1. Introduction
Diabetes constitutes an increasingly prevalent pathology affecting more than 5 percent of the world’s total population [
1]. This pathology is closely associated with vascular dysfunctions and atherosclerotic cardiovascular disease (ACVD), which represents the leading cause of mortality of diabetic persons [
2,
3]. Mortality due to heart attacks and strokes is increased two- to three-fold in diabetic persons [
4]. Enhanced ACVD occurrence in diabetes is due to microvascular and macrovascular complications such as coronary heart disease and ischemic strokes [
5]. Although the specific mechanisms underlying ACVD in diabetes remain unknown, research studies are increasingly identifying the vascular endothelium as a key target of circulating stressors [
6].
Erythrocytes represent vital abundant circulating elements that may be transformed into active stressors toward the vascular endothelium in ACVD [
7]. A direct relationship was established between the erythrocyte shape (width) and coronary artery disease rate [
8]. In ACVD, enhanced neovascularization of atherosclerotic plaque suggested an intense participation of erythrocytes in the progression of the pathology [
9]. More recently, a higher susceptibility for intraplaque hemorrhages has characterized atherosclerotic lesions issued from diabetics versus non-diabetics [
10].
Endothelium barrier and microvessel disruption may bring large amounts of erythrocytes into the plaque, participating in its progression and complication [
11]. Then, extravasated erythrocytes, as a source of cholesterol, hemoglobin and iron, can generate a deleterious environment contributing to oxidative stress and plaque rupture [
11,
12,
13].
Oxidative stress (OS) and damage can also occur in the vascular compartment and imply impaired/oxidized erythrocytes as central stressors in the promotion of diabetic complications [
3,
12,
14]. In hyperglycemic conditions, erythrocytes are subject to glycation. This glycation phenomenon leads to the occurrence of deleterious compounds termed advanced glycation end-products (AGEs) associated with diabetic vascular complications [
14]. Circulating AGEs can interact with vascular cells through a specific receptor family named RAGE. Very recently, our group identified the impaired structure of erythrocytes following glycation associated with an enhanced capacity to interact with and be phagocytosed by endothelial cells [
15,
16].
If accumulating evidence implicates erythrocytes as active contributors in diabetes disorder development and vascular complications, too few data concern glycated erythrocyte impact on endothelial cell functioning via phagocytosis. Hence, we hypothesized that glycation may induce damage in erythrocytes, rendering them more prone to altering endothelial cell functioning.
2. Materials and Methods
2.1. Reagents
Bicinchoninic acid (BCA), D-glucose, Drabkin’s reagent, ferrous sulfate (FeSO4), fluorescein isothiocyanate-dextran (FITCDextran), formic acid, mannitol and PKH67 (green fluorescent cell linker) were all purchased from Sigma-Aldrich (St. Louis, MO, USA). Fetal bovine serum (FBS), L-glutamine, penicillin/streptomycin, amphotericin B and HAT (hypoxanthine/aminopterin/thymidine) were obtained from Pan Biotech (Aidenbach, Germany). 3-(2-Pyridyl)-5,6-diphenyl-1,2,4-triazine-4′,4′′-disulfonic acid sodium salt (ferrozine) and 2,9-Dimethyl-1,10-phenanthroline(neocuproine) were purchased from Alfa Aesar (Kandel, Germany). MitoSOX Red mitochondrial superoxide indicator and pHrodo Red were purchased from Thermo Fisher (Carlsbad, CA, USA).
2.2. Erythrocyte Preparations
Erythrocytes were obtained from the French blood national agency (EFS-LR agreement number # 2018001378). Blood was collected in EDTA tubes (BD vacutainer
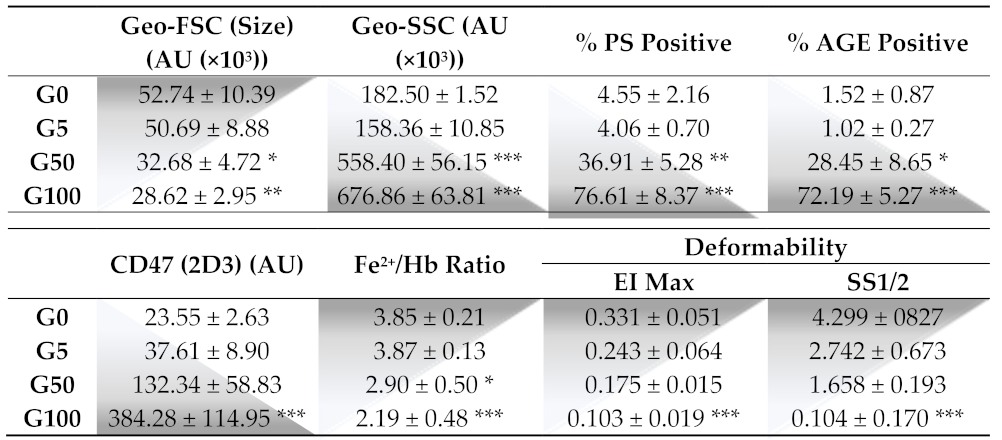
®, Franklin Lakes, NJ, USA). Erythrocytes were washed 3 times with an isotonic Ringer-lactate solution (B. Braun, Melsungen, Germany) and prepared at 20% hematocrit in phosphate buffered saline solution/5 mM glucose (PBS/0.1% glucose). Glycation was induced by incubating erythrocytes in the absence (G0) or presence of D-glucose at three increasing concentrations (5 (G5), 50 (G50) and 100 mM (G100)). A previous study, performed in our laboratory, showed that such in vitro conditions represent a suitable erythrocyte glycation model, inducing HbA1c percentages similar to those that can be measured in diabetic patients [
16]. After 5 days of incubation at 37 °C, erythrocytes were washed 3 times with Ringer-lactate solution before a direct use by FACS, ektacytometry and for endothelial cells stimulation. Quantitative determination of hemoglobin in erythrocytes was performed with Drabkin’s colorimetric assay.
