
Impact of Industrial Hemp (Cannabis sativa L.) Extracts on Seed Germination and Seedling Growth: Evaluating Allelopathic Activity Across Various Extraction Methods

 , ,
, ,  , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Raw Material
2.2. Chemicals
2.3. Conventional Methods of Extracting Plant Raw Materials
2.3.1. Maceration
2.3.2. Hydrodistillation
2.4. Green Methods of Extracting Hemp Panicles
2.4.1. Microwave-Assisted Extraction (MAE)
2.4.2. Ultrasound-Assisted Extraction (UAE)
2.5. Identification and Quantification of Phenolic Compounds
2.6. Identification and Quantification of Cannabinoids
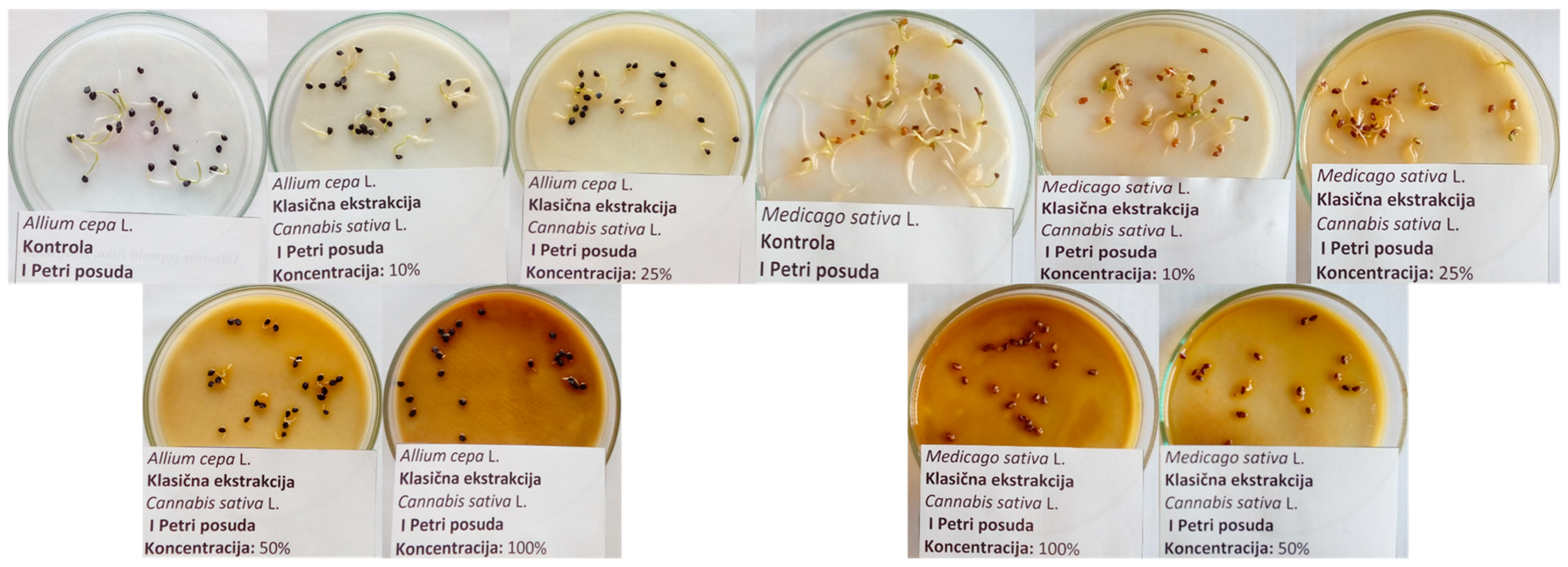
2.7. Seed Germination Protocol
2.8. Statistical Analysis
3. Results
3.1. Phenolic Content in Conventional and Green Hemp Extracts
3.2. Cannabinoids Content in Conventional and Green Hemp Extracts
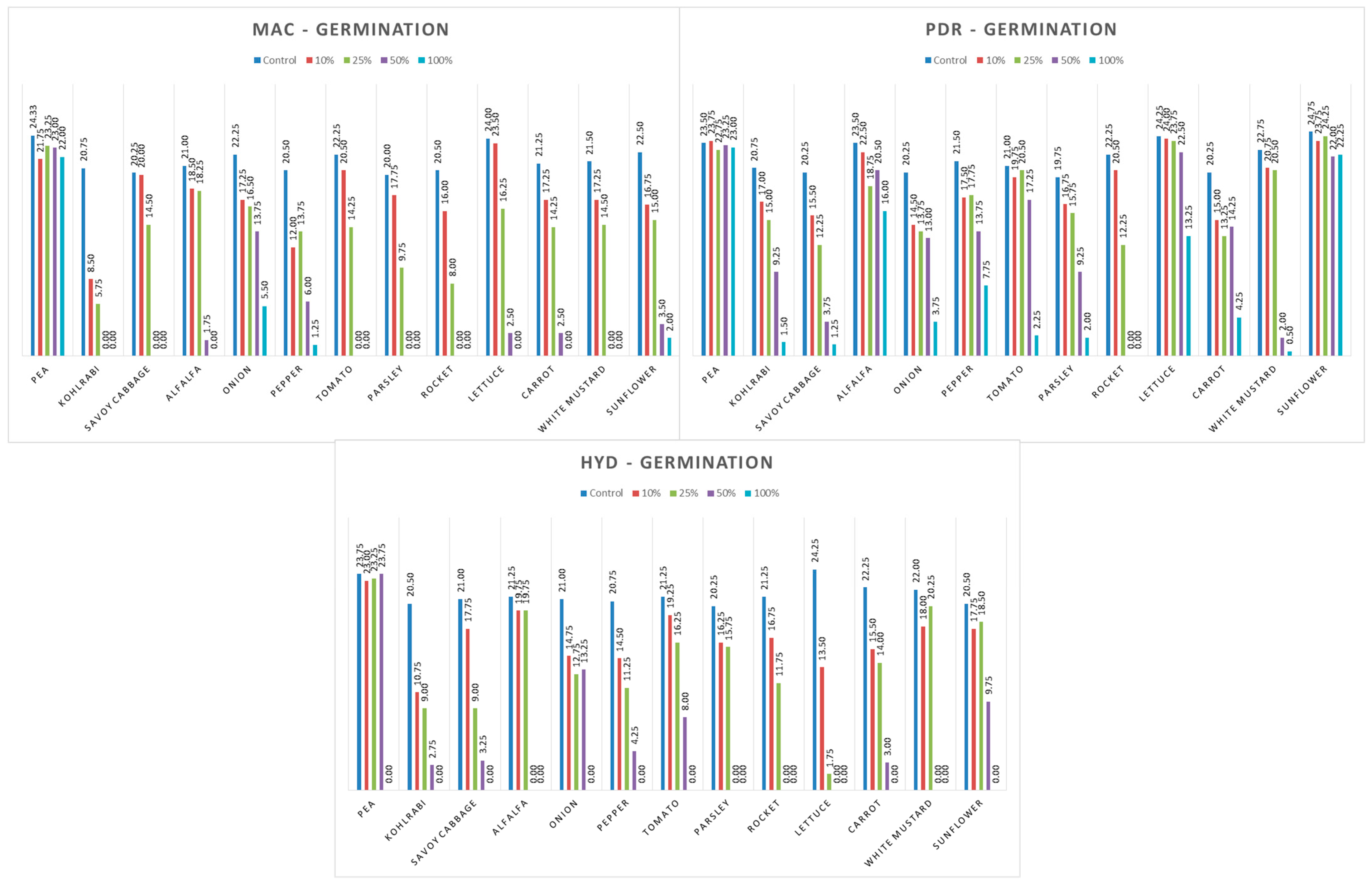
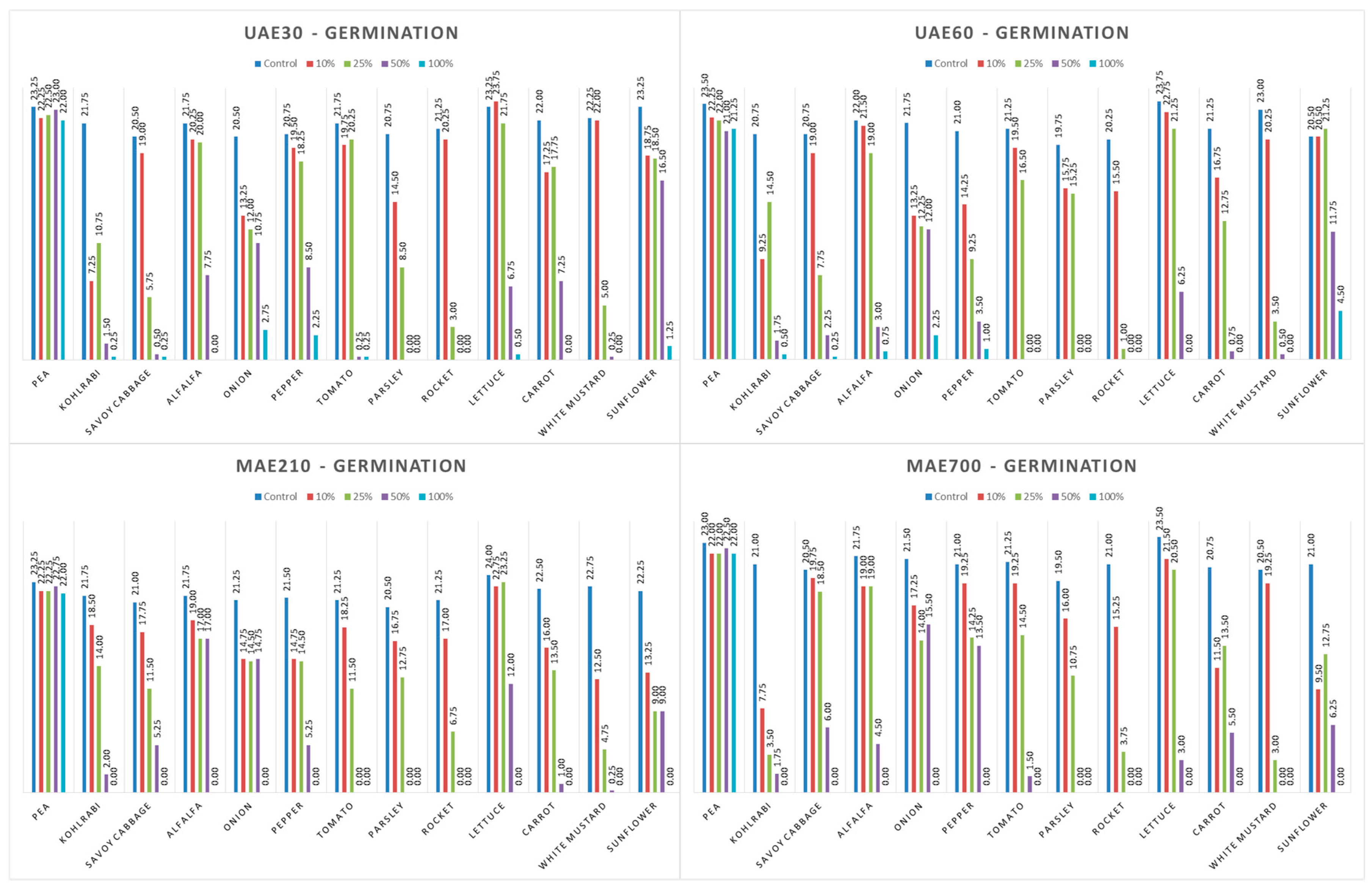
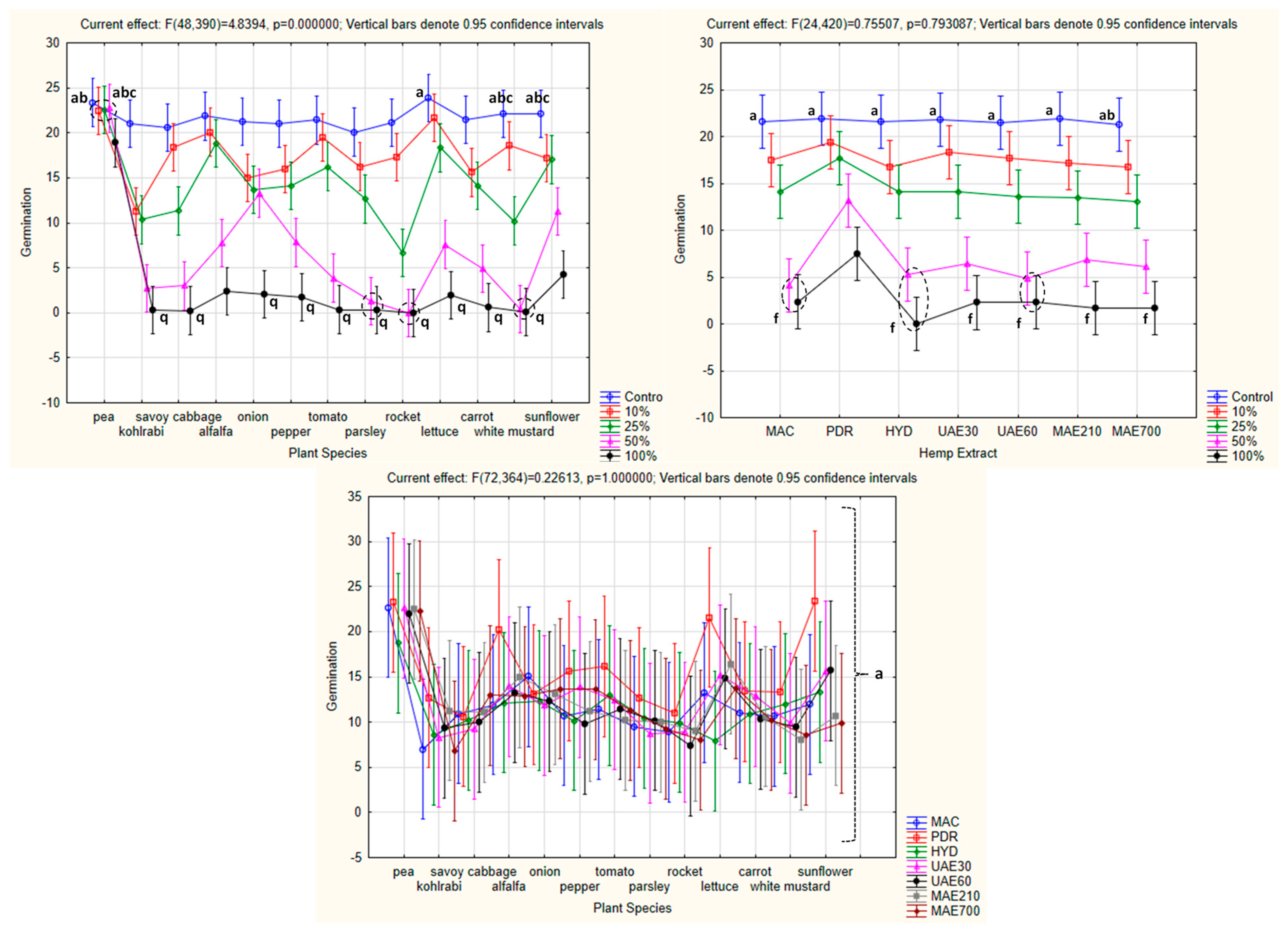
3.3. Influence of Hemp Extracts on Seed Germination: Conventional vs. Green Extracts (CE vs. GE)
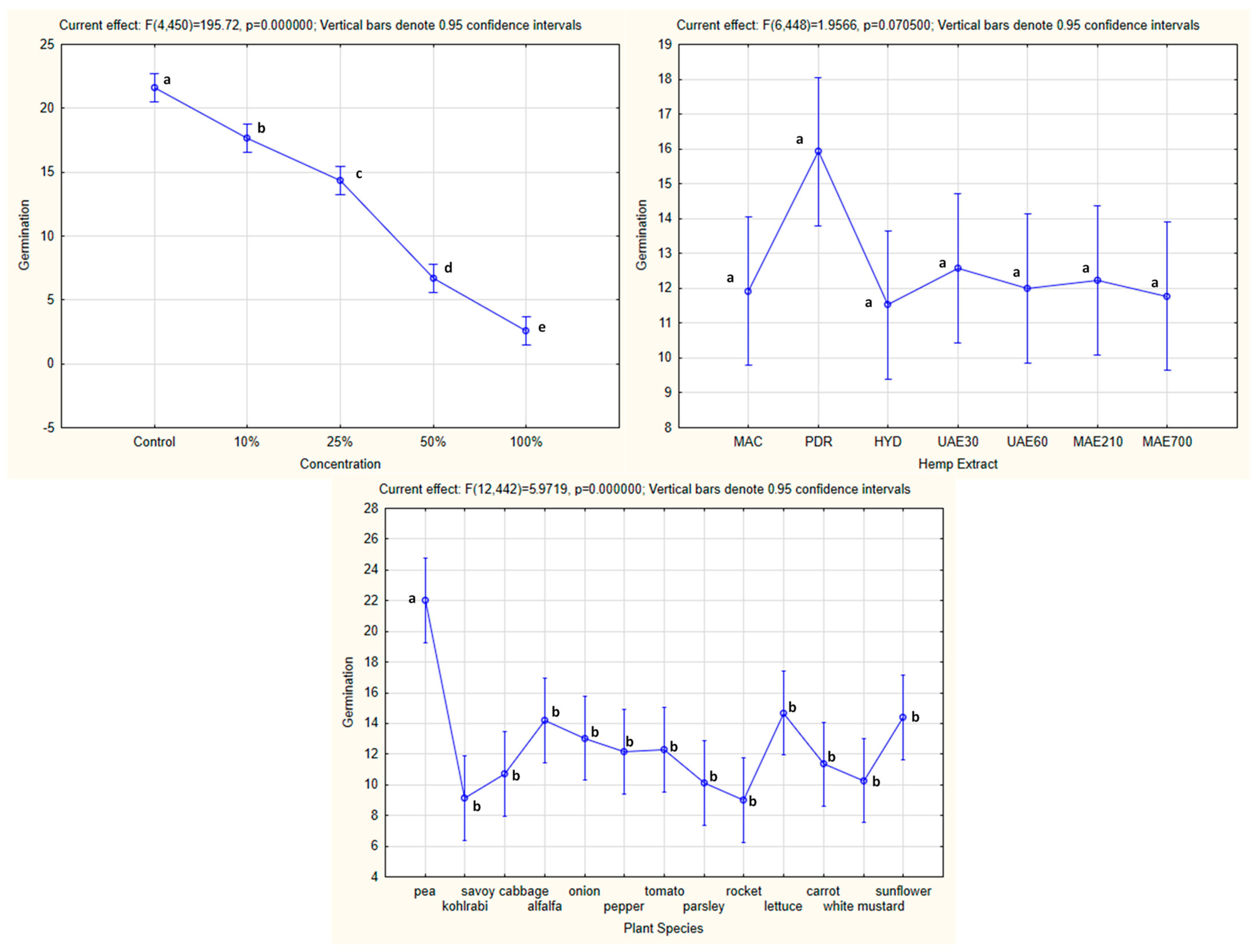
3.3.1. One-Way ANOVA: The Effect of Concentration on Seed Germination of the Tested Plants
3.3.2. One-Way ANOVA: The Effect of Hemp Extract on Seed Germination of the Tested Plants
3.3.3. One-Way ANOVA: The Effect of Plant Species on Seed Germination of the Tested Plants
3.3.4. Factorial ANOVA: The Combined Effect of Concentration and Plant Species on Seed Germination of the Tested Plants
3.3.5. Factorial ANOVA: The Combined Effect of Hemp Extract and Concentration on Seed Germination of the Tested Plants
3.3.6. Factorial ANOVA: The Combined Effect of Hemp Extract and Plant Species on Seed Germination of the Tested Plants
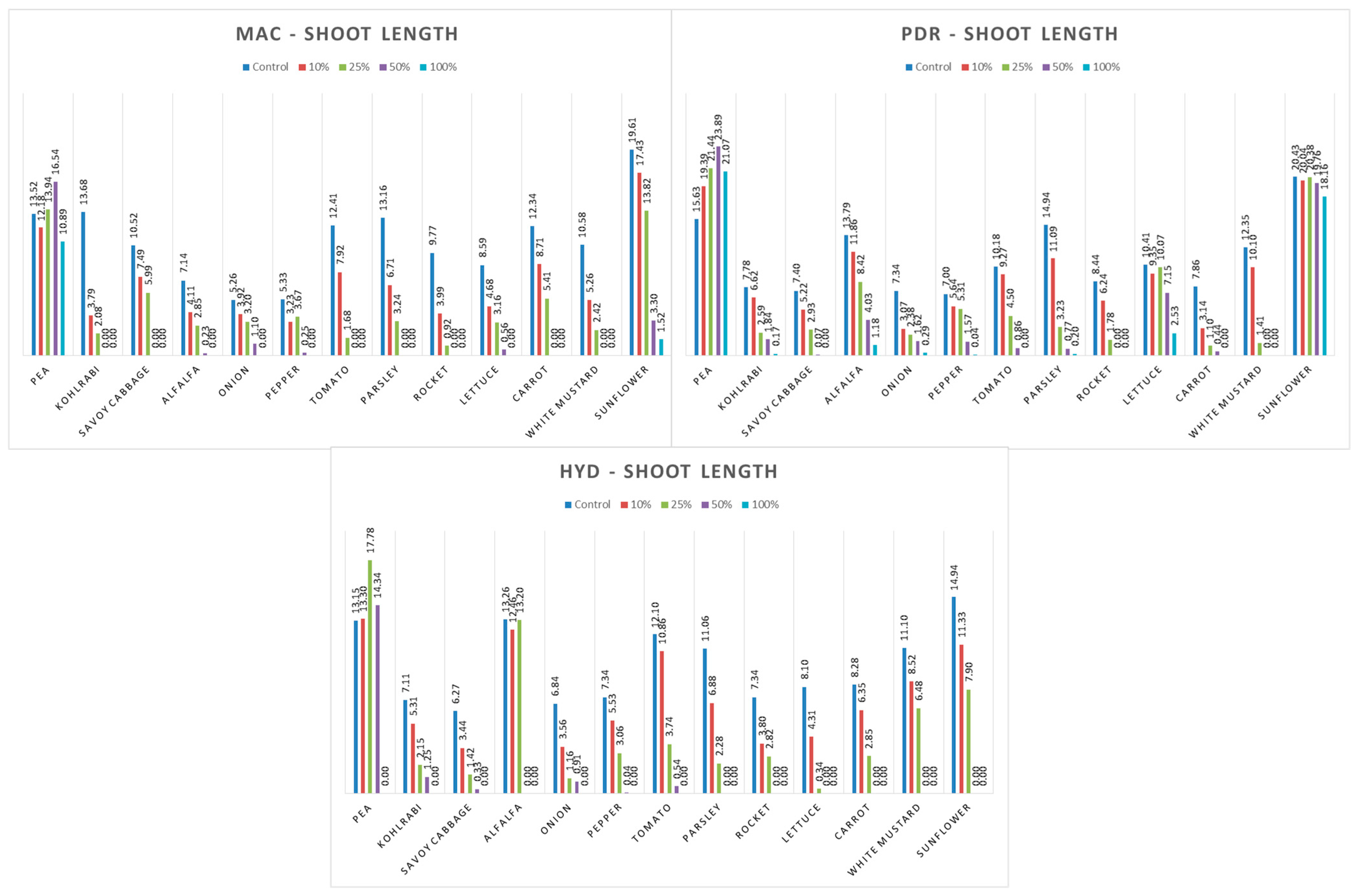
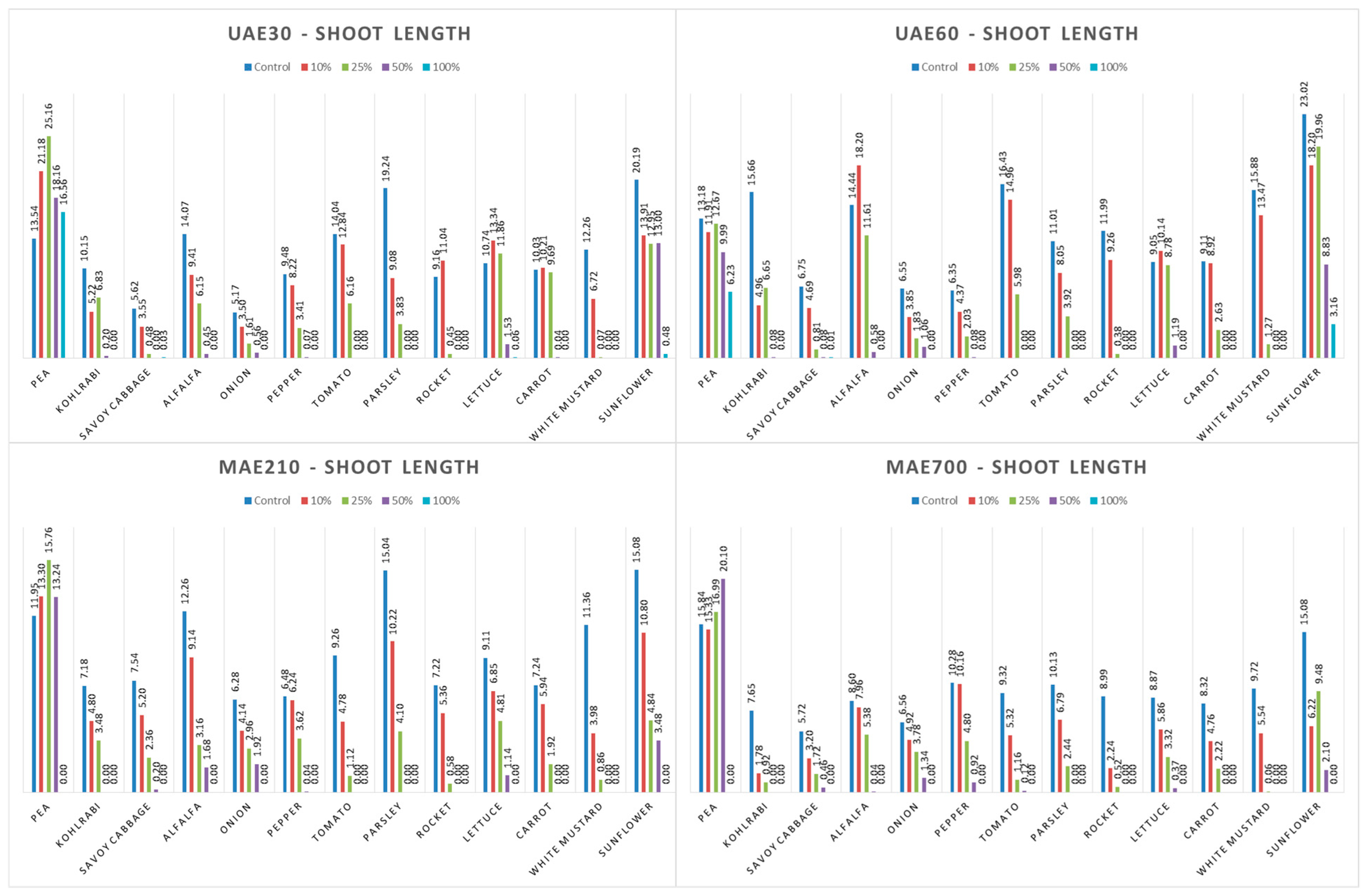
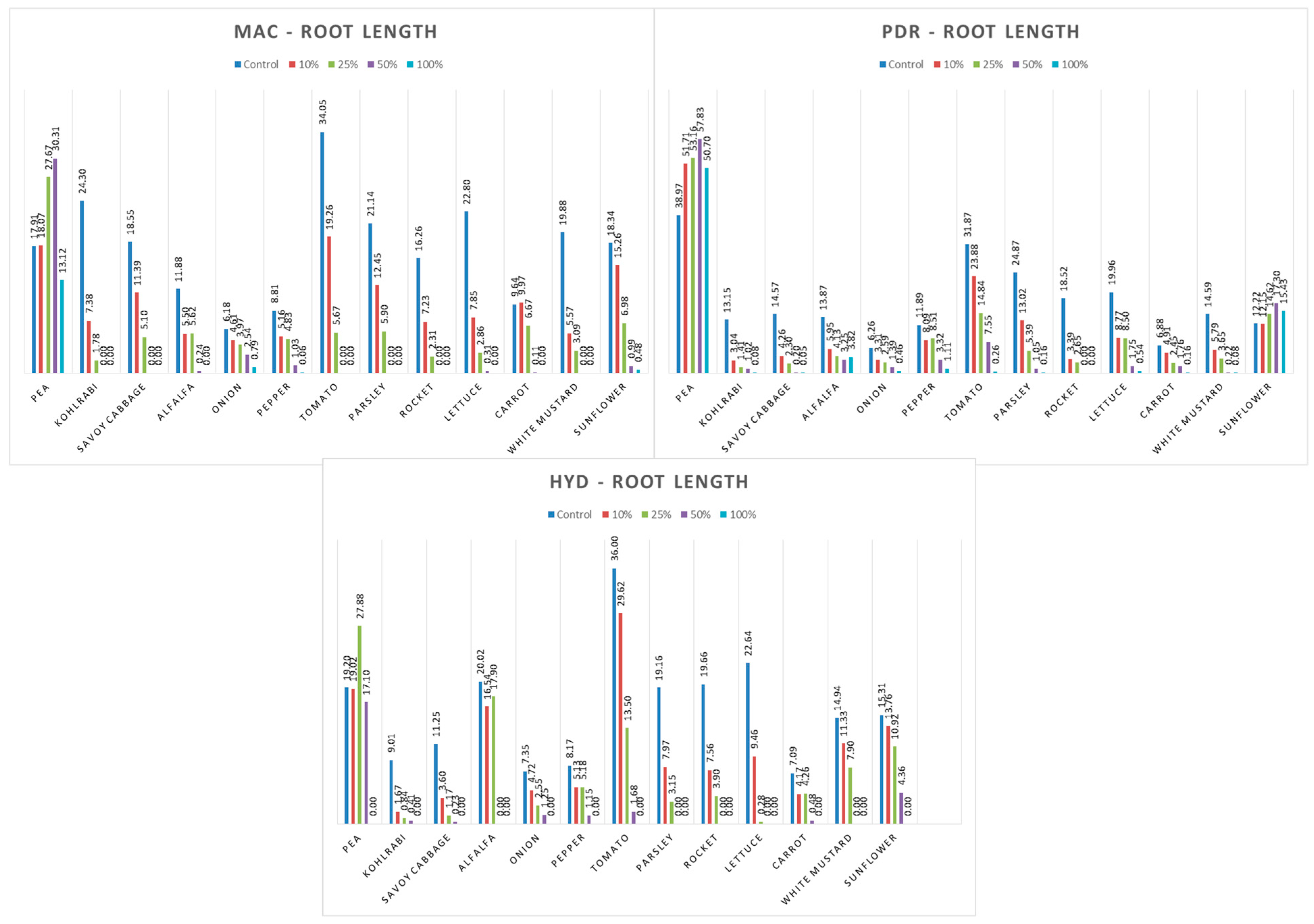
3.4. Influence of Hemp Extracts on Root/Shoot Length: CE vs. GE
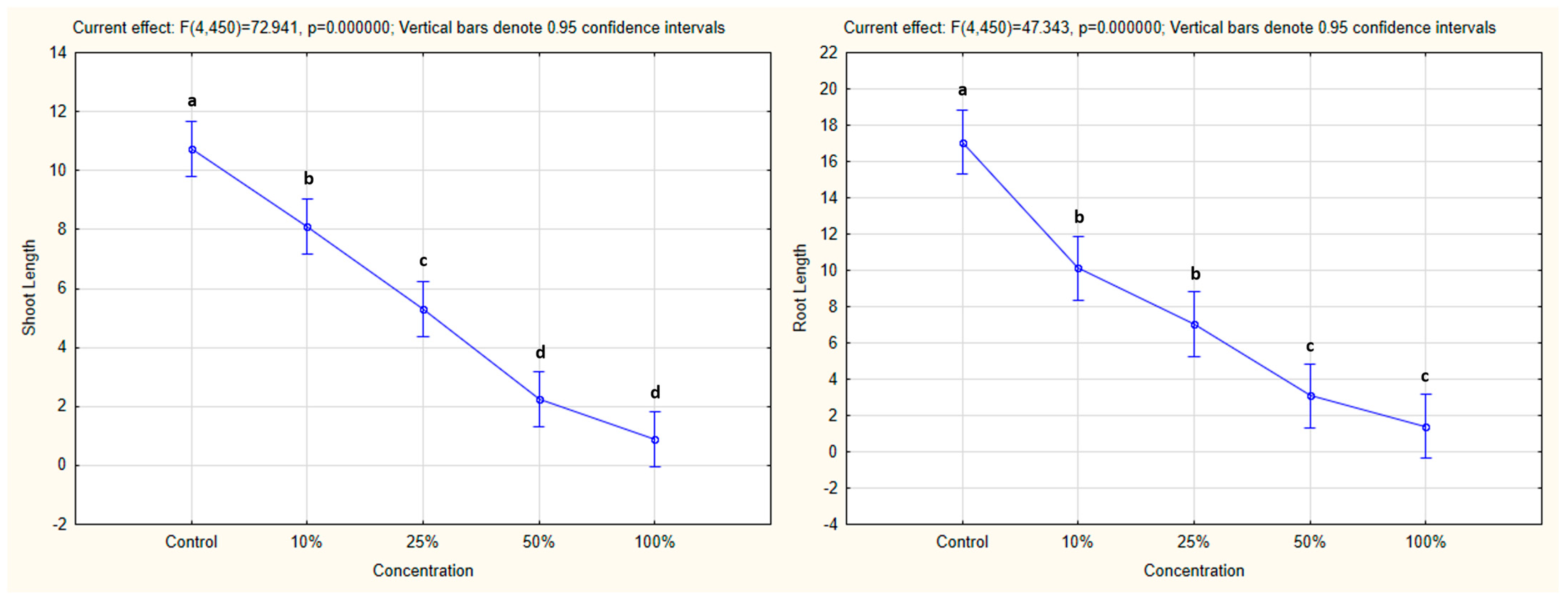
3.4.1. One-Way ANOVA: The Effect of Concentration on Shoot and Root Lengths of the Tested Plants
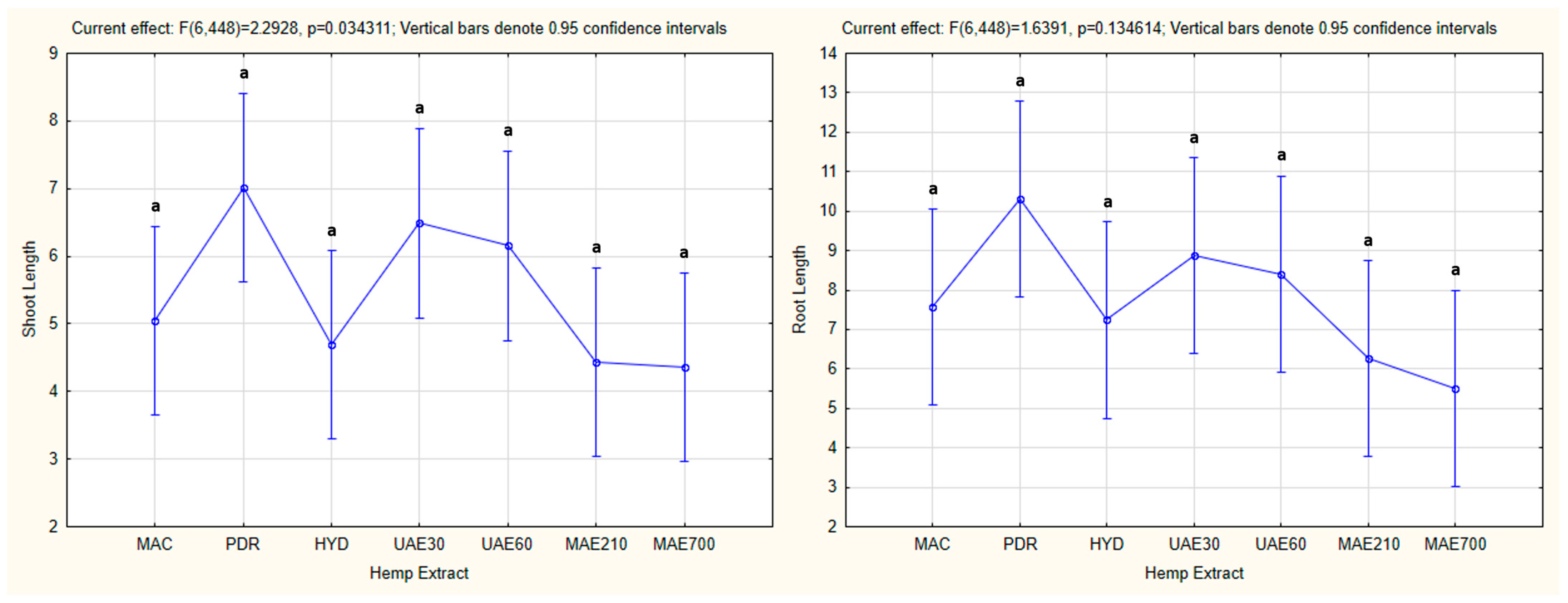
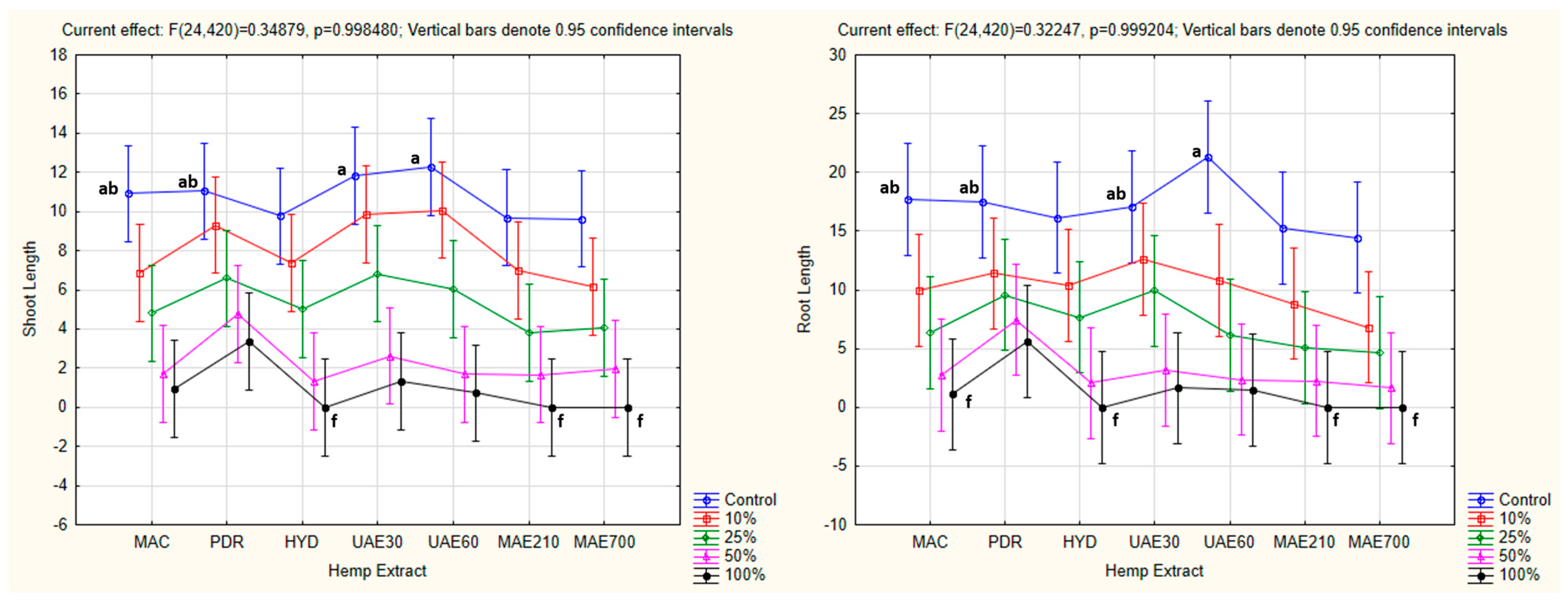
3.4.2. One-Way ANOVA: The Effect of Hemp Extract on Shoot and Root Lengths of the Tested Plants
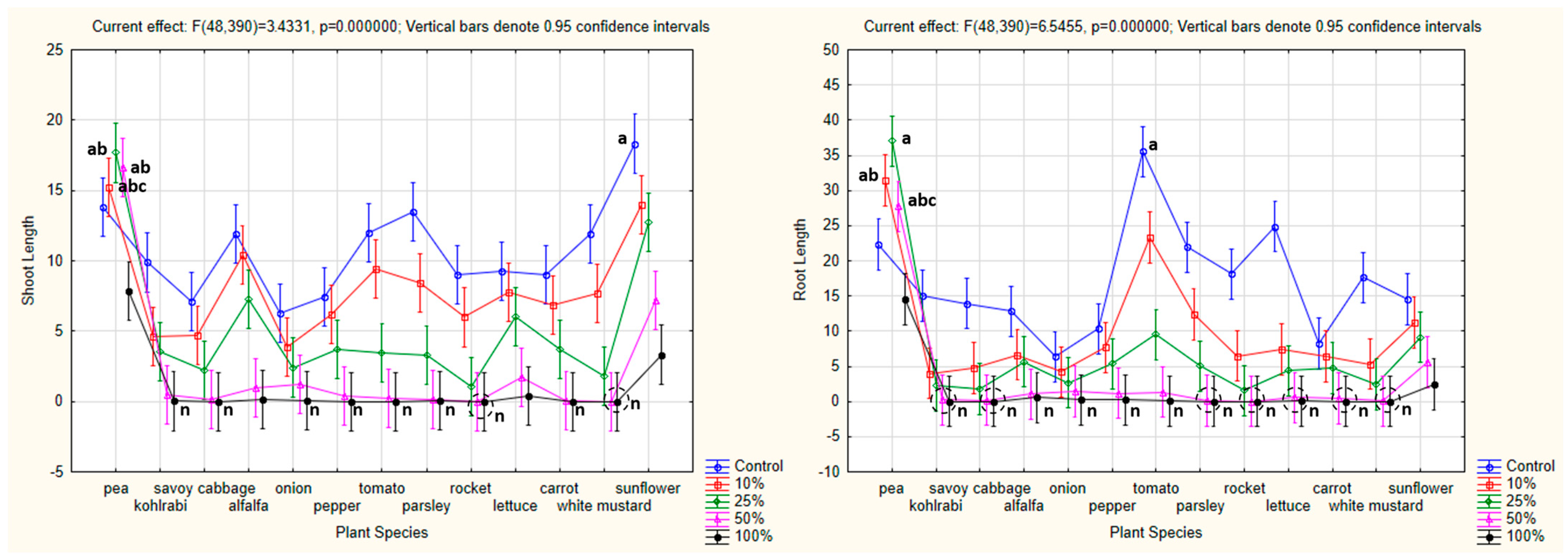
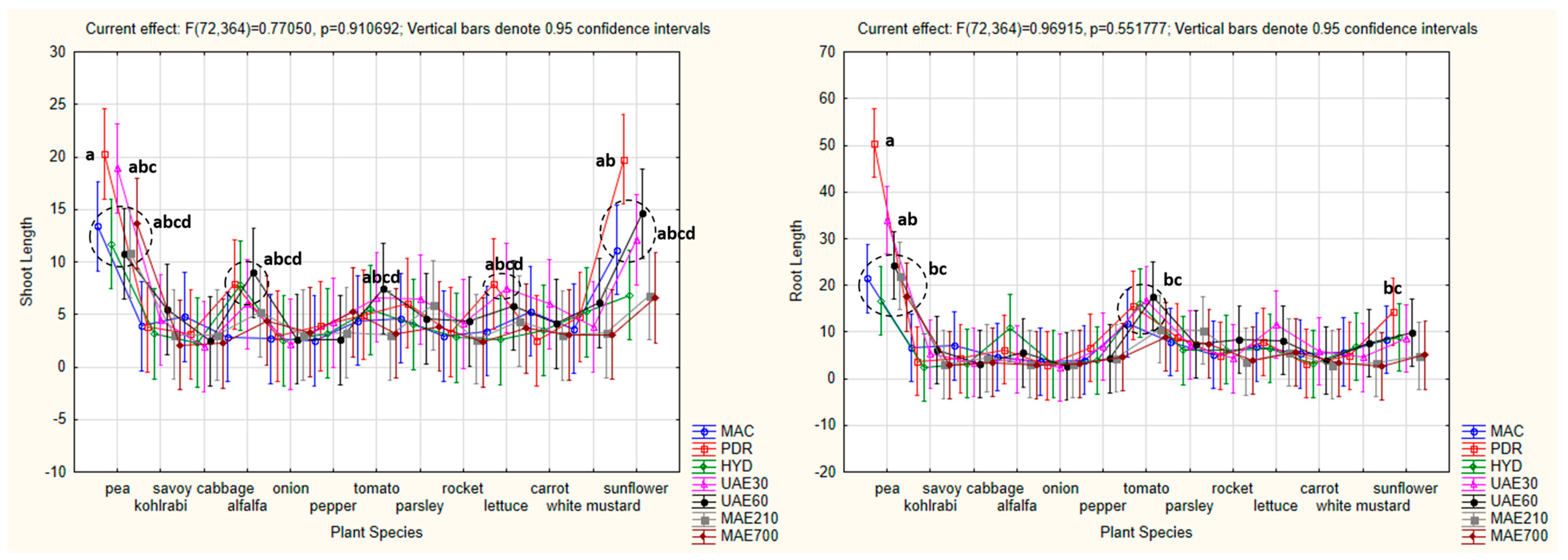
3.4.3. One-Way ANOVA: The Effect of Plant Species on Shoot and Root Lengths of the Tested Plants
3.4.4. Factorial ANOVA: The Combined Effect of Concentration and Plant Species on Shoot and Root Lengths of the Tested Plants
