
Exploring the Relationship between Nitrogen Use Efficiency and Protein Concentrations in Potato Genotypes

Abstract
1. Introduction
2. Materials and Methods
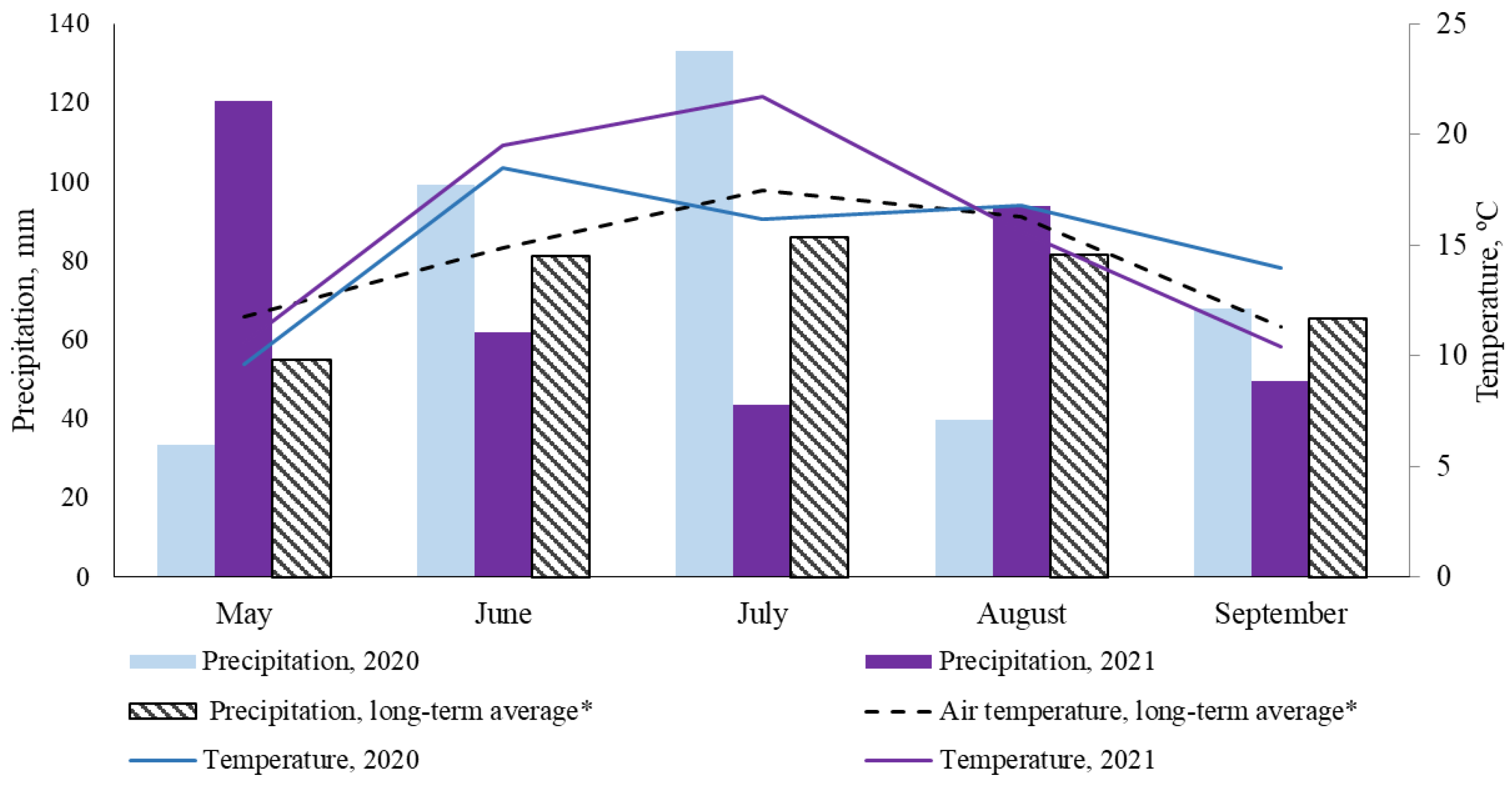
2.1. Growth Conditions
2.2. Plant Material Used in the Trials
2.3. Protein Analysis
2.4. NUE Calculation
2.5. Statistical Analysis
3. Results and Discussion
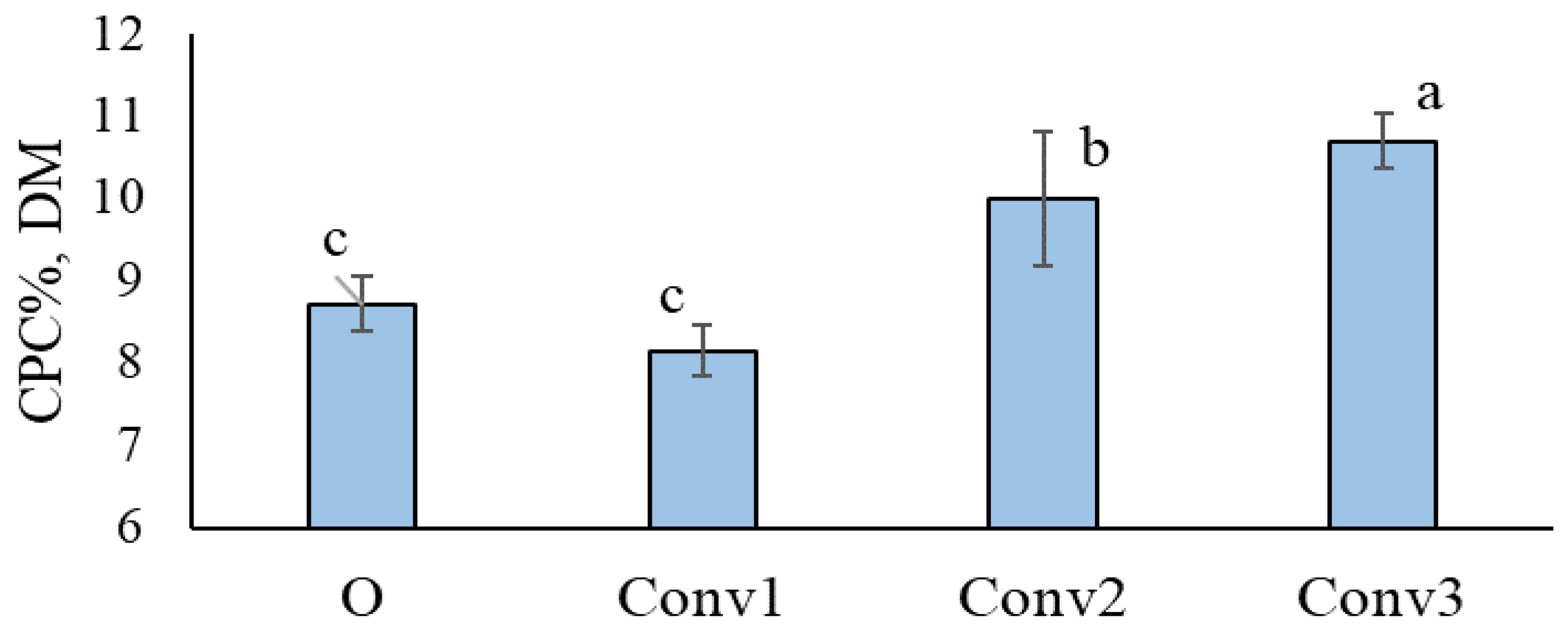
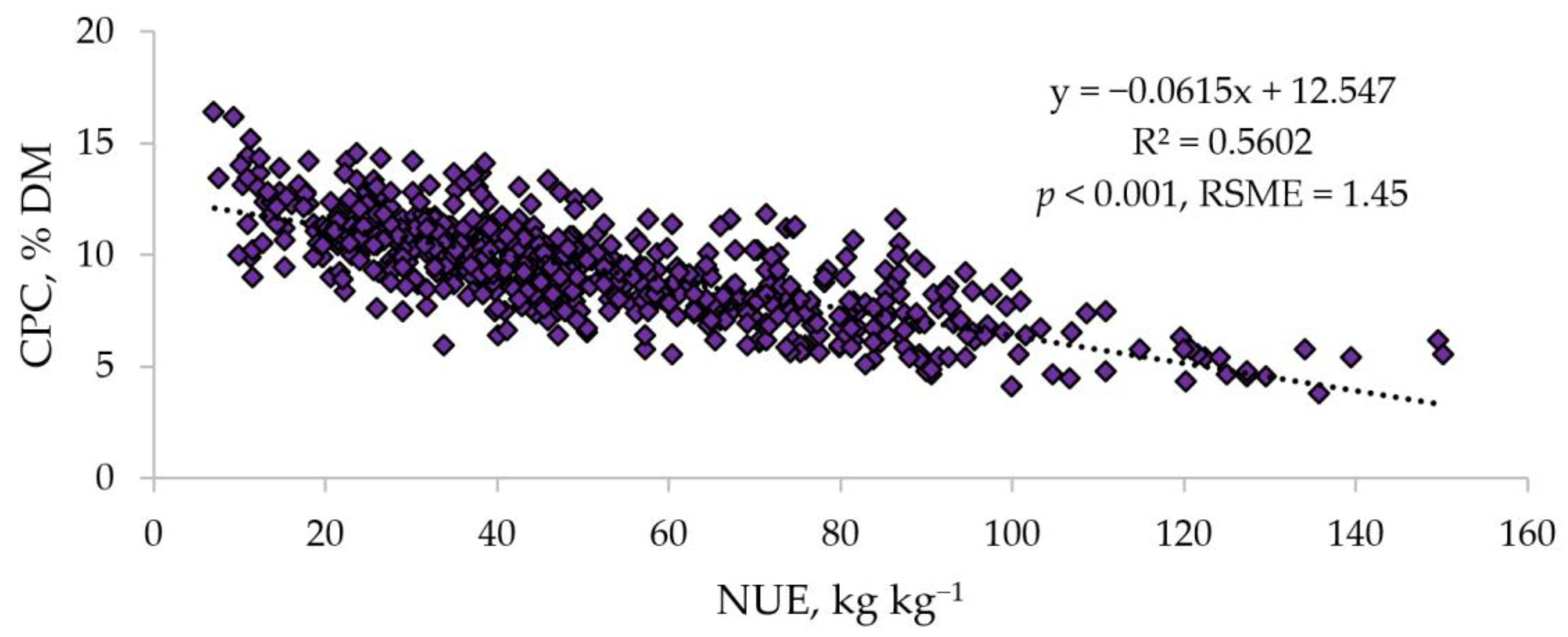
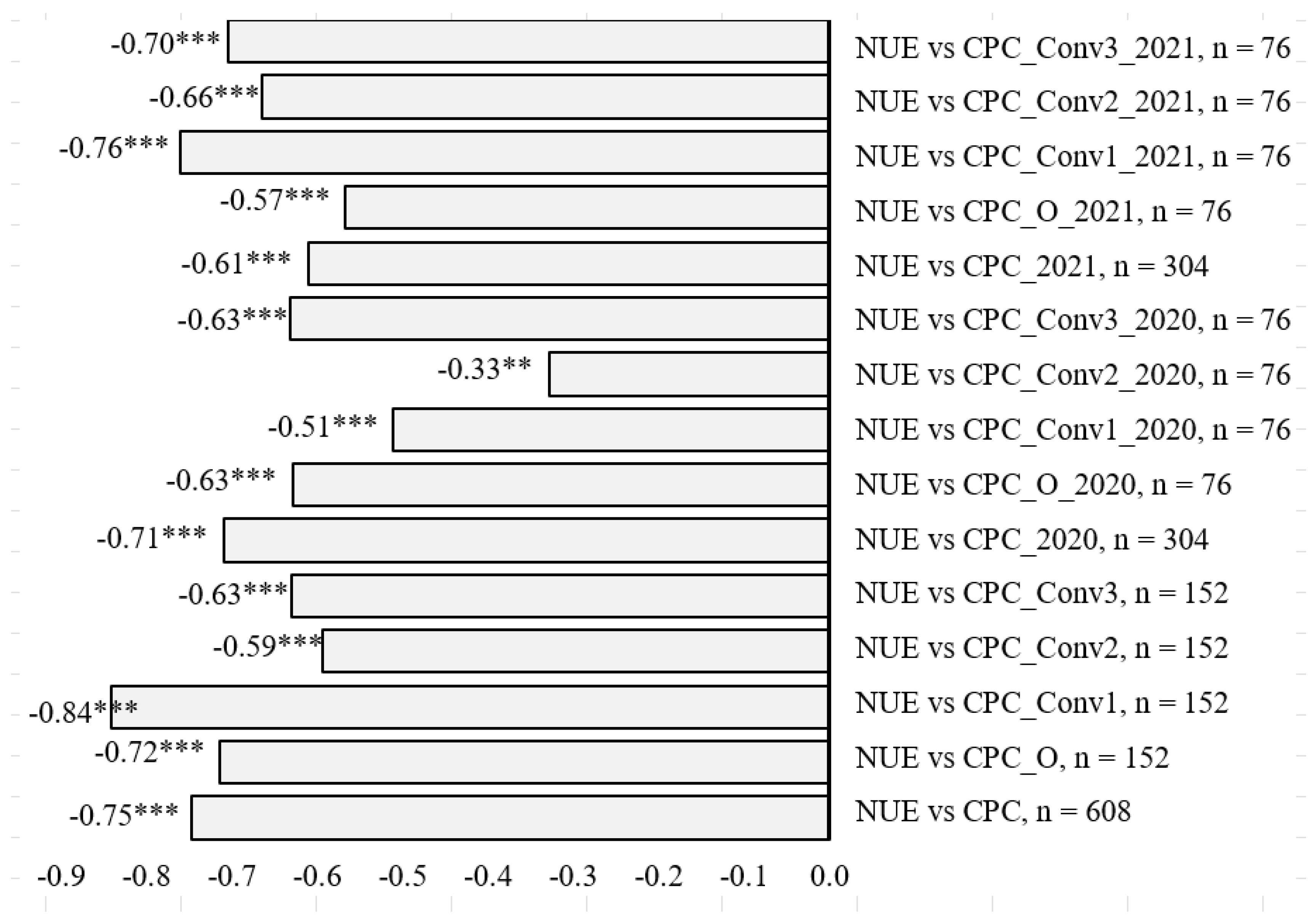
3.1. Crude Protein Concentration (CPC) in Tubers and Its Relationship with NUE
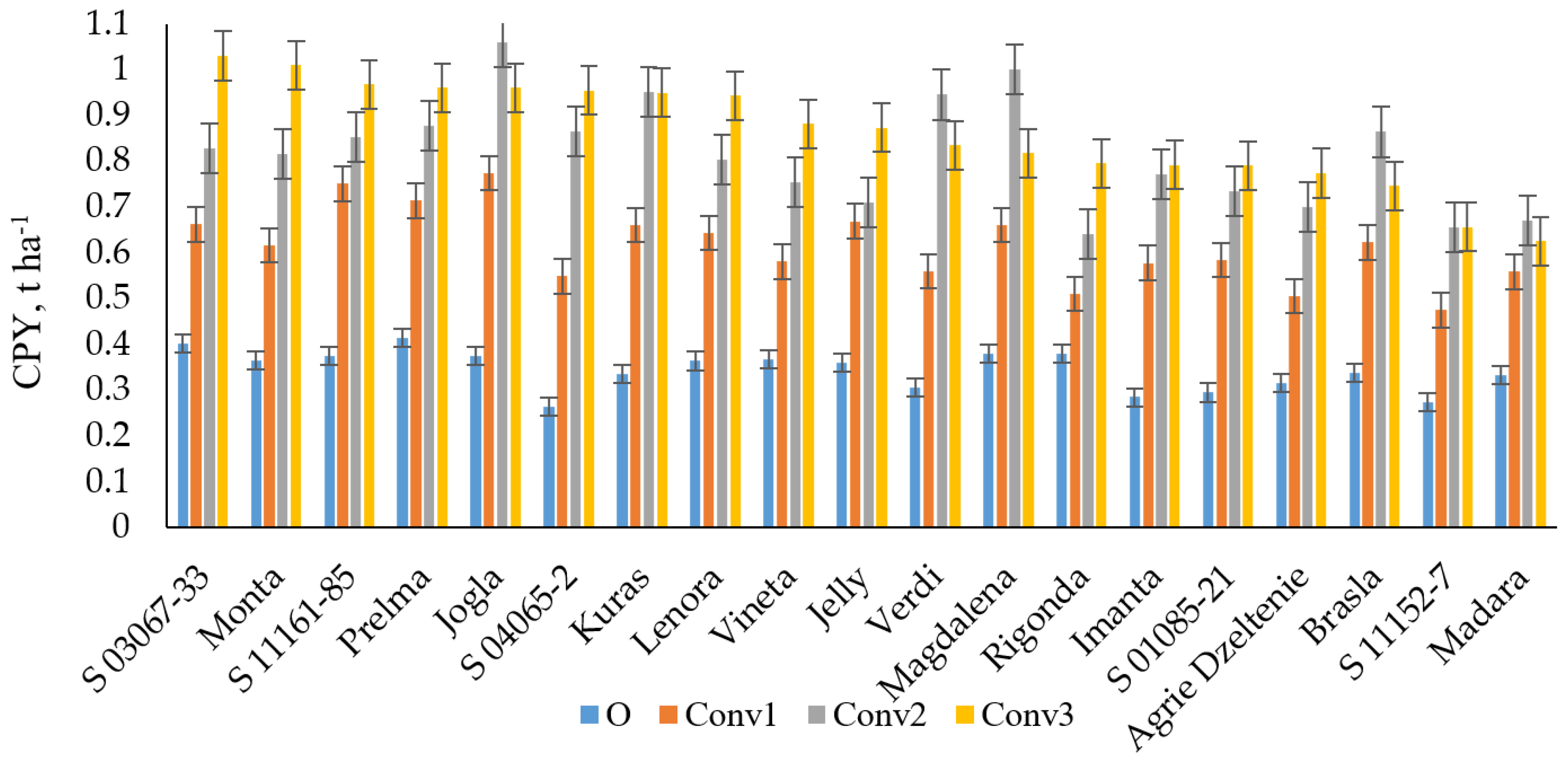
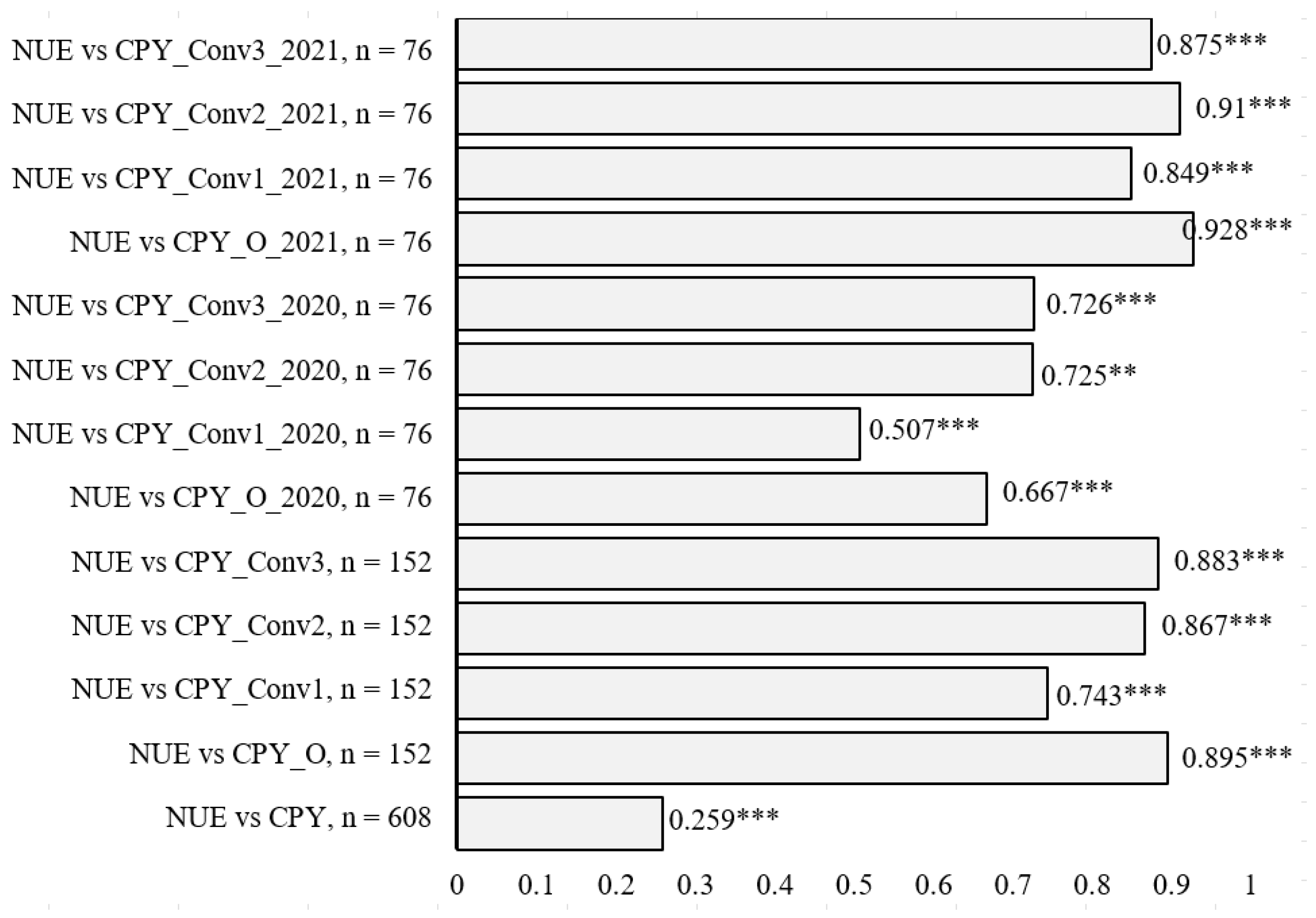
3.2. Crude Protein Yield (CPY) and Its Relationship with NUE
3.3. Patatin Relative Abundance (PRA) in Pure Protein and Its Relationship to NUE
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Camire, M.E.; Kubow, S.; Donnelly, D.J. Potatoes and Human Health. Crit. Rev. Food Sci. Nutr. 2009, 49, 823–840. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Qayum, A.; Xiuxiu, Z.; Liu, L.; Hussain, K.; Yue, P.; Yue, S.; Koko, M.Y.F.; Hussain, A.; Li, X. Potato Protein: An Emerging Source of High Quality and Allergy Free Protein, and Its Possible Future Based Products. Food Res. Int. 2021, 148, 110583. [Google Scholar] [CrossRef] [PubMed]
- Haverkort, A.J. Potato Handbook: Crop of the Future; Aardappelwereld BV: The Hague, The Netherlands, 2018; ISBN 9082897407. [Google Scholar]