2.3. Erythrocyte Senescence Characterization
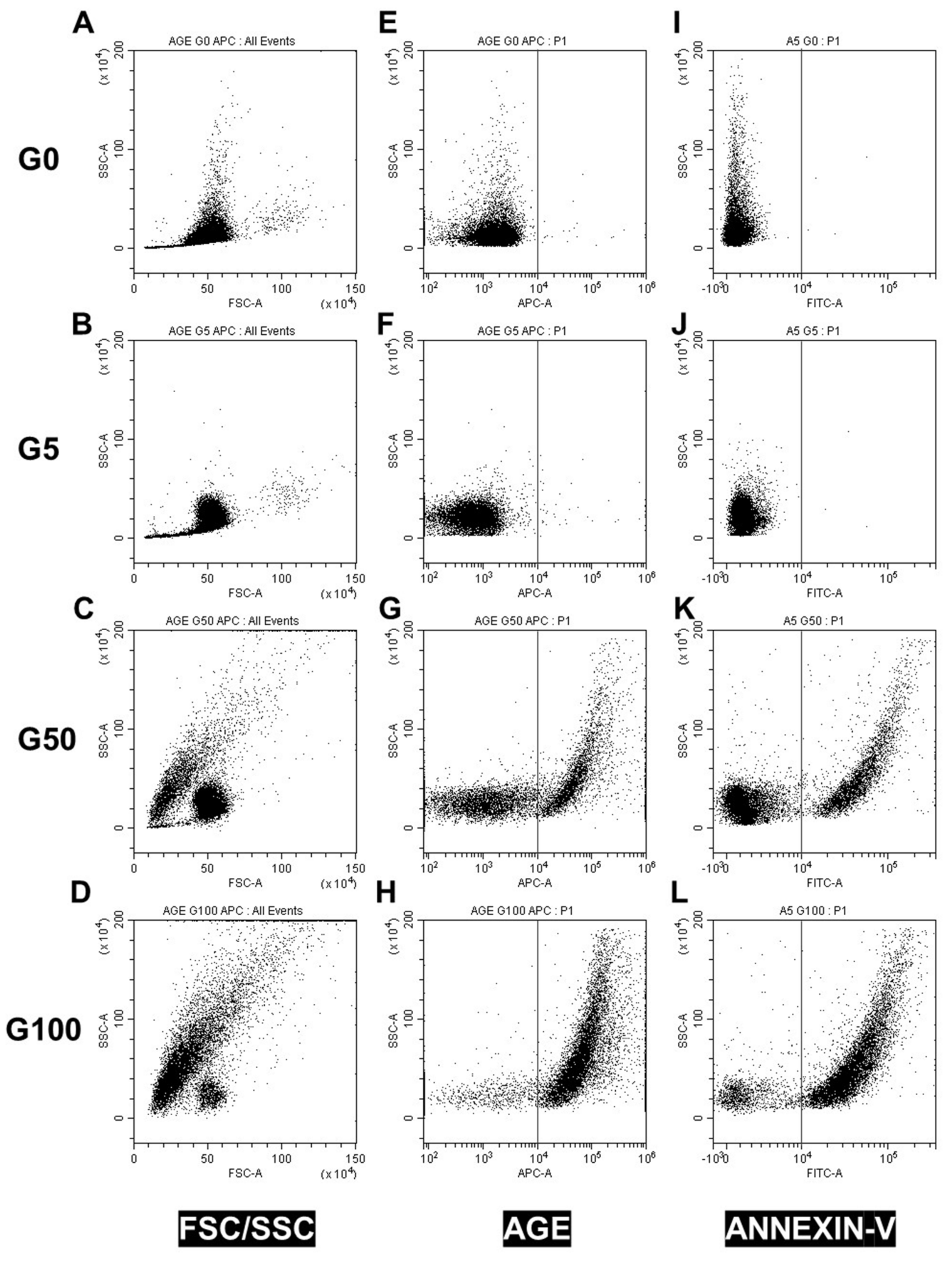
Erythrocyte shape, AGE level, CD47 expression and eryptosis were measured by flow cytometry using Beckman Coulter CytoFLEX and Cytexpert software (v2.1, Beckman Coulter, Brea, CA, USA). A specific erythrocyte cell population was selected by gating and could be characterized by its typical location in a forward scatter (FSC) versus a side scatter (SSC) parameter graph.
For AGE level determination, washed erythrocytes were incubated with AGE antibody (Abcam, 1:200 dilution) for 1 h, followed by an incubation with anti-rabbit Alexa 647-conjugated secondary IgG (Thermo Fisher, 1:200 dilution).
For conformational changes of CD47 on erythrocytes membrane, washed samples were double stained with two distinct CD47 antibodies (10 μg/mL in HEPES) directed against two different epitopes: an APC-conjugated conformation-independent antibody (clone B6H12) and a FITC-conjugated conformation-dependent (clone 2D3) antibody (Thermo Fisher, 1:200 dilution).
For phosphatidylserine (PS) exposure determination, 100 μL of erythrocytes (1:50 dilution) was incubated for 30 min at RT with 2 μg/mL annexin V-FITC in the commercial buffer (BioLegend, San Diego, CA, USA) before flow cytometry analysis. Annexin V protein, which exhibits a high affinity for PS, was measured with an excitation wavelength of 488 nm and an emission wavelength of 530 nm.
2.4. Shear Stress Gradient Ektacytometry
The determination of erythrocyte membrane deformability was performed using the deformability module of the Laser-assisted Optical Rotational Deformability Cell Analyser (LoRRca MaxSis. RR Mechatronics, Zwaag, The Netherlands), as previously described [
17]. The maximum elongation index (EI
max) and the shear stress at half maximal deformation (SS
1/2) can be obtained from deformability curves. Both parameters appeared to be relevant indicators of erythrocyte deformability capacity [
18].
2.5. Endothelial Cell Culture
EA.hy926 endothelial cell line was obtained from the American Type Culture Collection (ATCC, USA, CRL-2922™) and cultured in Dulbecco’s Modified Eagle Medium (DMEM; Pan Biotech, Aidenbach, Germany) supplemented with 25 mM glucose, 10% heat-inactivated fetal bovine serum (FBS), 2 mM L-glutamine, 100 units/mL penicillin, 100 μg/mL streptomycin, 250 μg/mL amphotericin B and HAT (100 μmol/L hypoxanthine, 0.4 μmol/L aminopterin and 16 μmol/L thymidine). Cells were maintained at 37 °C with 5% CO2 in a humidified atmosphere.
Human umbilical vein endothelial cells (HUVECs) were obtained from human umbilical cords of patients who delivered babies at Sainte-Clotilde Clinic (Saint-Denis de La Réunion, France), with informed consent following the method previously described [
19]. All procedures were approved according to French Law L.1243-3 modified by articles R1243-49 to 56, requiring the declaration of “Biobanking and preparation of cells and tissues from human body for research purpose” to MESR (French higher education and research ministry), Inserm (French National Institute for Health and Medical Research) and ANSM (French National Agency for Medicines and Health Products Safety) with the following references: C19-23 (Inserm), 2019-A01137-50 (IDRCB), and DC-2016-2614 (MESR). HUVECs were cultured on 0.2% (
w/
v) gelatin (Sigma-Aldrich) pre-coated flasks, grown in complete endothelial cell growth medium ENDOPAN-3 (Pan Biotech, Aidenbach, Germany) at 37 °C in 5% CO
2 and maintained using standard cell culture.
For the flow assay (performed in Spain), HUVECs were isolated from freshly obtained human umbilical cords following the method previously described [
20]. Umbilical cords were donated under written informed consent from mothers. All the procedures were approved by the Ethics Committee for Clinical Research at Galicia (Spain), according to the World Medical Association Declaration of Helsinki (Internal code 2013/326). Briefly, HUVECs were cultured on 0.2% (
w/
v) gelatin (Sigma-Aldrich; Merck Life Science S.L.U., Madrid, Spain) pre-coated flasks or dishes (Corning, New York, NY, USA) and grown in complete EGM-2 media (Endothelial Growth Medium-2, Lonza, Basel, Switzerland), containing 2% FBS between other components, in a humidity-saturated atmosphere with 5% CO
2 at 37 °C. Cells for the experiments were used between the second and seventh passages.