3.4.5. Factorial ANOVA: The Combined Effect of Hemp Extract and Concentration on Shoot and Root Lengths of the Tested Plants
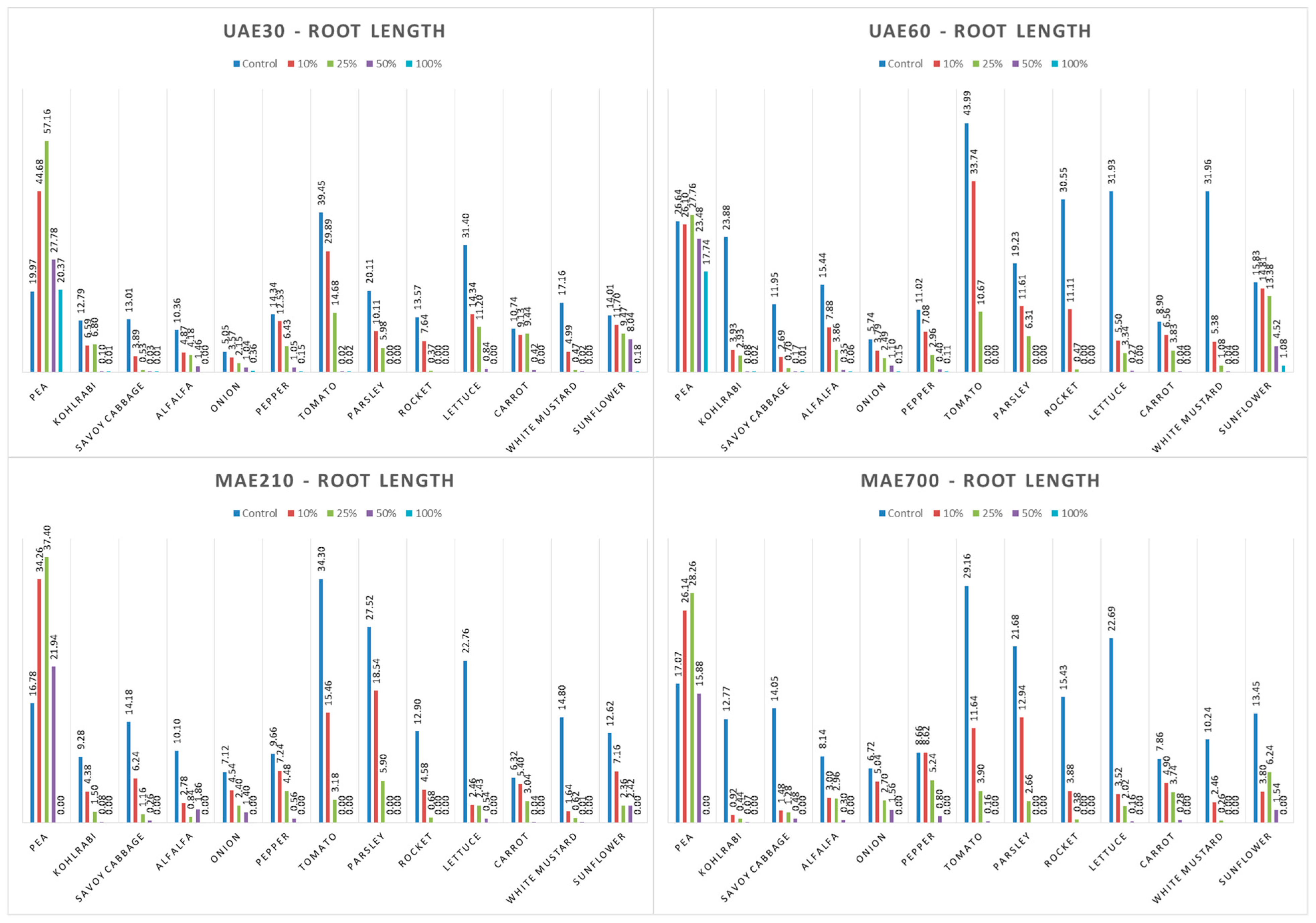
3.4.6. Factorial ANOVA: The Combined Effect of Hemp Extract and Plant Species on Shoot and Root Lengths of the Tested Plants
4. Discussion
4.1. Phenolic Content in Conventional and Green Hemp Extracts
4.2. Cannabinoids Content in Conventional and Green Hemp Extracts
4.3. Influence of Hemp Extracts on Seed Germination: Conventional vs. Green Extracts (CE vs. GE)
4.4. Influence of Hemp Extracts on Root/Shoot Length: CE vs. GE
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Owusu, N.O.; Arthur, B.; Aboagye, E.M. Industrial hemp as an agricultural crop in Ghana. J. Cannabis Res. 2021, 3, 9. [Google Scholar] [CrossRef] [PubMed]
- Drinić, Z.; Vladić, J.; Koren, A.; Zeremski, T.; Stojanov, N.; Tomić, M.; Vidović, S. Application of conventional an high-pressure extraction techniques for the isolation of bioactive compounds from the aerial part of hemp (Cannabis sativa L.) assortment Helena. Ind. Crops Prod. 2021, 171, 113908. [Google Scholar] [CrossRef]
- Kumar, P.; Mahato, D.K.; Kamle, M.; Borah, R.; Sharma, B.; Pandhi, S.; Vijay, T.; Yadav, H.S.; Devi, S.; Patil, U.; et al. Pharmacological properties, therapeutic potential, and legal status of Cannabis sativa L.: An overview. Phytother. Res. 2021, 35, 6010–6029. [Google Scholar] [CrossRef] [PubMed]
- Shikanai, A.; Gage, K.L. Allelopathic Potential of Hemp: Implications for Integrated Weed Management. Sec. Weed Manag. 2022, 4, 832471. [Google Scholar] [CrossRef]
- Mushtaq, W.; Siddiqui, M.B.; Hakeem, K.R. Allelopathy: Potential for Green Agriculture; Springer: Cham, Switzerland, 2020; pp. 1–25. [Google Scholar] [CrossRef]
- Bachheti, A.; Sharma, S.; Bachheti, R.K.; Husen, A.; Pandey, D.P. Plant allelochemicals and their various applications. In Co-Evolution of Secondary Metabolites; Mérillon, J.M., Ramawat, K., Eds.; Springer: Cham, Switzerland, 2020; pp. 441–457. [Google Scholar] [CrossRef]
- Scavo, A.; Restuccia, A.; Mauromicale, G. Allelopathy: Principles and basic aspects for agroecosystem control. In Sustainable Agriculture Reviews 28: Ecology for Agriculture; Springer: Cham, Switzerland, 2018; pp. 47–101. [Google Scholar] [CrossRef]
- Palanivel, H.; Tilaye, G.; Belliathan, S.K.; Benor, S.; Abera, S.; Kamaraj, M. Allelochemicals as natural herbicides for sustainable agriculture to promote a cleaner environment. In Strategies and Tools for Pollutant Mitigation; Aravind, J., Kamaraj, M., Devi, M.P., Rajakumar, S., Eds.; Springer: Cham, Switzerland, 2021; pp. 93–110. [Google Scholar] [CrossRef]
