Abstract
Italian ryegrass is one of the most important temperate forage grasses on a global basis. Improvement of both dry matter yield and quality of herbage have been major objectives of pasture grass breeding over the last century. F1 and F2 progeny sets derived from controlled pair-crosses between selected Italian ryegrass genotypes have been evaluated for yield and nutritive quality under field conditions. Linear regression of the performance of F1 families under sward conditions on parental genotype means in a spaced plant trial was significant for quality characteristics, but not for herbage yield. This result suggests that phenotypic selection of individual plants from spaced plant nursery is feasible for improvement of nutritive quality traits, but not for yield. The presence of significant heterosis within F1 populations was demonstrated by reduced herbage production in subsequent F2 populations (generated by one cycle of full-sib mating), an up to 22.1% total herbage yield in fresh weight, and a 30.5% survival rate at the end of the second reproductive cycle. Potential optimal crosses for exploiting such heterosis are discussed, based on construction and the inter-mating of complementary parental pools, for the implementation of a novel F1 hybrid production strategy.
1. Introduction
Italian ryegrass (L. multiflorum Lam.) is one of the most important temperate forage grasses on a global basis, providing a high quality and cost-effective option for the supply of forage during the winter and spring seasons. Forage productivity traits such as yield, quality, and persistence have been the core targets of modern ryegrass breeding [1]. Improvements have been made in yield, digestibility, water soluble carbohydrate content for perennial, and Italian ryegrasses [2,3]. However, the magnitude of genetic gains made for dry matter production have been small relative to gains in cereal crops [4,5]. In some instances, no significant changes of forage production have been observed [6].
Despite the widespread use of hybrid cultivars (based on intercrossing of inbred lines) for crop species such as maize and rice, most varieties of forage crops are synthetic populations that do not express maximum heterosis for important agronomic traits such as yield [7]. Three types of methods for first filial generation (F1) hybrid production have been proposed as a means to capture and exploit heterosis in forage grasses. These include varietal hybrid production [8,9,10,11], the use of cytoplasmic male sterility (CMS) [12], and a restriction of allelic diversity at self-incompatibility loci by selfing [13,14] or by genotypic classification and selection [15]. The theoretical expectation from crossing of perennial ryegrass lines of divergent geographic origin is that 50% of progeny individuals would be intervarietal F1 hybrids and the remainder would arise from intra-varietal mating, although several factors may produce bias in the proportions of hybrids and non-hybrids [8]. An empirical study of heterosis between 15 varieties of perennial ryegrass was conducted in spaced [9], simulated sward [10], and sown-sward [11] settings. Heterosis was highly specific to the test conditions, varying from up to 31% in spaced trial to only 3.6% in sown sward to the higher parent for herbage yield. Pure hybrids obtained by experimental pair-crossing outperformed the corresponding intervarietal hybrid by 8.4% in terms of yield [8].
Inbred lines were developed from several cycles of self-pollination in perennial ryegrass [14], and crosses between such lines identified a range of heterosis values, from negative to approximately. 20% of the mid-parental value. The author suggested that only the most vigorous individuals within inbred lines are able to survive under sward conditions, thus increasing the parental means and reducing the level of heterosis. Nonetheless, slightly lower levels of heterosis (up to 13% for yield) were observed in diallele crosses between eight distinct populations with distant geographic origins in Europe [16]. Considering the low levels of genetic gain that have been characteristic of ryegrass breeding programs based on individual or family-based phenotypic selection [2,3,5], access to even modest levels of heterosis would prove valuable for ryegrass improvement.
Recently, a F1 hybrid breeding strategy for perennial ryegrass was developed, based on restriction of allelic diversity at the S and Z gametophytic self-incompatibility loci in complementary germplasm pools and crossing between pools (alleles) [15]. Since a similar self-incompatibility mechanism is active in Italian ryegrass, the same strategy would be applicable. From the perspective of seed production, Italian ryegrass may provide a larger potential market than perennial ryegrass due to more frequent sowing. If levels of heterosis in Italian ryegrass prove adequate, F1 hybrid production would be an economically viable strategy. However, Italian ryegrass has a comparatively narrow genetic base compared to perennial ryegrass [17], and evidence for heterosis is relatively lacking. As a consequence, a crucial first step would be to identify heterotic combinations between Italian ryegrass germplasm sources.