- Govindasamy, P.; Muthusamy, S.K.; Bagavathiannan, M.; Mowrer, J.; Jagannadham, P.T.K.; Maity, A.; Halli, H.M.; Sujayananad, G.K.; Vadivel, R.; Das, T.K.; et al. Nitrogen Use Efficiency—A Key to Enhance Crop Productivity under a Changing Climate. Front. Plant Sci. 2023, 14, 1121073. [Google Scholar] [CrossRef] [PubMed]
- Hirel, B.; Tétu, T.; Lea, P.J.; Dubois, F. Improving Nitrogen Use Efficiency in Crops for Sustainable Agriculture. Sustainability 2011, 3, 1452–1485. [Google Scholar] [CrossRef]
- Lammerts van Bueren, E.T.; Struik, P.C. Diverse Concepts of Breeding for Nitrogen Use Efficiency. A Review. Agron. Sustain. Dev. 2017, 37, 50. [Google Scholar] [CrossRef]
- Getahun, B.B.; Kassie, M.M.; Visser, R.G.F.; van der Linden, C.G. Genetic Diversity of Potato Cultivars for Nitrogen Use Efficiency Under Contrasting Nitrogen Regimes. Potato Res. 2020, 63, 267–290. [Google Scholar] [CrossRef]
- Wang, C.; Zang, H.; Liu, J.; Shi, X.; Li, S.; Chen, F.; Chu, Q. Optimum Nitrogen Rate to Maintain Sustainable Potato Production and Improve Nitrogen Use Efficiency at a Regional Scale in China. A Meta-Analysis. Agron. Sustain. Dev. 2020, 40, 37. [Google Scholar] [CrossRef]
- Ospina, C.A.; Lammerts van Bueren, E.T.; Allefs, J.J.H.M.; Engel, B.; van der Putten, P.E.L.; van der Linden, C.G.; Struik, P.C. Diversity of Crop Development Traits and Nitrogen Use Efficiency among Potato Cultivars Grown under Contrasting Nitrogen Regimes. Euphytica 2014, 199, 13–29. [Google Scholar] [CrossRef]
