Effects of Brassinosteroids on Photosynthetic Performance and Nitrogen Metabolism in Pepper Seedlings under Chilling Stress



Abstract
1. Introduction
2. Materials and Methods
2.1. Growth Conditions and Treatment
2.2. Plant Growth and Biomass Measurement
2.3. Gas Exchange and Chlorophyll Fluorescence Measurement
2.4. Chlorophyll Content Determination
2.5. Determination and Histochemical Localization of H2O2, O2−· and Lipid Peroxidation
2.6. Determination of Nitrate Nitrogen and Ammonium Nitrogen Concentrations
2.7. Determination of Nitrogen Metabolism Enzyme Activity and Free Amino Acids
2.8. Statistical Analyses
3. Results
3.1. Effects of Exogenous EBR on the Plant Growth and Foliar Pigment Contents of Pepper Seedlings
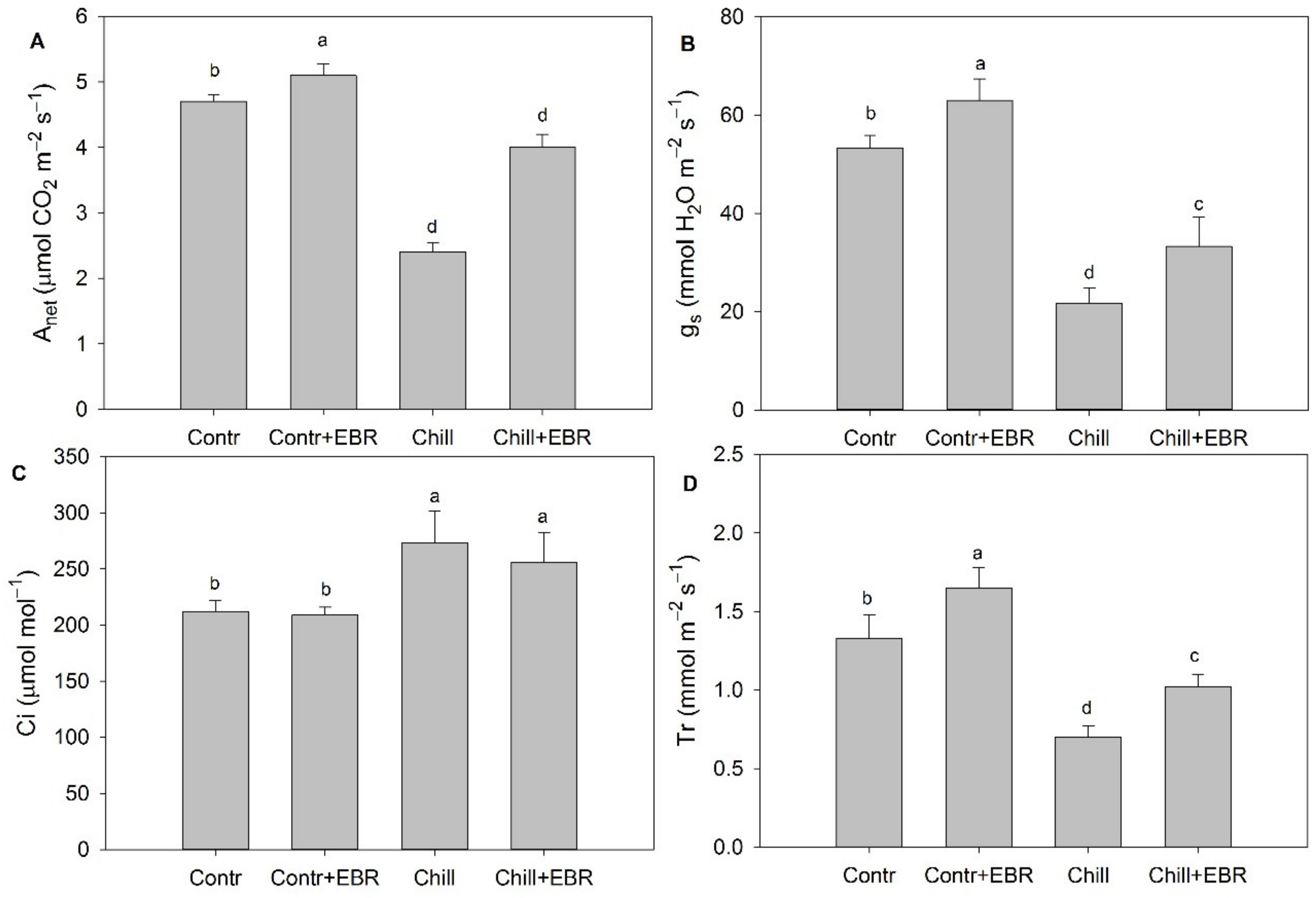
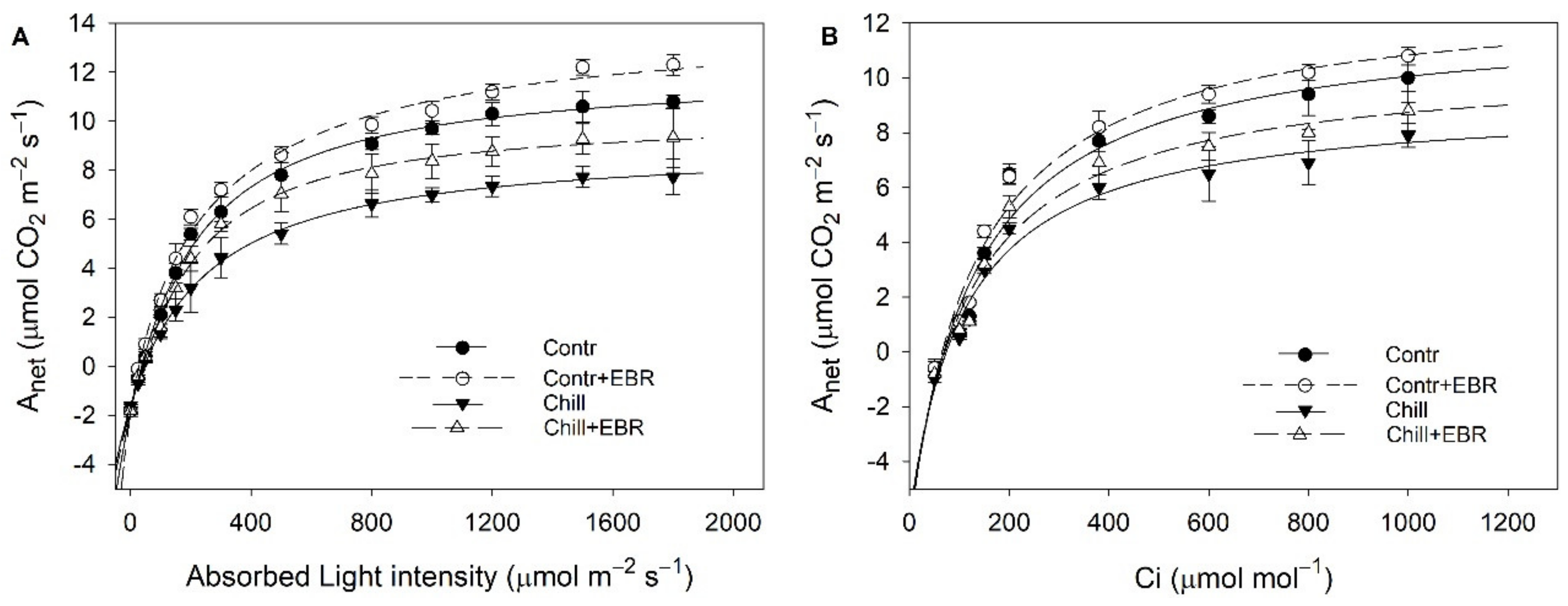
3.2. Exogenous EBR Enhances Photosynthesis of Pepper Leaves under Chilling Stress
3.3. Exogenous EBR Maintains High Photosynthesis Capability and Photosystem Balance in Pepper Leaves under Chilling Stress
3.4. Exogenous EBR Reduces ROS Accumulation and Mitigates Lipid Peroxidation in Pepper Leaves under Chilling Stress
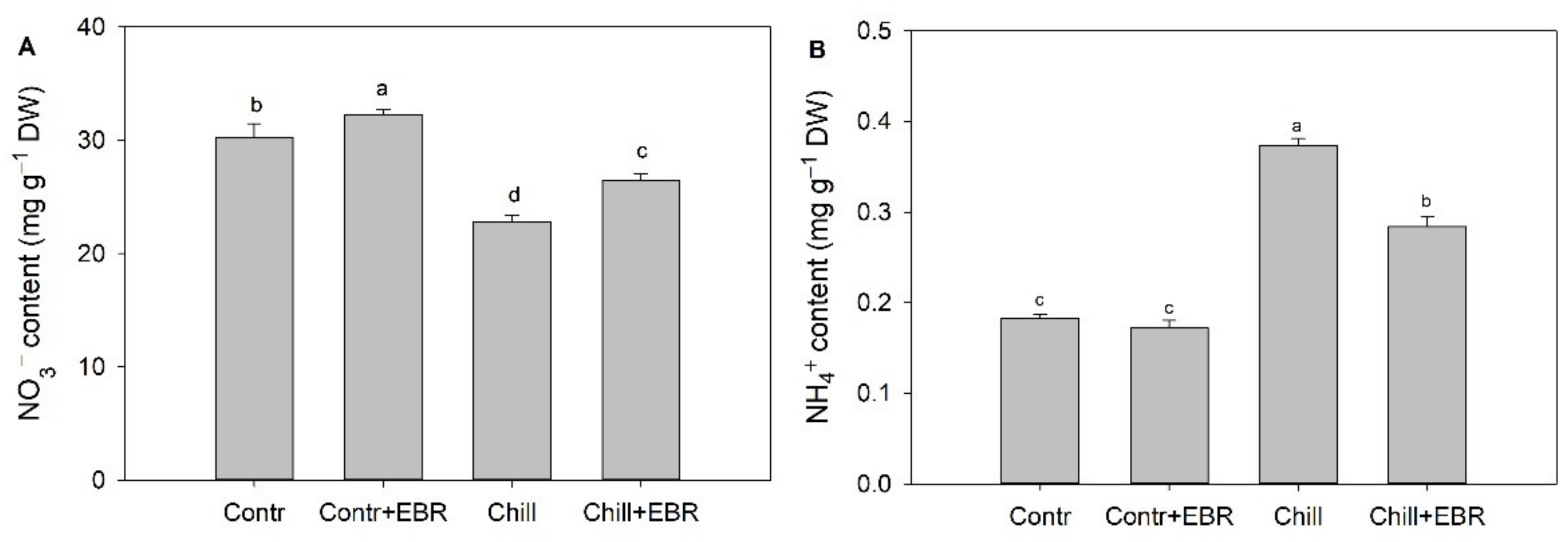
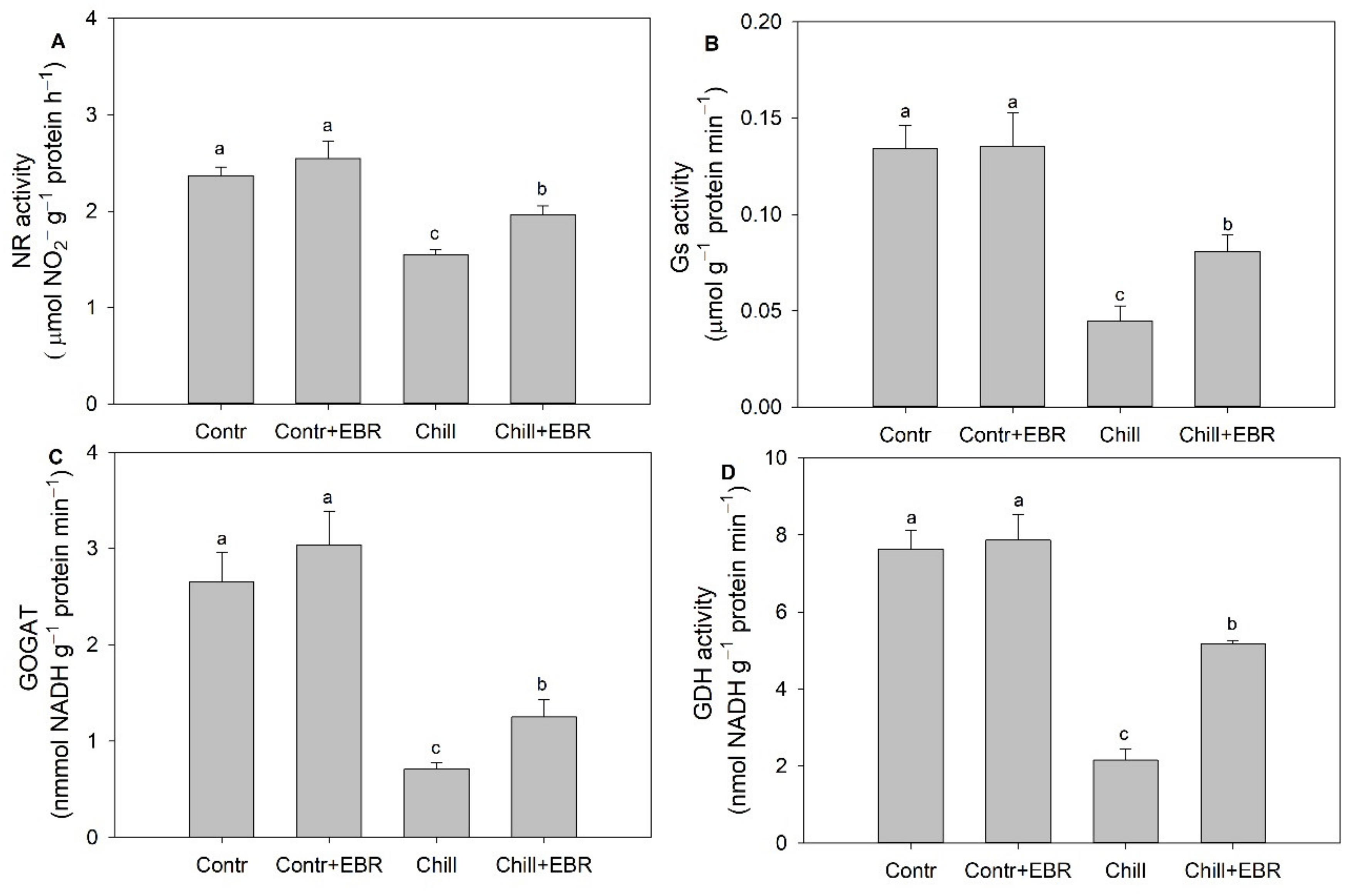
3.5. Exogenous EBR Promotes Nitrogen Metabolism via Increasing the Activities of NR, GS, GOGAT, and GDH in the Leaves of Pepper Seedlings
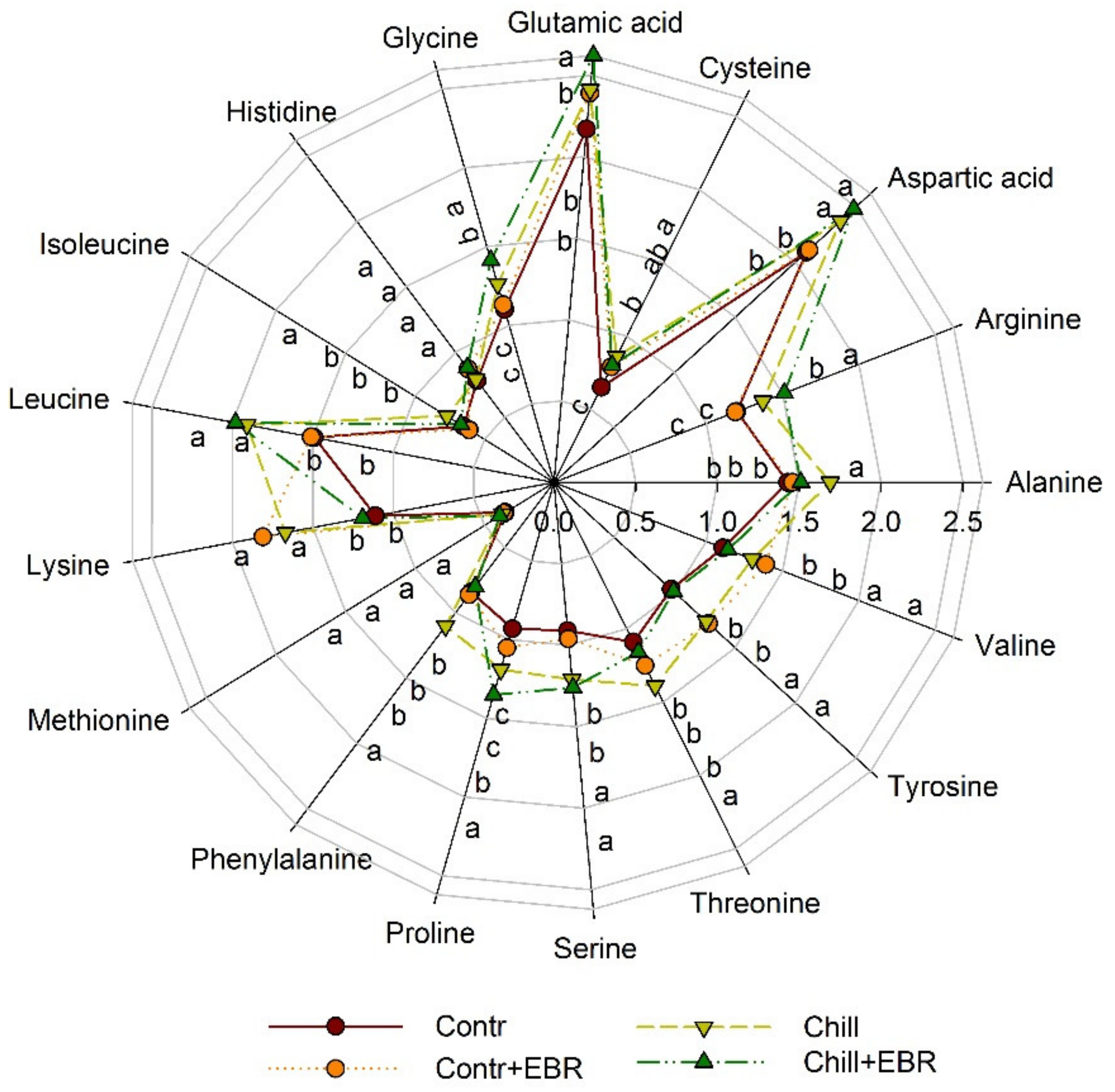
3.6. Exogenous EBR Results in an Increase in Amino Acid Accumulation in the Leaves of Pepper Seedlings under Chilling Stress
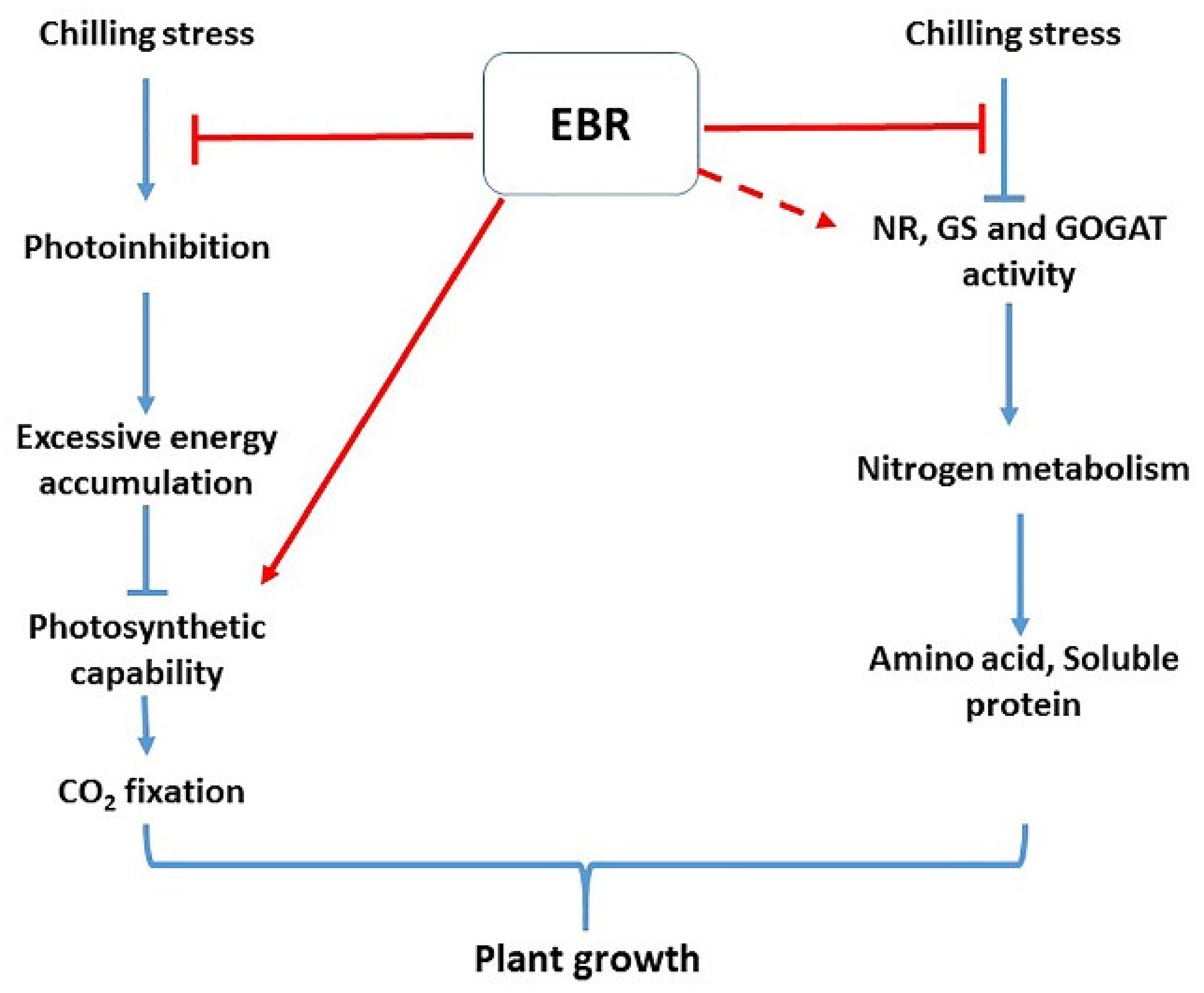
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Wang, P.; Wang, L.; Sun, G.; Zhao, J.; Zhang, H.; Du, N. The influence of facility agriculture production on phthalate esters distribution in black soils of northeast China. China. Sci. Total Environ. 2015, 506, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Yu, J.; Chen, B.; Feng, Z.; Li, J.; Zhao, C.; Lyu, J.; Hu, L.; Gan, Y.; Siddique, K.H. Facility cultivation systems “设施农业”: A Chinese model for the planet. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2017; Volume 145, pp. 1–42. [Google Scholar]