In a previous study, the extent of trait variation and broad-sense heritability was reported for yield, nutritive values, and morphological traits evaluated in a spaced plant nursery trial containing 960 genotypes of Italian ryegrass [18]. The plants were also analysed for trait-marker associations using single nucleotide polymorphism (SNP) genetic marker data. In the present paper, the results of selection from those genotypes and the evaluation of the first and second generations derived from bi-parental crosses are described. The objective of this study was to estimate narrow-sense heritability as an indicator of feasibility for spaced plant selection for improvement of yield and quality, and to quantify the presence of heterosis in F1 progeny sets and its subsequent erosion in succeeding generations.
2. Results
2.1. Field Evaluation of F1 Progeny Sets
Changes in yield and quality parameters were observed across multiple harvests (Figure S1). Yield from the first harvest exhibited the broadest range and highest average value, representing the establishment phase of growth in the first three months after planting. Yield gradually declined following harvest-2 and havest-3, and recovered during harvest-4 in November, when plants were developing flowering tillers. Growth slowed again for harvest-5 and havest-6, and reached the lowest point during harvest-7, corresponding to the aftermath regrowth in summer. Yield from harvest-8 (conducted in July) represented autumn recovery following a dry summer and was slightly higher on average than the yields from harvest-6 and harvest-7. Herbage quality also altered according to harvest. Water-soluable carbohydrate (WSC) content was lowest for harvest-3 and harvest-8, while crude protein (CP) content was highest for these two harvests. Acid detergent fibre (ADF) and netural detergent fibre (NDF) contents were high for harvest-4 and havest-5, while dry matter digestibility (DMD) was low for both harvests.
Simple linear regression of quality parameters for the F1 progeny sets for each harvest and mean quality parameters across all harvests on those of the parental genotype means resulted in regression coefficients of 0.32 (p < 0.001), 0.21 (p = 0.008), 0.34 (p < 0.001) , 0.23 (p = 0.002), and 0.26 (p = 0.029) for mean WSC, CP, NDF, ADF, and DMD, respectively (Table 1).

Table 1.
Regression of F1 family performances on the parental means for quality parameters (b: regression coefficient; S.E.: standard error; WSC: water-soluble carbohydrate; CP: crude protein; NDF: neutral detergent fibre; ADF: acid detergent fibre; DMD: dry matter digestibility).
Total yield from F1 swards was not significantly associated with the parental mean herbage yield from the spaced plant trial. Survival rate was highly correlated with fresh weight from harvest-8 (FW-8) (r = 0.70, p < 0.001), which was therefore taken as an indicator of persistence. Significant positive correlations were observed between parental mean plant height and F1 plot total dry matter yield in 2012 (r = 0.27, p = 0.0088). However, parental plant height was also negatively correlated with F1 plot persistence, as indicated by FW-8 (r = −0.22, p = 0.03). Genetic distance [17] between pairs of parental genotypes calculated on the basis of shared SNP markers was not correlated with the total herbage yield of the corresponding F1 family (r = 0.06, p = 0.57).
2.2. Field Evaluation of F2 Progeny Sets
Changes of yield and quality parameters across harvests in the sward trial of F2 families (Figure S2) showed a similar trend to that observed for the F1 sward trial. The first harvest showed the lowest ADF and NDF contents, and the highest DMD and WSC contents.
Residual maximum likelihood estimation (REML) analysis showed that different pair-wise combinations of parental genotypes produced significant effects on the yield and quality of offspring (p < 0.001). Identity of generation exerted a significant effect on herbage yield (p < 0.001), but not on quality. The total herbage yield of F1 populations was significantly higher than that of F2 populations (Figure 1A). The mean reduction of total herbage FW for F2, as compared to F1, populations was 3.96 kg, equivalent to 22.1% of the mean value across F1 families. For the individual family-harvest combinations, the corresponding value varied from negative in nature (such that higher yields were observed in the F2 sets) to reductions of up to 56.1%. There was no significant difference between the yield measurements for F2B and F2W populations, which suggested that selection for vigor within-family at an intensity of 20 in 100 plants did not significantly further improve yield. Some F1 families (1804 and 1836) were higher yielding than the best reference cultivars (Figure 1C). The total herbage yield of the best F1 family (1836) was significantly higher than Hulk (14.4%) and Tabu (17.3%), and slightly higher than LM299 (4.3%) but not significant. It was also observed that the reduction of F2 as compared to F1 yield was larger during the first year (25.5%) than the second year (19.3%). F1 populations also showed significantly higher survival rates than F2 populations (Figure 1B). The absolute reduction of survival rate was 12.3% (from 40.3% for F1 to 28.0% for F2 families), and the proportion of inbreeding depression was 30.5%. This value varied across different families from 16.7% (1901) to 54.5% (1812). Survival of some F1 families were superior to the reference cultivars (Figure 1 D). Within-family selection slightly increased the survival rate from 28.0% (for F2B) to 32.1% (for F2W), but the difference was not significant.
