
Transcriptome and Physio-Biochemical Profiling Reveals Differentially Expressed Genes in Seedlings from Aerial and Subterranean Seeds Subjected to Drought Stress in Amphicarpaea edgeworthii Benth

 , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Drought Treatment
2.2. Physiological Index Measurements Under Drought Stress
2.3. RNA Extraction and Illumina RNA-Seq
2.4. Raw Data Processing, Read Mapping, and Differentially Expressed Gene Analysis
2.5. Functional Annotation of DEGs
2.6. Quantitative Real-Time PCR (RT-qPCR)
2.7. Statistical Analysis
3. Results
3.1. Phenotypic and Physiological Alterations in A. edgeworthii Under Drought Stress
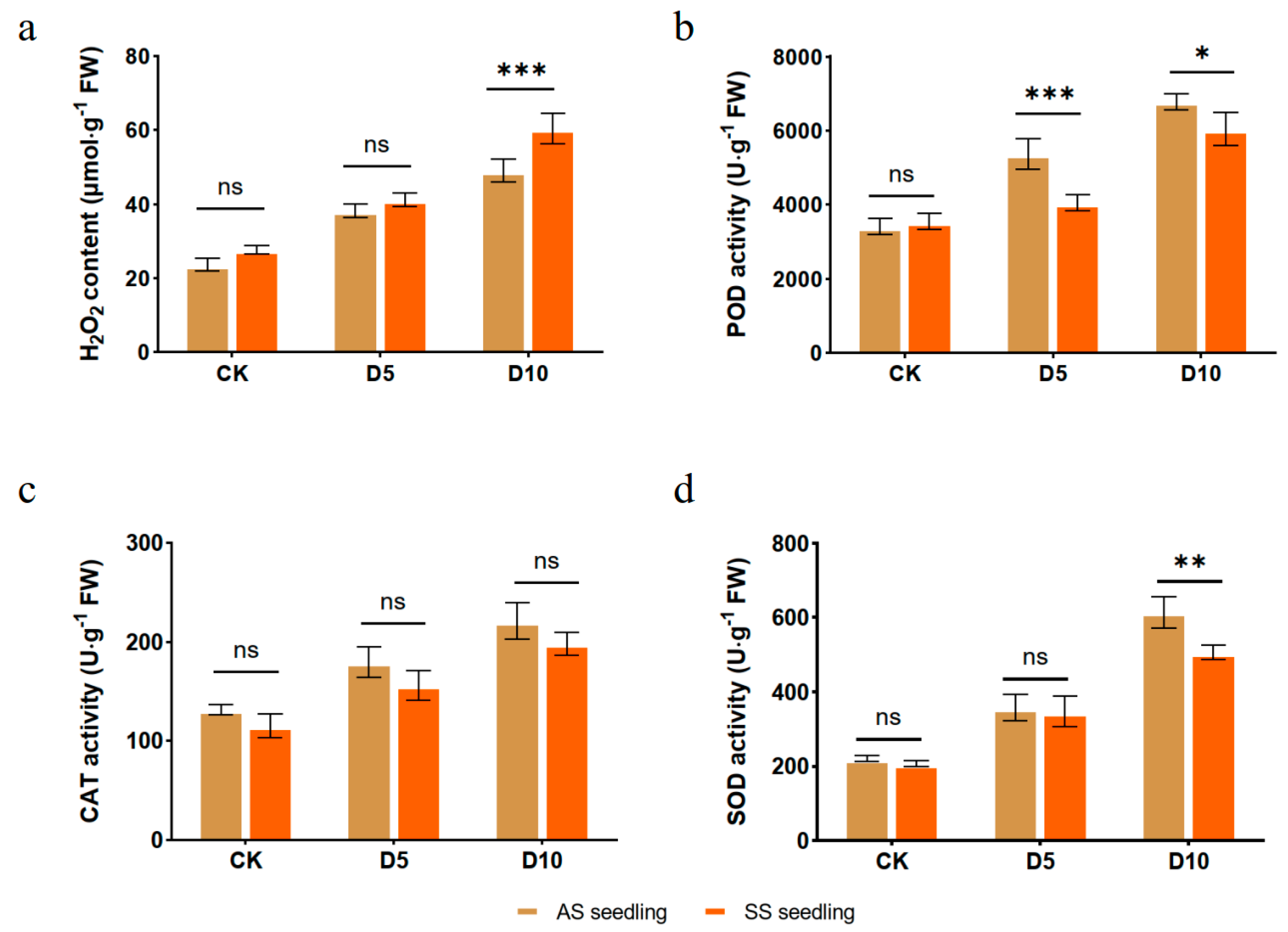
3.2. Effect of Drought Stress on ROSs and Antioxidant Activities in A. edgeworthii
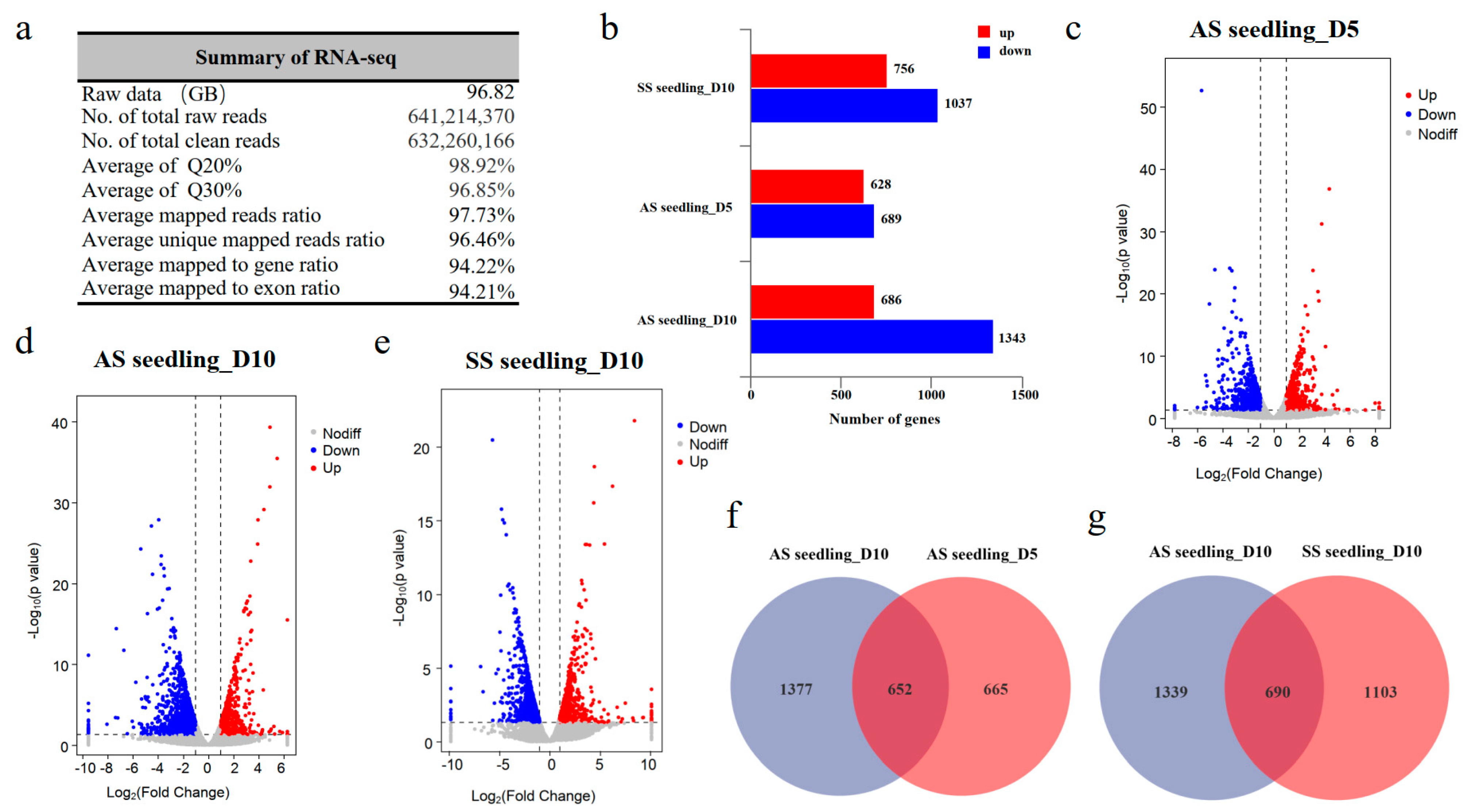
3.3. RNA-Seq Analysis and Screening of DEGs in AS and SS Seedlings Under Drought Stress
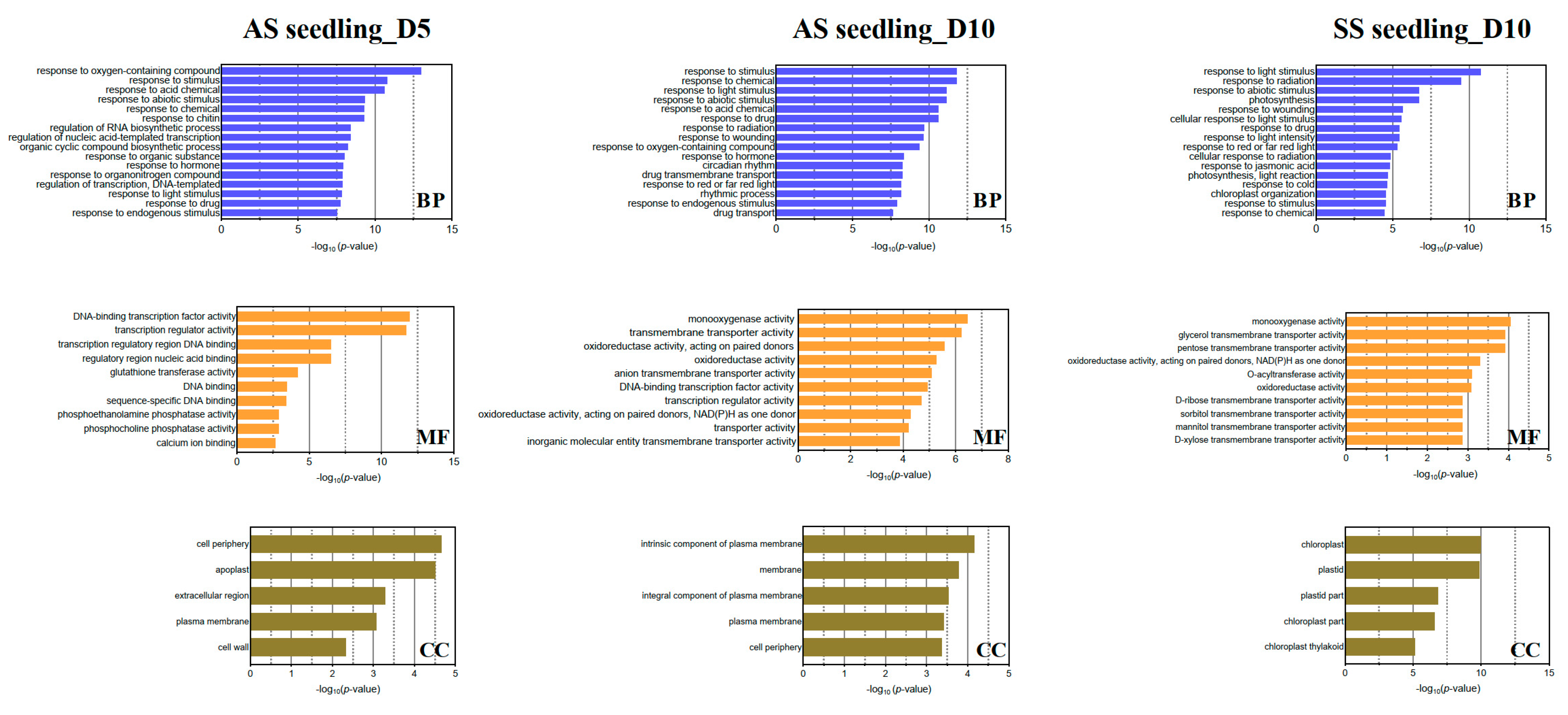
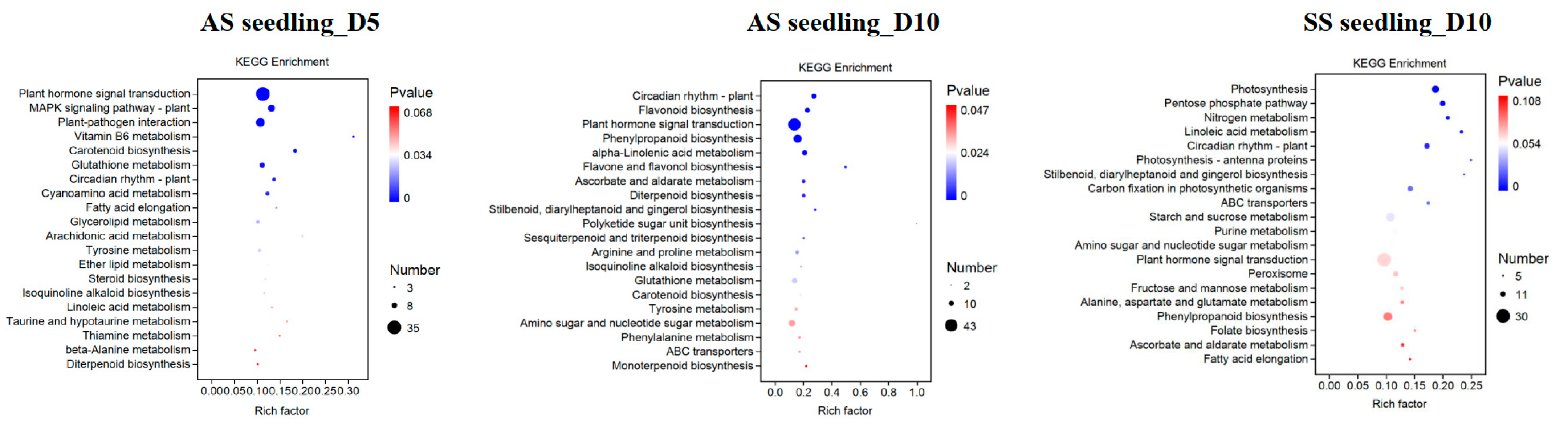
3.4. Functional Enrichment Analyses of Drought-Responsive DEGs
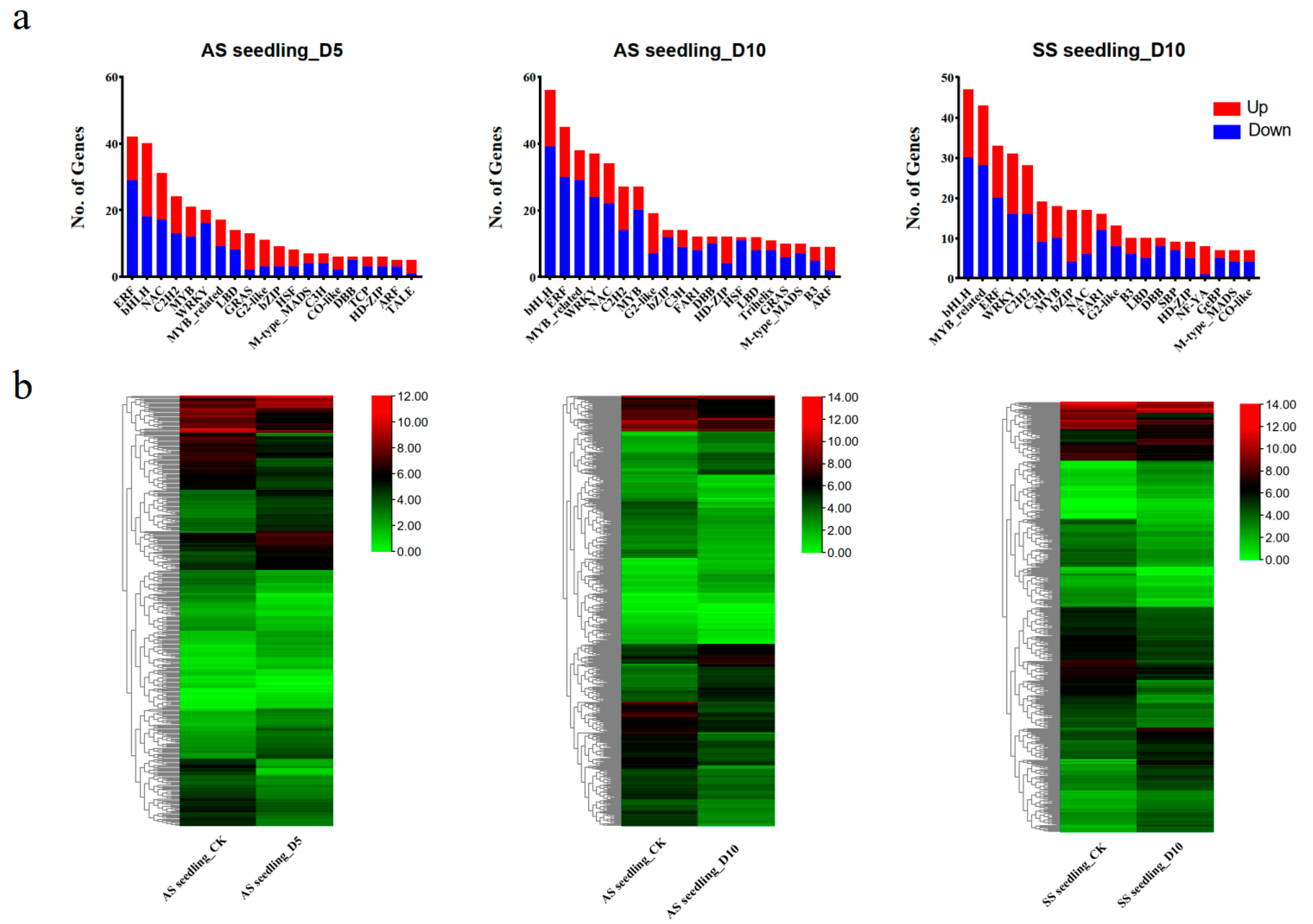
3.5. Distribution of Differentially Expressed Transcription Factors
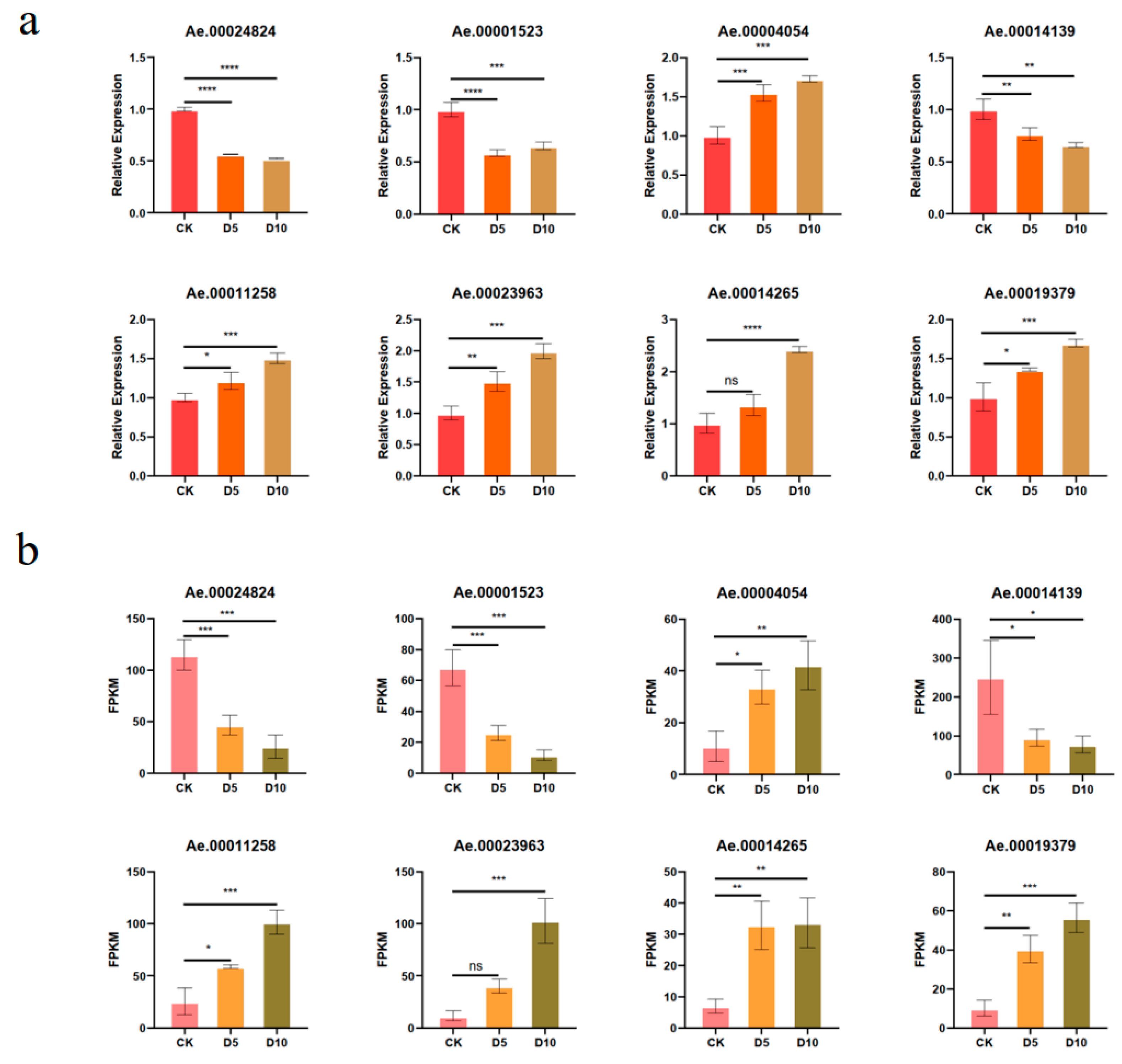
3.6. Validation of DEG Expression Patterns via RT-qPCR Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AS | Aerial seeds |
| SS | Subterranean seeds |
| RNA-seq | RNA sequencing |
| DEG | Differentially expressed gene |
| TF | Transcription factor |
| ROS | Reactive oxygen species |
References
- Zhang, Y.; Yang, J.; Rao, G.-Y. Genetic Diversity of an Amphicarpic Species, Amphicarpaea edgeworthii Benth. (Leguminosae) Based on RAPD Markers. Biochem. Syst. Ecol. 2005, 33, 1246–1257. [Google Scholar] [CrossRef]