Monkey kidney normal Vero cells (ATCC, CCL-81), used for only erythrophagocytosis assay, were cultured at 37 °C under a 5% CO2 atmosphere in complete Dulbecco’s Modified Eagle Medium (DMEM) (same medium composition as that used for EA.hy926 cells).
2.6. Real-Time Monitoring of HUVECs and EA.hy926 Cells
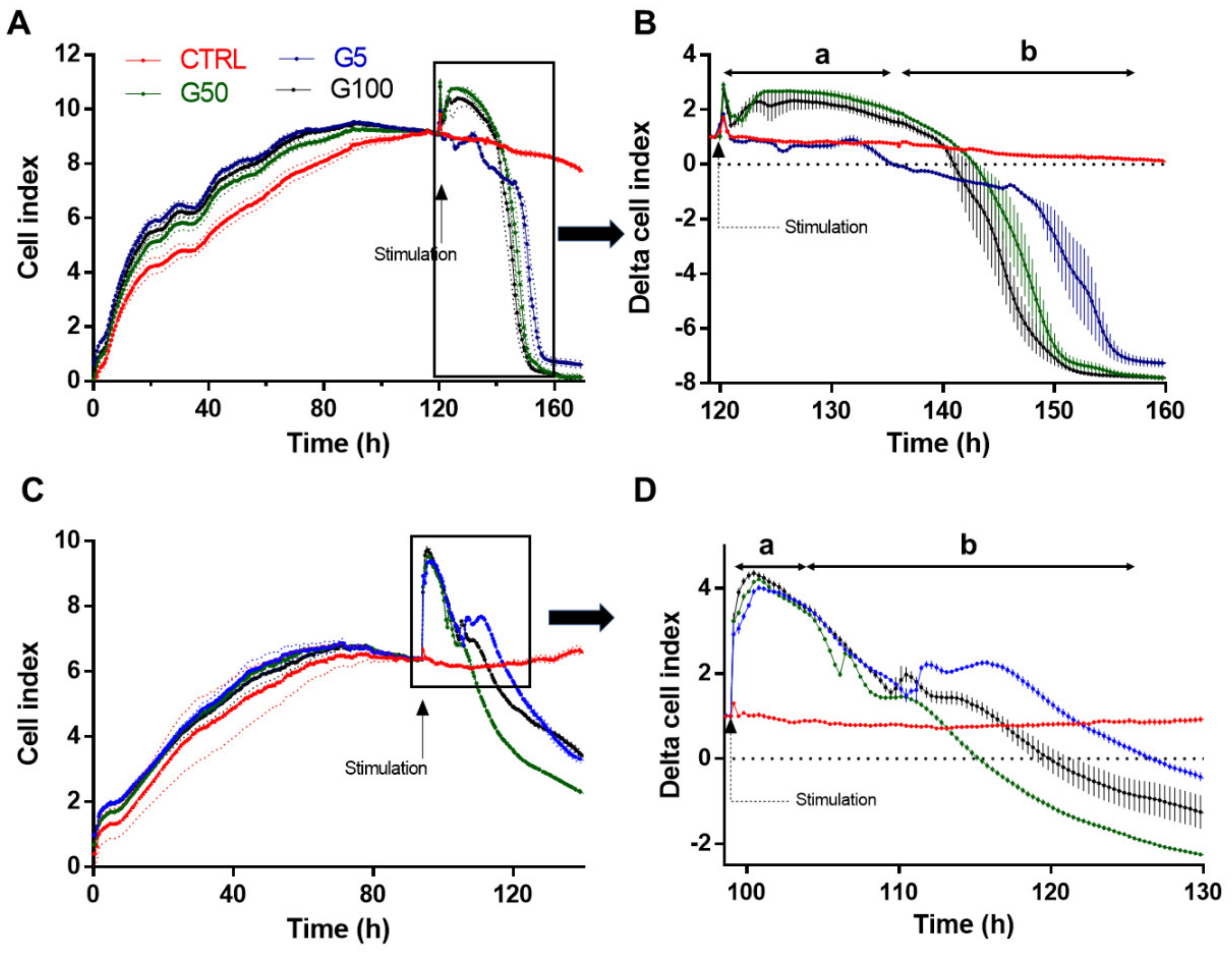
HUVECs or EA.hy926 cells were seeded at a concentration of 25,000 cells/well on xCELLigence 16-well E-plates (Acea Biosciences, San Diego, CA, USA) (gelatin-coated wells for HUVECs). The cell index was recorded continuously during cell growth until confluence, when a plateau was reached (until 90 h); cells were stimulated with erythrocyte samples at a density of 18.6 × 106 cell/cm2. Cell index measurements were automatically collected every 15 min from 0 to 160 h.
2.7. Necrosis and Apoptosis Assay
For apoptosis/necrosis study, HUVECs were collected by trypsinization and washed with PBS after 6 h stimulation with erythrocytes. Then, cells were successively incubated with 2 μg/mL annexin V-FITC and 2 μg/mL propidium iodide (Sigma) in 100 μL binding buffer (BioLegend) for 30 min at RT before analysis by cytometry.
2.8. Protein Extraction from HUVECs and EA.hy926 Cells
For total iron and heme determination in HUVECs and EA.hy926, 6-well plates were used for cell culture. After 6 h or 12 h stimulation with erythrocyte preparations, cells were washed 3 times with PBS and non-specifically bound erythrocytes were lysed with distilled water. Cells were then detached by trypsin and lysed in 50 mM Tris buffer (pH 7.5), 0.1 mM EDTA and 8 M urea and total cell proteins were quantified by BCA assay.
2.9. Erythrophagocytosis Assays Using pHrodo and PKH67 Probes
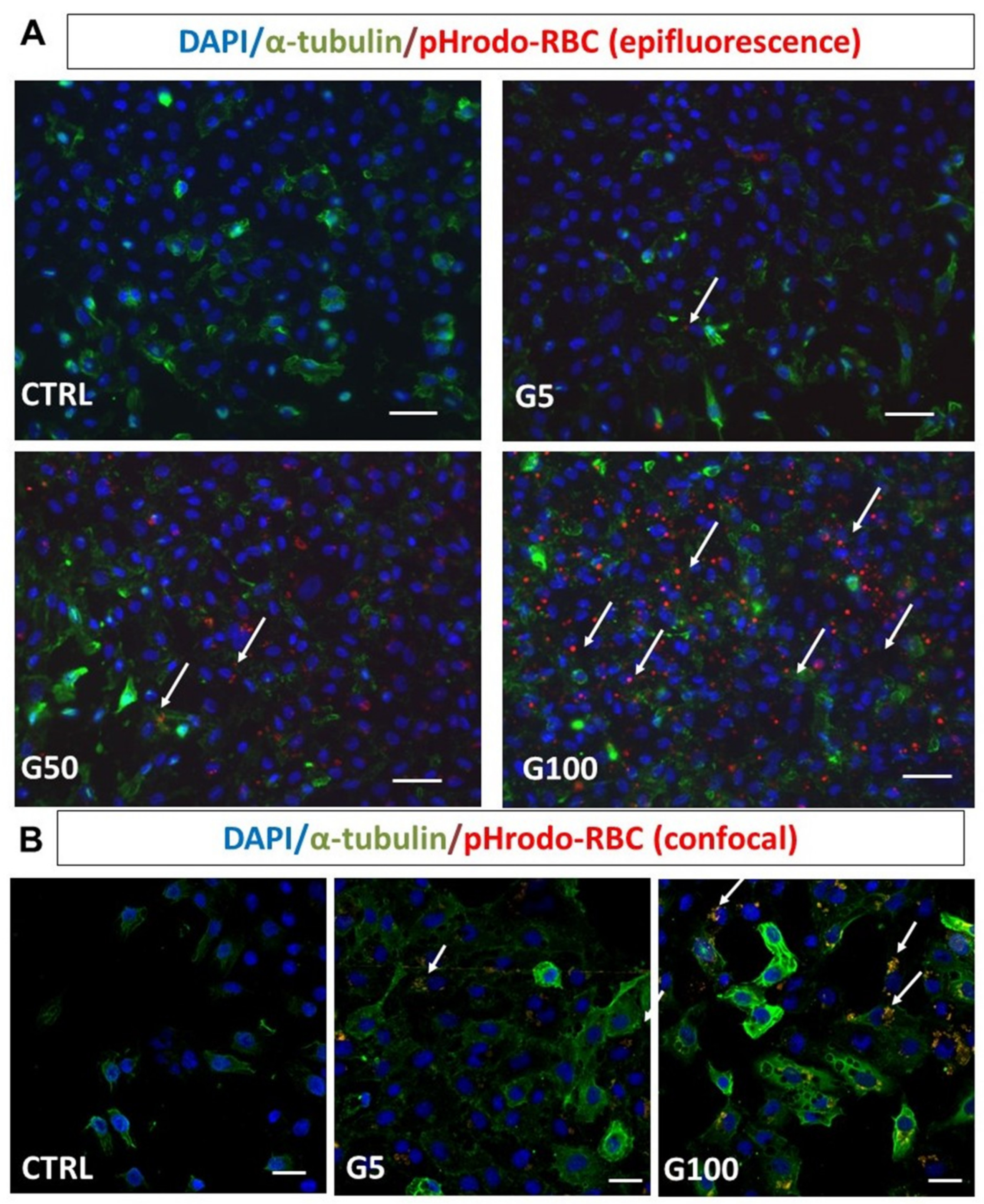
One of the common ways to assess erythrophagocytosis consists of erythrocyte labeling with fluorescent probes such as pHrodo or PKH67 (Turpin, 2022 unpublished manuscript). Erythrocyte samples are resuspended at 20% hematocrit and labeled with the fluorescent markers pHrodo or PKH67, according to the manufacturer’s instructions. For pHrodo, 109 erythrocytes were labeled with 2 µL pHrodo Red succinimidyl ester in a final volume of 1 mL PBS for 60 min at 37 °C. For PKH67, 50 µL of erythrocytes (corresponding to 2.5 × 107 cells) resuspended in 1 mL diluent buffer (supplied in the kit), were mixed to 2 μM PKH67 solution. The mixture was incubated at 37 °C for 10 min. Probe in excess was removed by several washing steps with PBS followed with 10 min centrifugation at 2000× g. Labeled erythrocytes were suspended with PBS at a final 20% of hematocrit prior to direct incubation with cultured endothelial cells for 6 h or 12 h. Endothelial cells treated in the same manner, but with unlabeled erythrocytes, were used as negative controls. After incubation and an intensive cell washing (three times with PBS) to get rid of non-phagocytosed erythrocytes, cells were subjected to flow-cytometry analysis, epi-fluorescence microscopy (Nikon Eclipse 80i) or confocal microscopy (Nikon Eclipse Ti2).
2.10. Human Umbilical Artery—Histological Analysis
From both umbilical cord arteries of approximately 6 cm in length, residual blood was removed by PBS flushing steps using a syringe equipped with a (23G) needle. Cleaned endothelial veins were filled either with glycated erythrocytes (G50 at a 20% hematocrit) or PBS (control) and sealed at each end. The cords were then fully immersed in PBS/ethanol solution (50:50, v/v) and incubated at 37 °C for 3 days. Following incubation, umbilical arteries were intensively washed with 4–5 volumes of PBS. After Wharton’s jelly removal, arteries were fixed in 4% paraformaldehyde and then embedded in OCT (Tissue Tek, Sakura Finetek, Flemingweg, The Netherlands) following standard histological protocols. Six micrometer-thick sections were prepared using a cryotome (Shandon™ cryotome FE, Thermo Scientific, Waltham, MA, USA) and were washed twice in PBS/0.1% Tween 20 before being blocked in PBS/0.1% Tween 20 containing 2% BSA. Next, sections were incubated with the following primary antibodies: anti-glycophorin A at 2.6 μg/mL (Dako, 1:500 dilution) combined with anti-PECAM (CD31) (Sigma, 1:500 dilution) or anti-von Willebrand factor at 3.17 μg/mL (Dako, 1:500 dilution) overnight at 4 °C. After two washing steps with PBS/0.1% Tween 20, sections were incubated with DAPI (4′,6′-diamidino-2-phenylindole) combined with secondary antibodies: donkey anti-rabbit Alexa Fluor 488 (Abcam, 1:500 dilution) for glycophorin A and von Willebrand factor and goat anti-mouse Alexa Fluor 594 (Abcam, 1:500 dilution) for CD31. After 2 h incubation at room temperature, sections were rinsed and the slides were mounted in Fluoromount and microscopy analysis was performed with a NanoZoomer S60 digital slide scanner (Hamamatsu).
2.11. Ferrozine Assay (Iron Level)
The quantitation of iron in endothelial cultured cells and in erythrocytes was performed by using the ferrozine-based colorimetric assay [
21]. For iron concentration in erythrocytes, an aliquot of erythrocyte samples was mixed with an equal volume of a specific buffer inducing iron release (equal volume of 1.4 M HCl and 4,5% KMnO
4 in H
2O). After 2 h incubation at 60 °C and a cooling step at room temperature, 30 µL of iron detection reagent was added (6.5 mM ferrozine, 6.5 mM neocuproine, 2.5 M ammonium acetate and 1 M ascorbic acid dissolved in water). After 30 min of incubation at room temperature, 200 µL of samples was transferred in a well of a 96-well plate and the absorbance was read at 550 nm with a microplate reader (FLUOstar Optima, BMG Labtech, Ortenberg, Germany). The total erythrocyte iron was calculated with respect to the calibration curve established with the ferrous sulfate (FeSO
4, Sigma-Aldrich, Darmstadt, Germany) (calibration range 25–400 μM) and normalized to hemoglobin content. Results were expressed as µmol of Fe
2+ per µmol of hemoglobin.
For total iron content determination in HUVECs and EA.hy926, the same protocol was applied on 100 μL of cell lysates. The total intracellular iron was normalized according to total protein content. Results were expressed as µmol of Fe2+ per µg of protein.
2.12. Heme Assay
Heme content in endothelial cells (HUVECs and EA.hy926) was assessed following the protocol previously described [
22]. After stimulation with erythrocyte samples in a 6-well plate, confluent cells were washed three times with PBS and cells were “solubilized” in 1 mL of concentrated formic acid. The heme concentration in the formic acid solution was determined with a spectrophotometer (Secoman Uvline 9600) at 398 nm. Intracellular heme content was calculated with respect to the erythrocyte solution calibration curve (calibration range: 0–2.2 × 10
7 erythrocytes). The result was expressed as a phagocytic index, corresponding to the quantity of phagocytosed erythrocytes reported to the treated endothelial cell number.
2.13. Mitochondrial Oxidative Stress
Mitochondrial oxidative stress was assessed on HUVECs by flow cytometry using Beckman Coulter’s CytoFLEX with MitoSOX Red dye as fluorescent probe, as previously described [
23]. Experiments were done in a 12-well plate with each condition performed in triplicate. HUVECs were incubated with 5 mM MitoSOX Red solution after 1 h treatment with modified erythrocytes. Results are expressed as the percentage of labeled cells, relative to the signal obtained for the vehicle control (HUVECs incubated with complete medium).
2.14. Permeability Assay
For endothelium permeability determination, HUVECs (3 × 105) were seeded on Millicell® 12-well hanging cell culture inserts (PET membrane with 0.4 μm pores)(Millipore, Burlington, MA, USA). Confluent cells were loaded with isothiocyanate (FITC)-labeled dextran (4 kDa and 70 kDa), prior to and during cell incubation in the absence (control) or the presence of erythrocytes (18.6 × 106 cell/cm2). Mannitol hyperosmotic solution (1.4 M) was used as a positive control for endothelial barrier opening. After 6 h of incubation and three successive washes with PBS, 800 μL of 10 kDa or 70 kDa dextran solutions (prepared at 0.385 mg/mL in serum-free RPMI 1640 (without phenol red)) were added on cells (apical chamber). The basolateral chamber was composed of serum-free RPMI 1640 (without dextran at the beginning of the test). Medium from the basolateral chamber was collected and replenished with fresh medium every 15 min for 1 h. The concentrations of FITC-dextran in medium from the basolateral chamber were determined by fluorescence measurements in collected samples with a microplate reader (Fluostar Optima; emission wavelength: 485 nm, excitation wavelength: 520 nm). The concentrations of FITC-dextran in medium from the basolateral chamber were calculated with respect to the FITC-dextran calibration curve (calibration range: 0.001–1 mg/mL). Results were expressed as a permeability increase calculated with the formula Ps/Pctrl where Ps and Pctrl are the slope of the curve corresponding to cumulated FITC-dextran concentration in medium against time for sample and control, respectively.
2.15. Immunochemistry Analysis
For fluorescence detection of erythrophagocytosis with pHrodo and PKH67 probes or for immunofluorescence detection of conjugated antibodies, EA.hy926 or HUVECs were seeded on coverslips and incubated for 6 h with or without erythrocytes at a density of 18.6 × 106 cell/cm2.
For immunofluorescence, HUVECs were washed three times with PBS and fixed for 20 min in 4% paraformaldehyde solution. Cells were then permeabilized with PBS/0.05% Triton X-100 for 10 min, blocked with PBS/1% BSA for 30 min and were stained with anti-CD31 (Sigma; 1:200 dilution), α-tubulin (Sigma; 1:200 dilution), mouse anti-human VE-cadherin (Santa-Cruz Biotechnology, 1:50 dilution) or mouse anti-human E-selectin (R&D Systems, 1:200 dilution). After 1 h incubation, cells were washed three times with PBS and incubated with secondary antibodies for 2 h. The secondary antibodies used were either a donkey anti-mouse antibody conjugated to Alexa Fluor A488 (Abcam, 1:500), a goat anti-rabbit conjugated to Alexa Fluor 488 (Life technologies; 1:500 dilution) or a rabbit anti-mouse antibody conjugated to Alexa Fluor 488 (Life technologies; 1:500 dilution). Endothelial cells nuclei were stained with Hoechst 33,342 (NucBlue® Live ReadyProbes® Reagent; R37605; Molecular Probes, Thermo Fisher, Madrid, Spain) or with DAPI (Sigma, 0.1 μg/mL) in PBS at room temperature for 20 min. Controls using a non-relevant IgG were included in each set of experiments. Images were acquired using an inverted fluorescence microscope (IX70, Olympus Iberia, L’Hospitalet de Llobregat, Spain), equipped with the CellSens software (Olympus).
2.16. Fabrication of a 3D Device Mimicking Microvessels
The microfluidic device used for the in vitro experiments was a Y-shaped channel with an internal circular diameter of 2 mm made by soft lithography of polydimethylsiloxane (PDMS, Sylgard 184; Dow Chemical Co., Midland, MI, USA). A detailed description of the method of fabrication is included in the
Supplementary Materials. In brief, the master consists of a rectangular box that features a Y-shaped outward channel with a semicircular profile 2 mm in diameter at its base. The master was printed with a stereolithographic 3D printer (Form 3B printer; Formlabs, Somerville, MA, USA) in Clear V4 resin (Formlabs), because of its high precision and good performance for replication [
24]. The master was post-cured by washing it with isopropanol (10 min in the Form Wash tank; Formlabs), dried and cured with UV light (405 nm wavelength) for 30 min at 60 °C. Two masters were needed to conform the two halves of the final device (
Supplementary Figure S1).
PDMS was used as the material to replicate the channel thanks to its optical transparency, permeability to gasses, elasticity and biocompatibility [
25]. Replication was made by soft lithography of PDMS (curing agent in a ratio of 10:1). PDMS was degassed in a vacuum chamber (40 min at 400 mbar) and cured in an oven (12 h at 60 °C). The two semicircular channels were bonded in a plasma cleaner under oxygen atmosphere (Diener Zepto; Diener, Ebhausen, Germany) and then thermally treated (20 min at 100 °C) to favor full bonding and improve optical quality.
2.17. Flow Assay
A functional assay was designed to investigate the possible effects of glycated erythrocytes in human endothelial cells under biomimetical conditions. The flow devices were sterilized following autoclaving at 121 °C for 1 h. After that, the channels of the devices were coated with fibronectin (Gibco) at 5 µg/mL in 0.02% gelatin solution at 37 °C for 3 h. This solution was removed immediately before seeding the cells. Confluent HUVECs were trypsinized (0.25% in Hank’s balanced salt solution with 1 mM EDTA; Gibco, Thermo Fisher Scientific, Waltham, MA, USA) and seeded in the pre-coated PDMS channels at a concentration of 1.5 × 106 cells/mL and maintained in a humidified incubator at 37 °C for 3 h. Then, the devices were flipped over and the seeding process was repeated to coat the cells on the entire channel surface. Once covered with HUVEC, the devices were maintained overnight in the incubator until the start of the fluidic experiment. Perfusion started at 0.5 mL/min with EGM-2 medium to avoid damage to the monolayer, and velocity was doubled every 60 min until reaching 3 mL/min with a peristaltic pump (Ismatec ISM596D Reglo, Ismatec, Wertheim, Germany).