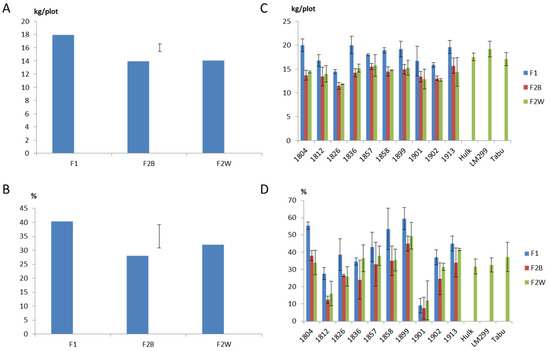
Figure 1.
(A) Predict mean of different generations on total fresh herbage yield including LSD bar at p = 0.05. (B) Predict mean of different generations on survival rate including LSD bar at p = 0.05. (C) Total fresh herbage yield of different generation family combinations and reference cultivars. (D) Survival rate of different generation family combinations and reference cultivars.
3. Discussion
3.1. Effectiveness of Phenotypic Selection from Spaced Plant Nursery
The observed seasonal changes of herbage yield and quality are consistent with previous reports [19]. The herbage yield of hybrid progeny in a sward setting was not significantly associated with parental mean herbage yield in a spaced plant trial. Current practices of phenotypic recurrent selection also involve selection from a spaced plant nursery, followed by testing progenies in sward conditions [20]. This raises the issue of whether parental genotypic performances in a spaced plant trial may be trusted to be correlated with the performance of the progeny in a sward setting. The results of the present study suggest that selection of high-yielding individual Italian ryegrass genotypes from a spaced plant nursery cannot guarantee high-yielding progeny populations in a competitive sward condition. Spaced plant evaluation was also found to be unpredictive of sward yield in tall fescue [21]. Nonetheless, the observed correlation of progeny yield with mean parental plant height indicates that indirect selection for yield may be achievable by direct selection for plant height, which displays a higher heritability value in Italian ryegrass. This correlation effect may be more relevant for Italian ryegrass than perennial ryegrass, as the former typically shows a more erect growth habit in which the vertical length of tillers will more accurately reflect total biomass. However, plant height was also found to be negatively correlated with persistence, such that any increases in yield would probably be achievable in the first growing season. Selection heavily and solely dependent on plant height would consequently not be ideal in the long term. Genotypic recurrent selection using full-sib or half-sib families [22] coupled with genomic selection [23] may hence be a necessary part of the breeding system if forage yield is the target.
The observed significant correlation between parental mean WSC content in a spaced plant trial and the WSC content of progeny under sward condition indicated that direct selection based on the former would be effective. The significant regression coefficient may arise because WSC content has never been the target of selection in the development histories of the particular varieties used in the present study. In such cases, there would be presumably a higher degree of genetic variability for the unselected character than for those traits (such as herbage yield) that have been subject to strong directional selection. This would ensure a broad range of WSC content in the regression analysis. In contrast, if the genetic variability was small and environmental effects were prominent, the regression analysis would probably not obtain significant results. Another possible explanation is that WSC content may be less sensitive to the inhibitory effects of plant neighbour interactions than a morphophysiological trait such as yield.
Based on these observations, phenotypic selection from spaced plant trials would be more effective for breeding of quality parameters than for yield. Since the quality parameters were affected by plant maturity, it would be important to measure at comparable stages of vegetative growth. In perennial grasses, selection is typically conducted on a single-plant basis and usually displays a realised heritability for digestibility of about 0.2 to 0.3 [24], in agreement with the results of the present study. Of the quality parameters, CP displayed lower heritability than for WSC content and NDF, consistent with reported sensitivity to environmental factors such as fertiliser applications [25].