- Dhankher, O.P.; Foyer, C.H. Climate resilient crops for improving global food security and safety. Plant Cell Environ. 2018, 41, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Bilska, A.; Sowiński, P. Closure of plasmodesmata in maize (Zea mays) at low temperature: A new mechanism for inhibition of photosynthesis. Ann. Bot. 2010, 106, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Airaki, M.; Leterrier, M.; Mateos, R.M.; Valderrama, R.; Chaki, M.; Barroso, J.B.; Del Rio, L.A.; Palma, J.M.; Corpas, F.J. Metabolism of reactive oxygen species and reactive nitrogen species in pepper (Capsicum annuum L.) plants under low temperature stress. Plant Cell Environ. 2012, 35, 281–295. [Google Scholar] [CrossRef]
- Shu, S.; Tang, Y.; Yuan, Y.; Sun, J.; Zhong, M.; Guo, S. The role of 24-epibrassinolide in the regulation of photosynthetic characteristics and nitrogen metabolism of tomato seedlings under a combined low temperature and weak light stress. Plant Physiol. Biochem. 2016, 107, 344–353. [Google Scholar] [CrossRef]
- Khripach, V.; Zhabinskii, V.; de Groot, A. Twenty years of brassinosteroids: Steroidal plant hormones warrant better crops for the XXI century. Ann. Bot. 2000, 86, 441–447. [Google Scholar] [CrossRef]
- Beevers, L.; Hageman, R. Nitrate reduction in higher plants. Annu. Rev. Plant Physiol. 1969, 20, 495–522. [Google Scholar] [CrossRef]
- Giagnoni, L.; Pastorelli, R.; Mocali, S.; Arenella, M.; Nannipieri, P.; Renella, G. Availability of different nitrogen forms changes the microbial communities and enzyme activities in the rhizosphere of maize lines with different nitrogen use efficiency. Appl. Soil Ecol. 2016, 98, 30–38. [Google Scholar] [CrossRef]
- Chow, F.; Pedersén, M.; Oliveira, M.C. Modulation of nitrate reductase activity by photosynthetic electron transport chain and nitric oxide balance in the red macroalga Gracilaria chilensis (Gracilariales, Rhodophyta). J. Appl. Phycol. 2013, 25, 1847–1853. [Google Scholar] [CrossRef]
- Suzuki, N.; Miller, G.; Morales, J.; Shulaev, V.; Torres, M.A.; Mittler, R. Respiratory burst oxidases: The engines of ROS signaling. Curr. Opin. Plant Biol. 2011, 14, 691–699. [Google Scholar] [CrossRef]
- Gapper, C.; Dolan, L. Control of plant development by reactive oxygen species. Plant Physiol. 2006, 141, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Gouia, H.; Suzuki, A.; Brulfert, J.; Ghorbal, M.H. Effects of cadmium on the coordination of nitrogen and carbon metabolism in bean seedlings. J. Plant Physiol. 2003, 160, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Khana, T.A.; Yusufb, M.; Ahmadc, A.; Zoobia, B.; Saeede, T.; Fariduddinf, Q.; Hayatf, S.; Mocka, H.; Wu, T.Q. Proteomic and physiological assessment of stress sensitive and tolerant variety of tomato treated with brassinosteroids and hydrogen peroxide under low-temperature stress. Food Chem. 2019, 289, 500–511. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.Q.; Huang, L.F.; Hu, W.H.; Zhou, Y.H.; Mao, W.H.; Ye, S.F.; Nogués, S. A role for brassinosteroids in the regulation of photosynthesis in Cucumis sativus. J. Exp. Bot. 2004, 55, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.H.; Yan, X.H.; Xiao, Y.A.; Zeng, J.J.; Qi, H.J.; Ogweno, J.O. 24-Epibrassinosteroid alleviate drought-induced inhibition of photosynthesis in Capsicum annuum. Sci. Hortic. 2013, 150, 232–237. [Google Scholar] [CrossRef]
- Ali, Q.; Ashraf, M. Modulation of growth, photosynthetic capacity and water relations in salt stressed wheat plants by exogenously applied 24-epibrassinolide. Plant Growth Regul. 2008, 56, 107–116. [Google Scholar] [CrossRef]
- Fang, P.; Yan, M.; Chi, C.; Wang, M.; Zhou, Y.; Zhou, J.; Shi, K.; Xia, X.; Foyer, C.H.; Yu, J. Brassinosteroids act as a positive regulator of photoprotection in response to chilling stress. Plant Physiol. 2019, 180, 2061–2076. [Google Scholar] [CrossRef]
- Li, J.; Yang, P.; Gan, Y.T.; Yu, J.H.; Xie, J.M. Brassinosteroid alleviates chilling-induced oxidative stress in pepper by enhancing antioxidation systems and maintenance of photosystem II. Acta Physiol. Plant. 2015, 37, 222. [Google Scholar] [CrossRef]
- Liu, Y.; Jiang, H.; Zhao, Z.; An, L. Abscisic acid is involved in brassinosteroids-induced chilling tolerance in the suspension cultured cells from Chorispora bungeana. J. Plant Physiol. 2011, 168, 853–862. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, Z.; Si, J.; Di, C.; Han, J.; An, L. Brassinosteroids alleviate chilling-induced oxidative damage by enhancing antioxidant defense system in suspension cultured cells of Chorispora bungeana. Plant Growth Regul. 2009, 59, 207–214. [Google Scholar] [CrossRef]
- Chen, X.B.; Yao, Q.F.; Gao, X.H.; Jiang, C.F.; Harberd Nicholas, P.; Fu, X.D. Shoot-to-root mobile transcription factor HY5 coordinates plant carbon and nitrogen acquisition. Curr. Biol. 2016, 26, 640–646. [Google Scholar] [CrossRef]
- Li, J.; Yang, P.; Xie, J.; Yu, J. Effects of 24-epibrassinolide on growth and antioxidant enzymes system in pepper roots under chilling stress. J. Nucl. Agric. Sci. 2015, 29, 1001–1008. [Google Scholar]
- Hu, W.; Wu, Y.; Zeng, J.; He, L.; Zeng, Q. Chill-induced inhibition of photosynthesis was alleviated by 24-epibrassinolide pretreatment in cucumber during chilling and subsequent recovery. Photosynthetica 2010, 48, 537–544. [Google Scholar] [CrossRef]
