
Adaptation of Diverse Maize Germplasm to Spring Season Conditions in Northeast China



Abstract
1. Introduction
2. Materials and Methods
2.1. Genetic Materials
2.2. Experimental Design and Data Collection
2.3. Statistical Analysis
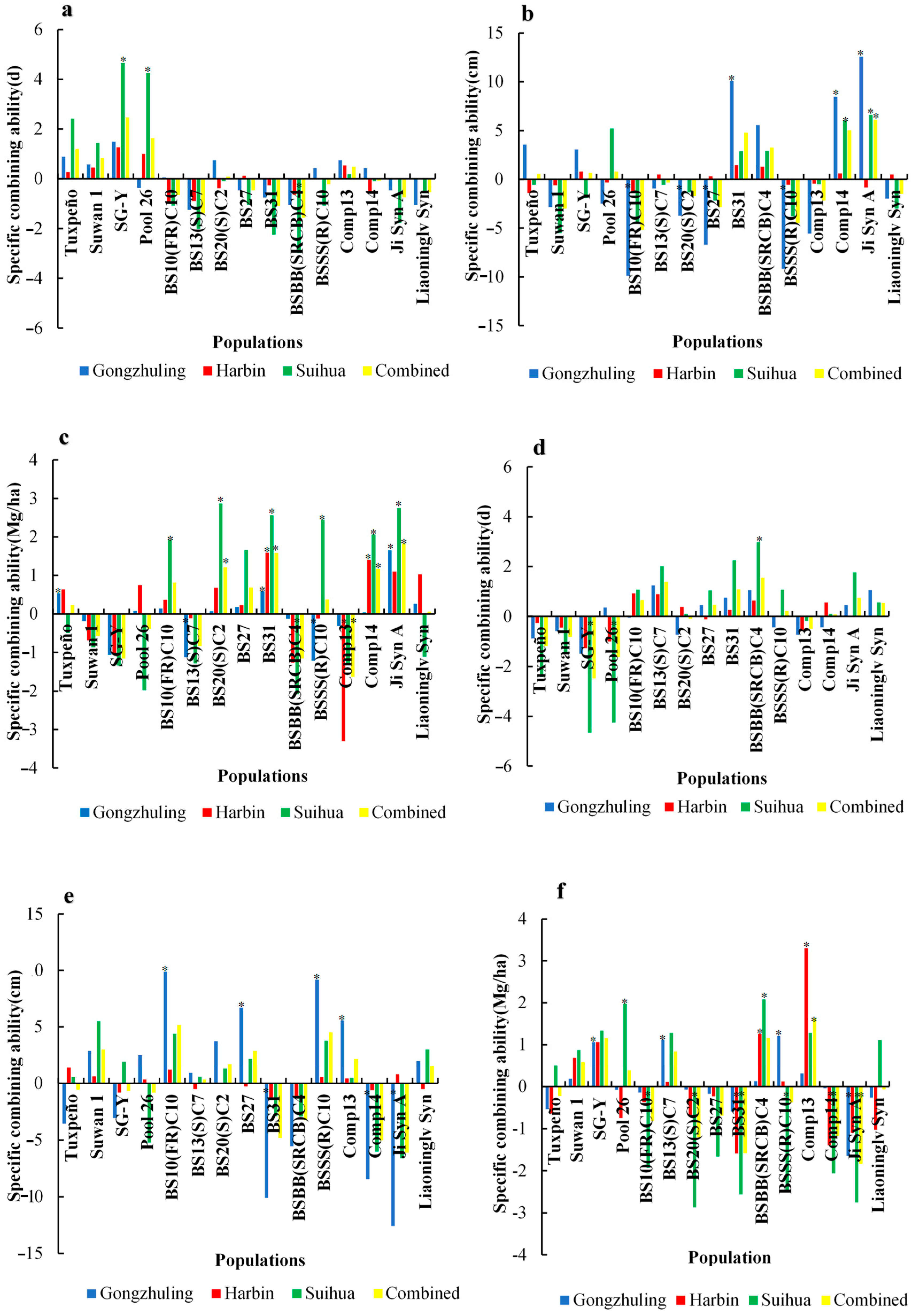
3. Results
3.1. Analysis of Variance
3.2. Days to Silking
3.3. Ear Height
3.4. Stalk Lodging
3.5. Grain Yield
4. Discussion
4.1. Combining Ability and Heterosis of 15 Diverse Populations
4.2. Germplasm Improvement and Utilization
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Source of Variation | d.f. | DS | EH | SL | GY | Source of Variation | d.f. | DS | EH | SL | GY |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Combined | Harbin | ||||||||||
| Genotypes | 29 | 4.91 *** | 3.08 *** | 2.61 *** | 2.51 *** | Genotypes | 29 | 2.07 ** | 2.62 *** | 1.63 ** | 2.64 *** |
| Environment (Envi.) | 5 | 364.39 * | 47.71 ** | 9.76 ** | 102.19 ** | Environment (Envi.) | 1 | 12.35 * | 81.83 ** | 6.19 | 192.00 * |
| Population | 14 | 3.20 *** | 3.04 *** | 2.18 ** | 3.45 *** | Population | 14 | 2.97 *** | 3.43 *** | 1.84 ** | 2.27 ** |
| Tester | 1 | 3.74 * | 0.91 * | 13.79 *** | 3.63 * | Tester | 1 | 3.24 ** | 14.48 *** | 4.15 * | 13.22 *** |
| Population × Tester | 14 | 3.30 ** | 2.24 *** | 1.12 | 1.15 *** | Population × Tester | 14 | 0.49 *** | 0.43 *** | 1.17 | 0.76 ** |
| Genotypes × Envi. | 145 | 2.72 *** | 2.35 ** | 1.63 ** | 1.65 *** | Genotypes × Envi. | 29 | 1.05 ** | 0.37 *** | 0.45 ** | 1.37 ** |
| Population × Envi. | 70 | 1.40 ** | 0.77 * | 0.79 * | 1.62 ** | Population × Envi. | 14 | 1.02 ** | 0.61 ** | 0.37 ** | 1.35 *** |
| Tester × Envi. | 5 | 18.45 ** | 26.48 ** | 13.93 * | 11.02 *** | Tester × Envi. | 1 | 1.54 ** | 0.44 ** | 1.58 | 4.98 * |
| Population × Tester × Envi. | 14 | 0.70 ** | 0.41 | 0.86 | 0.63 ** | Population × Tester × Envi. | 14 | 0.81 ** | 0.19 ** | 0.43 ** | 0.63 ** |
| δ2A | 6.94 | 3.95 | 15.97 | 7.08 | δ2A | 6.21 | 17.91 | 5.99 | 15.49 | ||
| δ2D | 3.30 | 2.24 | 1.12 | 1.15 | δ2D | 0.49 | 0.43 | 1.17 | 0.76 | ||
| H2b | 0.07 | 0.27 | 0.61 | 0.12 | H2b | 0.83 | 0.48 | 0.88 | 0.29 | ||
| H2n | 0.02 | 0.13 | 0.36 | 0.09 | H2n | 0.62 | 0.40 | 0.56 | 0.17 | ||
| Gongzhuling | Suihua | ||||||||||
| Genotypes | 29 | 3.79 *** | 3.52 *** | 2.31 ** | 1.89 * | Genotypes | 29 | 7.10 *** | 2.18 ** | 1.86 ** | 1.94 ** |
| Environment (Envi.) | 1 | 101.21 * | 90.16 * | 0.05 | 6.44 ** | Environment (Envi.) | 1 | 7.28 ** | 84.74 ** | 65.94 * | 4.00 * |