- Clarke, R.C.; Merlin, M.D. Cannabis domestication, breeding history, present-day genetic diversity, and future prospects. Crit. Rev. Plant Sci. 2016, 35, 293–327. [Google Scholar] [CrossRef]
- Ferus, P.; Mencik, K.; Konôpková, J. Allelopathic potential of Juglans nigra L. to control the invasive tree-of-heaven (Ailanthus altissima (Mill.) Swingle). Allelopath. J. 2020, 49, 177–188. [Google Scholar] [CrossRef]
- Ravlić, M.; Baličević, R.; Lucić, I. Allelopathic effect of parsley (Petroselinum crispum Mill.) cogermination, water extracts and residues on hoary cress (Lepidium draba (L.) Desv.). Poljoprivreda 2014, 20, 22–26. [Google Scholar]
- Mahmoodzadeh, H.; Ghasemi, M.; Zanganeh, H. Allelopathic effect of medicinal plant Cannabis sativa L. on Lactuca sativa L. seed germination. Acta Agric. Slov. 2015, 105, 233–239. [Google Scholar] [CrossRef]
- Das, C.; Dey, A.; Bandyopadhyay, A. Allelochemicals: An emerging tool for weed management. In Evidence Based Validation of Traditional Medicines: A Comprehensive Approach; Springer: Cham, Switzerland, 2021; pp. 249–259. [Google Scholar] [CrossRef]
- Šćepanović, M.; Sarić-Krsmanović, M.; Šoštarčić, V.; Brijačak, E.; Lakić, J.; Špirović Trifunović, B.; Gajić Umiljendić, J.; Radivojević, L.J. Inhibitory Effects of Brassicaceae Cover Crop on Ambrosia artemisiifolia Germination and Early Growth. Plants 2021, 10, 794. [Google Scholar] [CrossRef]
- Prvulović, D.; Gvozdenac, S.; Latković, D.; Peić Tukuljac, M.; Sikora, V.; Kiprovski, B.; Mišan, A.; Chrysargyris, A.; Tzortzakis, N.; Ovuka, J. Phytotoxic and insecticidal activity of industrial hemp (Cannabis sativa L.) extracts against Plodia interpunctella Hübner—A potential sunflower grain protectant. Agronomy 2023, 13, 2456. [Google Scholar] [CrossRef]
- Patanè, C.; Pellegrino, A.; Cosentino, S.L.; Testa, G. Allelopathic effects of Cannabis sativa L. aqueous leaf extracts on seed germination and seedling growth in durum wheat and barley. Agronomy 2023, 13, 454. [Google Scholar] [CrossRef]
- Pudełko, K.; Majchrzak, L.; Narožna, D. Allelopathic effect of fibre hemp (Cannabis sativa L.) on monocot and dicot plant species. Ind. Crops Prod. 2014, 56, 191–199. [Google Scholar] [CrossRef]
- Poonsawat, T.; Srilasak, N.; Koodkaew, I. Allelopathic characterization and allelochemicals identification of hemp (Cannabis sativa L.) leaf residue. Ind. Crops Prod. 2024, 222, 120003. [Google Scholar] [CrossRef]