- Dickson, A.; Leaf, A.L.; Hosner, J.F. Quality appraisal of white spruce and white pine seedling stock in nurseries. For. Chron. 1960, 36, 10–13. [Google Scholar] [CrossRef]
- Thornley, J.H. Mathematical Models in Plant Physiology; Academic Press (Inc.) Ltd.: London, UK, 1976. [Google Scholar]
- Trouwborst, G.; Hogewoning, S.W.; Harbinson, J.; van Ieperen, W. Photosynthetic acclimation in relation to nitrogen allocation in cucumber leaves in response to changes in irradiance. Physiol. Plant. 2011, 142, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.V.; von Caemmerer, S.V.; Berry, J. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, T.D.; Bernacchi, C.J.; Farquhar, G.D.; Singsaas, E.L. Fitting photosynthetic carbon dioxide response curves for C3 leaves. Plant Cell Environ. 2007, 30, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of Q A redox state and excitation energy fluxes. Photosynth. Res. 2004, 79, 209. [Google Scholar] [CrossRef]
- Hussain, M.I.; Reigosa, M.J. Allelochemical stress inhibits growth, leaf water relations, PSII photochemistry, non-photochemical fluorescence quenching, and heat energy dissipation in three C3 perennial species. J. Exp. Bot. 2011, 62, 4533–4545. [Google Scholar] [CrossRef]
- Bian, Z.; Yang, Q.; Li, T.; Cheng, R.; Barnett, Y.; Lu, C. Study of the beneficial effects of green light on lettuce grown under short-term continuous red and blue light-emitting diodes. Physiol. Plant. 2018, 164, 226–240. [Google Scholar] [CrossRef] [PubMed]
- Li, X.G.; Meng, Q.W.; Jiang, G.Q.; Zou, Q. The susceptibility of cucumber and sweet pepper to chilling under low irradiance is related to energy dissipation and water-water cycle. Photosynthetica 2003, 41, 259–265. [Google Scholar] [CrossRef]
- Bian, Z.H.; Cheng, R.F.; Yang, Q.C.; Wang, J.; Lu, C. Continuous light from red, blue, and green light-emitting diodes reduces nitrate content and enhances phytochemical concentrations and antioxidant capacity in lettuce. J. Am. Soc. Hortic. Sci. 2016, 141, 186–195. [Google Scholar] [CrossRef]
- Yang, F.; Wang, Y.; Miao, L.F. Comparative physiological and proteomic responses to drought stress in two poplar species originating from different altitudes. Physiol. Plant. 2010, 139, 388–400. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, A.; Baker, D. The effect of water stress on the activities of key regulatory enzymes of the sucrose to starch pathway in wheat. Plant Growth Regul. 2001, 35, 81–91. [Google Scholar] [CrossRef]