- Zhang, K.; Baskin, J.M.; Baskin, C.C.; Yang, X.; Huang, Z. Lack of Divergence in Seed Ecology of Two Amphicarpaea (Fabaceae) Species Disjunct between Eastern Asia and Eastern North America. Am. J. Bot. 2015, 102, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, X.; Han, K.; Li, R.; Xu, G.; Han, Y.; Cui, F.; Fan, S.; Seim, I.; Fan, G.; et al. Insights into Amphicarpy from the Compact Genome of the Legume Amphicarpaea edgeworthii. Plant Biotechnol. J. 2021, 19, 952–965. [Google Scholar] [CrossRef]
- Zhang, K.; Baskin, J.M.; Baskin, C.C.; Yang, X.; Huang, Z. Effect of Seed Morph and Light Level on Growth and Reproduction of the Amphicarpic Plant Amphicarpaea edgeworthii (Fabaceae). Sci. Rep. 2017, 7, 39886. [Google Scholar] [CrossRef]
- González, E.M. Drought Stress Tolerance in Plants. Int. J. Mol. Sci. 2023, 24, 6562. [Google Scholar] [CrossRef]
- Valliyodan, B.; Nguyen, H.T. Understanding Regulatory Networks and Engineering for Enhanced Drought Tolerance in Plants. Curr. Opin. Plant Biol. 2006, 9, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Kakumanu, A.; Ambavaram, M.M.R.; Klumas, C.; Krishnan, A.; Batlang, U.; Myers, E.; Grene, R.; Pereira, A. Effects of Drought on Gene Expression in Maize Reproductive and Leaf Meristem Tissue Revealed by RNA-Seq. Plant Physiol. 2012, 160, 846–867. [Google Scholar] [CrossRef]
- Zhang, A.; Liu, M.; Gu, W.; Chen, Z.; Gu, Y.; Pei, L.; Tian, R. Effect of Drought on Photosynthesis, Total Antioxidant Capacity, Bioactive Component Accumulation, and the Transcriptome of Atractylodes lancea. BMC Plant Biol. 2021, 21, 293. [Google Scholar] [CrossRef]