| Population | 14 | 2.43 ** | 3.22 *** | 1.74 * | 1.92 ** | Population | 14 | 9.15 *** | 3.24 *** | 1.99 ** | 3.00 *** |
| Tester | 1 | 29.77 *** | 19.46 *** | 15.21 * | 10.37 ** | Tester | 1 | 15.29 *** | 1.36 ** | 0.71 * | 0.06 ** |
| Population × Tester | 14 | 0.80 ** | 0.72 * | 1.46 * | 0.69 ** | Population × Tester | 14 | 0.98 ** | 0.13 * | 0.41 | 0.27 ** |
| Genotypes × Envi. | 29 | 3.54 *** | 2.07 * | 1.01 * | 3.56 ** | Genotypes × Envi. | 29 | 1.47 *** | 0.47 * | 1.93 ** | 1.61 *** |
| Population × Envi. | 14 | 0.48 ** | 0.44 ** | 0.32 ** | 3.84 *** | Population × Envi. | 14 | 2.84 ** | 0.30 ** | 2.40 ** | 2.52 ** |
| Tester × Envi. | 1 | 47.91 ** | 29.71 *** | 10.35 ** | 36.91 *** | Tester × Envi. | 1 | 0.12 ** | 0.04 *** | 6.02 ** | 0.04 ** |
| Population × Tester × Envi. | 14 | 1.61 ** | 0.48 ** | 0.98 * | 0.47 ** | Population × Tester × Envi. | 14 | 0.18 ** | 0.71 ** | 0.75 ** | 1.03 ** |
| δ2A | 32.2 | 22.68 | 16.95 | 12.29 | δ2A | 24.44 | 4.60 | 2.70 | 3.06 | ||
| δ2D | 0.80 | 0.72 | 1.46 | 0.69 | δ2D | 0.98 | 0.13 | 0.41 | 0.27 | ||
| H2b | 0.52 | 0.53 | 0.99 | 0.89 | H2b | 0.97 | 0.43 | 0.45 | 0.93 | ||
| H2n | 0.30 | 0.34 | 0.55 | 0.61 | H2n | 0.67 | 0.32 | 0.24 | 0.70 |
| Population | DS (d) | EH (cm) | ||||||||||||||
| Gongzhuling | Harbin | Suihua | Combined | Gongzhuling | Harbin | Suihua | Combined | |||||||||
| PH4CV | PH6WC | PH4CV | PH6WC | PH4CV | PH6WC | PH4CV | PH6WC | PH4CV | PH6WC | PH4CV | PH6WC | PH4CV | PH6WC | PH4CV | PH6WC | |
| Tuxpeño | 85.83 # | 81.50 ¶ | 87.83 ¶ | 87.00 ¶ | 96.33 # | 96.00 # | 90.00 # | 88.17 ¶ | 148.28 # | 134.73 ¶ | 146.12 # | 139.42 ¶ | 131.63 ¶ | 136.53 ¶ | 136.68 ¶ | 136.89 ¶ |
| Suwan 1 | 85.16 ¶ | 81.00 ¶ | 88.17 ¶ | 89.33 # | 95.17 # | 99.00 # | 89.50 # | 89.78 # | 151.22 # | 127.82 ¶ | 145.00 # | 152.92 # | 142.62 # | 147.32 # | 142.61 ¶ | 142.69 ¶ |
| SG-Y | 87.16 # | 83.00 ¶ | 89.67 # | 90.00 # | 99.00 # | 100.17 # | 91.94 # | 91.06 # | 156.55 # | 131.65 ¶ | 145.22 # | 135.57 ¶ | 142.53 # | 139.81 ¶ | 144.77 ¶ | 135.68 ¶ |
| PooL 26 | 82.67 ¶ | 81.98 ¶ | 90.00 # | - † | 99.33 # | - | 90.67 # | - | 156.10 # | 158.41 # | 142.67 ¶ | - | 157.33 # | - | 148.70 # | - |
| BS10(FR)C10 | 84.00 ¶ | 83.67 ¶ | 85.67 ¶ | 87.33 ¶ | 92.17 ¶ | 95.33 # | 87.28 ¶ | 88.78 ¶ | 133.08 ¶ | 127.95 ¶ | 132.08 ¶ | 130.75 ¶ | 125.97 ¶ | 136.12 ¶ | 124.38 ¶ | 131.61 ¶ |
| BS13(S)C7 | 79.67 ¶ | 81.02 ¶ | 85.00 ¶ | - | 90.67 ¶ | - | 85.11 ¶ | - | 140.83 ¶ | 143.63 ¶ | 134.67 ¶ | - | 116.43 ¶ | - | 123.98 ¶ | - |