- Elstner, E.F.; Heupel, A. Inhibition of nitrite formation from hydroxylammoniumchloride: A simple assay for superoxide dismutase. Anal. Biochem. 1976, 70, 616–620. [Google Scholar] [CrossRef]
- Khokon, M.A.R.; Okuma, E.; Hossain, M.A.; Munemasa, S.; Uraji, M.; Nakamura, Y.; Mori, I.C.; Murata, Y. Involvement of extracellular oxidative burst in salicylic acid-induced stomatal closure in Arabidopsis. Plant Cell Environ. 2011, 34, 434–443. [Google Scholar] [CrossRef]
- Pompella, A.; Maellaro, E.; Casini, A.; Comporti, M. Histochemical detection of lipid peroxidation in the liver of bromobenzene-poisoned mice. Am. J. Pathol. 1987, 129, 295. [Google Scholar]
- He, X.; Mulvaney, R.L.; Banwart, W. A rapid method for total nitrogen analysis using microwave digestion. Soil Sci. Soc. Am. J. 1990, 54, 1625–1629. [Google Scholar] [CrossRef]
- Solorzano, L. Determination of ammonia in natural water by the phenolhypochlorite method Limnol. Oceanography 1969, 14, 799–801. [Google Scholar]
- Gangwar, S.; Singh, V.P. Indole acetic acid differently changes growth and nitrogen metabolism in Pisum sativum L. seedlings under chromium (VI) phytotoxicity: Implication of oxidative stress. Sci. Hortic. 2011, 129, 321–328. [Google Scholar] [CrossRef]
- Jiao, D.; Huang, X.; Li, X.; Chi, W.; Kuang, T.; Zhang, Q.; Ku, M.S.; Cho, D. Photosynthetic characteristics and tolerance to photo-oxidation of transgenic rice expressing C4 photosynthesis enzymes. Photosynth. Res. 2002, 72, 85–93. [Google Scholar] [CrossRef]
- Shapiro, B.; Stadtman, E. The regulation of glutamine synthesis in microorganisms. Annu. Rev. Microbiol. 1970, 24, 501–524. [Google Scholar] [CrossRef]
- Majerowicz, N.; Kerbauy, G.; Nievola, C.; Suzuki, R. Growth and nitrogen metabolism of Catasetum fimbriatum (Orchidaceae) grown with different nitrogen sources. Environ. Exp. Bot. 2000, 44, 195–206. [Google Scholar] [CrossRef]
- Aurisano, N.; Bertani, A.; Reggiani, R. Involvement of calcium and calmodulin in protein and amino acid metabolism in rice roots under anoxia. Plant Cell Physiol. 1995, 36, 1525–1529. [Google Scholar] [CrossRef]
- Bajguz, A.; Hayat, S. Effects of brassinosteroids on the plant responses to environmental stresses. Plant Physiol. Biochem. 2009, 47, 1–8. [Google Scholar] [CrossRef]