- Zebarth, B.J.; Tai, G.; Tarn, R.; de Jong, H.; Milburn, P.H. Nitrogen Use Efficiency Characteristics of Commercial Potato Cultivars. Can. J. Plant Sci. 2004, 84, 589–598. [Google Scholar] [CrossRef]
- Tiwari, J.K.; Devi, S.; Buckseth, T.; Ali, N.; Singh, R.K.; Zinta, R.; Dua, V.K.; Chakrabarti, S.K. Precision Phenotyping of Contrasting Potato (Solanum tuberosum L.) Varieties in a Novel Aeroponics System for Improving Nitrogen Use Efficiency: In Search of Key Traits and Genes. J. Integr. Agric. 2020, 19, 51–61. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Y.; Zhao, Y.; Zhang, Y.; Zhang, J.; Ma, H.; Han, Y. Erratum: Transcriptome Analysis Reveals Nitrogen Deficiency Induced Alterations in Leaf and Root of Three Cultivars of Potato (Solanum tuberosum L.). PLoS ONE 2020, 15, e0240662, Erratum in PLoS ONE 2021, 16, e0253994. https://doi.org/10.1371/journal.pone.0253994. [Google Scholar] [CrossRef]
- Gálvez, J.H.; Tai, H.H.; Lagüe, M.; Zebarth, B.J.; Strömvik, M.V. The Nitrogen Responsive Transcriptome in Potato (Solanum tuberosum L.) Reveals Significant Gene Regulatory Motifs. Sci. Rep. 2016, 6, 26090. [Google Scholar] [CrossRef]
- Baligar, V.C.; Fageria, N.K.; He, Z.L. Nutrient Use Efficiency in Plants. Commun. Soil Sci. Plant Anal. 2001, 32, 921–950. [Google Scholar] [CrossRef]
- Tiemens-Hulscher, M.; Lammerts van Bueren, E.T.; Struik, P.C. Identifying Nitrogen-Efficient Potato Cultivars for Organic Farming. Euphytica 2014, 199, 137–154. [Google Scholar] [CrossRef]
