
Genomics-Assisted Breeding: A Powerful Breeding Approach for Improving Plant Growth and Stress Resilience

 , ,
, ,  and
and
Abstract
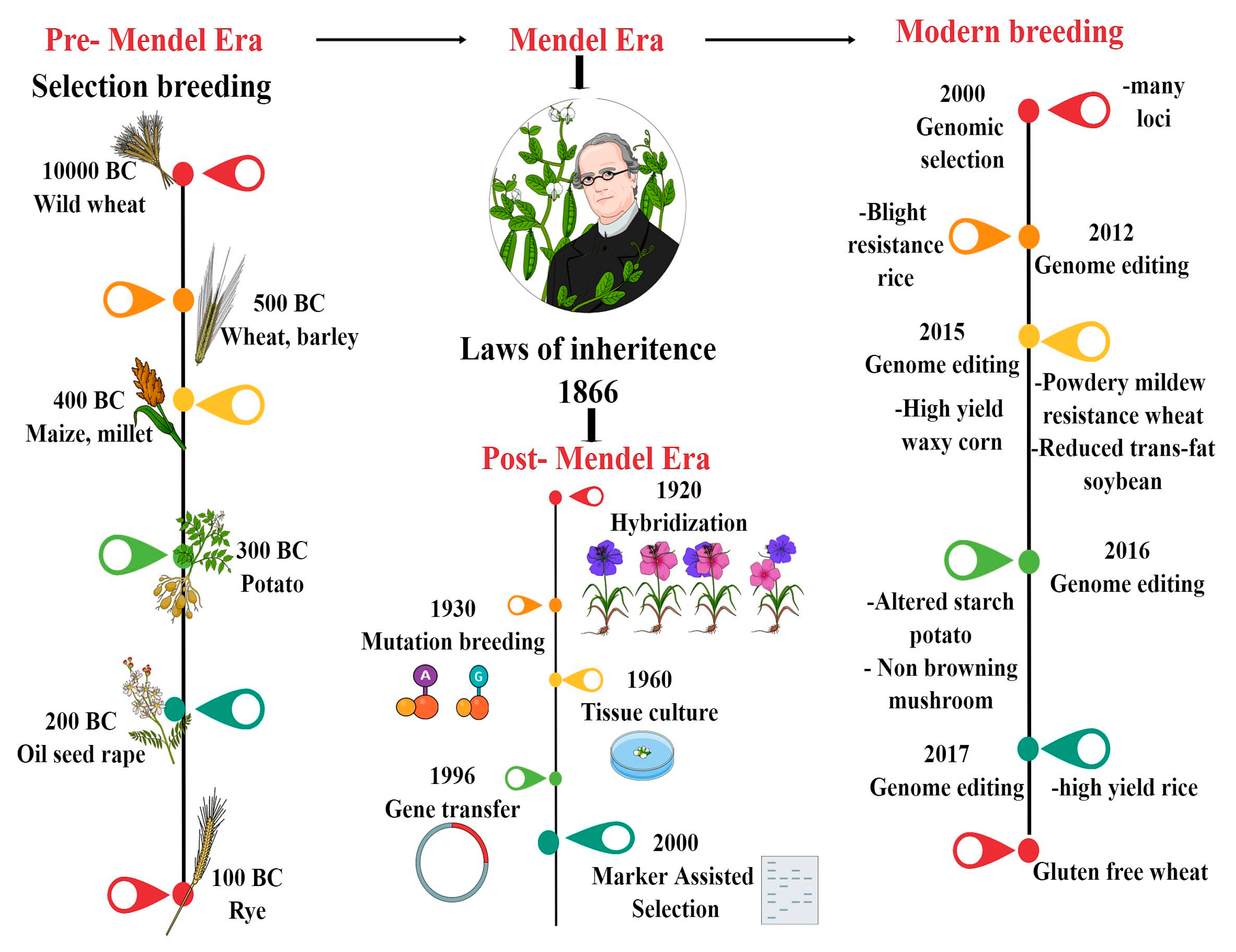
1. Introduction
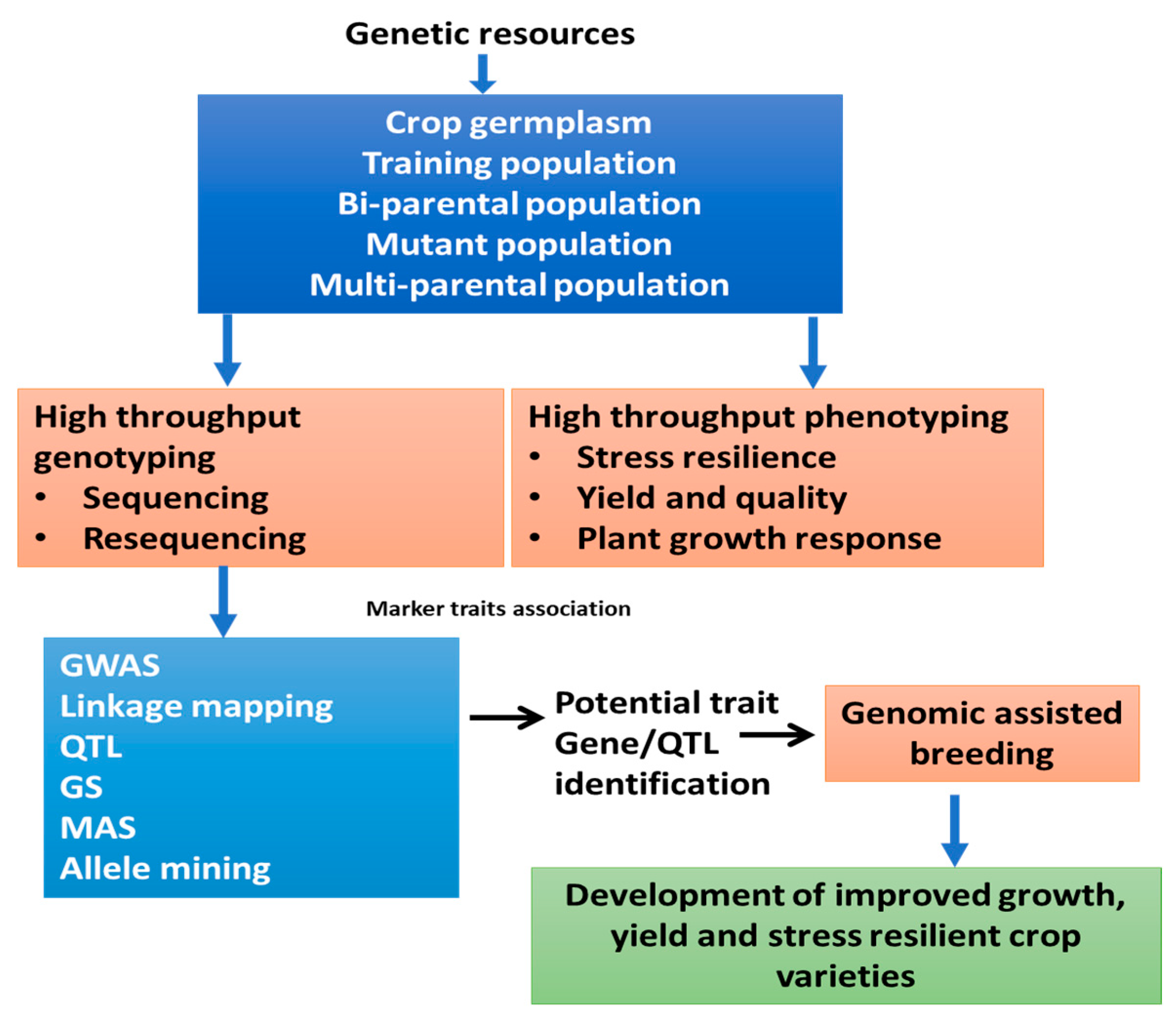
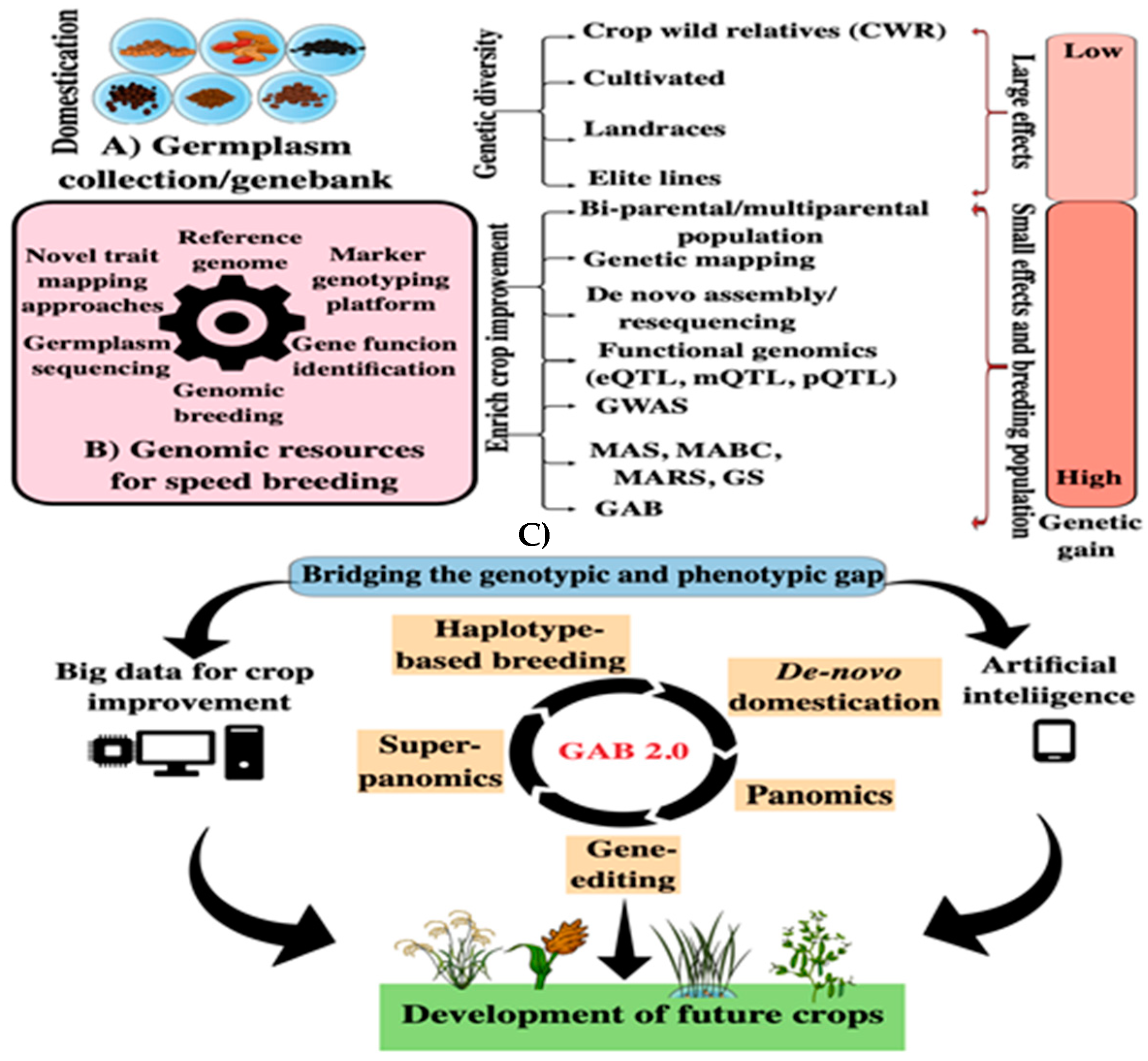
2. How to Use Genomic Information to Maximize Genetic Gain in Plant Breeding Program?
3. GxE in Genomic Predictions
4. GAB for Improving Stress Resilience and Quality Traits in Food Crops
4.1. GAB for Improving Disease Resistance in Crops
4.2. Utilizing GAB for Improving Abiotic Stress Tolerance in Plants
4.3. Application of GAB for Improving Growth and Quality Traits
5. Omics-Assisted Breeding (OAB): Toward More Efficient Food Production
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ali, S.; Tyagi, A.; Rajarammohan, S.; Mir, Z.A.; Bae, H. Revisiting Alternaria-host interactions: New insights on its pathogenesis, defense mechanisms and control strategies. Sci. Hortic. 2023, 322, 112424. [Google Scholar] [CrossRef]
- Gou, C.; Zafar, S.; Hasnain, Z.; Aslam, N.; Iqbal, N.; Abbas, S.; Li, H.; Li, J.; Chen, B.; Ragauskas, A.J.; et al. Machine and Deep Learning: Artificial Intelligence Application in Biotic and Abiotic Stress Management in Plants. Front. Biosci. 2024, 29, 20. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.; Sun, J.; Shabbir, S.; Khattak, W.A.; Ren, G.; Nie, X.; Sonne, C. A review of plants strategies to resist biotic and abiotic environmental stressors. Sci. Total Environ. 2023, 900, 165832. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Reynolds, M.; Xu, Y. Climate change challenges plant breeding. Curr. Opin. Plant Biol. 2022, 70, 102308. [Google Scholar] [CrossRef] [PubMed]
- Benitez-Alfonso, Y.; Soanes, B.K.; Zimba, S.; Sinanaj, B.; German, L.; Sharma, V.; Foyer, C.H. Enhancing climate change resilience in agricultural crops. Curr. Biol. 2023, 33, R1246–R1261. [Google Scholar] [CrossRef]
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2022; FAO: Rome, Italy, 2022; Available online: https://data.unicef.org/resources/sofi-2022/ (accessed on 1 March 2024).
- Krishna, T.P.A.; Veeramuthu, D.; Maharajan, T.; Soosaimanickam, M. The Era of Plant Breeding: Conventional Breeding to Genomics-assisted Breeding for Crop Improvement. Curr. Genom. 2023, 24, 24–35. [Google Scholar] [CrossRef]
- Razzaq, A.; Kaur, P.; Akhter, N.; Wani, S.H.; Saleem, F. Next-generation breeding strategies for climate-ready crops. Front. Plant Sci. 2021, 12, 620420. [Google Scholar] [CrossRef]
- Ahmad, M. Plant breeding advancements with “CRISPR-Cas” genome editing technologies will assist future food security. Front. Plant Sci. 2023, 14, 1133036. [Google Scholar] [CrossRef]
- Li, J.-F.; Norville, J.E.; Aach, J.; McCormack, M.; Zhang, D.; Bush, J.; Church, G.M.; Sheen, J. Multiplex and homologous recombination–mediated genome editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9. Nat. Biotechnol. 2013, 31, 688–691. [Google Scholar] [CrossRef]
- Shan, Q.; Wang, Y.; Li, J.; Zhang, Y.; Chen, K.; Liang, Z.; Zhang, K.; Liu, J.; Xi, J.J.; Qiu, J.L.; et al. Targeted genome modification of crop plants using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 686–688. [Google Scholar] [CrossRef]
- Bohra, A.; Chand Jha, U.; Godwin, I.D.; Kumar Varshney, R. Genomic interventions for sustainable agriculture. Plant Biotechnol. J. 2020, 18, 2388–2405. [Google Scholar] [CrossRef] [PubMed]
- Pazhamala, L.; Saxena, R.K.; Singh, V.K.; Sameerkumar, C.V.; Kumar, V.; Sinha, P.; Patel, K.; Obala, J.; Kaoneka, S.R.; Tongoona, P.; et al. Genomics-assisted breeding for boosting crop improvement in pigeonpea (Cajanus cajan). Front. Plant Sci. 2015, 6, 50. [Google Scholar] [CrossRef] [PubMed]
- Leng, P.F.; Lübberstedt, T.; Xu, M.L. Genomics-assisted breeding–a revolutionary strategy for crop improvement. J. Integr. Agric. 2017, 16, 2674–2685. [Google Scholar] [CrossRef]
- Sinha, P.; Singh, V.K.; Bohra, A.; Kumar, A.; Reif, J.C.; Varshney, R.K. Genomics and breeding innovations for enhancing genetic gain for climate resilience and nutrition traits. Theor. Appl. Genet. 2021, 134, 1829–1843. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.; Messina, C.D.; Podlich, D.; Totir, L.R.; Baumgarten, A.; Hausmann, N.J.; Wright, D.; Graham, G. Predicting the future of plant breeding. Complementing empirical evaluation with genetic prediction. Crop Pasture Sci. 2014, 65, 311–336. [Google Scholar] [CrossRef]
- Varshney, R.K.; Bohra, A.; Yu, J.; Graner, A.; Zhang, Q.; Sorrells, M.E. Designing future crops: Genomics-assisted breeding comes of age. Trends Plant Sci. 2021, 26, 631–649. [Google Scholar] [CrossRef] [PubMed]
- Qutub, M.; Chandran, S.; Rathinavel, K.; Sampathrajan, V.; Rajasekaran, R.; Manickam, S.; Adhimoolam, K.; Muniyandi, S.J.; Natesan, S. Improvement of a Yairipok Chujak maize landrace from North Eastern Himalayan region for β-carotene content through molecular marker-assisted backcross breeding. Genes 2021, 12, 762. [Google Scholar] [CrossRef]
- Sundaram, R.M.; Vishnupriya, M.R.; Biradar, S.K.; Laha, G.S.; Reddy, G.A.; Rani, N.S.; Sarma, N.P.; Sonti, R.V. Marker assisted introgression of bacterial blight resistance in Samba Mahsuri, an elite indica rice variety. Euphytica 2008, 160, 411–422. [Google Scholar] [CrossRef]
- Ratna Madhavi, K.; Rambabu, R.; Abhilash Kumar, V.; Vijay Kumar, S.; Aruna, J.; Ramesh, S.; Sundaram, R.M.; Laha, G.S.; Sheshu Madhav, M.; Ravindra Babu, V.; et al. Marker assisted introgression of blast (Pi-2 and Pi-54) genes in to the genetic background of elite, bacterial blight resistant indica rice variety, Improved Samba Mahsuri. Euphytica 2016, 212, 331–342. [Google Scholar] [CrossRef]
- Yi, M.; Vanavichit, A.; Chai-arree, W.; Toojinda, T. Marker assisted backcross breeding to improve cooking quality traits in Myanmar rice cultivar Manawthukha. Field Crops Res. 2009, 113, 178–186. [Google Scholar] [CrossRef]
- Janila, P.; Pandey, M.K.; Shasidhar, Y.; Variath, M.T.; Sriswathi, M.; Khera, P.; Manohar, S.S.; Nagesh, P.; Vishwakarma, M.K.; Mishra, G.P.; et al. Molecular breeding for introgression of fatty acid desaturase mutant alleles (ahFAD2A and ahFAD2B) enhances oil quality in high and low oil containing peanut genotypes. Plant Sci. 2016, 242, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Yaduru Shasidhar, Y.S.; Variath, M.T.; Vishwakarma, M.K.; Manohar, S.S.; Gangurde, S.S.; Manda Sriswathi, M.S.; Sudini, H.K.; Dobariya, K.L.; Bera, S.K.; Thankappan Radhakrishnan, T.R.; et al. Improvement of three popular Indian groundnut varieties for foliar disease resistance and high oleic acid using SSR markers and SNP array in marker-assisted backcrossing. Plant Biol. J. 2020, 8, 1–15. [Google Scholar] [CrossRef]
- Mitrofanova, O.P.; Khakimova, A.G. New genetic resources in wheat breeding for increased grain protein content. Russ. J. Genet. Appl. Res. 2017, 7, 477–487. [Google Scholar] [CrossRef]
- Shamshad, M.; Sharma, A. The usage of genomic selection strategy in plant breeding. Next Gener. Plant Breed. 2018, 26, 93–108. [Google Scholar]
- Huang, J.; Hu, T.; Yasir, M.; Gao, Y.; Chen, C.; Zhu, R.; Wang, X.; Yuan, H.; Yang, J. Root growth dynamics and yield responses of rice (Oryza sativa L.) under drought—Flood abrupt alternating conditions. Environ. Exp. Bot. 2019, 157, 11–25. [Google Scholar] [CrossRef]
- Shikha, M.; Kanika, A.; Rao, A.R.; Mallikarjuna, M.G.; Gupta, H.S.; Nepolean, T. Genomic selection for drought tolerance using genome-wide SNPs in maize. Front. Plant Sci. 2017, 8, 550. [Google Scholar] [CrossRef]
- Meuwissen, T.H.; Hayes, B.J.; Goddard, M. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef] [PubMed]
- Yugander, A.; Sundaram, R.M.; Singh, K.; Ladhalakshmi, D.; Subba Rao, L.V.; Madhav, M.S.; Badri, J.; Prasad, M.S.; Laha, G.S. Incorporation of the novel bacterial blight resistance gene Xa38 into the genetic background of elite rice variety Improved Samba Mahsuri. PLoS ONE 2018, 13, e0198260. [Google Scholar] [CrossRef] [PubMed]
- Khanna, A.; Sharma, V.; Ellur, R.K.; Shikari, A.B.; Gopala Krishnan, S.; Singh, U.D.; Prakash, G.; Sharma, T.R.; Rathour, R.; Variar, M.; et al. Development and evaluation of near-isogenic lines for major blast resistance gene(s) in Basmati rice. Theor. Appl. Genet. 2015, 128, 1243–1259. [Google Scholar] [CrossRef]
- Ellur, R.K.; Khanna, A.; Yadav, A.; Pathania, S.; Rajashekara, H.; Singh, V.K.; Krishnan, S.G.; Bhowmick, P.K.; Nagarajan, M.; Vinod, K.K.; et al. Improvement of Basmati rice varieties for resistance to blast and bacterial blight diseases using marker assisted backcross breeding. Plant Sci. 2016, 242, 330–341. [Google Scholar] [CrossRef]
- Neeraja, C.N.; Maghirang-Rodriguez, R.; Pamplona, A.; Heuer, S.; Collard, B.C.; Septiningsih, E.M.; Vergara, G.; Sanchez, D.; Xu, K.; Ismail, A.M.; et al. A marker-assisted backcross approach for developing submergence-tolerant rice cultivars. Theor. Appl. Genet. 2017, 115, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Lang, N.T.; Tao, N.V.; Buu, B.C. Marker-assisted backcrossing (MAB) for rice submergence tolerance in Mekong delta. Omonrice 2011, 18, 11–21. [Google Scholar]
- Hasan, M.M.; Rafii, M.Y.; Ismail, M.R.; Mahmood, M.; Rahim, H.A.; Alam, M.A.; Ashkani, S.; Malek, M.A.; Latif, M.A. Marker-assisted backcrossing: A useful method for rice improvement. Biotechnol. Biotechnol. Equip. 2015, 29, 237–254. [Google Scholar] [CrossRef]
- Waziri, A.; Kumar, P.; Purty, R.S. Saltol QTL and their role in salinity tolerance in rice. Austin J. Biotechnol. Bioeng. 2016, 3, 1067. [Google Scholar]
- Dixit, S.; Yadaw, R.B.; Mishra, K.K.; Kumar, A. Marker-assisted breeding to develop the drought-tolerant version of Sabitri, a popular variety from Nepal. Euphytica 2017, 213, 184. [Google Scholar] [CrossRef] [PubMed]
- Das, G.; Rao, G.J.; Varier, M.; Prakash, A.; Prasad, D. Improved Tapaswini having four BB resistance genes pyramided with six genes/QTLs, resistance/tolerance to biotic and abiotic stresses in rice. Sci. Rep. 2018, 8, 2413. [Google Scholar] [CrossRef] [PubMed]
- Kuraparthy, V.; Sood, S.; See, D.R.; Gill, B.S. Development of a PCR assay and marker-assisted transfer of leaf rust and stripe rust resistance genes Lr57 and Yr40 into hard red winter wheats. Crop Sci. 2009, 49, 120–126. [Google Scholar] [CrossRef]
- Vasistha, N.K.; Balasubramaniam, A.; Mishra, V.K.; Srinivasa, J.; Chand, R.; Joshi, A.K. Molecular introgression of leaf rust resistance gene Lr34 validates enhanced effect on resistance to spot blotch in spring wheat. Euphytica 2017, 213, 262. [Google Scholar] [CrossRef]
- Rai, K.N.; Hash, C.T.; Singh, A.K.; Velu, G. Adaptation and quality traits of a germplasm-derived commercial seed parent of pearl millet. Plant Genet. Resour. Newsl. 2008, 154, 20–24. [Google Scholar]
- Majka, M.; Kwiatek, M.; Korbas, M.; Danielewicz, J.; Gawlowska, M.; Góral, T.; Wisniewska, H. Eyespot resistance of winter wheat breeding lines evaluated with marker-assisted selection and inoculation tests at the seedling and adult plant stages. J. Plant Prot. Res. 2018, 58, 387–394. [Google Scholar] [CrossRef]
- Pellio, B.; Streng, S.; Bauer, E.; Stein, N.; Perovic, D.; Schiemann, A.; Friedt, W.; Ordon, F.; Graner, A. High-resolution mapping of the Rym4/Rym5 locus conferring resistance to the barley yellow mosaic virus complex (BaMMV, BaYMV, BaYMV-2) in barley (Hordeum vulgare ssp. vulgare L.). Theor. Appl. Genet. 2005, 110, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Yaeno, T.; Wahara, M.; Nagano, M.; Wanezaki, H.; Toda, H.; Inoue, H.; Eishima, A.; Nishiguchi, M.; Hisano, H.; Kobayashi, K.; et al. RACE1, a Japanese Blumeria graminis f. sp. hordei isolate, is capable of overcoming partially mlo-mediated penetration resistance in barley in an allele-specific manner. PLoS ONE 2021, 16, e0256574. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, B.M.; Palacios-Rojas, N.; Hossain, F.; Muthusamy, V.; Menkir, A.; Dhliwayo, T.; Ndhlela, T.; San Vicente, F.; Nair, S.K.; Vivek, B.S.; et al. Molecular breeding for nutritionally enriched maize: Status and prospects. Front. Genet. 2020, 10, 1392. [Google Scholar] [CrossRef]
- Hossain, F.; Muthusamy, V.; Pandey, N.; Vishwakarma, A.K.; Baveja, A.; Zunjare, R.U.; Thirunavukkarasu, N.; Saha, S.; Manjaiah, K.M.; Prasanna, B.M.; et al. Marker-assisted introgression of opaque2 allele for rapid conversion of elite hybrids into quality protein maize. J. Genet. 2018, 97, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Arelli, P.R.; Young, L.D.; Mengistu, A. Registration of high yielding and multiple disease-resistant soybean germplasm JTN-5503. Crop Sci. 2006, 46, 2723. [Google Scholar] [CrossRef]
- Arelli, P.R.; Young, L.D.; Concibido, V.C. Inheritance of resistance in soybean PI 567516C to LY1 nematode population infecting cv. Hartwig. Euphytica 2009, 165, 1–4. [Google Scholar] [CrossRef]
- Smith, J. USDA, ARS, National Genetic Resources Program. In Germplasm Resources Information Network-(GRIN); National Germplasm Resources Laboratory: Beltsville, MA, USA, 2010. [Google Scholar]
- Varshney, R.K.; Pandey, M.K.; Janila, P.; Nigam, S.N.; Sudini, H.; Gowda, M.V.C.; Sriswathi, M.; Radhakrishnan, T.; Manohar, S.S.; Nagesh, P. Marker-assisted introgression of a QTL region to improve rust resistance in three elite and popular varieties of peanut (Arachis hypogaea L.). Theor. Appl. Genet. 2014, 127, 1771–1781. [Google Scholar] [CrossRef]
- Varshney, R.K.; Penmetsa, R.V.; Dutta, S.; Kulwal, P.L.; Saxena, R.K.; Datta, S.; Sharma, T.R.; Rosen, B.; Carrasquilla-Garcia, N.; Farmer, A.D.; et al. Pigeonpea genomics initiative (PGI): An international effort to improve crop productivity of pigeonpea (Cajanus cajan L.). Mol. Breed. 2009, 26, 393–408. [Google Scholar] [CrossRef]
- Gautami, B.; Foncéka, D.; Pandey, M.K.; Moretzsohn, M.C.; Sujay, V.; Qin, H.; Hong, Y.; Faye, I.; Chen, X.; BhanuPrakash, A. An international reference consensus genetic map with 897 marker loci based on 11 mapping populations for tetraploid groundnut (Arachis hypogaea L.). PLoS ONE 2012, 7, e41213. [Google Scholar] [CrossRef]
- Khedikar, Y.P.; Gowda, M.V.; Sarvamangala, C.; Patgar, K.V.; Upadhyaya, H.D.; Varshney, R.K. A QTL study on late leaf spot and rust revealed one major QTL for molecular breeding for rust resistance in groundnut (Arachis hypogaea L.). Theor. Appl. Genet. 2010, 121, 971–984. [Google Scholar] [CrossRef]
- Sujay, V.; Gowda, M.V.; Pandey, M.K.; Bhat, R.S.; Khedikar, Y.P.; Nadaf, H.L.; Gautami, B.; Sarvamangala, C.; Lingaraju, S.; Radhakrishan, T. Quantitative trait locus analysis and construction of consensus genetic map for foliar disease resistance based on two recombinant inbred line populations in cultivated groundnut (Arachis hypogaea L.). Mol. Breed. 2012, 30, 773–788. [Google Scholar] [CrossRef] [PubMed]
- Sarvamangala, C.; Gowda, M.V.C.; Varshney, R.K. Identification of quantitative trait loci for protein content oil content and oil quality for groundnut (Arachis hypogaea L.). Field Crops Res. 2011, 122, 49–59. [Google Scholar] [CrossRef]
- Chu, Y.; Wu, C.L.; Holbrook, C.C.; Tillman, B.L.; Person, G.; Ozias-Akins, P. Marker-assisted selection to pyramid nematode resistance and the high oleic trait in peanut. Plant Genome 2011, 4, 2. [Google Scholar] [CrossRef]
- Varshney, R.K.; Mohan, S.M.; Gaur, P.M.; Chamarthi, S.K.; Singh, V.K.; Srinivasan, S.; Swapna, N.; Sharma, M.; Pande, S.; Singh, S.; et al. Marker-assisted backcrossing to introgress resistance to Fusarium wilt race 1 and Ascochyta blight in C 214, an elite cultivar of chickpea. Plant Genome 2014, 7, 1–11. [Google Scholar] [CrossRef]
- Anbessa, Y.; Taran, B.; Warkentin, T.D.; Tullu, A.; Vandenberg, A. Genetic analyses and conservation of QTL for ascochyta blight resistance in chickpea (Cicer arietinum L.). Theor. Appl. Genet. 2009, 119, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Anuradha, C.; Gaur, P.M.; Pande, S.; Gali, K.K.; Ganesh, M.; Kumar, J.; Varshney, R.K. Mapping QTL for resistance to botrytis grey mould in chickpea. Euphytica 2011, 182, 1–9. [Google Scholar] [CrossRef]
- Development of Two Superior Chickpea Varieties by Genomics Assisted Breeding. Available online: https://www.icar.org.in/node/12172 (accessed on 1 March 2024).
- Rehman, A.U.; Malhotra, R.S.; Bett, K.; Tar’an, B.; Bueckert, R.; Warkentin, T.D. Mapping QTL associated with traits affecting grain yield in chickpea (Cicer arietinum L.) under terminal drought stress. Crop Sci. 2011, 51, 450–463. [Google Scholar] [CrossRef]
- Vadez, V.; Krishnamurthy, L.; Thudi, M.; Anuradha, C.; Colmer, T.D.; Turner, N.C.; Siddique, K.H.; Gaur, P.M.; Varshney, R.K. Assessment of ICCV 2 × JG 62 chickpea progenies shows sensitivity of reproduction to salt stress and reveals QTL for seed yield and yield components. Mol. Breed. 2012, 30, 9–21. [Google Scholar] [CrossRef]
- Cobos, M.J.; Winter, P.; Kharrat, M.; Cubero, J.I.; Gil, J.; Milian, T.; Rubino, J. Genetic analysis of agronomic traits in a wide cross of chickpea. Field Crops Res. 2009, 111, 130–136. [Google Scholar] [CrossRef]
- Bajaj, D.; Saxena, M.S.; Kujur, A.; Das, S.; Badoni, S.; Tripathi, S.; Upadhyaya, H.D.; Gowda, C.L.L.; Sharma, S.; Singh, S.; et al. Genome-wide conserved non-coding microsatellite (CNMS) marker-based integrative genetical genomics for quantitative dissection of seed weight in chickpea. J. Exp. Bot. 2014, 66, 1271–1290. [Google Scholar] [CrossRef]
- Bajaj, D.; Upadhyaya, H.D.; Khan, Y.; Das, S.; Badoni, S.; Shree, T.; Kumar, V.; Tripathi, S.; Gowda, C.L.L.; Singh, S.; et al. A combinatorial approach of comprehensive QTL-based comparative genome mapping and transcript profiling identified a seed weight-regulating candidate gene in chickpea. Sci. Rep. 2015, 5, 9264. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Upadhyaya, H.D.; Bajaj, D.; Kujur, A.; Badoni, S.; Laxmi; Kumar, V.; Tripathi, S.; Gowda, C.L.; Sharma, S.; et al. Deploying QTL-seq for rapid delineation of a potential candidate gene underlying major trait-associated QTL in chickpea. DNA Res. 2015, 22, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Kujur, A.; Upadhyaya, H.D.; Shree, T.; Bajaj, D.; Das, S.; Saxena, M.S.; Badoni, S.; Kumar, V.; Tripathi, S.; Gowda, C.L.L.; et al. Ultra-high density intra-specific genetic linkage maps accelerate identification of functionally relevant molecular tags governing important agronomic traits in chickpea. Sci. Rep. 2015, 5, 9468. [Google Scholar] [CrossRef]
- Kujur, A.; Bajaj, D.; Upadhyaya, H.D.; Das, S.; Ranjan, R.; Shree, T.; Saxena, M.S.; Badoni, S.; Kumar, V.; Tripathi, S.; et al. A genome-wide SNP scan accelerates trait-regulatory genomic loci identification in chickpea. Sci. Rep. 2015, 5, 11166. [Google Scholar] [CrossRef] [PubMed]
- Daetwyler, H.D.; Villanueva, B.; Woolliams, J.A. Accuracy of predicting the genetic risk of disease using a genome-wide approach. PLoS ONE 2008, 3, e3395. [Google Scholar] [CrossRef] [PubMed]
- Goddard, M. Genomic selection: Prediction of accuracy and maximisation of long term response. Genetica 2009, 136, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Lu, Y.; Xie, C.; Gao, S.; Wan, J.; Prasanna, B.M. Whole-genome strategies for marker-assisted plant breeding. Mol. Breed. 2012, 29, 833–854. [Google Scholar] [CrossRef]
- Varshney, R.K.; Graner, A.; Sorrells, M.E. Genic microsatellite markers in plants: Features and applications. TRENDS Biotechnol. 2005, 23, 48–55. [Google Scholar] [CrossRef]
- Varshney, R.K.; Ribaut, J.M.; Buckler, E.S.; Tuberosa, R.; Rafalski, J.A.; Langridge, P. Can genomics boost productivity of orphan crops? Nat. Biotechnol. 2012, 30, 1172–1176. [Google Scholar] [CrossRef]
- Lucas, J.A. Advances in plant disease and pest management. J. Agric. Sci. 2011, 149, 91–114. [Google Scholar] [CrossRef]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Madhavi, K.R.; Prasad, M.S.; Madhav, M.S.; Laha, G.S.; Mohan, M.K.; Sundaram, R.M.; Jahanavi, B.; Vijitha, S.; Rao, P.R.; Virktamath, B.C. Introgression of blast resistance gene Pi-kh into elite indica rice variety Improved Samba Mahsuri. Ind. J. Plant Prot. 2012, 40, 52–56. [Google Scholar]
- Miedaner, T.; Korzun, V. Marker-assisted selection for disease resistance in wheat and barley breeding. Phytopathology 2012, 102, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Koenning, S.R.; Wrather, J.A. Suppression of soybean yield potential in the continental United States by plant diseases from 2006 to 2009. Plant Health Prog. 2010, 11, 5. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Fukao, T.; Ronald, P.; Ismail, A.; Heuer, S.; Mackill, D. Submergence tolerant rice: SUB1’s journey from landrace to modern cultivar. Rice 2010, 3, 138–147. [Google Scholar] [CrossRef]
- Ismail, A.M.; Singh, U.S.; Singh, S.; Dar, M.H.; Mackill, D.J. The contribution of submergence-tolerant (Sub1) rice varieties to food security in flood-prone rainfed lowland areas in Asia. Field Crops Res. 2013, 152, 83–93. [Google Scholar] [CrossRef]
- Thomson, M.J.; de Ocampo, M.; Egdane, J.; Rahman, M.A.; Sajise, A.G.; Adorada, D.L.; Tumimbang-Raiz, E.; Blumwald, E.; Seraj, Z.I.; Singh, R.K.; et al. Characterizing the Saltol quantitative trait locus for salinity tolerance in rice. Rice 2010, 3, 148–160. [Google Scholar] [CrossRef]
- Singh, A.K.; Gopalakrishnan, S.; Singh, V.P.; Prabhu, K.V.; Mohapatra, T.; Singh, N.K.; Sharma, T.R.; Nagarajan, M.; Vinod, K.K.; Singh, D.; et al. Marker assisted selection: A paradigm shift in Basmati breeding. Ind. J. Genet. Plant Breed. 2011, 71, 120. [Google Scholar]
- Luu, T.N.H.; Luu, M.C.; Abdelbagi, M.I.; Le, H.H. Introgression the salinity tolerance QTLs Saltol into AS996, the elite rice variety of Vietnam. Am. J. Plant Sci. 2012, 3, 981–987. [Google Scholar]
- Linh, L.H.; Linh, T.H.; Xuan, T.D.; Ham, L.H.; Ismail, A.M.; Khanh, T.D. Molecular breeding to improve salt tolerance of rice (Oryza sativa L.) in the Red River Delta of Vietnam. Int. J. Plant Genom. 2012, 2012, 949038. [Google Scholar] [CrossRef] [PubMed]
- Thu, N.T.; Quang, V.H.; Van Tan, M.; Vu, Ä.; Toshitsugu, N.; Hue, N.T.; Hue, N.T.; Long, N.V.; Van Hoan, N.; Van Liet, V.; et al. Marker-assisted pyramiding of Xa21 and Xa7 genes conferring resistance to bacterial leaf blight in indica cultivar Bacthom7. Afr. J. Biotechnol. 2018, 17, 1389–1396. [Google Scholar]
- Huyen, L.T.N.; Cuc, L.M.; Ham, L.H.; Khanh, T.D. Introgression the SALTOL QTL into Q5DB, the elite variety of Vietnam using marker-assisted-selection (MAS). Am. J. BioSci. 2013, 1, 80–84. [Google Scholar] [CrossRef]
- Hoque, A.B.M.Z.; Haque, M.A.; Sarker, M.R.A.; Rahman, M.A. Marker-assisted introgression of saltol locus into genetic background of BRRI Dhan-49. Int. J. BioSci. 2015, 6, 71–80. [Google Scholar]
- Usatov, A.V.; Alabushev, A.V.; Kostylev, P.I.; Azarin, K.V.; Makarenko, M.S.; Usatova, O.A. Introgression the saltol QTL into the elite rice variety of Russia by marker-assisted selection. Am. J. Agric. Biol. Sci. 2015, 10, 165–169. [Google Scholar] [CrossRef]
- Sharma, A.; Garg, S.; Sheikh, I.; Vyas, P.; Dhaliwal, H.S. Effect of wheat grain protein composition on end-use quality. J. Food Sci. Technol. 2020, 57, 2771–2785. [Google Scholar] [CrossRef]
- Brevis, J.C.; Dubcovsky, J. Effects of the chromosome region including the Gpc-B1 locus on wheat grain and protein yield. Crop Sci. 2010, 50, 93–104. [Google Scholar] [CrossRef]
- Abbai, R.; Singh, V.K.; Nachimuthu, V.V.; Sinha, P.; Selvaraj, R.; Vipparla, A.K.; Singh, A.K.; Singh, U.M.; Varshney, R.K.; Kumar, A. Haplotype analysis of key genes governing grain yield and quality traits across 3K RG panel reveals scope for the development of tailor-made rice with enhanced genetic gains. Plant Biotechnol. J. 2019, 17, 1612–1622. [Google Scholar] [CrossRef]
- Tanger, P.; Klassen, S.; Mojica, J.P.; Lovell, J.T.; Moyers, B.T.; Baraoidan, M.; Naredo, M.E.B.; McNally, K.L.; Poland, J.; Bush, D.R.; et al. Field-based high throughput phenotyping rapidly identifies genomic regions controlling yield components in rice. Sci. Rep. 2017, 7, 42839. [Google Scholar] [CrossRef]
- Xie, K.; Yang, Y. RNA-Guided Genome Editing in Plants Using a CRISPR-Cas System. Mol. Plant 2013, 6, 1975–1980. [Google Scholar] [CrossRef]
- Fuentes, A.; Yoon, S.; Kim, S.C.; Park, D.S. A robust deep-learning-based detector for real-time tomato plant diseases and pests’ recognition. Sensors 2017, 17, 2022. [Google Scholar] [CrossRef]
- Aquino, A.; Diago, M.P.; Millán, B.; Tardáguila, J. A new methodology for estimating the grapevine-berry number per cluster using image analysis. Biosyst. Eng. 2017, 156, 80–95. [Google Scholar] [CrossRef]
- Sinha, P.; Singh, V.K.; Saxena, R.K.; Khan, A.W.; Abbai, R.; Chitikineni, A.; Desai, A.; Molla, J.; Upadhyaya, H.D.; Kumar, A.; et al. Superior haplotypes for haplotype-based breeding for drought tolerance in pigeonpea (Cajanus cajan L.). Plant Biotechnol. J. 2020, 18, 2482–2490. [Google Scholar] [CrossRef]
- Brinton, J.; Ramirez-Gonzalez, R.H.; Simmonds, J.; Wingen, L.; Orford, S.; Griffiths, S.; Haberer, G.; Spannagl, M.; Walkowiak, S.; Pozniak, C.; et al. A haplotype-led approach to increase the precision of wheat breeding. Commun. Biol. 2020, 3, 712. [Google Scholar] [CrossRef] [PubMed]
- Silva-Perez, V.; Molero, G.; Serbin, S.P.; Condon, A.G.; Reynolds, M.P.; Furbank, R.T.; Evans, J.R. Hyperspectral reflectance as a tool to measure biochemical and physiological traits in wheat. J. Exp. Bot. 2017, 69, 483–496. [Google Scholar] [CrossRef] [PubMed]
- Montesinos-López, A.; Montesinos-López, O.A.; Cuevas, J.; Mata-López, W.A.; Burgueño, J.; Mondal, S.; Huerta, J.; Singh, R.; Autrique, E.; González-Pérez, L.; et al. Genomic Bayesian functional regression models with interactions for predicting wheat grain yield using hyper-spectral image data. Plant Methods 2017, 13, 62. [Google Scholar] [CrossRef]
- Jain, M.; Srivastava, A.K.; Joon, R.K.; McDonald, A.; Royal, K.; Lisaius, M.C.; Lobell, D.B. Mapping smallholder wheat yields and sowing dates using micro-satellite data. Remote Sens. 2016, 8, 860. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, X.; Shan, Q.; Zhang, Y.; Liu, J.; Gao, C.; Qiu, J.L. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat. Biotechnol. 2014, 32, 947–951. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.; Ghosh, S.; Williams, M.J.; Cuddy, W.S.; Simmonds, J.; Rey, M.D.; Asyraf Md Hatta, M.; Hinchliffe, A.; Steed, A.; Reynolds, D.; et al. Speed breeding is a powerful tool to accelerate crop research and breeding. Nat. Plants 2018, 4, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Araus, J.L.; Kefauver, S.C.; Zaman-Allah, M.; Olsen, M.S.; Cairns, J.E. Translating high-throughput phenotyping into genetic gain. Trends Plant Sci. 2018, 23, 451–466. [Google Scholar] [CrossRef]
- Miller, N.D.; Haase, N.J.; Lee, J.; Kaeppler, S.M.; de Leon, N.; Spalding, E.P. A robust, high-throughput method for computing maize ear, cob, and kernel attributes automatically from images. Plant J. 2017, 89, 169–178. [Google Scholar] [CrossRef]
- Quemada, M. Airborne hyperspectral images and ground-level optical sensors as assessment tools for maize nitrogen fertilization. Remote Sens. 2014, 6, 2940–2962. [Google Scholar] [CrossRef]
- Elazab, A.; Ordóñez, R.A.; Savin, R.; Slafer, G.A.; Araus, J.L. Detecting interactive effects of N fertilization and heat stress on maize productivity by remote sensing techniques. Eur. J. Agron. 2016, 73, 11–24. [Google Scholar] [CrossRef]
- Yendrek, C.R.; Tomaz, T.; Montes, C.M.; Cao, Y.; Morse, A.M.; Brown, P.J.; McIntyre, L.M.; Leakey, A.D.; Ainsworth, E.A. High-throughput phenotyping of maize leaf physiological and biochemical traits using hyperspectral reflectance. Plant Physiol. 2017, 173, 614–626. [Google Scholar] [CrossRef] [PubMed]
- Zaman-Allah, M.; Vergara, O.; Araus, J.L.; Tarekegne, A.; