lee, Y.; Yamaguchi, K.; Matsuoka, K.; Lee, K.W.; Oda, T. Detection of nitric oxide (NO) in marine phytoplankters. J. Biosci. Bioeng. 2008, 105, 414–417. [Google Scholar] [CrossRef]
- Chow, F.; Pedersén, M.; Oliveira, M.C. Modulation of nitrate reductase activity by photosynthetic electron transport chain and nitric oxide balance in the red macroalga Gracilaria chilensis (Gracilariales, Rhodophyta). J. Appl. Phycol. 2013, 25, 1847–1853. [Google Scholar] [CrossRef]
- Martínez-Ruiz, A.; Cadenas, S.; Lamas, S. Nitric oxide signaling: Classical, less classical, and nonclassical mechanisms. Free Radic. Biol. Med. 2011, 51, 17–29. [Google Scholar] [CrossRef]
- Shen, Q.; Zhan, X.; Yang, P.; Li, J.; Chen, J.; Tang, B.; Hong, Y. Dual activities of plant cGMP-dependent protein kinase and its roles in gibberellin signaling and salt stress. Plant Cell 2019, 31, 3073–3091. [Google Scholar] [CrossRef] [Green Version]
- Gross, I.; Durner, J. In search of enzymes with a role in 3′, 5′-cyclic guanosine monophosphate metabolism in plants. Front. Plant Sci. 2016, 7, 576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neill, S.J.; Desikan, R.; Clarke, A.; Hurst, R.D.; Hancock, J.T. Hydrogen peroxide and nitric oxide as signalling molecules in plants. J. Exp. Bot. 2002, 53, 1237–1247. [Google Scholar] [CrossRef] [PubMed]
- Qiao, W.; Li, C.; Fan, L.M. Cross-talk between nitric oxide and hydrogen peroxide in plant responses to abiotic stresses. Environ. Exp. Bot. 2014, 100, 84–93. [Google Scholar] [CrossRef]
- Bright, J.; Desikan, R.; Hancock, J.T.; Weir, I.S.; Neill, S.J. ABA-induced NO generation and stomatal closure in Arabidopsis are dependent on H2O2 synthesis. Plant J. 2006, 45, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Delledonne, M.; Zeier, J.; Marocco, A.; Lamb, C. Signal interactions between nitric oxide and reactive oxygen intermediates in the plant hypersensitive disease resistance response. Proc. Natl. Acad. Sci. USA 2001, 98, 13454–13459. [Google Scholar] [CrossRef] [Green Version]
- Desikan, R.; Cheung, M.K.; Bright, J.; Henson, D.; Hancock, J.T.; Neill, S.J. ABA, hydrogen peroxide and nitric oxide signalling in stomatal guard cells. J. Exp. Bot. 2004, 55, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Petřivalský, M.; Luhová, L. Nitrated nucleotides: New players in signaling pathways of reactive nitrogen and oxygen species in plants. Front. Plant. Sci. 2020, 11, 598. [Google Scholar] [CrossRef]
- Fujiwara, T.; Hirooka, S.; Ohbayashi, R.; Onuma, R.; Miyagishima, S.Y. Relationship between cell cycle and diel transcriptomic changes in metabolism in a unicellular red alga. Plant. Physiol. 2020, 183, 1484–1501. [Google Scholar] [CrossRef]
- Čížková, M.; Slavková, M.; Vítová, M.; Zachleder, V.; Bišová, K. Response of the green alga Chlamydomonas reinhardtii to the DNA damaging agent zeocin. Cells 2019, 8, 735. [Google Scholar] [CrossRef] [Green Version]