- Koch, M.; Naumann, M.; Pawelzik, E.; Gransee, A.; Thiel, H. The Importance of Nutrient Management for Potato Production Part I: Plant Nutrition and Yield. Potato Res. 2020, 63, 97–119. [Google Scholar] [CrossRef]
- Stefaniak, T.R.; Fitzcollins, S.; Figueroa, R.; Thompson, A.L.; Schmitz Carley, C.; Shannon, L.M. Genotype and Variable Nitrogen Effects on Tuber Yield and Quality for Red Fresh Market Potatoes in Minnesota. Agronomy 2021, 11, 255. [Google Scholar] [CrossRef]
- Ruza, A.; Skrabule, I.; Vaivode, A. Influence of Nitrogen on Potato Productivity and Nutrient Use Efficiency. Proc. Latv. Acad. Sci. 2013, 67, 247–253. [Google Scholar]
- Zhang, H.; Liu, X.; Song, B.; Nie, B.; Zhang, W.; Zhao, Z. Effect of Excessive Nitrogen on Levels of Amino Acids and Sugars, and Differential Response to Post-Harvest Cold Storage in Potato (Solanum tuberosum L.) Tubers. Plant Physiol. Biochem. 2020, 157, 38–46. [Google Scholar] [CrossRef]
- Barta, J.; Bartova, V. Patatin, the Major Protein of Potato (Solanum tuberosum L.) Tubers, and Its Occurrence as Genotype Effect: Processing versus Table Potatoes. Czech J. Food Sci. 2008, 26, 347–359. [Google Scholar] [CrossRef]
- Bártová, V.; Bárta, J.; Brabcová, A.; Zdráhal, Z.; Horáčková, V. Amino Acid Composition and Nutritional Value of Four Cultivated South American Potato Species. J. Food Compos. Anal. 2015, 40, 78–85. [Google Scholar] [CrossRef]