Preparations of erythrocytes, subjected to 5 days glycation with 100 mM D-glucose, were compared to preparations incubated with a physiological concentration of glucose (5 mM). Erythrocytes were introduced in the perfusion circuit under sterile conditions at a concentration of 2.8 × 10
5 erythrocytes/µL and they circulated at a flow of 3 mL/min for 5 h through the channel device coated with an endothelial cells’ monolayer on the inner wall. Afterwards, the microfluidic chambers were washed with PBS for 5 min to remove any remaining erythrocyte in circulation. The cells were then fixed using paraformaldehyde (PFA) 4% (Ted Pella, INC., Redding, CA, USA) for 10 min and were then washed with PBS. Immunofluorescence staining of protein markers or nucleus staining was then performed. The nuclei of the endothelial cells were stained with Hoechst 33,342 for cell counting on each part of the bifurcation channel and cells were also stained with VE-cadherin and quantified by fluorescence intensity. Three to five fluorescent and contrast phase images (2200 × 1600 µm) from each channel were taken from at least three independent experiments (Olympus IX51, Olympus, Shinjuku, Tokyo, Japan). Images were processed with ImageJ software [
26] to calculate the number of cells in the images by counting the number of nuclei.
2.18. Numerical Simulation
A detailed description of the numerical methods is included in the
Supplementary Materials. In brief, fluid dynamics was analyzed with in silico numerical simulations using Star-CCM+ software [Star-CCM+ documentation] to design geometries, build a grid and carry out the experiments. Fluid dynamics was simulated by the incompressible Navier–Stokes equations described below:
where ρ is the fluid density, μ is the fluid viscosity, v
→ is the fluid velocity and p is the pressure.
Computations were run until a steady state was reached and convergence monitors were set at 10
−5. The domain used to perform the numerical simulation was a bifurcation with an opening angle α equal to 90° and sharp vertex (carina). Medium was considered as fluid in experiments and in numerical simulations with a density of 1025 kg/m
3 and a viscosity equal to 0.0015 Pa·s. Zero pressure was set at the outlets and the inlet boundary condition was defined in order to achieve the same Reynolds number as those reached in the experiments, and thus the same flow behavior. Reynolds number was always less than 100 and thus laminar flow was guaranteed for all the simulations. Rigid erythrocytes were modeled with a biconcave shape with a diameter of 6 µm. The forces acting on erythrocytes were drag and lift force and gravity (see the
Supplementary Materials for their mathematical description). An appropriate numeric mesh was established in order to ensure that the results obtained do not depend on the discretization of the geometry (see the
Supplementary Materials for a complete description).
2.19. Statistical Analysis
All the results are expressed as the mean +/− SEM of multiple experiments. Statistical analysis was achieved using Prism software (Prism software, GraphPad, San Diego, CA, USA). Significant differences (
p < 0.05) between the means were determined by one-way analysis of variance (ANOVA) procedures followed by Tukey’s multiple comparison test, respectively. In the numerical simulations, the independence of the geometrical mesh employed was assessed using four different types of mesh that showed less than 5% of variation in the magnitude measured (see the
Supplementary Materials). This allowed us to consider the differences observed in the results about the time of residence and number of impacts of erythrocytes as statistically significant.
4. Discussion
In this study, a glycated erythrocyte model was established following pathological conditions that can be encountered in diabetic patients. Incubation with high concentrations of glucose (until 100 mM) at 37 °C for a period of 5 days was chosen to reproduce conditions occurring during blood stasis in the neovessels of atherothrombotic plaques. Recent publications from our group revealed that such in vitro conditions induced HbA1c percentages and cell characteristics quite similar to those that can be measured in diabetic patients [
15,
16].
Here, we confirm that erythrocyte morphology following glycation was characterized by a reduced size and heterogeneous shapes associated with a drastic loss of deformability. Among the key cytoskeleton proteins involved in the membrane flexibility, β-actin was shown to be glycated upon exposure to high glucose concentrations, and associated with a decrease in erythrocyte deformability [
28]. Other important membrane proteins, such as ankyrin, band 3 or glycophorin, are closely linked to erythrocyte deformability and are also prone to glycation [
29]. As indicated by the reduced FSC value and enhanced SSC value, glycation causes cellular shrinkage in erythrocytes and membrane blebbing, which correspond to peripheral cell irregularities. This explains the heterogeneity in the morphology of erythrocytes [
30]. Cell shrinkage and membrane blebbing constitute typical hallmarks of senescent or eryptotic erythrocytes [
31].
As for senescent cells, we showed that glycated erythrocytes expose “eat-me” signals on their surface for their recognition and subsequent elimination by phagocytes. In particular, a high AGE expression was featured on erythrocytes when glycated. AGEs were shown to promote erythrocyte interaction with many cells such as endothelial cells, expressing their receptors such as RAGE or CD36 [
32]. In parallel, we showed that glycation triggers eryptosis as evidenced by the enhanced phosphatidylserine exposure at the erythrocyte surface. Glucose-mediated glycation can impair erythrocyte pump activity, and specifically the Ca
2+-ATPase pump allowing the active transport of Ca
2+ across the erythrocyte cell membrane. The activation of this pump upon glycation induces a massive intracellular calcium accumulation causing cell shrinkage and phosphatidylserine exposure [
33,
34]. Interactions between glycated erythrocytes and endothelial cells can occur through phosphatidylserine recognition as reported in numerous studies [
35,
36]. CXCL16 represents the main scavenger receptor for phosphatidylserines involved in eryptotic erythrocyte clearance by endothelial cells, but many other phosphatidylserine receptors have been identified [
37]. In addition, following erythrocyte glycation, the CD47 senescence marker has been shown to undergo a conformational change as indicated by the increased expression in CD47 clone 2D6 reflecting a variable domain of the protein. This conformational change in CD47 promotes the specific recognition for glycated erythrocytes as an “eat me” signal by potential phagocytic cells [
38]. Indeed, the conformational change of CD47 allows its binding to thrombospondin-1 (TSP-1) and the subsequent recognition of the complex formed (CD47-TSP1) by the signal regulatory protein α (SIRPα), regulating phagocytosis [
38].