- Singh, N.B.; Thapar, R. Allelopathic influence of Cannabis sativa on growth and metabolism of Parthenium hysterophorus. Allelopath. J. 2003, 12, 61–70. [Google Scholar]
- Wu, H.; Haig, T.; Pratley, J.; Lemerle, D.; An, M. Distribution and exudation of allelochemicals in wheat Triticum aestivum. J. Chem. Ecol. 2000, 26, 2141–2154. [Google Scholar] [CrossRef]
- Vinatoru, M.; Mason, T.J.; Calinescu, I. Ultrasonically assisted extraction (UAE) and microwave assisted extraction (MAE) of functional compounds from plant materials. TrAC Trends Anal. Chem. 2017, 97, 159–178. [Google Scholar] [CrossRef]
- Wang, C.; Qi, J.; Liu, Q.; Wang, Y.; Wang, H. Allelopathic potential of aqueous extracts from fleagrass (Adenosma buchneroides Bonati) against two crop and three weed species. Agriculture 2022, 12, 1103. [Google Scholar] [CrossRef]
- Bocsa, I.; Karus, M. The Cultivation of Hemp: Botany, Varieties, Cultivation and Harvesting; Hemptech: Sebastopol, CA, USA, 1998; pp. 1–184. ISBN 978-1-886874-03-9. [Google Scholar]
- Callaway, J.C. A more reliable evaluation of hemp THC levels is necessary and possible. J. Ind. Hemp. 2008, 13, 117–144. [Google Scholar] [CrossRef]
- Mišan, A.; Mimica-Dukić, N.; Mandić, A.; Sakač, M.; Milovanović, I.; Sedej, I. Development of a rapid resolution HPLC method for the separation and determination of 17 phenolic compounds in crude plant extracts. Open Chem. 2011, 9, 133–142. [Google Scholar] [CrossRef]
- Poortman-Van Der Meer, A.J.; Huizer, H. A contribution to the improvement of accuracy in the quantitation of THC. Forensic Sci. Int. 1999, 101, 1–8. [Google Scholar] [CrossRef]
- Beleggia, R.; Iannucci, A.; Menga, V.; Quitadamo, F.; Suriano, S.; Citti, C.; Pecchioni, N.; Trono, D. Impact of chitosan-based foliar application on the phytochemical content and the antioxidant activity in hemp (Cannabis sativa L.) inflorescences. Plants 2023, 12, 3692. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Henry, P.; Shan, J.; Chen, J. Identification of chemotypic markers in three chemotype categories of cannabis using secondary metabolites profiled in inflorescences, leaves, stem bark, and roots. Front. Plant Sci. 2021, 12, 699530. [Google Scholar] [CrossRef] [PubMed]
- Lorizola, I.M.; Furlan CP, B.; Portovedo, M.; Milanski, M.; Botelho, P.B.; Bezerra RM, N.; Sumere, B.R.; Rostagno, M.A.; Capitani, C.D. Beet stalks and leaves (Beta vulgaris L.) protect against high-fat diet-induced oxidative damage in the liver in mice. Nutrients 2018, 10, 872. [Google Scholar] [CrossRef]