3.2. Heterosis and Inbreeding Depression
In the present study, F1 families were generated from pair crosses between two unique genotypes. As a consequence, mean values for a parental population in a field setting could not be obtained, except by vegetative propagation to produce multiple ramets from the selected genotype, which is logistically unfeasible, so the performance of F1 families cannot be compared to those of parents. However, comparisons can be made between F1 and F2 progeny sets derived from a particular pair-wise genotypic combination in order to deduce the presence of heterosis in the F1 generation and erosion by inbreeding depression in subsequent generations. High levels of inbreeding depression in parental lines has been shown to be associated with heterosis in hybrids of other plant species [26]. The magnitude of the observed inbreeding depression in the present study was on average 22.1% for total fresh herbage yield, with a range from negative values to 56.1% for different family-harvest combinations. In other studies of ryegrass species, yield in Italian ryegrass declined by 6% from Syn1 to Syn2, and remained constant from Syn2 and Syn3 [27], while that of perennial ryegrass declined by 11% from Syn1 to Syn3 of five-parent synthetic [28]. Dry matter yield in tall fescue declined by 6% from the Syn1 to Syn2 generations of four-parent synthetic [29]. In smooth bromegrass (Bromus inermis L.), the average level of heterosis for herbage yield in diallele crosses between seven parents was 14% with a range of −4 to 39%, and the level of inbreeding depression based comparison of selfed progenies with parents was 18%–33% [30]. The difference between species may be attributable to the number of genotypes used for crossing and different mechanisms of genetic control.
Of the 10 different families obtained in the present study, 8 were derived from pair-crosses of genotypes from different cultivars. If they were assumed to be unrelated, the inbreeding coefficient would be 0 and 0.25 for F1 and F2 progeny sets, respectively. For the other two families (1857 and 1826), the two parental genotypes were obtained from the same cultivar (“Tabu” for 1857 and “Warrior” for 1826), and the two parents within each family, due to an origin from a single polycross, must by definition be to some extent related. “Tabu” was bred from a mass selection made from an 18-month old field trial of variety “Flanker” [31], while “Warrior” was bred from breeding pools obtained from various sources [32]. It is therefore likely that the two parental genotypes of family 1857 were more closely related than those of family 1826; consequently, the increase of inbreeding coefficient from the F1 to F2 generation for these two families will be less than 0.25, especially for 1857. A lesser degree of inbreeding depression would hence be expected in these two families. Results were consistent with this expectation, as the decline in total herbage yield for 1857 was 13.5%, the least among the 10 families. On average, heterosis increases as the genetic disparity of the parental stocks increases and interspecific crosses show greater heterosis than intraspecific crosses [33]. However, the extent of correlations between molecular marker-based genetic distance and heterosis are not conclusive. There are several prerequisites for positive correlations, such as high trait heritability and close marker linkage [34]. It is not a surprise that the present study revealed no correlation between yield in the F1 generation and genetic distance between the parents, since yield showed low heritability and random low-density distribution of markers used for diversity analysis. Cultivars of the interspecific hybrid between perennial and Italian ryegrass species (L. x boucheanum), which combine advantages from each species, are available in the commercial marketplace. However, they are not F1 hybrids, and any heterosis in the initial crosses would have been eroded through seed multiplication during the variety development process and so would not be expected to exploit the full potential of heterosis.
A 30.5% reduction in survival rate was seen in the F2B generations as compared to F1 generations across the various families, consistent with heterotic effects. Survival rate is a measure of persistence in forage grasses. It may be regarded as a fitness-related trait, which generally exhibit lower heritability values than morphological traits [35]. Nonetheless, heterosis for fitness-related traits is commonly observed [27]. Hybrid vigour for persistency in Italian ryegrass would be beneficial to farmers to allow the growth of more productive pastures for longer periods, with reducing re-sowing and establishment costs. Although inbreeding depression was observed for yield and persistence, no such effects were seen for quality parameters. Similarly, there is little evidence of heterosis for WSC content in perennial ryegrass [36] or for total soluble solid and crude protein content in sorghum [26]. Population hybrids (generated through controlled pair-crossing between varieties) of orchardgrass (Dactylis glomerata L.) exhibited higher mean phenotypic values than source cultivars for some trait-harvest combinations (such as increased CP, WSC, and decreased NDF); however, the magnitudes of difference were very small [37]. Based on these results, the feasibility of exploiting hybrids for improvement of quality traits in ryegrasses appears limited.