| BS20(S)C2 | 85.50 # | 84.00 ¶ | 86.67 ¶ | 87.33 ¶ | 93.33 # | 94.00 # | 88.50 ¶ | 88.44 ¶ | 140.31 ¶ | 122.27 ¶ | 123.40 ¶ | 124.47 ¶ | 132.98 ¶ | 138.62 ¶ | 132.23 ¶ | 128.45 ¶ |
| BS27 | 82.33 ¶ | 82.12 ¶ | 87.67 ¶ | - | 92.00 ¶ | - | 87.33 ¶ | - | 149.50 # | 149.84 # | 133.00 ¶ | - | 134.07 ¶ | - | 128.86 ¶ | - |
| BS31 | 81.33 ¶ | 80.98 ¶ | 86.67 ¶ | - | 90.33 ¶ | - | 86.11 ¶ | - | 141.33 ¶ | 143.33 ¶ | 128.67 ¶ | - | 131.50 ¶ | - | 138.17 ¶ | - |
| BSBB(SRCB)C4 | 80.33 ¶ | 80.17 ¶ | 85.67 ¶ | - | 89.33 ¶ | - | 85.11 ¶ | - | 137.17 ¶ | 138.37 ¶ | 127.33 ¶ | - | 134.27 ¶ | - | 129.59 ¶ | - |
| BSSS(R)C10 | 84.83 ¶ | 82.00 ¶ | 87.33 ¶ | 87.50 ¶ | 92.17 ¶ | 93.80 # | 88.11 ¶ | 87.77 ¶ | 133.90 ¶ | 124.78 ¶ | 129.40 ¶ | 124.12 ¶ | 127.38 ¶ | 131.62 ¶ | 126.89 ¶ | 126.84 ¶ |
| Comp13 | 85.50 # | 82.00 ¶ | 88.33 ¶ | 89.83 # | 93.67 # | 96.83 # | 89.17 # | 89.55 # | 143.60 ¶ | 133.73 ¶ | 127.60 ¶ | 133.98 ¶ | 134.70 ¶ | 143.20 # | 135.30 ¶ | 136.97 ¶ |
| Comp14 | 84.83 ¶ | 81.33 ¶ | 86.33 ¶ | 88.00 ¶ | 93.33 # | 94.17 # | 88.16 ¶ | 87.83 ¶ | 136.82 ¶ | 132.40 ¶ | 126.07 ¶ | 129.05 ¶ | 131.05 ¶ | 137.08 ¶ | 131.31 ¶ | 132.84 ¶ |
| Ji Syn A | 82.33 ¶ | 81.78 ¶ | 87.33 ¶ | - | 91.00 ¶ | - | 86.89 ¶ | - | 161.67 # | 162.70 # | 121.67 ¶ | - | 159.47 # | - | 144.27 # | - |
| Liaoninglv Syn | 80.33 ¶ | 80.19 ¶ | 87.33 ¶ | 87.00 ¶ | 92.67 ¶ | - | 86.78 ¶ | - | 142.03 ¶ | 144.19 ¶ | 131.00 ¶ | 126.33 ¶ | 125.37 ¶ | - | 132.80 ¶ | - |
| SD | 82.24 ± 2.29 | 82.00 ± 2.69 | 66.99 ± 37.22 | 72.59 ± 38.02 | 87.01 ± 6.34 | 91.04 ± 7.15 | 88.04 ± 1.97 | 88.92 ± 1.14 | 131.02 ± 21.49 | 134.75 ± 21.67 | 123.86 ± 17.77 | 137.59 ± 19.74 | 116.70 ± 16.32 | 126.69 ± 21.01 | 134.70 ± 7.71 | 133.99 ± 5.14 |
| Xianyu 335 | 85.33 | 88.67 | 92.80 | 88.93 | 145.17 | 144.67 | 142.33 | 144.05 | ||||||||
| Population | SL (%) | GY (t/ha) | ||||||||||||||
| Gongzhuling | Harbin | Suihua | Combined | Gongzhuling | Harbin | Suihua | Combined | |||||||||
| PH4CV | PH6WC | PH4CV | PH6WC | PH4CV | PH6WC | PH4CV | PH6WC | PH4CV | PH6WC | PH4CV | PH6WC | PH4CV | PH6WC | PH4CV | PH6WC | |
| Tuxpeño | 5.00 # | 1.00 ¶ | 9.00 # | 8.00 # | 6.00 # | 4.00 # | 8.67 # | 4.67 # | 9.20 ¶ | 9.74 ¶ | 8.20 ¶ | 8.03 ¶ | 6.37 ¶ | 6.64 ¶ | 7.92 ¶ | 8.14 ¶ |
| Suwan 1 | 3.00 # | 1.00 ¶ | 4.00 # | 4.00 # | 3.00 # | 2.00 # | 3.67 # | 2.33 # | 10.01 ¶ | 9.14 ¶ | 8.58 ¶ | 9.55 ¶ | 7.00 ¶ | 6.97 ¶ | 8.53 ¶ | 8.55 ¶ |
| SG-Y | 2.00 # | 3.00 # | 6.00 # | 7.00 # | 4.00 # | 2.00 # | 4.00 # | 4.00 # | 9.61 ¶ | 8.77 ¶ | 8.10 ¶ | 7.79 ¶ | 5.94 ¶ | 6.31 ¶ | 7.88 ¶ | 7.62 ¶ |