- Fisher, N.L.; Campbell, D.A.; Hughes, D.J.; Kuzhiumparambil, U.; Halsey, K.H.; Ralph, P.J.; Suggett, D.J. Divergence of photosynthetic strategies amongst marine diatoms. PLoS ONE 2020, 15, e0244252. [Google Scholar] [CrossRef]
- Carvalho, A.M.; Neto, A.M.; Tonon, A.P.; Pinto, E.; Cardozo, K.H.; Brigagao, M.R.; Guaratini, T. Circadian protection against oxidative stress in marine algae. Hypnos 2004, 1, 142–157. [Google Scholar]
- Zones, J.M.; Blaby, I.K.; Merchant, S.S.; Umen, J.G. High-resolution profiling of a synchronized diurnal transcriptome from Chlamydomonas reinhardtii reveals continuous cell and metabolic differentiation. Plant Cell 2015, 27, 2743–2769. [Google Scholar] [PubMed]
- Miyagishima, S.Y.; Fujiwara, T.; Sumiya, N.; Hirooka, S.; Nakano, A.; Kabeya, Y.; Nakamura, M. Translation-independent circadian control of the cell cycle in a unicellular photosynthetic eukaryote. Nat. Commun. 2014, 5, 3807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapuy, O.; Vinod, P.K.; Bánhegyi, G.; Novák, B. Systems-level feedback regulation of cell cycle transitions in Ostreococcus tauri. Plant. Physiol. Biochem. 2018, 126, 39–46. [Google Scholar] [CrossRef]
- Tamiya, H. Synchronous cultures of algae. Annu. Rev. Plant Physiol. 1966, 17, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Bišová, K.; Zachleder, V. Cell-cycle regulation in green algae dividing by multiple fission. J. Exp. Bot. 2014, 466, 128–132. [Google Scholar] [CrossRef]
- Ivanov, I.N.; Vítová, M.; Bišová, K. Growth and the cell cycle in green algae dividing by multiple fission. Folia Microbiol. 2019, 64, 663–672. [Google Scholar] [CrossRef]
- Zachleder, V.; Ivanov, I.N.; Kselíková, V.; Bialevich, V.; Vítová, M.; Ota, S.; Bišová, K. Characterization of Growth and Cell Cycle Events Affected by Light Intensity in the Green Alga Parachlorella kessleri: A New Model for Cell Cycle Research. Biomolecules 2021, 11, 891. [Google Scholar] [CrossRef]
- Grossman, A.R. Paths toward algal genomics. Plant Physiol. 2005, 137, 410–427. [Google Scholar] [CrossRef] [Green Version]
- Levine, R.P. Genetic control of photosynthesis in Chlamydomonas reinhardi. Proc. Natl. Acad. Sci. USA 1960, 46, 972. [Google Scholar] [CrossRef] [Green Version]
- Wakao, S.; Niyogi, K.K. Chlamydomonas as a model for reactive oxygen species signaling and thiol redox regulation in the green lineage. Plant Physiol. 2021, 187, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Cross, F.R.; Umen, J.G. The Chlamydomonas cell cycle. Plant J. 2015, 82, 370–392. [Google Scholar] [CrossRef]
- De Veylder, L.; Joubès, J.; Inzé, D. Plant cell cycle transitions. Curr. Opin. Plant Biol. 2003, 6, 536–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumura, K.; Yagi, T.; Yasuda, K. Differential analysis of cell cycle stability in Chlamydomonas using on-chip single-cell cultivation system. Jpn. J. Appl. Phys. 2003, 42, L784. [Google Scholar] [CrossRef]