- de Freitas Marinho, L.; Sganzerla, W.G.; Ferreira, V.C.; Moreno, J.A.J.; Rostagno, M.A.; Forster-Carneiro, T. Advances in green extraction methods, biological properties, and applications of betanin and vitexin: An updated review and bibliometric analysis. Biocatal. Agric. Biotechnol. 2023, 51, 102744. [Google Scholar] [CrossRef]
- Izzo, L.; Pacifico, S.; Piccolella, S.; Castaldo, L.; Narváez, A.; Grosso, M.; Ritieni, A. Chemical analysis of minor bioactive components and cannabidiolic acid in commercial hemp seed oil. Molecules 2020, 25, 3710. [Google Scholar] [CrossRef]
- Abdalla, M.M. The potential of Moringa oleifera extract as a biostimulant in enhancing the growth, biochemical and hormonal contents in rocket (Eruca vesicaria subsp. sativa) plants. Int. J. Plant Physiol. Biochem. 2013, 5, 42–49. [Google Scholar] [CrossRef]
- Drinić, Z.; Vidović, S.; Vladić, J.; Koren, A.; Kiprovski, B.; Sikora, V. Effect of extraction solvent on total polyphenols content and antioxidant activity of Cannabis sativa L. Lek. Sirovine 2018, 38, 17–21. [Google Scholar] [CrossRef]
- Gallo, M.; Formato, A.; Ciaravolo, M.; Formato, G.; Naviglio, D. Study of the kinetics of extraction process for the production of hemp inflorescences extracts by means of conventional maceration (CM) and rapid solid-liquid dynamic extraction (RSLDE). Separations 2020, 7, 20. [Google Scholar] [CrossRef]
- Palmieri, S.; Pellegrini, M.; Ricci, A.; Compagnone, D.; Lo Sterzo, C. Chemical composition and antioxidant activity of thyme, hemp and coriander extracts: A comparison study of maceration, Soxhlet, UAE and RSLDE techniques. Foods 2020, 9, 1221. [Google Scholar] [CrossRef]
- Lewis-Bakker, M.M.; Yang, Y.; Vyawahare, R.; Kotra, L.P. Extractions of medical cannabis cultivars and the role of decarboxylation in optimal receptor responses. Cannabis Cannabinoid Res. 2019, 4, 183–194. [Google Scholar] [CrossRef]
- Drinić, Z.; Vladić, J.; Koren, A.; Zeremski, T.; Stojanov, N.; Kiprovski, B.; Vidović, S. Microwave-assisted extraction of cannabinoids and antioxidants from Cannabis sativa aerial parts and process modeling. J. Chem. Technol. Biotechnol. 2020, 95, 831–839. [Google Scholar] [CrossRef]
- Agarwal, C.; Máthé, K.; Hofmann, T.; Csóka, L. Ultrasound-assisted extraction of cannabinoids from Cannabis sativa L. optimized by response surface methodology. J. Food Sci. 2018, 83, 700–710. [Google Scholar] [CrossRef]
- Dong, X.; Li, X.; Ruan, X.; Kong, L.; Wang, N.; Gao, W.; Wang, R.; Sun, Y.; Jin, M. A deep insight into the structure-solubility relationship and molecular interaction mechanism of diverse flavonoids in molecular solvents, ionic liquids, and molecular solvent/ionic liquid mixtures. J. Mol. Liq. 2023, 385, 122359. [Google Scholar] [CrossRef]