- Li, J.; Yang, P.; Kang, J.; Gan, Y.T.; Yu, J.H.; Calderón-Urrea, A.; Lyu, J.; Zhang, G.B.; Feng, Z.; Xie, J.M. Transcriptome analysis of pepper revealed a role of 24-epibrassinolide in response to chilling. Front. Plant Sci. 2016, 7, 1281. [Google Scholar] [CrossRef]
- Ogweno, J.O.; Song, X.S.; Shi, K.; Hu, W.H.; Mao, W.H.; Zhou, Y.H.; Salvador, N. Brassinosteroids alleviate heat-induced inhibition of photosynthesis by increasing carboxylation efficiency and enhancing antioxidant systems in Lycopersicon esculentum. J. Plant Growth Regul. 2008, 27, 49–57. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Xia, X.J.; Li, X.; Shi, K.; Yu, J.Q.; Zhou, Y.H. Role of Brassinosteroid in Plant Adaptation to Abiotic Stresses and its Interplay with Other Hormones. Curr. Protein Pept. Sci. 2015, 16, 462–473. [Google Scholar] [CrossRef]
- Almansa, E.M.; Espín, A.; Chica, R.M.; Lao, M.T. Nutritional response of seedling tomato plants under different lighting treatments. J. Plant Nutr. 2017, 40, 467–475. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed]
- Nixon, P.J.; Michoux, F.; Yu, J.; Boehm, M.; Komenda, J. Recent advances in understanding the assembly and repair of photosystem II. Ann. Bot. 2010, 106, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Velez-Ramirez, A.I.; van Ieperen, W.; Vreugdenhil, D.; Millenaar, F.F. Plants under continuous light. Trends Plant Sci. 2011, 16, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.J.; Yu, L.; Wang, L.P.; Guo, S.R. Bottle gourd root stock-grafting promotes photosynthesis by regulating the stomata and non-stomata performances in leaves of watermelon seedlings under NaCl stress. Plant Physiol. 2015, 186, 50–58. [Google Scholar] [CrossRef]
- Xiong, D.L.; Douthe, C.; Flexas, J. Differential coordination of stomatal conductance, mesophyll conductance and leaf hydraulic conductance in response to changing light across species. Plant Cell Environ. 2018, 41, 436–450. [Google Scholar] [CrossRef]
- Zait, Y.; Shtein, I.; Schwartz, A. Long-Term Acclimation to Different Stress Types: Revealing Tradeoffs between Mesophyll and Stomatal Conductance. Available online: https://www.biorxiv.org/content/10.1101/340786v1.full (accessed on 11 November 2019).
- Shahbaz, M.; Ashraf, M.; Athar, H.R. Does exogenous application of 24-epibrassinolide ameliorate salt induced growth inhibition in wheat (Triticum aestivum L.). Plant Growth Regul. 2008, 55, 51–64. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, J.J.; Wang, K.X.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Yu, J.Q.; Zhou, J. BZR1 mediates brassinosteroid-induced autophagy and nitrogen starvation in tomato. Plant Physiol. 2019, 179, 671–685. [Google Scholar] [CrossRef]