The differential expressions of surface markers such AGEs, PS and CD47 in erythrocytes following glycation can clearly trigger their recognition by endothelial cells. Here, we showed that interaction between glycated erythrocytes and endothelial cells occurs rapidly and impacts cell homeostasis as indicated by cell impedance results performed on both EA.hy926 and HUVECs. Indeed, cell stimulation by erythrocytes produces a significant increase in the cell index in the first 6 h, suggesting an interaction of glycated erythrocytes with the endothelial cells. This interaction cannot be attributed to a simple adhesion of erythrocytes with endothelium, but rather to an enhanced erythrophagocytosis phenomenon in endothelial cells observed after only 6 h of exposition with glycated erythrocytes. Although in direct contact with circulating blood compounds, endothelial cells were not considered as specialized phagocytic cells involved in senescent erythrocyte clearance. However, like other cell types such as smooth muscle cells (SMCs), endothelial cells can bind and ingest aged or senescent erythrocytes [
15,
32]. In addition, and by using DAF as a colorimetric method of detection of internalized erythrocytes, recent studies from our group reported endothelial cell (EA.hy926) capacity to preferentially phagocytose glycated erythrocytes after 24 h incubation [
15,
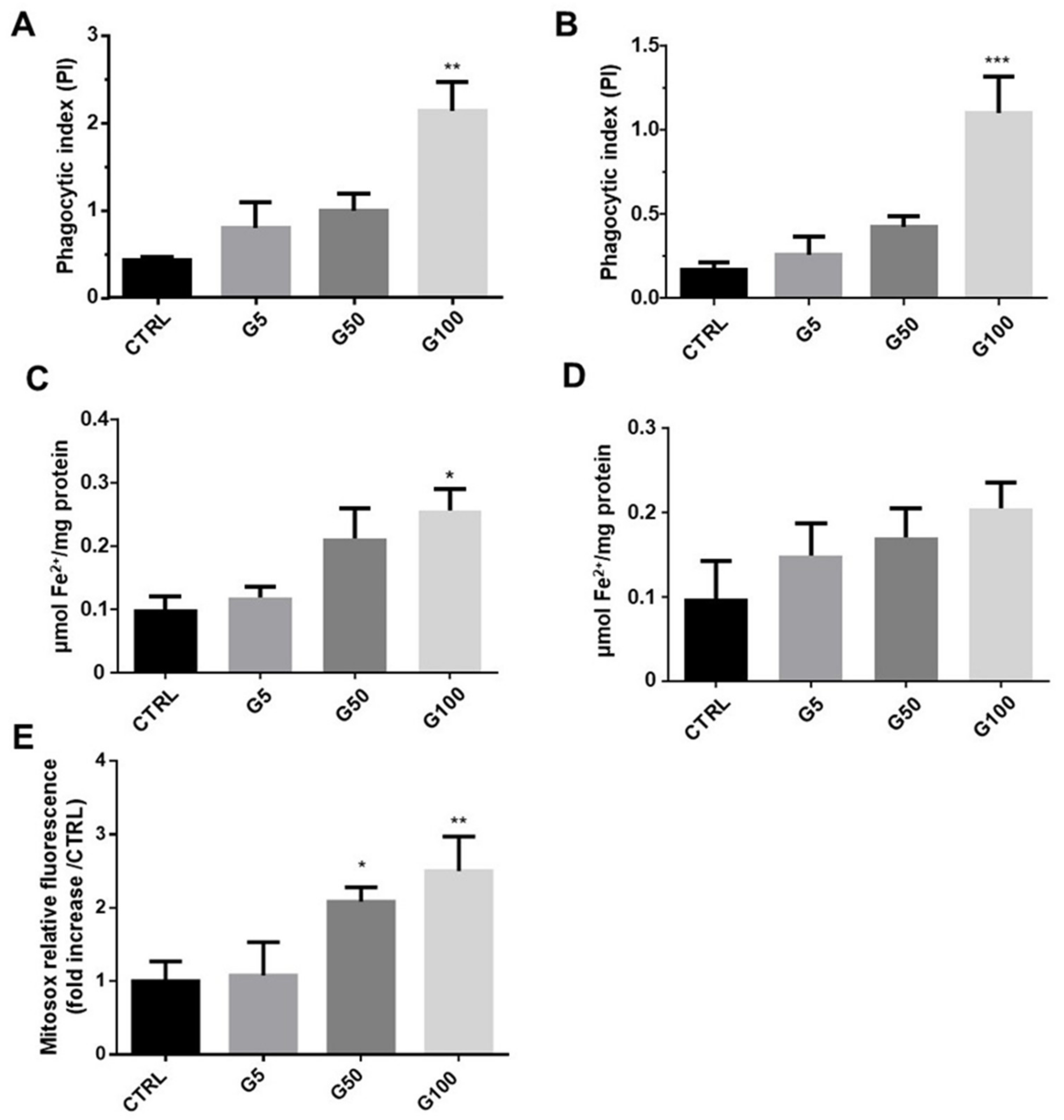
16]. In the present work, we highlighted the internalization of erythrocytes using fluorescence probes (pHrodo and PKH67), which allow a more quantitative and specific detection of erythrophagocytosis than DAF reagent.
In relation to the different markers identified on glycated erythrocytes, many pathways may be involved in the phagocytic mechanism. Erythrocyte–endothelium interaction can be mediated via the AGE–RAGE axis as shown by Wautier et al. on HUVEC primary cultures [
32]. The recognition of phosphatidylserine from eryptotic erythrocytes by their specific cell receptors may be a parallel mechanism for the erythrophagocytic process. Finally, the CD47-TSP1-SIRPa axis constitutes a possible pathway requiring in depth investigations.
In our experimental conditions, increased and early erythrophagocytosis of glycated erythrocytes led to deleterious impacts on the endothelial layer integrity. Indeed, upon glycated erythrocyte incubation, the endothelium became permeable to proteins of 10 kDa or below. VE-cadherin is a component of endothelial cell-to-cell adherens junctions that are associated with the impermeability along with tight and gap junction proteins [
39]. Though VE-cadherin is known to be involved in the endothelial barrier functioning, its expression was not altered in our experimental conditions. Other junction proteins such as ZO-1 or occludin for tight junctions or connexins for gap junctions could be investigated. This endothelial activation and dysfunction, observed through the prism of altered permeability, could also induce the expression and secretion of adhesion molecules such as VCAM-1, ICAM-1 and E-selectin [
40]. To extend this study, it would therefore be relevant to determine whether endothelial activation occurs with erythrophagocytosis and exacerbates erythrocyte adhesion at the endothelial surface.
Iron is known to play a key role in the atherosclerosis process, particularly in the activation of the endothelium [
41,
42]. Erythrocyte hemoglobin contains most of the iron in the body (70%), which allows erythrocyte the capacity to carry oxygen. The four iron atoms present in each hemoglobin molecule can bind four oxygen molecules. In our experimental conditions, we showed a drastic decrease in iron content together with a marked reduction in the iron/hemoglobin molar ratio in glycated erythrocytes. The overall loss of iron can easily be explained by the increased hemolysis due to erythrocyte fragility following glycation. By contrast, the decrease of the iron/hemoglobin molar ratio in glycated erythrocytes described here for the first time is quite unexpected. To explain this result, we could hypothesize that glycation impacts the attachment of heme groups to globin units. Indeed, we observed that the heme/hemoglobin molar ratio was reduced following erythrocyte glycation (data not shown). It is worth noting that the lack of heme iron in glycated erythrocytes could lead to deleterious functional consequences.
Despite iron deficiency in glycated erythrocytes, an enhanced intracellular accumulation of the metal was measured in endothelial cells following erythrophagocytosis. Indeed, we showed an intracellular increase in heme associated with iron accumulation in EA.hy926 and HUVECs incubated with glycated erythrocytes. It is worth noting that the measured increase in intracellular iron content was consistent with the heme content of endothelial treated cells. Indeed, this accumulation of iron results from heme degradation by heme oxygenase following erythrophagocytosis [
43,
44]. Then, the iron released from the heme molecule may not be taken up by ferroportin to be exported extracellularly [
45]. In glycated erythrocyte-treated endothelial cells, these high levels of cellular iron probably participate in the overproduction of ROS via the Fenton reaction. Here, we report an overproduction of mitochondrial superoxide radical anion, the precursor of many other ROS. As described in numerous studies, this production of superoxide radical anion and other related ROS constitute relevant contributors to the endothelial dysfunction we observed [
46,
47]. Indeed, ROS can react with NO produced by endothelial cells, forming the highly oxidant species ONOO. NO bioavailability then becomes dramatically reduced, causing platelet aggregation and enhanced vasoconstriction [
48]. Enhanced oxidative stress was also shown to induce expression of adhesion molecules by endothelial cells promoting immune cell recruiting and inflammation [
49].