To date, very little information has been generated on the extent and prevalence of heterosis in Italian ryegrass. As the genetic base of Italian ryegrass is known to be narrow, it may be assumed that lower levels of differentiation between different germplasm pools are present, compared with the closely related species, perennial ryegrass. If so, a lesser extent of heterosis might be anticipated. However, the results of the present study suggest that the magnitude of heterosis in Italian ryegrass, for comparable traits, is no less than for perennial ryegrass. The significant heterosis observed in herbage yield and survival rate provided foundation data for the implementation of the F1 hybrid breeding strategy in Italian ryegrass. However, controlled pair-crossing between specific genotypes is not a feasible approach for commercial seed production, especially as any advance for seed multiplication by reproduction will cause inbreeding depression. As a consequence, the development and maintenance of divergent and compatible parental population pools will be critical for F1 hybrid breeding. Nonetheless, if an average of 20% yield advantages from F1 hybrids could be achieved in commercial production, the gain would be enormous compared with the less than 1% of annual genetic gain in yield that has been achieved via conventional breeding programs.
4. Materials and Methods
4.1. Selection of Individual Plants as Parents
A total of 960 genotypes from 7 cultivars/breeding lines (“Accelerate”, “AristocratII”, “Hulk”, “LM299”, “LM414”, “Tabu”, and “Warrior”) of Italian ryegrass were phenotypically assessed for herbage yield and quality in a spaced plant nursery trial with 4 replicates and genotyped with 384 SNP markers [18]. To conserve allelic diversity, an in-house R script was developed that selected a minimum core set of genotypes that contained all the alleles observed in the original population/nursery. This core set was made up of 29 individual plants, with representation from each of the cultivars/breeding lines. The core set was then expanded to 150 genotypes, weighted by phenotypic performance, i.e., herbage yield.
4.2. Generation of F1 Full Sib Families and Field Test of Performance
The core set of 29 plants, along with 21 of the most elite genotypes (herbage yield) were propagated into two clonal ramets by a division of tillers. Each replicate of these 50 genotypes was then pair-crossed randomly with one of the remaining 100 selected genotypes. A total of 100 pair-crosses were conducted in glasshouses with pollen-proof bags. Seeds of each full-sib family were bulk-collected from each parent. Seeds were cleaned and germinated in a glasshouse, and 94 of the families obtained more than 100 seedlings each. Along with the original reference cultivars/breeding lines (2–4 entries each), the F1 families were tested in a simulated sward setting during 2012–2013 with a total of 110 plots in a 10 (row) × 11 (column) layout. Each plot consisted of 100 plants in a 10 × 10 grid configuration. The distance between plots was 1 m, and the space between plants within a plot was 15 cm. The seedlings were transplanted into the field on the research farm of the Department of Economic Development, Transport, Jobs and Resources, Hamilton, Victoria, in May 2012. Apart from natural precipitation, irrigation was applied to the trial during the summer months (December, January, and February) in order to minimise plant death. Fertilizer “Grow Plus” (30 N, 10 P, 25 K, 10 S) was applied at a rate of 150 kg/ha on 18 April 2012 (before planting) and on 13 November 2012. A total of 8 harvests were conducted. The first 7 harvests were conducted every month from August 2012 onwards until February 2013 for production traits, and the last harvest was conducted in July 2013 for persistence. Fresh weight (FW) of each plot was recorded for every harvest, and herbage quality data was collected from 6 of the 8 harvests, apart from harvest-6 and harvest-7 which were performed in January and February 2013, respectively. When herbage quality data was collected, 200–300 g sub-samples from each plot were dried in a fan-forced oven at 60 °C, and dry matter yield was calculated. Herbage quality parameters including contents of WSC, CP, NDF, ADF, and DMD were assessed as described previously [38].