- Ma, Y.; Guo, S.; Shan, X.; Sun, J.; Wang, L.; Wang, Y.; Shu, S. Effect of exogenous 24-epibrassinolide on nitrogen assimilation of cucumber seedlings under hypoxia stress. J. Nanjing Agric. Univ. 2015, 38, 538–545. [Google Scholar]
- Diab, H.; Limami, A.M. Metabolism upon Hypoxia Stress and Recovery: Roles of Alanine Aminotransferase (AlaAT) and Glutamate Dehydrogenase (GDH) Alternative route for nitrogen assimilation in higher plants. Plants 2016, 5, 25. [Google Scholar] [CrossRef]
- Masclaux-Daubresse, C.; Reisdorf-Cren, M.; Pageau, K.; Lelandais, M.; Grandjean, O.; Kronenberger, J.; Valadier, M.H.; Feraud, M.; Jouglet, T.; Suzuki, A. Glutamine synthetase-glutamate synthase pathway and glutamate dehydrogenase play distinct roles in the sink-source nitrogen cycle in tobacco. Plant Physiol. 2006, 140, 444–456. [Google Scholar] [CrossRef]
- Liu, L.; Ji, H.; An, J.; Shi, K.; Ma, J.; Liu, B.; Tang, L.; Cao, W.; Zhu, Y. Response of biomass accumulation in wheat to low-temperature stress at jointing and booting stages. Environ. Exp. Bot. 2019, 157, 46–57. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Vardhini, B.V.; Rao, S.S.R. Amelioration of osmotic stress by brassinosteroids on seed germination and seedling growth of three varieties of sorghum. Plant Growth Regul. 2003, 41, 25–31. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Plant Height (cm) | Stem Diameter (cm) | Fresh Weight (FW, g) | Dry Weight (DW, g) | Index of Seedling Quality (QI) |
|---|---|---|---|---|---|
| Contr | 11.80 ± 0.20 a | 2.45 ± 0.05 b | 2.07 ± 0.13 a | 0.22 ± 0.01 a | 0.045 ± 0.004 b |
| Contr + EBR | 12.23 ± 0.25 a | 2.77 ± 0.02 a | 2.34 ± 0.31 a | 0.24 ± 0.05 a | 0.057 ± 0.003 a |
| Chill | 9.03 ± 0.46 d | 2.22 ± 0.07 c | 0.96 ± 0.18 c | 0.09 ± 0.02 c | 0.022 ± 0.007 d |
| Chill + EBR | 10.60 ± 0.26 c | 2.36 ± 0.02 b | 1.67 ± 0.17 b | 0.16 ± 0.02 d | 0.036 ± 0.002 c |
| Treatment | Chl a (mg g−1 FW) | Chl b (mg g−1 FW) | Chl a + b (mg g−1 FW) | Chl a/b | Carotenoids (mg g−1 FW) |
|---|---|---|---|---|---|
| Contr | 1.17 ± 0.0713 a | 0.28 ± 0.0406 ab | 1.48 ± 0.3041 a | 4.22 ± 0.8569 a | 0.28 ± 0.0180 ab |
| Contr + EBR | 1.20 ± 0.1541 a | 0.33 ± 0.0224 a | 1.50 ± 0.0936 a | 3.64 ± 0.4828 a | 0.30 ± 0.0197 a |