- Yang, H.; Wu, F.; Cheng, J. Reduced Chilling Injury in Cucumber by Nitric Oxide and the Antioxidant Response. Food Chem. 2011, 127, 1237–1242. [Google Scholar] [CrossRef]
- Nadeem, M.; Li, J.; Yahya, M.; Sher, A.; Ma, C.; Wang, X.; Qiu, L. Research Progress and Perspective on Drought Stress in Legumes: A Review. Int. J. Mol. Sci. 2019, 20, 2541. [Google Scholar] [CrossRef]
- Bhatnagar-Mathur, P.; Vadez, V.; Jyostna Devi, M.; Lavanya, M.; Vani, G.; Sharma, K.K. Genetic Engineering of Chickpea (Cicer arietinum L.) with the P5CSF129A Gene for Osmoregulation with Implications on Drought Tolerance. Mol. Breed. 2009, 23, 591–606. [Google Scholar] [CrossRef]
- Chen, Y.; Chi, Y.; Meng, Q.; Wang, X.; Yu, D. GmSK1, an SKP1 Homologue in Soybean, Is Involved in the Tolerance to Salt and Drought. Plant Physiol. Biochem. 2018, 127, 25–31. [Google Scholar] [CrossRef]
- Privitera, G.F.; Treccarichi, S.; Nicotra, R.; Branca, F.; Pulvirenti, A.; Lo Piero, A.R.; Sicilia, A. Comparative Transcriptome Analysis of B. oleracea L. Var. Italica and B. macrocarpa Guss. Genotypes under Drought Stress: De Novo vs Reference Genome Assembly. Plant Stress 2024, 14, 100657. [Google Scholar] [CrossRef]
- Behera, S.; Voshall, A.; Moriyama, E.N. Plant Transcriptome Assembly: Review and Benchmarking. In Bioinformatics; Nakaya, H.I., Ed.; Exon Publications: Brisbane, Australia, 2021; pp. 109–130. ISBN 978-0-6450017-1-6. [Google Scholar]