- Bouras, M.; Chadni, M.; Barba, F.J.; Grimi, N.; Bals, O.; Vorobiev, E. Optimization of microwave-assisted extraction of polyphenols from Quercus bark. Ind. Crops Prod. 2015, 77, 590–601. [Google Scholar] [CrossRef]
- He, M.; Min, J.W.; Kong, W.L.; He, X.H.; Li, J.X.; Peng, B.W. A review on the pharmacological effects of vitexin and isovitexin. Fitoterapia 2016, 115, 74–85. [Google Scholar] [CrossRef]
- Chen, F.; Zhang, Q.; Liu, J.; Gu, H.; Yang, L. An efficient approach for the extraction of orientin and vitexin from Trollius chinensis flowers using ultrasonic circulating technique. Ultrason. Sonochem. 2017, 37, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, A.; Turner, C. Pressurized liquid extraction as a green approach in food and herbal plants extraction: A review. Anal. Chim. Acta 2011, 703, 8–18. [Google Scholar] [CrossRef]
- Miron, T.L.; Plaza, M.; Bahrim, G.; Ibáñez, E.; Herrero, M. Chemical composition of bioactive pressurized extracts of Romanian aromatic plants. J. Chromatogr. A 2011, 1218, 4918–4927. [Google Scholar] [CrossRef]
- Balachandran, S.; Kentish, S.E.; Mawson, R.; Ashokkumar, M. Ultrasonic enhancement of the supercritical extraction from ginger. Ultrason. Sonochem. 2006, 13, 471–479. [Google Scholar] [CrossRef]
- Meng, Q.; Buchanan, B.; Zuccolo, J.; Poulin, M.M.; Gabriele, J.; Baranowski, D.C. A reliable and validated LC-MS/MS method for the simultaneous quantification of 4 cannabinoids in 40 consumer products. PLoS ONE 2018, 13, e0196396. [Google Scholar] [CrossRef]
- De Martino, L.D.; Mancini, E.; de Almeida LF, R.; Feo, V.D. The antigerminative activity of twenty-seven monoterpenes. Molecules 2010, 15, 6630–6637. [Google Scholar] [CrossRef] [PubMed]
- Shikanai, A. Understanding the Role of Secondary Metabolites in Hemp-Weed Interactions. Master’s Thesis, Southern Illinois University, Carbondale, IL, USA, 2021. [Google Scholar]
- Sirikantaramas, S.; Taura, F.; Tanaka, Y.; Ishikawa, Y.; Morimoto, S.; Shoyama, Y. Tetrahydrocannabinolic acid synthase, the enzyme controlling marijuana psychoactivity, is secreted into the storage cavity of the glandular trichomes. Plant Cell Physiol. 2005, 46, 1578–1582. [Google Scholar] [CrossRef]
- De Vita, D.; Madia, V.N.; Tudino, V.; Saccoliti, F.; De Leo, A.; Messore, A.; Roscilli, P.; Botto, A.; Pindinello, I.; Santilli, G.; et al. Comparison of different methods for the extraction of cannabinoids from cannabis. Nat. Prod. Res. 2020, 34, 2952–2958. [Google Scholar] [CrossRef]