- Murniece, I.; Karklina, D.; Galoburda, R.; Santare, D.; Skrabule, I.; Costa, H.S. Nutritional Composition of Freshly Harvested and Stored Latvian Potato (Solanum tuberosum L.) Varieties Depending on Traditional Cooking Methods. J. Food Compos. Anal. 2011, 24, 699–710. [Google Scholar] [CrossRef]
- Gorissen, S.H.M.; Crombag, J.J.R.; Senden, J.M.G.; Waterval, W.A.H.; Bierau, J.; Verdijk, L.B.; van Loon, L.J.C. Protein Content and Amino Acid Composition of Commercially Available Plant-Based Protein Isolates. Amino Acids 2018, 50, 1685–1695. [Google Scholar] [CrossRef]
- Waglay, A.; Karboune, S.; Khodadadi, M. Investigation and Optimization of a Novel Enzymatic Approach for the Isolation of Proteins from Potato Pulp. LWT Food Sci. Technol. 2016, 65, 197–205. [Google Scholar] [CrossRef]
- Bernal, J.; Mouzo, D.; López-Pedrouso, M.; Franco, D.; García, L.; Zapata, C. The Major Storage Protein in Potato Tuber Is Mobilized by a Mechanism Dependent on Its Phosphorylation Status. Int. J. Mol. Sci. 2019, 20, 1889. [Google Scholar] [CrossRef]
- Pots, A.M.; Gruppen, H.; Van Diepenbeek, R.; Van Der Lee, J.J.; Van Boekel, M.A.J.S.; Wijngaards, G.; Voragen, A.G.J. The Effect of Storage of Whole Potatoes of Three Cultivars on the Patatin and Protease Inhibitor Content; A Study Using Capillary Electrophoresis and MALDI-TOF Mass Spectrometry. J. Sci. Food Agric. 1999, 79, 1557–1564. [Google Scholar] [CrossRef]
- de Souza Cândido, E.; Pinto, M.F.S.; Pelegrini, P.B.; Lima, T.B.; Silva, O.N.; Pogue, R.; Grossi-de-Sá, M.F.; Franco, O.L. Plant Storage Proteins with Antimicrobial Activity: Novel Insights into Plant Defense Mechanisms. FASEB J. 2011, 25, 3290–3305. [Google Scholar] [CrossRef]