- Ding, N.; Zhao, Y.; Wang, W.; Liu, X.; Shi, W.; Zhang, D.; Chen, J.; Ma, S.; Sun, Q.; Wang, T.; et al. Transcriptome Analysis in Contrasting Maize Inbred Lines and Functional Analysis of Five Maize NAC Genes under Drought Stress Treatment. Front. Plant Sci. 2023, 13, 1097719. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.; Dun, B.; Li, L.; Ma, X.; Zhang, H.; Su, Y.; Wu, D. Metabolomic and Transcriptomic Responses of Adiantum (Adiantum nelumboides) Leaves under Drought, Half-Waterlogging, and Rewater Conditions. Front. Genet. 2023, 14, 1113470. [Google Scholar] [CrossRef]
- Singh, V.; Gupta, K.; Singh, S.; Jain, M.; Garg, R. Unravelling the Molecular Mechanism Underlying Drought Stress Response in Chickpea via Integrated Multi-Omics Analysis. Front. Plant Sci. 2023, 14, 1156606. [Google Scholar] [CrossRef]
- Azzouz-Olden, F.; Hunt, A.G.; Dinkins, R. Transcriptome Analysis of Drought-Tolerant Sorghum Genotype SC56 in Response to Water Stress Reveals an Oxidative Stress Defense Strategy. Mol. Biol. Rep. 2020, 47, 3291–3303. [Google Scholar] [CrossRef]
- Li, M.; Li, H.; Sun, A.; Wang, L.; Ren, C.; Liu, J.; Gao, X. Transcriptome Analysis Reveals Key Drought-Stress-Responsive Genes in Soybean. Front. Genet. 2022, 13, 1060529. [Google Scholar] [CrossRef]
- Aleem, M.; Raza, M.M.; Haider, M.S.; Atif, R.M.; Ali, Z.; Bhat, J.A.; Zhao, T. Comprehensive RNA-seq Analysis Revealed Molecular Pathways and Genes Associated with Drought Tolerance in Wild Soybean (Glycine soja Sieb. and Zucc.). Physiol. Plant. 2021, 172, 707–732. [Google Scholar] [CrossRef]
- Abdeen, A.; Schnell, J.; Miki, B. Transcriptome Analysis Reveals Absence of Unintended Effects in Drought-Tolerant Transgenic Plants Overexpressing the Transcription Factor ABF3. BMC Genom. 2010, 11, 69. [Google Scholar] [CrossRef]