- Malingre, T.; Hendriks, H.; Batterman, S.; Bos, R.; Visser, J. The essential oil of Cannabis sativa. Planta Med. 1975, 28, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Mediavilla, V.; Steinemann, S. Essential oil of Cannabis sativa L. strains. J. Int. Hemp Assoc. 1997, 4, 80–82. [Google Scholar] [CrossRef]
- Chang, C.W.; Yen, C.C.; Wu, M.T.; Hsu, M.C.; Wu, Y.T. Microwave-assisted extraction of cannabinoids in hemp nut using response surface methodology: Optimization and comparative study. Molecules 2017, 22, 1894. [Google Scholar] [CrossRef]
- Brighenti, V.; Pellati, F.; Steinbach, M.; Maran, D.; Benvenuti, S. Development of a new extraction technique and HPLC method for the analysis of non-psychoactive cannabinoids in fibre-type Cannabis sativa L.(hemp). J. Pharm. Biomed. Anal. 2017, 143, 228–236. [Google Scholar] [CrossRef]
- Tanase, C.; Bujor, O.C.; Popa, V.I. Influence on physiological processes in plants. In Polyphenols in Plants: Isolation, Purification and Extract Preparation; Watson, R.R., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 45–55. [Google Scholar] [CrossRef]
- Hegab, M.M.; Khodary SE, A.; Hammouda, O.; Ghareib, H.R. Autotoxicity of chard and its allelopathic potentiality on germination and some metabolic activities associated with growth of wheat seedlings. Afr. J. Biotechnol. 2008, 7, 884–892. [Google Scholar]
- Gharib, F.A.; Hegazi, A.Z. Salicylic acid ameliorates germination, seedling growth, phytohormone and enzymes activity in bean (Phaseolus vulgaris L.) under cold stress. J. Am. Sci. 2010, 6, 675–683. [Google Scholar]
- Putnam, A.R. Allelochemicals from plants as herbicides. Weed Technol. 1988, 2, 510–518. [Google Scholar] [CrossRef]
- Scavo, A.; Pandino, G.; Restuccia, A.; Lombardo, S.; Pesce, G.R.; Mauromicale, G. Allelopathic potential of leaf aqueous extracts from Cynara cardunculus L. on the seedling growth of two cosmopolitan weed species. Ital. J. Agron. 2019, 14, 1373. [Google Scholar] [CrossRef]
- Sadeghi, H.; Khazaei, F.; Sheidaei, S.; Yari, L. Effect of seed size on seed germination behavior of safflower (Carthamus tinctorius L.). ARPN J. 2011, 6, 5–8. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hemp Extracts | Content of Phenolic Compounds (mg L−1) | |||||||
|---|---|---|---|---|---|---|---|---|
| Ferulic Acid | Orientin | Vitexin | Luteolin Derivative 1 | Isovitexin | Apigenin Derivative | Luteolin Derivative 2 | Total Polyphenols | |
| MAC | 4.648 | - | 5.42 | - | - | - | - | 10.068 |
| ±SD | 0.004 | 0.043 | ||||||
| PDR | 5.798 | 9.913 | 10.171 | 12.997 | - | 8.076 | 5.830 | 52.785 |
| ±SD | 0.205 | 0.02 | 0.002 | 0.055 | 0.005 | 0.016 | ||
| HYD | - | - | - | - | - | - | - | - |
| UAE30 | 5.059 | 6.717 | 15.051 | 4.554 | 2.9 | 4.48 | 4.887 | 43.648 |
| ±SD | 0 | 0.119 | 0.413 | 0.084 | 0.102 | 0.047 | 0.017 | |
| UAE60 | 4.635 | 12.607 | 18.047 | 8.668 | 2.156 | 6.57 | 5.062 | 57.745 |
| ±SD | 0.075 | 0.034 | 0.094 | 0.105 | 0.049 | 0.048 | 0.068 | |
| MAE210 | 5.227 | 14.923 | 20.093 | 17.194 | 1.881 | 14.475 | 7.705 | 81.498 |
| ±SD | 0.012 | 0.420 | 0.199 | 0.01 | 0 | 0.042 | 0.157 | |
| MAE700 | 5.716 | 16.777 | 20.175 | 19.126 | 2.096 | 14.632 | 8.597 | 87.119 |
| ±SD | 0.185 | 0.119 | 0.224 | 0.021 | 0.052 | 0.013 | 0.008 | |
| Hemp Extracts | CBD Content (mg mL−1) | THC Content (mg mL−1) |
|---|---|---|
| MAC | 0.189 ± 0.012 | 0.098 ± 0.011 |
| PDR | 0.081 ± 0.005 | 0.050 ± 0.003 |
| HYD | 0.010 ± 0.003 | - |
| UAE30 | 0.075 ± 0.004 | 0.045 ± 0.001 |
| UAE60 | 0.215 ± 0.012 | 0.097 ± 0.003 |
| MAE210 | 0.072 ± 0.006 | 0.039 ± 0.004 |
| MAE700 | 0.074 ± 0.006 | 0.045 ± 0.005 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kojić, M.; Samardžić, N.; Popov, M.; Gavarić, A.; Vidović, S.; Teslić, N.; Zeremski, T.; Koren, A.; Konstantinović, B. Impact of Industrial Hemp (Cannabis sativa L.) Extracts on Seed Germination and Seedling Growth: Evaluating Allelopathic Activity Across Various Extraction Methods. Agronomy 2025, 15, 684. https://doi.org/10.3390/agronomy15030684
Kojić M, Samardžić N, Popov M, Gavarić A, Vidović S, Teslić N, Zeremski T, Koren A, Konstantinović B. Impact of Industrial Hemp (Cannabis sativa L.) Extracts on Seed Germination and Seedling Growth: Evaluating Allelopathic Activity Across Various Extraction Methods. Agronomy. 2025; 15(3):684. https://doi.org/10.3390/agronomy15030684
Chicago/Turabian StyleKojić, Mirjana, Nataša Samardžić, Milena Popov, Aleksandra Gavarić, Senka Vidović, Nemanja Teslić, Tijana Zeremski, Anamarija Koren, and Bojan Konstantinović. 2025. "Impact of Industrial Hemp (Cannabis sativa L.) Extracts on Seed Germination and Seedling Growth: Evaluating Allelopathic Activity Across Various Extraction Methods" Agronomy 15, no. 3: 684. https://doi.org/10.3390/agronomy15030684
APA StyleKojić, M., Samardžić, N., Popov, M., Gavarić, A., Vidović, S., Teslić, N., Zeremski, T., Koren, A., & Konstantinović, B. (2025). Impact of Industrial Hemp (Cannabis sativa L.) Extracts on Seed Germination and Seedling Growth: Evaluating Allelopathic Activity Across Various Extraction Methods. Agronomy, 15(3), 684. https://doi.org/10.3390/agronomy15030684