4.3. Generation of F2 Families and Field Test of Performance
Based on the performance of the F1 sward trial as described, 10 F1 families were selected for high yield, high WSC content, and good persistence (Table 2). Within each of the 10 F1 families, 20 plants were further selected based on visual assessment of vigour, and half of each selected plant was taken from the field and potted in a glasshouse in order to allow polycrossing (exclusive to a particular family) between extracted individuals (so providing the basis for between- and within-family selection, designated as F2W). The 10 selected in-field plots were also covered with pollen proof cages to generate F2 families based on inter-mating of members of the same F1 family (so providing the basis for between-family selection, designated as F2B). Seeds from these 20 polycrosses were harvested and cleaned, and used for in-field evaluation.
Table 2.
Selected 10 bi-parental F1 families.
A simulated sward trial was conducted in 2014–2016 to test the performance of the F2 families along with F1 progeny sets and reference cultivars. The trial entries included 10 F1 families, 10 F2B families, 10 F2W families, and the reference cultivars, “Tabu”, “LM299” and “Hulk”. The trial was basically designed as a split-plot design with family as whole plot and generation (F1, F2B, and F2W) as sub-plot, with 2 replicates. Reference cultivars were represented by 4 replicates in the trial. A total of 72 plots were organized in a 12 (row) × 6 (column) layout. Each of the plots contained 100 plants, and the space between plots and the distances between plants within a plot were the same as for the F1 sward trial. Seeds from each entry were germinated in seedling trays in the glasshouse, and seedlings were transplanted to the field in May 2014. Irrigation was applied for the trial in the first summer months in order to keep plants alive. The trial was fertilized six times with nitrogen fertilizer and “Grow Plus” at the same rate as that of the F1 trial at 2–3 month intervals, and a total of 12 harvests were conducted. The first 10 harvests were conducted every month starting from September 2014 to June 2015, and harvest-11 and harvest-12 were conducted in August and November 2015, respectively. Quality data were collected from harvests 1, 2, 7, 9, 10, and 11 following the same procedure as described in the F1 trial. Plant survival rate was recorded on 4 March 2016.
4.4. Statistical Analysis
Statistical analysis was carried out using GenStat [39]. Correlation coefficients between pairs of traits were calculated using the correlation command in GenStat. Narrow-sense heritability was estimated by simple linear regression of the F1 progeny on parental mean for each trait. The narrow-sense heritability for the corresponding traits is equal to the regression coefficient [40]. Significance testing of family and generation differences was conducted using REML. The level of inbreeding depression from comparison of the F1 and F2 generations was calculated as the deviation of the mean performance from the mean of the previous generation.
Supplementary Materials
The following are available online at www.mdpi.com/2073-4395/6/4/49/s1. Figure S1: Scatter plot of yield and quality across harvests from the F1 field trial (FW: Fresh weight (g); WSC: water-soluble carbohydrate (% dry matter); CP: crude protein (% dry matter); NDF: neutral detergent fibre (% dry matter); ADF: acid detergent fibre (% dry matter); DMD: dry matter digestibility (%)), Figure S2: Scatter plot of yield and quality across harvests from the F2 field trial (trait abbreviations and units see Figure S1).
Acknowledgments
The authors acknowledge financial support from the Victorian Department of Economic Development, Jobs, Transport and Resources, Geoffrey Gardiner Foundation, Meat and Livestock Australia and New Zealand Agriseeds, Christchurch, New Zealand, through the Dairy Futures Cooperative Research Centre.
Author Contributions
Noel O. I. Cogan and John W. Forster conceived the experiments. Junping Wang and Luke W. Pembleton designed and performed the experiments; Junping Wang analyzed the data; Junping Wang, Luke W. Pembleton, and John W. Forster prepared the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Stewart, A.; Hayes, R. Ryegrass breeding-balancing trait priorities. Irish J. Agric. Food Res. 2011, 50, 31–46. [Google Scholar]
- Chaves, B.; de Vliegher, A.; van Waes, J.; Carlier, L.; Marynissen, B. Change in agronomic performance of Lolium perenne and Lolium multiflorum varieties in the past 40 years based on date from Belgian VCU trials. Plant Breed. 2009, 128, 680–690. [Google Scholar] [CrossRef]
- Wilkins, P.W.; Lovatt, J.A. Gains in dry matter yield and herbage quality from breeding perennial ryegrass. Irish J. Agric. Food Res. 2011, 50, 23–30. [Google Scholar]
- Woodfield, D.R. Genetic improvement in New Zealand forage cultivars. In Proceedings of the New Zealand Grasslands Association, Hawkes Bay, New Zealand, 1999; Volume 61, pp. 3–7.