- Nasmyth, K. Control of the yeast cell cycle by the Cdc28 protein kinase. Curr. Opin. Cell Biol. 1993, 5, 166–179. [Google Scholar] [CrossRef]
- Harper, J.V.; Brooks, G. The mammalian cell cycle: An overview. Methods Mol. Biol. 2005, 296, 113–153. [Google Scholar] [PubMed]
- Bišová, K.; Krylov, D.M.; Umen, J.G. Genome-wide annotation and expression profiling of cell cycle regulatory genes in Chlamydomonas reinhardtii. Plant Physiol. 2005, 137, 475–491. [Google Scholar] [CrossRef] [Green Version]
- Umen, J.G.; Goodenough, U.W. Control of cell division by a retinoblastoma protein homolog in Chlamydomonas. Genes Dev. 2001, 15, 1652–1661. [Google Scholar] [CrossRef] [Green Version]
- Goodenough, U.W. Chloroplast division and pyrenoid formation in Chlamydomonas reinhardi 1. J. Phycol. 1970, 6, 1–6. [Google Scholar]
- Miyagishima, S.Y.; Suzuki, K.; Okazaki, K.; Kabeya, Y. Expression of the nucleus-encoded chloroplast division genes and proteins regulated by the algal cell cycle. Mol. Biol. Evol. 2012, 29, 2957–2970. [Google Scholar] [CrossRef] [Green Version]
- Lau, K.W.; Ren, J.; Wu, M. Redox modulation of chloroplast DNA replication in Chlamydomonas reinhardtii. Antioxid. Redox Signal. 2000, 2, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Schnaubelt, D.; Queval, G.; Dong, Y.; Diaz-Vivancos, P.E.D.R.O.; Makgopa, M.E.; Howell, G.; Foyer, C.H. Low glutathione regulates gene expression and the redox potentials of the nucleus and cytosol in A rabidopsis thaliana. Plant Cell Environ. 2015, 38, 266–279. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.C.; Chung, C.L.; Chen, C.H.; Lopez-Paz, C.; Umen, J.G. Defects in a new class of sulfate/anion transporter link sulfur acclimation responses to intracellular glutathione levels and cell cycle control. Plant Physiol. 2014, 166, 1852–1868. [Google Scholar] [CrossRef] [Green Version]
- Kabeya, Y.; Miyagishima, S.Y. Chloroplast DNA replication is regulated by the redox state independently of chloroplast division in Chlamydomonas reinhardtii. Plant Physiol. 2013, 161, 2102–2112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potters, G.; Pasternak, T.P.; Guisez, Y.; Jansen, M.A. Different stresses, similar morphogenic responses: Integrating a plethora of pathways. Plant Cell Environ. 2009, 32, 158–169. [Google Scholar] [CrossRef] [PubMed]
- De Schutter, K.; Joubès, J.; Cools, T.; Verkest, A.; Corellou, F.; Babiychuk, E.; De Veylder, L. Arabidopsis WEE1 kinase controls cell cycle arrest in response to activation of the DNA integrity checkpoint. Plant. Cell 2007, 19, 211–225. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Wang, P.; Hou, H.; Zhang, H.; Wang, Y.; Yan, S.; Li, L. Transcriptional regulation of cell cycle genes in response to abiotic stresses correlates with dynamic changes in histone modifications in maize. PLoS ONE 2014, 9, e106070. [Google Scholar] [CrossRef]