| Chill | 0.82 ± 0.0378 b | 0.21 ± 0.0053 c | 1.05 ± 0.0613 b | 3.70 ± 0.2729 a | 0.22 ± 0.0053 c |
| Chill + EBR | 1.12 ± 0.0343 a | 0.27 ± 0.0127 b | 1.40 ± 0.0499 a | 4.58 ± 0.8391 a | 0.27 ± 0.0127 b |
| Contr | Contr + EBR | Chill | Chill + EBR | |
|---|---|---|---|---|
| Measured parameters | ||||
| Fv/Fm | 0.84 ± 0.021 a | 0.85 ± 0.017 a | 0.79 ± 0.010 c | 0.81 ± 0.006 b |
| ΦPSII | 0.74 ± 0.009 a | 0.74 ± 0.008 a | 0.58 ± 0.005 c | 0.66 ± 0.004 b |
| qP | 0.97 ± 0.002 a | 0.97 ± 0.009 a | 0.83 ± 0.013 c | 0.88 ± 0.014 b |
| Fv′/Fm′ | 0.76 ± 0.009 a | 0.76 ± 0.003 a | 0.70 ± 0.008 c | 0.73 ± 0.006 b |
| E | 0.021 ± 0.001 c | 0.022 ± 0.007 c | 0.12 ± 0.010 a | 0.092 ± 0.012 b |
| β/α − 1 | 0.028 ± 0.002 c | 0.029 ± 0.009 c | 0.20 ± 0.019 a | 0.14 ± 0.018 b |
| Fitting parameters | ||||
| Amax | 14.0 ± 0.25 b | 15.7 ± 0.37 a | 10.6 ± 0.15 d | 11.9 ± 0.10 c |
| Rd | 1.95 ± 12.9 ab | 1.98 ± 4.65 a | 1.38 ± 6.22 c | 1.60 ± 0.67 bc |
| Jmax | 82.3 ± 0.83 a | 85.1 ± 1.33 a | 71.1 ± 3.42 c | 76.2 ± 1.02 b |
| Vmax | 47.5 ± 0.87 b | 51.4 ± 0.87 a | 38.8 ± 1.32 d | 44.3 ± 0.76 c |
| Treatment. | H2O2 (μmol g−1, FW) | O2−· (μmol min−1 g−1, FW) | MDA (μmol g−1, FW) |
|---|---|---|---|
| Contr | 0.34 ± 0.017 c | 0.22 ± 0.011 c | 0.32 ± 0.009 c |
| Contr + EBR | 0.33 ± 0.021 c | 0.21 ± 0.025 c | 0.32 ± 0.008 c |
| Chill | 0.78 ± 0.025 a | 0.40 ± 0.027 a | 0.43 ± 0.011 a |
| Chill + EBR | 0.59 ± 0.010 b | 0.31 ± 0.017 b | 0.36 ± 0.005 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, P.; Wang, Y.; Li, J.; Bian, Z. Effects of Brassinosteroids on Photosynthetic Performance and Nitrogen Metabolism in Pepper Seedlings under Chilling Stress. Agronomy 2019, 9, 839. https://doi.org/10.3390/agronomy9120839
Yang P, Wang Y, Li J, Bian Z. Effects of Brassinosteroids on Photosynthetic Performance and Nitrogen Metabolism in Pepper Seedlings under Chilling Stress. Agronomy. 2019; 9(12):839. https://doi.org/10.3390/agronomy9120839
Chicago/Turabian StyleYang, Ping, Yu Wang, Jie Li, and Zhonghua Bian. 2019. "Effects of Brassinosteroids on Photosynthetic Performance and Nitrogen Metabolism in Pepper Seedlings under Chilling Stress" Agronomy 9, no. 12: 839. https://doi.org/10.3390/agronomy9120839
APA StyleYang, P., Wang, Y., Li, J., & Bian, Z. (2019). Effects of Brassinosteroids on Photosynthetic Performance and Nitrogen Metabolism in Pepper Seedlings under Chilling Stress. Agronomy, 9(12), 839. https://doi.org/10.3390/agronomy9120839