| PooL 26 | 4.00 # | 4.00 # | 3.00 # | 0.00 ¶ | 0.00 ¶ | 0.00 ¶ | 4.67 # | 1.33 ¶ | 10.31 ¶ | 10.46 ¶ | 7.97 ¶ | 10.89 # | 6.81 ¶ | - | 8.36 ¶ | - |
| BS10(FR)C10 | 2.00 # | 0.00 ¶ | 3.00 # | 3.00 # | 8.00 # | 3.00 # | 4.33 # | 2.00 # | 10.01 ¶ | 8.36 ¶ | 8.92 ¶ | 9.77 ¶ | 7.03 ¶ | 8.12 ¶ | 8.65 ¶ | 8.75 ¶ |
| BS13(S)C7 | 2.00 # | 2.00 # | 2.00 # | 0.00 ¶ | 0.00 ¶ | 0.00 ¶ | 1.33 ¶ | 0.67 ¶ | 9.11 ¶ | 9.23 ¶ | 9.40 ¶ | 11.82 # | 7.65 ¶ | - | 8.72 ¶ | - |
| BS20(S)C2 | 6.00 # | 2.00 # | 12.00 # | 5.00 # | 3.00 # | 4.00 # | 7.00 # | 3.67 # | 10.19 ¶ | 9.57 ¶ | 9.32 ¶ | 9.40 ¶ | 7.95 ¶ | 7.84 ¶ | 9.15 ¶ | 8.94 ¶ |
| BS27 | 9.00 # | 9.00 # | 23.00 # | 0.00 ¶ | 0.00 ¶ | 0.00 ¶ | 10.67 # | 3.00 # | 9.77 ¶ | 9.87 ¶ | 8.33 ¶ | 11.77 # | 7.54 ¶ | - | 8.55 ¶ | - |
| BS31 | 1.00 ¶ | 1.00 ¶ | 2.00 # | 0.00 ¶ | 0.00 ¶ | 0.00 ¶ | 1.00 ¶ | 0.33 ¶ | 10.05 ¶ | 10.21 ¶ | 9.16 ¶ | 11.62 # | 8.82 # | - | 9.34 ¶ | - |
| BSBB(SRCB)C4 | 5.00 # | 5.00 # | 7.00 # | 0.00 ¶ | 0.00 ¶ | 0.00 ¶ | 4.00 # | 1.67 # | 8.24 ¶ | 8.37 ¶ | 9.67 ¶ | 11.86 # | 8.34 ¶ | - | 8.75 ¶ | - |
| BSSS(R)C10 | 10.00 # | 1.00 ¶ | 12.00 # | 4.00 # | 8.00 # | 5.00 # | 10.00 # | 3.33 # | 9.08 ¶ | 9.29 ¶ | 8.14 ¶ | 9.24 ¶ | 7.90 ¶ | 8.11 ¶ | 8.37 ¶ | 8.88 ¶ |
| Comp13 | 4.00 # | 1.00 ¶ | 6.00 # | 7.00 # | 2.00 # | 1.00 ¶ | 4.00 # | 3.00 # | 11.10 ¶ | 9.49 ¶ | 9.43 ¶ | 8.38 ¶ | 7.15 ¶ | 7.21 ¶ | 9.23 ¶ | 8.36 ¶ |
| Comp14 | 7.00 # | 0.00 ¶ | 9.00 # | 1.00 ¶ | 4.00 # | 1.00 ¶ | 6.67 # | 0.67 ¶ | 10.28 ¶ | 9.98 ¶ | 8.64 ¶ | 9.52 ¶ | 8.11 ¶ | 8.13 ¶ | 9.01 ¶ | 9.21 ¶ |
| Ji Syn A | 1.00 ¶ | 1.00 ¶ | 7.00 # | 0.00 ¶ | 0.00 ¶ | 0.00 ¶ | 2.67 # | 0.33 ¶ | 11.27 ¶ | 11.39 ¶ | 9.62 ¶ | 11.61 # | 8.19 ¶ | - | 9.69 ¶ | - |
| Liaoninglv Syn | 7.00 # | 7.00 # | 4.00 # | 8.00 # | 0.00 ¶ | 0.00 ¶ | 3.67 # | 5.00 # | 9.59 ¶ | 10.01 ¶ | 8.82 ¶ | 11.43 # | 7.08 ¶ | - | 8.50 ¶ | - |
| SD | 3.13 ± 4.92 | 1.47 ± 3.06 | 5.86 ± 7.74 | 2.21 ± 5.14 | 5.36 ± 7.75 | 2.20 ± 5.46 | 5.09 ± 2.92 | 2.40 ± 1.56 | 9.36 ± 1.69 | 9.43 ± 1.54 | 8.82 ± 2.15 | 7.53 ± 1.50 | 8.22 ± 1.97 | 7.68 ± 1.51 | 8.71 ± 0.50 | 9.54 ± 1.81 |
| Xianyu 335 | 1.41 | 1.53 | 1.63 | 1.52 | 11.49 | 10.61 | 8.72 | 10.27 | ||||||||
References
- Kandil, E.E.; Abdelsalam, N.R.; Mansour, M.A.; Ali, H.M.; Siddiqui, M.H. Potentials of organic manure and potassium forms on maize (Zea mays L.) growth and production. Sci. Rep. 2020, 10, 8752. [Google Scholar] [CrossRef]
- National Bureau of Statistics of China (NSBC). China Statistics Yearbook; China Statistics Press: Beijing, China, 2021. [Google Scholar]