- Pęksa, A.; Miedzianka, J. Potato Industry By-Products as a Source of Protein with Beneficial Nutritional, Functional, Health-Promoting and Antimicrobial Properties. Appl. Sci. 2021, 11, 3497. [Google Scholar] [CrossRef]
- Kowalczewski, P.Ł.; Olejnik, A.; Białas, W.; Rybicka, I.; Zielińska-Dawidziak, M.; Siger, A.; Kubiak, P.; Lewandowicz, G. The Nutritional Value and Biological Activity of Concentrated Protein Fraction of Potato Juice. Nutrients 2019, 11, 1523. [Google Scholar] [CrossRef]
- Liu, Y.-W.; Han, C.-H.; Lee, M.-H.; Hsu, F.-L.; Hou, W.-C. Patatin, the Tuber Storage Protein of Potato (Solanum tuberosum L.), Exhibits Antioxidant Activity in Vitro. J. Agric. Food Chem. 2003, 51, 4389–4393. [Google Scholar] [CrossRef]
- Fu, Y.; Liu, W.N.; Soladoye, O.P. Towards Potato Protein Utilisation: Insights into Separation, Functionality and Bioactivity of Patatin. Int. J. Food Sci. Technol. 2020, 55, 2314–2322. [Google Scholar] [CrossRef]
- Bartova, V.; Barta, J.; Divis, J.; Svajiner, J.; Peterka, J. Crude Protein Content in Tubers of Starch Processing Potato Cultivars in Dependence on Different Agro-Ecological Conditions Vliv Agroekologických Podmínek Na Obsah Hrubého Proteinu V Hlízách. Cent. Eur. Agric. 2009, 10, 57–66. [Google Scholar]
- Mitrus, J.; Stankiewicz, C.; Steć, E.; Kamecki, M.; Starczewski, J. The Influence of Selected Cultivation on the Content of Total Protein and Amino Acids in the Potato Tubers. Plant Soil Environ. 2003, 49, 131–134. [Google Scholar] [CrossRef]
- Öztürk, E.; Kavurmaci, Z.; Kara, K.; Polat, T. The Effects of Different Nitrogen and Phosphorus Rates on Some Quality Traits of Potato. Potato Res. 2010, 53, 309–312. [Google Scholar] [CrossRef]
- Assunção, N.S.; Fernandes, A.M.; Soratto, R.P.; Mota, L.H.S.O.; Ribeiro, N.P.; Leonel, M. Tuber Yield and Quality of Two Potato Cultivars in Response to Nitrogen Fertilizer Management. Potato Res. 2021, 64, 147–166. [Google Scholar] [CrossRef]
- Zarzecka, K.; Gugala, M.; Mystkowska, I.; Sikorska, A. Total and True Protein Content in Potato Tubers Depending on Herbicides and Biostimulants. Agronomy 2020, 10, 1106. [Google Scholar] [CrossRef]
- Skrabule, I.; Rābante-hāne, L.; Dimante, I.; Taškova, I. The Effect of Nitrogen Use Efficiency on Significant Traits of Potato Starch Production. Zemdirb. Agric. 2023, 110, 329–338. [Google Scholar] [CrossRef]
- Berga, D.; Sterna, V.; Sokolova, E.; Taskova, I.; Seile, S.; Dimante, I.; Skrabule, I. Evaluation of Patatin Content in Proteins of Potato Genotypes Grown in Latvia. Rural Sustain. Res. 2021, 46, 125–132. [Google Scholar] [CrossRef]
- Hawkesford, M.J.; Griffiths, S. Exploiting Genetic Variation in Nitrogen Use Efficiency for Cereal Crop Improvement. Curr. Opin. Plant Biol. 2019, 49, 35–42. [Google Scholar] [CrossRef]
- Jamovi The Jamovi Project 2021. Available online: https://www.jamovi.org/ (accessed on 14 June 2024).
- Leszczyński, W.; Lisińska, G. Influence of Nitrogen Fertilization on Chemical Composition of Potato Tubers. Food Chem. 1988, 28, 45–52. [Google Scholar] [CrossRef]
- Bártová, V.; Diviš, J.; Bárta, J.; Brabcová, A.; Švajnerová, M. Variation of Nitrogenous Components in Potato (Solanum tuberosum L.) Tubers Produced under Organic and Conventional Crop Management. Eur. J. Agron. 2013, 49, 20–31. [Google Scholar] [CrossRef]
- Tong, C.; Ma, Z.; Chen, H.; Gao, H. Toward an Understanding of Potato Starch Structure, Function, Biosynthesis, and Applications. Food Front. 2023, 4, 980–1000. [Google Scholar] [CrossRef]
- Bernhard, T.; Truberg, B.; Friedt, W.; Snowdon, R.; Wittkop, B. Development of Near-Infrared Reflection Spectroscopy Calibrations for Crude Protein and Dry Matter Content in Fresh and Dried Potato Tuber Samples. Potato Res. 2016, 59, 149–165. [Google Scholar] [CrossRef]
- Leonel, M.; do Carmo, E.L.; Fernandes, A.M.; Soratto, R.P.; Ebúrneo, J.A.M.; Garcia, É.L.; dos Santos, T.P.R. Chemical Composition of Potato Tubers: The Effect of Cultivars and Growth Conditions. J. Food Sci. Technol. 2017, 54, 2372–2378. [Google Scholar] [CrossRef]
- Milroy, S.P.; Wang, P.; Sadras, V.O. Defining Upper Limits of Nitrogen Uptake and Nitrogen Use Efficiency of Potato in Response to Crop N Supply. Field Crops Res. 2019, 239, 38–46. [Google Scholar] [CrossRef]
- Bárta, J.; Bártová, V.; Zdráhal, Z.; Šedo, O. Cultivar Variability of Patatin Biochemical Characteristics: Table versus Processing Potatoes (Solanum tuberosum L.). J. Agric. Food Chem. 2012, 60, 4369–4378. [Google Scholar] [CrossRef]
- Lehesranta, S.J.; Koistinen, K.M.; Massat, N.; Davies, H.V.; Shepherd, L.V.T.; McNicol, J.W.; Cakmak, I.; Cooper, J.; Lück, L.; Kärenlampi, S.O. Effects of Agricultural Production Systems and Their Components on Protein Profiles of Potato Tubers. Proteomics 2007, 7, 597–604. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicators | Organic Field | Integrated Field | ||
|---|---|---|---|---|
| 2020 | 2021 | 2020 | 2021 | |
| pHKCl | 5.7 | 5.6 | 5.5 | 5.0 |
| Organic matter, g kg−1 | 23 | 27 | 23 | 21 |
| K2O, mg kg−1 | 56 | 132 | 137 | 190 |
| P2O5, mg kg−1 | 101 | 164 | 179 | 175 |
| Estimated plant-available N in the soil kg ha−1 (before fertilization) | 71 | 83 | 69 | 58 |
| Cultivar | PRA, % | ||
|---|---|---|---|
| 2020 | 2021 | Average | |
| Verdi | 34.2 | 36.8 | 35.5 a |
| Brasla | 22.4 | 27.3 | 24.9 b |
| Imanta | 23.2 | 26.6 | 24.9 b |
| Lenora | 21.2 | 25.8 | 23.5 bc |
| S 11152-7 | 23.9 | 22.7 | 23.3 bc |
| Rigonda | 23.0 | 21.5 | 22.3 bcd |
| S 11161-85 | 20.0 | 24.5 | 22.2 bcd |
| Monta | 24.0 | 19.2 | 21.6 bcde |
| Jogla | 21.1 | 18.6 | 19.8 bcde |
| Madara | 21.2 | 17.7 | 19.5 bcdef |
| S 04065-2 | 19.5 | 19.3 | 19.4 bcdef |
| Jelly | 13.3 | 18.9 | 16.1 cdefg |
| Agrie Dzeltenie | 17.2 | 14.5 | 15.9 cdefg |
| Kuras | 17.1 | 13.7 | 15.4 cdefg |
| S 01085-21 | 12.2 | 17.9 | 15.1 cdefg |
| S 03067-33 | 12.5 | 14.5 | 13.5 efg |
| Prelma | 7.2 | 14.5 | 10.8 fg |
| Vineta | 8.7 | 11.4 | 10.0 g |
| Magdalena | 6.5 | 10.9 | 8.7 g |
| Average | 18.33 | 19.80 | 19.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dimante, I.; Skrabule, I.; Sokolova, E.; Taskova, I.; Berga, D.; Sterna, V. Exploring the Relationship between Nitrogen Use Efficiency and Protein Concentrations in Potato Genotypes. Agronomy 2024, 14, 1517. https://doi.org/10.3390/agronomy14071517
Dimante I, Skrabule I, Sokolova E, Taskova I, Berga D, Sterna V. Exploring the Relationship between Nitrogen Use Efficiency and Protein Concentrations in Potato Genotypes. Agronomy. 2024; 14(7):1517. https://doi.org/10.3390/agronomy14071517
Chicago/Turabian StyleDimante, Ilze, Ilze Skrabule, Elina Sokolova, Inese Taskova, Dace Berga, and Vita Sterna. 2024. "Exploring the Relationship between Nitrogen Use Efficiency and Protein Concentrations in Potato Genotypes" Agronomy 14, no. 7: 1517. https://doi.org/10.3390/agronomy14071517
APA StyleDimante, I., Skrabule, I., Sokolova, E., Taskova, I., Berga, D., & Sterna, V. (2024). Exploring the Relationship between Nitrogen Use Efficiency and Protein Concentrations in Potato Genotypes. Agronomy, 14(7), 1517. https://doi.org/10.3390/agronomy14071517