- Jia, X.; Sun, C.; Zuo, Y.; Li, G.; Li, G.; Ren, L.; Chen, G. Integrating Transcriptomics and Metabolomics to Characterise the Response of Astragalus membranaceus Bge. var. Mongolicus (Bge.) to Progressive Drought Stress. BMC Genom. 2016, 17, 188. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Alekseyev, Y.O.; Fazeli, R.; Yang, S.; Basran, R.; Maher, T.; Miller, N.S.; Remick, D. A Next-Generation Sequencing Primer—How Does It Work and What Can It Do? Acad. Pathol. 2018, 5, 2374289518766521. [Google Scholar] [CrossRef]
- Liu, E.; Xu, L.; Luo, Z.; Li, Z.; Zhou, G.; Gao, H.; Fang, F.; Tang, J.; Zhao, Y.; Zhou, Z.; et al. Transcriptomic Analysis Reveals Mechanisms for the Different Drought Tolerance of Sweet Potatoes. Front. Plant Sci. 2023, 14, 1136709. [Google Scholar] [CrossRef]
- Tamang, B.G.; Li, S.; Rajasundaram, D.; Lamichhane, S.; Fukao, T. Overlapping and Stress-specific Transcriptomic and Hormonal Responses to Flooding and Drought in Soybean. Plant J. 2021, 107, 100–117. [Google Scholar] [CrossRef]
- Chen, D.; Wang, S.; Cao, B.; Cao, D.; Leng, G.; Li, H.; Yin, L.; Shan, L.; Deng, X. Genotypic Variation in Growth and Physiological Response to Drought Stress and Re-Watering Reveals the Critical Role of Recovery in Drought Adaptation in Maize Seedlings. Front. Plant Sci. 2016, 6, 1241. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Ma, Y.; Xie, H.; Chang, F.; Guan, C.; Yang, B.; Ma, Y. Proline Metabolism in Response to Climate Extremes in Hairgrass. Plants 2024, 13, 1408. [Google Scholar] [CrossRef]
- Čakar, U.; Čolović, M.; Milenković, D.; Pagnacco, M.; Maksimović, J.; Krstić, D.; Đorđević, B. Strawberry and Drupe Fruit Wines Antioxidant Activity and Protective Effect Against Induced Oxidative Stress in Rat Synaptosomes. Antioxidants 2025, 14, 155. [Google Scholar] [CrossRef]
- Ahmad, M.; Waraich, E.A.; Zulfiqar, U.; Yong, J.W.H.; Ishfaq, M.; Din, K.U.; Ullah, A.; Abbas, A.; Awan, M.I.; Moussa, I.M.; et al. Thiourea Improves Yield and Quality Traits of Brassica napus L. by Upregulating the Antioxidant Defense System under High Temperature Stress. Sci. Rep. 2024, 14, 12195. [Google Scholar] [CrossRef]
- Das, D.; Chowdhury, N.; Sharma, M.; Suma, R.; Saikia, B.; Velmurugan, N.; Chikkaputtaiah, C. Screening for Brown-Spot Disease and Drought Stress Response and Identification of Dual-Stress Responsive Genes in Rice Cultivars of Northeast India. Physiol. Mol. Biol. Plants 2024, 30, 647–663. [Google Scholar] [CrossRef]
- Zhang, S.; Xu, X.; Sun, Y.; Zhang, J.; Li, C. Influence of Drought Hardening on the Resistance Physiology of Potato Seedlings under Drought Stress. J. Integr. Agric. 2018, 17, 336–347. [Google Scholar] [CrossRef]
- Chen, Q.; Hu, T.; Li, X.; Song, C.-P.; Zhu, J.-K.; Chen, L.; Zhao, Y. Phosphorylation of SWEET Sucrose Transporters Regulates Plant Root:Shoot Ratio under Drought. Nat. Plants 2022, 8, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yan, C.; Cao, Y.; Wang, C.; Sun, X.; Zhang, L.; Wang, W.; Song, S. Comparative Physiological and Transcriptomic Analysis of Two Contrasting Soybean Genotypes Reveals Complex Mechanisms Involved in Drought Avoidance. Crop Sci. 2024, 64, 788–802. [Google Scholar] [CrossRef]