- Varum, S.; Momčilović, O.; Castro, C.; Ben-Yehudah, A.; Ramalho-Santos, J.; Navara, C.S. Enhancement of human embryonic stem cell pluripotency through inhibition of the mitochondrial respiratory chain. Stem Cell Res. 2009, 3, 142–156. [Google Scholar] [CrossRef] [Green Version]
- Chiu, J.; Dawes, I.W. Redox control of cell proliferation. Trends Cell Biol. 2012, 22, 592–601. [Google Scholar] [CrossRef]
- Diaz-Vivancos, P.; de Simone, A.; Kiddle, G.; Foyer, C.H. Glutathione–linking cell proliferation to oxidative stress. Free. Radic. Biol. Med. 2015, 89, 1154–1164. [Google Scholar] [CrossRef]
- Fehér, A. Callus, dedifferentiation, totipotency, somatic embryogenesis: What these terms mean in the era of molec-ular plant biology? Front. Plant Sci. 2019, 10, 536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tognetti, V.B.; Bielach, A.; Hrtyan, M. Redox regulation at the site of primary growth: Auxin, cytokinin and ROS crosstalk. Plant Cell Environ. 2017, 40, 2586–2605. [Google Scholar] [CrossRef] [PubMed]
- Burhans, W.C.; Heintz, N.H. The cell cycle is a redox cycle: Linking phase-specific targets to cell fate. Free. Radic. Biol. Med. 2009, 47, 1282–1293. [Google Scholar] [CrossRef] [PubMed]
- Velappan, Y.; Signorelli, S.; Considine, M.J. Cell cycle arrest in plants: What distinguishes quiescence, dormancy and differentiated G1? Ann. Bot. 2017, 120, 495–509. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Fan, M.; Candas, D.; Zhang, T.Q.; Qin, L.; Eldridge, A.; Li, J.J. Cyclin B1/Cdk1 coordinates mitochondrial respiration for cell-cycle G2/M progression. Dev. Cell 2014, 29, 217–232. [Google Scholar] [CrossRef] [Green Version]
- Magyar, Z.; Bögre, L.; Ito, M. DREAMs make plant cells to cycle or to become quiescent. Curr. Opin. Plant Biol. 2016, 34, 100–106. [Google Scholar] [CrossRef] [Green Version]
- Dick, F.A.; Rubin, S.M. Molecular mechanisms underlying RB protein function. Nat. Rev. Mol. Cell Biol. 2013, 14, 297–306. [Google Scholar] [CrossRef] [Green Version]
- Harashima, H.; Sugimoto, K. Integration of developmental and environmental signals into cell proliferation and differentiation through RETINOBLASTOMA-RELATED 1. Curr. Opin. Plant Biol. 2016, 29, 95–103. [Google Scholar] [CrossRef]
- Horvath, B.M.; Kourova, H.; Nagy, S.; Nemeth, E.; Magyar, Z.; Papdi, C.; Scheres, B. Arabidopsis RETINOBLASTOMA RELATED directly regulates DNA damage responses through functions beyond cell cycle control. EMBO J. 2017, 36, 1261–1278. [Google Scholar] [CrossRef]
- Fehér, A. The initiation phase of somatic embryogenesis: What we know and what we don’t. Acta Biol. Szeged. 2008, 52, 53–56. [Google Scholar]
- Nowack, M.K.; Harashima, H.; Dissmeyer, N.; Bouyer, D.; Weimer, A.K.; De Winter, F.; Schnittger, A. Genetic framework of cyclin-dependent kinase function in Arabidopsis. Dev. Cell 2012, 22, 1030–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, T.; Benning, C. Nitrogen-dependent coordination of cell cycle, quiescence and TAG accumulation in Chlamydomonas. Biotechnol. Biofuels 2019, 12, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domenichini, S.; Benhamed, M.; De Jaeger, G.; Van De Slijke, E.; Blanchet, S.; Bourge, M.; Raynaud, C. Evidence for a role of Arabidopsis CDT1 proteins in gametophyte development and maintenance of genome integrity. Plant Cell 2012, 24, 2779–2791. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liu, D.; Lopez-Paz, C.; Olson, B.J.; Umen, J.G. A new class of cyclin dependent kinase in Chlamydomonas is required for coupling cell size to cell division. Elife 2016, 5, e10767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimotohno, A.; Aki, S.S.; Takahashi, N.; Umeda, M. Regulation of the plant cell cycle in response to hormones and the environment. Annu. Rev. Plant Biol. 2021, 72, 273–296. [Google Scholar] [CrossRef]
- Blaby, I.K.; Blaby-Haas, C.E.; Pérez-Pérez, M.E.; Schmollinger, S.; Fitz-Gibbon, S.; Lemaire, S.D.; Merchant, S.S. Genome-wide analysis on Chlamydomonas reinhardtii reveals the impact of hydrogen peroxide on protein stress responses and overlap with other stress transcriptomes. Plant J. 2015, 84, 974–988. [Google Scholar] [CrossRef] [Green Version]
- Reichheld, J.P.; Vernoux, T.; Lardon, F.; Van Montagu, M.; Inzé, D. Specific checkpoints regulate plant cell cycle progression in response to oxidative stress. Plant J. 1999, 17, 647–656. [Google Scholar] [CrossRef]
- Mangano, S.; Juárez, S.P.D.; Estevez, J.M. ROS regulation of polar growth in plant cells. Plant Physiol. 2016, 171, 1593–1605. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Wilson, M.H.; Wright, M.H. Redox regulation of cell proliferation: Bioinformatics and redox proteomics approaches to identify redox-sensitive cell cycle regulators. Free Radic. Biol. Med. 2018, 122, 137–149. [Google Scholar] [CrossRef]
- Foyer, C.H.; Shigeoka, S. Understanding oxidative stress and antioxidant functions to enhance photosynthesis. Plant Physiol. 2011, 155, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Dietz, K.J.; Turkan, I.; Krieger-Liszkay, A. Redox-and reactive oxygen species-dependent signaling into and out of the photosynthesizing chloroplast. Plant Physiol. 2016, 171, 1541–1550. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.F. Thylakoid protein phosphorylation, state 1-state 2 transitions, and photosystem stoichiometry adjustment: Redox control at multiple levels of gene expression. Physiol. Plant. 1995, 93, 196–205. [Google Scholar] [CrossRef]