- Gout, M.; Jones, S. Estimates of the annual sales of proprietary cultivars in Australia, and the value of pastures to the livestock and cropping industries. In Presentation to Pastures Australia, Adelaide, Australia, 23–24 February 2006.
- Redfearn, D.D.; Venuto, B.C.; Pitman, W.D.; Blouin, D.C.; Alison, M.W. Multilocation annual ryegrass cultivar performance over a twelve-year period. Crop Sci. 2005, 45, 2388–2393. [Google Scholar] [CrossRef]
- Brummer, E.C. Capturing heterosis in forage crop cultivar development. Crop Sci. 1999, 39, 943–954. [Google Scholar] [CrossRef]
- Foster, C.A. A study of the theoretical expectation of F1 hybridity resulting from bulk interpopulation hybridization in herbage grasses. J. Agric. Sci. 1971, 76, 295–300. [Google Scholar] [CrossRef]
- Foster, C.A. Interpopulational and intervarietal hybridization in Lolium perenne breeding: Heterosis under non-competitive conditions. J. Agric. Sci. 1971, 76, 107–130. [Google Scholar] [CrossRef]
- Foster, C.A. Interpopulational and intervarietal hybridization in Lolium perenne: Heterosis under simulated-sward conditions. J. Agric. Sci. 1971, 76, 401–409. [Google Scholar] [CrossRef]
- Foster, C.A. Interpopulational and intervarietal F1 hybrids in Lolium perenne: Performance in field sward conditions. J. Agric. Sci. 1973, 80, 463–477. [Google Scholar] [CrossRef]
- Islam, M.S.; Studer, B.; Møller, I.M.; Asp, T. Genetics and biology of cytoplasmic male sterility and its applications in forage and turf grass breeding. Plant Breed. 2014, 133, 299–312. [Google Scholar] [CrossRef]
- England, F. The use of incompatibility for the production of F1 hybrids in forage grasses. Heredity 1974, 32, 183–188. [Google Scholar] [CrossRef]
- Posselt, U.K. Hybrid production in Lolium perenne based on incompatibility. Euphytica 1993, 71, 29–33. [Google Scholar] [CrossRef]
- Pembleton, L.W.; Shinozuka, H.; Wang, J.; Spangenberg, G.C.; Forster, J.W.; Cogan, N.O.I. Design of an F1 hybrid breeding strategy for ryegrasses based on selection of self-incompatibility locus-specific alleles. Front. Plant Sci. 2015, 6, 764. [Google Scholar] [CrossRef] [PubMed]
- Posselt, U.K. Identification of heterotic patterns in perennial ryegrass. In Sustainable Use of Genetic Diversity in Forage and Turf Breeding; Huyghe, C., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 569–572. [Google Scholar]
- Wang, J.; Pembleton, L.W.; Baillie, R.C.; Drayton, M.C.; Hand, M.L.; Bain, M.; Sawbridge, T.I.; Spangenberg, G.C.; Forster, J.W.; Cogan, N.O.I. Development and implementation of a multiplexed single nucleotide polymorphism genotyping tool for differentiation of ryegrass species and cultivars. Mol. Breed. 2014, 33, 435–451. [Google Scholar] [CrossRef]
- Wang, J.; Cogan, N.O.I.; Pembleton, L.W.; Forster, J.W. Variance, inter-trait correlation, heritability and trait-marker association of herbage yield, nutritive values, and morphological characteristics in Italian ryegrass (Lolium multiflorum Lam.). Crop Pasture Sci. 2015, 66, 973–984. [Google Scholar]
- Radojevic, I.; Simpson, R.J.; St. John, J.A.; Humphreys, M.O. Chemical composition and in vitro digestibility of lines of Lolium perenne selected for high concentrations of water-soluble carbohydrate. Aust. J. Agric. Res. 1994, 45, 901–912. [Google Scholar] [CrossRef]
- Vogel, K.P.; Pederson, J.F. Breeding systems for cross pollinated perennial grasses. In Plant Breeding Reviews; Janick, J., Ed.; John Wiley & Sons, Inc.: Oxford, UK, 1993; Volume 11, pp. 251–274. [Google Scholar]
- Waldron, B.L.; Robins, J.G.; Peel, M.D.; Jensen, K.B. Predicted efficiency of spaced-plant selection to indirectly improve tall fescue sward yield and quality. Crop Sci. 2008, 48, 443–449. [Google Scholar] [CrossRef]