As far as the impact of glycated erythrocytes on endothelial cells is concerned, one should note that cells did not undergo any apoptotic or necrotic processes. We suppose that a prolonged exposure to glycated erythrocytes could impact the migration and proliferation capacity of endothelial cells as previously shown by our group [
15]. Cytotoxic effects of glycated erythrocytes occurred following 24 h incubation with EA.hy926 and more rapidly in HUVEC treated cells. Here, a rapid increased interaction between erythrocytes and endothelial cells and subsequent deleterious erythrophagocytosis phenomenon were observed following glycated erythrocyte incubations.
The cell impedance results highlight an early apoptotic process appearing in the first 24 h of incubation with glycated erythrocytes. When glycated erythrocytes rapidly induced endothelial cell apoptosis, cell mortality was detected after 24 h incubation. In our static model, the cytotoxicity observed may be due to the progressive sedimentation of erythrocytes on the endothelial cell layer, impairing an optimal access to oxygen. This prolonged contact between erythrocytes and endothelial cells mimics conditions that can be encountered in the intraplaque neovessel where blood flow is minimal [
15].
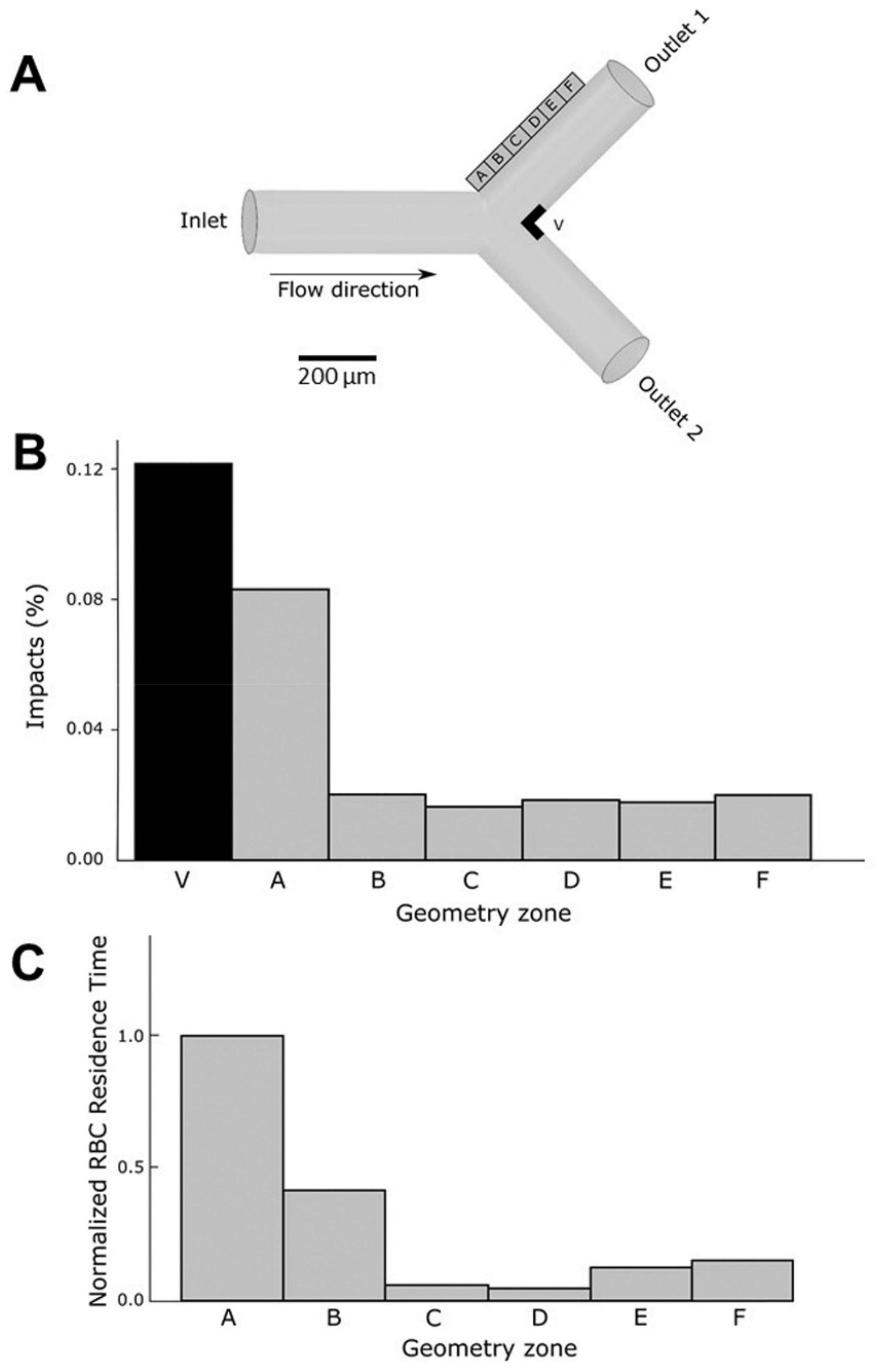
Still in an attempt to mimic conditions that can be encountered in vivo, effects of native or glycated erythrocytes on endothelial cells were tested under flow dynamic conditions. Therefore, we designed a flow assay to test the effects of our erythrocyte preparations on human endothelial cells covering the inner surface of a biomimetic channel, under flow conditions [
50]. To demonstrate the possible influence of hemodynamics, a channel mimicking a Y-shape bifurcation was designed. Compared to the main straight channel, bifurcations promote higher shear stress changes that can impact endothelial cell response [
51,
52]. In these experimental conditions, glycated, but not native, erythrocytes induced cell detachment from the inner surface of the channel after 5 h of flow exposure. Interestingly, endothelial cell detachment was localized at specific points of the bifurcation, but not in the main straight segment of the channel. The outer sides and the carina of the bifurcation are points of significant wall shear stress, making them particularly susceptible to circulating compounds [
51,
53]. To quantify the effects of circulating erythrocytes on the endothelial cells at the walls of the channel, in terms of number of impacts and time of residence, a numerical simulation approach was implemented. This simulation in silico reproduced the conditions of our in vitro flow assay. The results were consistent with fluid behavior at the bifurcations. These types of geometries are characterized by the presence of low-velocity zones at the outer wall [
51]. These low velocity values allow erythrocytes to interact for longer periods with the endothelial cells at the wall surfaces. At the carina level, the calculated number of impacts of erythrocytes was rather higher than in the rest of the bifurcation. The long exposure together with the high impact number of glycated erythrocytes correlate with the endothelial damage extent observed at the specific points of the bifurcation. For the first time, our results highlighted here a deleterious impact of glycated erythrocytes at specific zones of artificial microvessels, the bifurcation, rendering them more prone to the occurrence of vascular complications.


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}