- Abdel-Ghany, S.E.; Ullah, F.; Ben-Hur, A.; Reddy, A.S.N. Transcriptome Analysis of Drought-Resistant and Drought-Sensitive Sorghum (Sorghum bicolor) Genotypes in Response to PEG-Induced Drought Stress. Int. J. Mol. Sci. 2020, 21, 772. [Google Scholar] [CrossRef]
- Zhu, T.; Liu, T.; Kang, S.; Zhang, J.; Zhang, S.; Yang, B.; Ma, X.; Guo, L.; Li, M.; Jin, L. Integrated Physiological Characterisation and Transcriptomics Reveals Drought Tolerance Differences between Two Cultivars of A. Sinensis at Seedling Stage. Mol. Biol. Rep. 2025, 52, 283. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Feng, C. Genome-Wide Analysis of MYB Genes in Primulina eburnea (Hance) and Identification of Members in Response to Drought Stress. Int. J. Mol. Sci. 2023, 25, 465. [Google Scholar] [CrossRef]
- Tao, R.; Liu, Y.; Chen, S.; Shityakov, S. Meta-Analysis of the Effects of Overexpressed bZIP Transcription Factors in Plants under Drought Stress. Plants 2024, 13, 337. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wang, L.; Wang, S. Comprehensive Analysis and Discovery of Drought-Related NAC Transcription Factors in Common Bean. BMC Plant Biol. 2016, 16, 193. [Google Scholar] [CrossRef]
- Wu, J.; Chen, J.; Wang, L.; Wang, S. Genome-Wide Investigation of WRKY Transcription Factors Involved in Terminal Drought Stress Response in Common Bean. Front. Plant Sci. 2017, 8, 380. [Google Scholar] [CrossRef]