- Kühlbrandt, W.; Wang, D.N.; Fujiyoshi, Y. Atomic model of plant light-harvesting complex by electron crystallography. Nature 1994, 367, 614–621. [Google Scholar] [CrossRef]
- Bonardi, V.; Pesaresi, P.; Becker, T.; Schleiff, E.; Wagner, R.; Pfannschmidt, T.; Leister, D. Photosystem II core phosphorylation and photosynthetic acclimation require two different protein kinases. Nature 2005, 437, 1179–1182. [Google Scholar] [CrossRef]
- Bruick, R.K.; Mayfield, S.P. Light-activated translation of chloroplast mRNAs. Trends Plant Sci. 1999, 4, 190–195. [Google Scholar] [CrossRef]
- Pfannschmidt, T.; Liere, K. Redox regulation and modification of proteins controlling chloroplast gene expression. Antioxid. Redox Signal. 2005, 7, 607–618. [Google Scholar] [CrossRef]
- Shiina, T.; Tsunoyama, Y.; Nakahira, Y.; Khan, M.S. Plastid RNA polymerases, promoters, and transcription regulators in higher plants. Int. Rev. Cytol. 2005, 244, 1–68. [Google Scholar]
- Tadini, L.; Jeran, N.; Peracchio, C.; Masiero, S.; Colombo, M.; Pesaresi, P. The plastid transcription machinery and its coordination with the expression of nuclear genome: Plastid-Encoded Polymerase, Nuclear-Encoded Polymerase and the Genomes Uncoupled 1-mediated retrograde communication. Philos. Trans. R. Soc. B 2020, 375, 20190399. [Google Scholar] [CrossRef]
- Puthiyaveetil, S.; McKenzie, S.D.; Kayanja, G.E.; Ibrahim, I.M. Transcription initiation as a control point in plastid gene expression. Biochim. Biophys. Acta (BBA)—Gene Regul. Mech. 2021, 1864, 194689. [Google Scholar] [CrossRef]
- Lindahl, M.; Kieselbach, T. Disulphide proteomes and interactions with thioredoxin on the track towards understanding redox regulation in chloroplasts and cyanobacteria. J. Proteom. 2009, 72, 416–438. [Google Scholar] [CrossRef]
- Chotewutmontri, P.; Barkan, A. Multilevel effects of light on ribosome dynamics in chloroplasts program genome-wide and psbA-specific changes in translation. PLoS Genet. 2018, 14, e1007555. [Google Scholar] [CrossRef] [PubMed]
- Mullineaux, P.M.; Exposito-Rodriguez, M.; Laissue, P.P.; Smirnoff, N.; Park, E. Spatial chloroplast-to-nucleus signalling involving plastid–nuclear complexes and stromules. Philos. Trans. R. Soc. B 2020, 375, 20190405. [Google Scholar] [CrossRef] [PubMed]
- Beltrán, J.; Wamboldt, Y.; Sanchez, R.; LaBrant, E.W.; Kundariya, H.; Virdi, K.S.; Mackenzie, S.A. Specialized plastids trigger tissue-specific signaling for systemic stress response in plants. Plant. Physiol. 2018, 178, 672–683. [Google Scholar] [CrossRef] [Green Version]
- Rhoads, D.M.; Subbaiah, C.C. Mitochondrial retrograde regulation in plants. Mitochondrion 2007, 7, 177–194. [Google Scholar] [CrossRef] [PubMed]
- Zaffagnini, M.; Fermani, S.; Costa, A.; Lemaire, S.D.; Trost, P. Plant cytoplasmic GAPDH: Redox post-translational modifications and moonlighting properties. Front. Plant Sci. 2013, 4, 450. [Google Scholar] [CrossRef] [Green Version]
- Abat, J.K.; Deswal, R. Differential modulation of S-nitrosoproteome of Brassica juncea by low temperature: Change in S-nitrosylation of Rubisco is responsible for the inactivation of its carboxylase activity. Proteomics 2009, 9, 4368–4380. [Google Scholar] [CrossRef]
- De Simone, A.; Hubbard, R.; De La Torre, N.V.; Velappan, Y.; Wilson, M.; Considine, M.J.; Foyer, C.H. Redox changes during the cell cycle in the embryonic root meristem of Arabidopsis thaliana. Antioxid. Redox Signal. 2017, 27, 1505–1519. [Google Scholar] [CrossRef] [Green Version]
- Lai, A.G.; Doherty, C.J.; Mueller-Roeber, B.; Kay, S.A.; Schippers, J.H.; Dijkwel, P.P. CIRCADIAN CLOCK-ASSOCIATED 1 regulates ROS homeostasis and oxidative stress responses. Proc. Natl. Acad. Sci. USA 2012, 109, 17129–17134. [Google Scholar] [CrossRef] [Green Version]
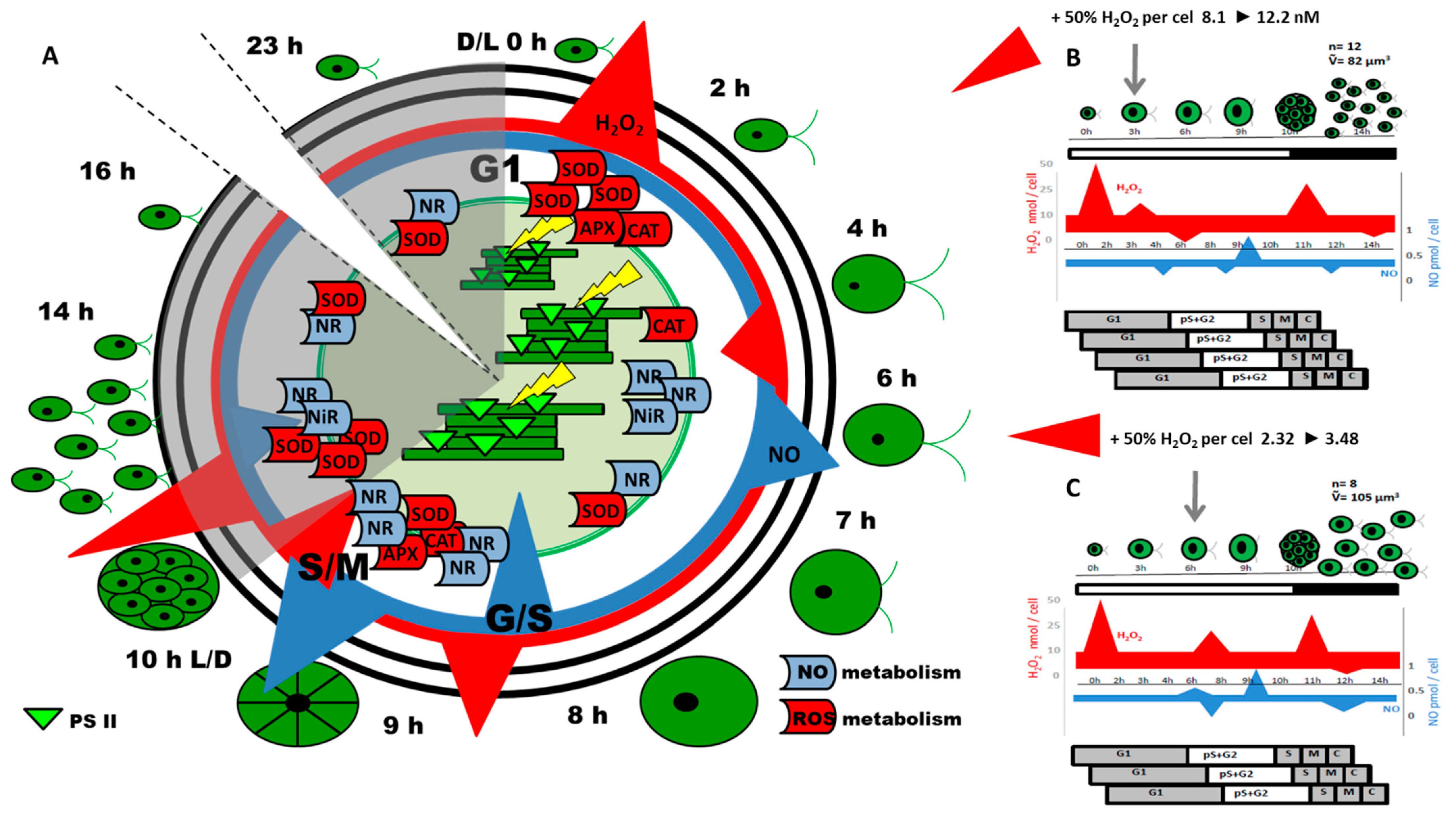

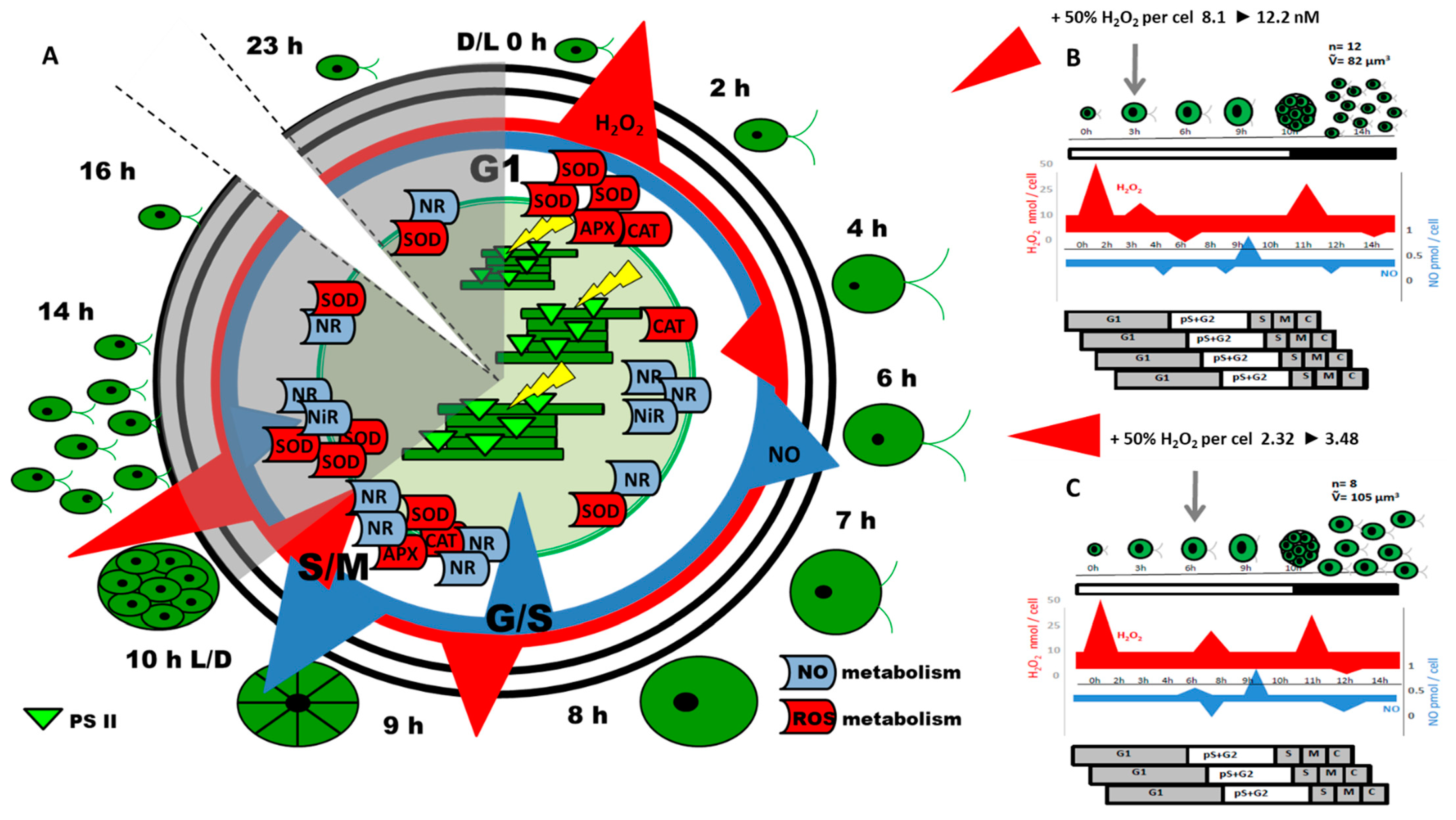{kind=link}
| Arabidopsis Thaiana | Chlamydomonas Reinhardtii | |||
|---|---|---|---|---|
| Name | AGI | Name | Locus ID | Note |
| ALY1 | AT5G27610 | REF1 | Cre10.g462250 | RNA Export Factor |
| APC6 | AT1G78770 | APC6 | Cre13.g562950 | Anaphase promoting complex subunit 6 |
| ARLA1A | AT5G37680 | ARL8 | Cre17.g708250 | ARF-like GTPase |
| ATM | AT3G48190 | ATM1 | Cre13.g564350 | ATM-like serine protein kinase |
| ATMYB3R1/PC-MYB1 | AT4G32730 | n.d. | Cre09.g399067 | –MYB-like DNA-binding protein MYB//ATMYB1 protein |
| ATXR5 | AT5G09790 | HLM3 | Cre01.g041100 | –Histone-lysine N-methyltransferase |
| CCS52A2 | AT4G11920 | CDH1 | Cre09.g406851 | Activator and specificity factor for anaphase promoting complex |
| CCS52B/FZR3 | AT5G13840 | CCS5 | Cre17.g702150 | Thioredoxin-like protein similar to Arabidopsis HCF164 |
| CDC20.2 | AT4G33260 | CDKH1 | Cre07.g355400 | Cyclin-dependent kinase |
| CDC20.4 | AT5G26900 | CDKA1 | Cre10.g465900 | Cre10.g465900–Cyclin-dependent kinase |
| CDC45 | AT3G25100 | CDC45 | Cre06.g270250 | Cell Division Cycle protein 45 |
| CDC48 | AT3G09840 | CDC48 | Cre06.g269950 | Cell Division Cycle protein 48 |
| CDC6 | AT2G29680 | CDC6 | Cre06.g292850 | Pre-initiation complex, subunit CDC6 |
| CDKC;1 | AT4g28980 | CDKC1 | Cre08.g385850 | Cyclin-dependent kinase |
| CDKC;2 | AT5g10270 | CDKC1 | Cre08.g385850_4532 | Cyclin-dependent kinase |
| CDKD1;1 | AT1G73690 | CDKD1 | Cre09.g388000 | CDK activating kinase |
| CDKG1 | AT5G63370 | CDKG1 | Cre06.g271100 | CDK activating kinase |
| CDKG2 | AT1G67580 | CDKG2 | Cre17.g742250 | CDK activating kinase |
| CDT1A | AT2G31270 | CDT1 | Cre03.g163300 | DNA replication initiation factor, CDT1-like |
| CYCA1;1 | AT1G44110 | CYCA1 | Cre03.g207900 | A-type cyclin |
| CYCB1;1 | AT4G37490 | CYCB1 | Cre08.g370401 | B-type cyclin |
| CYCD1;1 | AT1G70210 | CYCD1 | Cre11.g467772 | D-type cyclin |
| CYCD2;1 | AT2G22490 | CYCD2 | Cre06.g289750 | D-type cyclin |
| CYCD3;1 | AT4G34160 | CYCD4 | Cre09.g414416 | D-type cyclin |
| CYCD4;1 | AT5G65420 | CYCD5 | Cre04.g221301 | D-type cyclin |
| CYCD6;1 | AT4G03270 | CYCD3 | Cre06.g284350 | D-type cyclin |
| CYCT1;2 | AT4G19560 | CYCT1 | Cre14.g613900 | T-type cyclin |
| DPA | AT5G02470 | ROC59 | Cre10.g425050 | Rhythm Of Chloroplast protein 59 |
| E2F1 | AT5G22220 | E2F1 | Cre01.g052300 | Transcription factor, E2F and DP-related |
| E2F3 | AT2G36010 | E2F2 | Cre13.g572950 | Transcription factor, E2F and DP-related |
| MAPKKK13 | AT1G07150 | MAPKKK13 | Cre16.g649100 | Mitogen-Activated Protein Kinase Kinase Kinase |
| MAPKKK14 | AT2G30040 | MAPKKK14 | Cre01.g001200 | Mitogen-Activated Protein Kinase Kinase Kinase |
| MAPKKK5 | AT5G66850 | MAPKKK5 | Cre07.g339900 | Mitogen-Activated Protein Kinase Kinase Kinase |
| MAPKKK6 | AT3G07980 | MAPKKK6 | Cre03.g154250 | Mitogen-Activated Protein Kinase Kinase Kinase |
| MAPKKK7 | AT3G13530 | MAPKKK7 | Cre12.g517000 | Mitogen-Activated Protein Kinase Kinase Kinase |
| MAPKKK9 | AT4G08480 | MAPKKK9 | Cre12.g545950 | Mitogen-Activated Protein Kinase Kinase Kinase |
| MCM2 | AT1G44900 | n.d.2 | Cre10.g436600 | K06949–ribosome biogenesis GTPase [EC:3.6.1.-] (rsgA, engC) |
| MKK6 | AT5G56580 | n.d. | Cre02.g095099 | KOG0984–Mitogen-activated protein kinase (MAPK) kinase MKK3/MKK6 |
| MPK4 | AT4G01370 | MAPK8 | Cre01.g010000 | Mitogen-Activated Protein Kinase |
| ORC3 | AT5G16690 | n.d. | Cre17.g744247-PF07034 | (ORC) subunit 3 N-terminus (ORC3_N) |
| PCNA1 | AT1G07370 | PCN1 | Cre12.g515850 | Proliferating cell nuclear antigen homolog |
| SMR11 | AT2G28330 | PPR4 | Cre12.g511400 | PPR-Cyclin protein |
| SMR4 | AT5G02220 | PPR6 | Cre10.g437150 | PPR-Cyclin protein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pokora, W.; Tułodziecki, S.; Dettlaff-Pokora, A.; Aksmann, A. Cross Talk between Hydrogen Peroxide and Nitric Oxide in the Unicellular Green Algae Cell Cycle: How Does It Work? Cells 2022, 11, 2425. https://doi.org/10.3390/cells11152425
Pokora W, Tułodziecki S, Dettlaff-Pokora A, Aksmann A. Cross Talk between Hydrogen Peroxide and Nitric Oxide in the Unicellular Green Algae Cell Cycle: How Does It Work? Cells. 2022; 11(15):2425. https://doi.org/10.3390/cells11152425
Chicago/Turabian StylePokora, Wojciech, Szymon Tułodziecki, Agnieszka Dettlaff-Pokora, and Anna Aksmann. 2022. "Cross Talk between Hydrogen Peroxide and Nitric Oxide in the Unicellular Green Algae Cell Cycle: How Does It Work?" Cells 11, no. 15: 2425. https://doi.org/10.3390/cells11152425
APA StylePokora, W., Tułodziecki, S., Dettlaff-Pokora, A., & Aksmann, A. (2022). Cross Talk between Hydrogen Peroxide and Nitric Oxide in the Unicellular Green Algae Cell Cycle: How Does It Work? Cells, 11(15), 2425. https://doi.org/10.3390/cells11152425