- Conaghan, P.; Casler, M.D. A theoretical and practical analysis of optimum breeding system for perennial ryegrass. Irish J. Agric. Food Res. 2011, 50, 47–63. [Google Scholar]
- Hayes, B.; Cogan, N.O.I.; Pembleton, L.; Goddard, M.; Wang, J.; Spangenberg, G.C.; Forster, J.W. Prospects for genomic selection in forage plant species. Plant Breed. 2013, 132, 133–143. [Google Scholar] [CrossRef]
- Casler, M.D.; Vogel, K.P. Accomplishments and impact from breeding for increased forage nutritional value. Crop Sci. 1999, 39, 12–20. [Google Scholar] [CrossRef]
- McGrath, D. A note on the influence of nitrogen application and time of cutting on water soluble carbohydrate production by Italian ryegrass. Irish J. Agric. Food Res. 1992, 31, 189–192. [Google Scholar]
- Bhatt, A. Studies on heterosis and inbreeding depression in forage sorghum [Sorghum bicolor (L.) Moench]. Agric. Sci. Digest 2008, 28, 258–261. [Google Scholar]
- Becker, H.C. Breeding synthetic varieties of crop plants. Plant Genet. Breed. Rev. 1988, 1, 31–54. [Google Scholar]
- Hayward, M.D.; Abdultah, I.B. Selection and stability of synthetic varieties of Lolium perenne 1. The selected character and its expression over generations of multiplication. Theor. Appl. Genet. 1985, 70, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Piano, E.; Annicchiarico, P.; Romani, M.; Pecetti, L. Effect of the number of parents and their combining ability on the performance of synthetic varieties in tall fescue. Aust. J. Agric. Res. 2007, 58, 1100–1105. [Google Scholar] [CrossRef]
- Casler, M.D.; Diaby, M.; Stendal, C. Heterosis and inbreeding depression for forage yield and fiber concentration in smooth bromegrass. Crop Sci. 2005, 45, 44–50. [Google Scholar] [CrossRef]
- ‘Tabu’ in Plant Varieties J. 2002, 15, 60.
- ‘Warrior’ in Plant Varieties J. 2003, 20, 162.
- East, E.M. Heterosis. Genetics 1936, 21, 375–397. [Google Scholar] [PubMed]
- Bernardo, R. Relationship between single-cross performance and molecular marker heterozygosity. Theor. Appl. Genet. 1992, 83, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Visscher, P.M.; Hill, W.G.; Wray, N.R. Heritability in the genomics era—Concepts and misconceptions. Nat. Rev. Genet. 2008, 9, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, P.W.; Lovatt, J.A. Chromosome doubling and top-crossing as a means of exploiting heterosis in perennial ryegrass. In Proceedings of Breeding and Seed Production for Conventional and Organic Agriculture, Proceeding of the 26th EUCAPRIA Fodder Crops and Amenity Grasses Section and the 16th Medicago spp. Group Joint Meeting, Perugia, Italy, 3–7 September 2006; pp. 52–55.
- Robins, J.; Bushman, B.; Escribano, S.; Jensen, K. Heterosis for protein, digestibility, fiber, and water soluble carbohydrates in nine sources of orchardgrass germplasm. Euphytica 2015, 204, 503–511. [Google Scholar] [CrossRef]
- Pembleton, L.W.; Wang, J.; Cogan, N.O.I.; Pryce, J.E.; Ye, G.; Bandaranayake, C.K.; Hand, M.L.; Baillie, R.C.; Drayton, M.C.; Lawless, K.; et al. Candidate gene-based association genetics analysis of herbage quality traits in perennial ryegrass (Lolium perenne L.). Crop Pasture Sci. 2013, 64, 244–253. [Google Scholar] [CrossRef]
- Payne, R.W.; Murray, D.A.; Harding, S.A.; Baird, D.B.; Soutar, D.M. GenStat for Windows Introduction; VSN International: Hemel Hempstead, UK, 2009. [Google Scholar]
- Falconer, D.S.; Mackay, T.F.C. Introduction to Quantitative Genetics, 4th ed.; Pearson Education Limited: Harlow, UK, 1996; pp. 160–181. [Google Scholar]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).