- Li, H.; Wang, Y.; Wu, M.; Li, L.; Li, C.; Han, Z.; Yuan, J.; Chen, C.; Song, W.; Wang, C. Genome-Wide Identification of AP2/ERF Transcription Factors in Cauliflower and Expression Profiling of the ERF Family under Salt and Drought Stresses. Front. Plant Sci. 2017, 8, 946. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kou, J.; Su, Y.; Lei, T.; Hou, S.; Tian, J.; Li, M.; Zhang, S.; Ding, X.; Li, Q.; Xiao, J. Transcriptome and Physio-Biochemical Profiling Reveals Differentially Expressed Genes in Seedlings from Aerial and Subterranean Seeds Subjected to Drought Stress in Amphicarpaea edgeworthii Benth. Agronomy 2025, 15, 735. https://doi.org/10.3390/agronomy15030735
Kou J, Su Y, Lei T, Hou S, Tian J, Li M, Zhang S, Ding X, Li Q, Xiao J. Transcriptome and Physio-Biochemical Profiling Reveals Differentially Expressed Genes in Seedlings from Aerial and Subterranean Seeds Subjected to Drought Stress in Amphicarpaea edgeworthii Benth. Agronomy. 2025; 15(3):735. https://doi.org/10.3390/agronomy15030735
Chicago/Turabian StyleKou, Jiancheng, Yue Su, Tianyu Lei, Siqi Hou, Jiali Tian, Minglong Li, Shuzhen Zhang, Xiaodong Ding, Qiang Li, and Jialei Xiao. 2025. "Transcriptome and Physio-Biochemical Profiling Reveals Differentially Expressed Genes in Seedlings from Aerial and Subterranean Seeds Subjected to Drought Stress in Amphicarpaea edgeworthii Benth" Agronomy 15, no. 3: 735. https://doi.org/10.3390/agronomy15030735
APA StyleKou, J., Su, Y., Lei, T., Hou, S., Tian, J., Li, M., Zhang, S., Ding, X., Li, Q., & Xiao, J. (2025). Transcriptome and Physio-Biochemical Profiling Reveals Differentially Expressed Genes in Seedlings from Aerial and Subterranean Seeds Subjected to Drought Stress in Amphicarpaea edgeworthii Benth. Agronomy, 15(3), 735. https://doi.org/10.3390/agronomy15030735