- Gong, F.; Wu, X.; Zhang, H.; Chen, Y.; Wang, W. Making better maize plants for sustainable grain production in a changing climate. Front. Plant Sci. 2015, 6, 835. [Google Scholar] [CrossRef] [PubMed]
- Yong, H.J.; Tang, J.; Zhao, X.Z.; Zhang, F.Y.; Yang, Z.Y.; Li, Y.; Li, M.S.; Zhang, D.G.; Hao, Z.F.; Weng, J.F.; et al. Effect of five modified mass selection cycles on combining ability in two Chinese maize populations. Euphytica 2020, 216, 57. [Google Scholar] [CrossRef]
- Yong, H.J.; Zhang, X.C.; Zhang, D.G.; Wang, J.J.; Zhang, H.X.; Li, M.S.; Liu, W.G.; Weng, J.F.; Hao, Z.F.; Ci, X.K.; et al. Breeding potential of U.S. maize germplasm for utilization in Chinese temperate conditions. Euphytica 2013, 192, 435–451. [Google Scholar] [CrossRef]
- Ndoro, O.; Magorokosho, C.; Setimela, P.S.; Kamutando, C.N.; Labuschagne, M.T. Identification of exotic temperate maize inbreds for use in tropical breeding programs. Euphytica 2022, 218, 164. [Google Scholar] [CrossRef]
- Ke, F.; Ma, X. Responses of maize hybrids with contrasting maturity to planting date in Northeast China. Sci. Rep. 2021, 11, 15776. [Google Scholar] [CrossRef]
- Dong, M.; Zhao, J.; Li, E.; Liu, Z.; Guo, S.; Zhang, Z.; Cui, W.; Yang, X. Effects of changing climate extremes on maize grain yield in Northeast China. Agronomy 2023, 13, 1050. [Google Scholar] [CrossRef]
- Glover, M.A.; Willmot, D.B.; Darrah, L.L.; Hibbard, B.E.; Zhu, X. Diallel analyses of agronomic traits using Chinese and U.S. maize germplasm. Crop Sci. 2005, 45, 1096–1102. [Google Scholar] [CrossRef]
- Goodman, M.M. Broadening the U.S. maize germplasm base. Maydica 2005, 50, 203–214. [Google Scholar]
- Cupertino-Rodrigues, M.; Dhliwayo, T.; Trachsel, S.; Guo, R.; San, V.F. Evaluation of U.S. inbred lines with expired plant variety protection for mid-altitude tropical maize breeding. Euphytica 2020, 216, 44. [Google Scholar] [CrossRef]
- Rogers, A.R.; Bian, Y.; Krakowsky, M.; Peters, D.; Turnbull, C.; Nelson, P.; Holland, J.B. Genomic prediction for the germplasm enhancement of maize project. Plant Genome 2022, 15, e20267. [Google Scholar] [CrossRef] [PubMed]
- Yong, H.J.; Jin, Z.; Gao, L.; Zhang, L.; Liu, X.; Zhang, F.; Zhang, X.; Zhang, D.; Li, M.S.; Weng, J.F.; et al. Breeding potential of maize germplasm populations to improve yield and predominant heterotic pattern in Northeast China. Euphytica 2017, 213, 219. [Google Scholar] [CrossRef]
- Fan, X.M.; Zhang, Y.D.; Jeffers, D.P.; Bi, Y.Q.; Kang, M.S.; Yin, X.F. Combining ability of yellow lines derived from CIMMYT populations for use in subtropical and tropical midaltitude maize production environments. Crop Sci. 2018, 58, 169–179. [Google Scholar] [CrossRef]
- Azizdoost, H.; Shiri, M.; Dezhsetan, S. The performance of temperate maize testers for screening tropical and subtropical germplasm. Cereal Res. 2024, 13, 367–384. [Google Scholar] [CrossRef]
- Yong, H.J.; Zhang, F.Y.; Tang, J.; Yang, Z.Y.; Zhao, X.Z.; Li, M.S.; Zhang, D.G.; Hao, Z.F.; Weng, J.F.; Li, X.H. Breeding potential of inbred lines derived from five maize (Zea mays L.) populations. Euphytica 2019, 215, 1. [Google Scholar] [CrossRef]
- Smith, J.S.; Trevisan, W.; McCunn, A.; Huffman, W.E. Global dependence on Corn Belt Dent maize germplasm: Challenges and opportunities. Crop Sci. 2022, 62, 2039–2066. [Google Scholar] [CrossRef]
- Hallauer, A.R.; Carena, M.J. Maize breeding. In Handbook of Plant Breeding: Cereals; Carena, M.J., Ed.; Springer: New York, NY, USA, 2009; pp. 3–98. [Google Scholar]
- Rizzo, G.; Monzon, J.P.; Tenorio, F.A.; Réka, H.; Cassman, K.G.; Grassini, P. Climate and agronomy, not genetics, underpin recent maize yield gains in favorable environments. Proc. Natl. Acad. Sci. USA 2022, 119, e2113629119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.H.; Bonjean, A.P.A. Maize breeding and production in China. In Cereals in China; He, Z., Bonjean, A.P.A., Eds.; CIMMYT: El Batan, Mexico, 2010; pp. 35–49. [Google Scholar]
- Zhang, R.Y.; Xu, G.; Li, J.S.; Yan, J.B.; Li, H.H.; Yang, X.H. Patterns of genomic variation in Chinese maize inbred lines and implications for genetic improvement. Theor. Appl. Genet. 2018, 131, 1207–1221. [Google Scholar] [CrossRef] [PubMed]
- Alhaj, H.Y.; Shaghaleh, H.; Zhang, K.; Okla, M.K.; Alaraidh, I.A.; AbdElgawad, H.; Sheteiwy, M.S. Calcium lignosulfonate-induced modification of soil chemical properties improves physiological traits and grain quality of maize (Zea mays) under salinity stress. Front. Plant Sci. 2024, 15, 1397552. [Google Scholar] [CrossRef]
- Baloch, N.; Liu, W.M.; Hou, P.; Ming, B.; Xie, R.Z.; Wang, K.R.; Liu, Y.E.; Li, S.K. Effect of latitude on maize kernel weight and grain yield across China. Agron. J. 2020, 113, 1172–1182. [Google Scholar] [CrossRef]
- Yong, H.J.; Zhang, D.G.; Wang, J.J.; Li, M.S.; Liu, W.G.; Zhang, X.C.; Zhao, H.Y.; Weng, J.F.; Hao, Z.F.; Bai, L.; et al. Broadening the genetic base of Chinese maize heterotic pools with exotic germplasm. Crop Sci. 2013, 53, 1907–1916. [Google Scholar] [CrossRef]
- Prasanna, B.M.; Burgueño, J.; Beyene, Y.; Makumbi, D.; Asea, G.; Woyengo, V.; Tarekegne, A.; Magorokosho, C.; Wegary, D.; Ndhlela, T.; et al. Genetic trends in CIMMYT’s tropical maize breeding pipelines. Sci. Rep. 2022, 12, 20110. [Google Scholar] [CrossRef]
- Hallauer, A.R.; Carena, M.J. Adaptation of tropical maize germplasm to temperate environments. Euphytica 2014, 196, 1–11. [Google Scholar] [CrossRef]
- Olmos, S.E.; Lia, V.V.; Eyhérabide, G.H. Genetic diversity and linkage disequilibrium in the Argentine public maize inbred line collection. Plant Genet. Resour. 2017, 15, 515–526. [Google Scholar] [CrossRef]
- Melani, M.D.; Carena, M.J. Alternative maize heterotic patterns for the Northern Corn Belt. Crop Sci. 2005, 45, 2139–2677. [Google Scholar] [CrossRef]
- Hallauer, A.R. Registration of BS27 maize germplasm. Crop Sci. 1992, 32, 1512–1513. [Google Scholar] [CrossRef]
- Sen, H.; Kumar, A.; Janeja, H.S. Biofortification of major crops through conventional and modern biotechnological approaches to fight hidden hunger: An overview. J. Adv. Biol. Biotechnol. 2024, 27, 96–113. [Google Scholar] [CrossRef]
- Jumbo, M.B.; Carena, M.J. Combining ability, maternal, and reciprocal effects of elite early-maturing maize population hybrids. Euphytica 2008, 162, 325–333. [Google Scholar] [CrossRef]
- Edwards, J. Changes in plant morphology in response to recurrent selection in the Iowa stiff stalk synthetic maize population. Crop Sci. 2011, 51, 2352–2361. [Google Scholar] [CrossRef]
- Yong, H.J.; Wang, L.M.; Li, M.S.; Zhang, D.G.; Pan, G.T.; Zhang, S.H.; Li, X.H.; Rong, T.Z. Breeding potential analysis of eight populations in the application of maize breeding in northeast China. J. Nucl. Agric. Sci. 2014, 28, 2139–2147, (With Chinese Abstract). [Google Scholar]
- Yang, K.; Tan, J.; Zhang, Q.; Bai, T.; Zhou, S.; Hao, J.; Yu, X.; Zang, Z.; Zhang, D. Research on the genetic improvement effects of lodging resistance-related traits in maize core germplasm. Agronomy 2025, 15, 17. [Google Scholar] [CrossRef]
- China Meteorological Administration (CMA). Ground Meteorological Data in China. Available online: http://data.cma.cn/ (accessed on 28 August 2022).
- Butler, D.G.; Cullis, B.R.; Gilmour, A.R.; Gogel, B.J.; Thompson, R. ASReml-R Reference Manual, version 4; VSN International Ltd: Hemel Hempstead, UK, 2017; Available online: https://mmade.org/wp-content/uploads/2021/08/asremlRMfinal.pdf (accessed on 18 August 2021).
- Șimon, A.; Moraru, P.I.; Ceclan, A.; Russu, F.; Chețan, F.; Bărdaș, M.; Popa, A.; Rusu, T.; Pop, A.I.; Bogdan, I. The impact of climatic factors on the development stages of maize crop in the transylvanian plain. Agronomy 2023, 13, 1612. [Google Scholar] [CrossRef]
- Rasmussen, C.C.; Hallauer, A.R. Evaluation of heterotic patterns of Iowa stiff stalk synthetic and non-stiff stalk synthetic maize populations. Maydica 2006, 51, 177–186. [Google Scholar]
- Ku, L.X.; Meng, Q.L.; Hou, B.J.; Li, J.F.; Liu, H.J.; Chen, Y.H. Effificiency of recurrent selection for combining ability of yield trait in maize population Yu Syn5. Acta Agron. Sin. 2012, 38, 215–222, (With Chinese Abstract). [Google Scholar] [CrossRef]
- Koimbori, J.K.; Wang, S.; Pan, J.; Guo, L.; Li, K. Yield response of spring maize under future climate and the effects of adaptation measures in Northeast China. Plants 2022, 11, 1634. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, A.J.; Scott, M.P.; Lamkey, K.R. Genetic variation and breeding potential of phytate and inorganic phosphorus in a maize population. Crop Sci. 2008, 48, 79–84. [Google Scholar] [CrossRef]
- Scott, M.P.; Darrigues, A.; Stahly, T.S.; Lamkey, K.R. Recurrent selection to alter grain methionine concentration and improve nutritional value of maize. Crop Sci. 2008, 48, 1705–1713. [Google Scholar] [CrossRef]

| Population | Ecosystem Type | Germplasm Description | Literature |
|---|---|---|---|
| Tuxpeño | Tropical | Derived from CIMMYT Tuxpeño Planta Baja C11. | [25] |
| Suwan 1 | Tropical | Derived from 36 diverse germplasm sources and two sources with downy mildew resistance and high yield, Philippine DMR 1 and 5. | [25] |
| Stay green-yellow (SG-Y) | Tropical | Derived from 32 original and recycled lines with fairly good resistance to foliar disease. | [25] |
| PooL 26 | Tropical | Includes germplasm from Mexico, Colombia, the Caribbean, and Central America, and the U.S. Corn Belt. | [25] |
| BS10(FR)C10 | Temperate | BS10 was developed by intercrossing 10 inbred lines with prolific ear expression and after 10 cycles of reciprocal full-sib recurrent selection with BS11 as a tester. | [26] |
| BS13(S)C7 | Temperate | An improved breeding population that was developed from Iowa Stiff Stalk Synthetic (BSSS) after 14 cycles of recurrent selection for increased yield and seven cycles for combining ability for yield. | [27] |
| BS20(S)C2 | Temperate | An Iowa Late Rootworm Synthetic was developed by combining 12 inbred lines and after two cycles of recurrent selection and the evaluation of S1 lines in replicated experiments. | [28] |
| BS27 | Temperate | An adapted population of Antigua Composite adapted to temperate conditions initiated in 1977 and after six cycles of mass selection for earlier flowering. | [29] |
| BS31 | Temperate | Derived from a synthetic originally called FS8A(T)C4 developed by mass selection for earliness in a synthetic containing tropical germplasm. | [30] |
| BSBB(SRCB)C4 | Temperate | An Iowa Synthetic BSBB, developed by recombining 44 North Central Corn Belt inbred lines and after four cycles of mass selection. | [31] |
| BSSS(R)C10 | Temperate | BSSS is a synthetic variety derived from 16 inbred lines and five varieties, after seven cycles of reciprocal recurrent selection. | [32] |
| Comp13 | Temperate | Developed from Chinese populations Cysn5 and Cysn6, and CIMMYT population Suwan1 by the Chinese Academy of Agricultural Science. | [33] |
| Comp14 | Temperate | Developed from Chinese populations Cysn7 and Yu Syn5 by the Chinese Academy of Agricultural Sciences. | [33] |
| Ji Syn A | Temperate | Derived from 20 inbred lines with high combining ability by the Jilin Academy of Agricultural Sciences in China were selected from 93 inbred lines. | [33] |
| Liaoninglv Syn | Temperate | Derived from the Lvda Red Cob inbred line by the Liaoning Academy of Agricultural Sciences in China. | [33] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Yang, Z.; Shao, Y.; Jin, Z.; Gao, L.; Yu, Y.; Zhang, F.; Zhang, Y.; Nan, Y.; Li, M.; et al. Adaptation of Diverse Maize Germplasm to Spring Season Conditions in Northeast China. Agronomy 2025, 15, 170. https://doi.org/10.3390/agronomy15010170
Li Y, Yang Z, Shao Y, Jin Z, Gao L, Yu Y, Zhang F, Zhang Y, Nan Y, Li M, et al. Adaptation of Diverse Maize Germplasm to Spring Season Conditions in Northeast China. Agronomy. 2025; 15(1):170. https://doi.org/10.3390/agronomy15010170
Chicago/Turabian StyleLi, Yi, Zhiyuan Yang, Yong Shao, Zhenguo Jin, Li Gao, Yang Yu, Fengyi Zhang, Yuxing Zhang, Yuantao Nan, Mingshun Li, and et al. 2025. "Adaptation of Diverse Maize Germplasm to Spring Season Conditions in Northeast China" Agronomy 15, no. 1: 170. https://doi.org/10.3390/agronomy15010170
APA StyleLi, Y., Yang, Z., Shao, Y., Jin, Z., Gao, L., Yu, Y., Zhang, F., Zhang, Y., Nan, Y., Li, M., Zhang, D., Hao, Z., Weng, J., Li, X., & Yong, H. (2025). Adaptation of Diverse Maize Germplasm to Spring Season Conditions in Northeast China. Agronomy, 15(1), 170. https://doi.org/10.3390/agronomy15010170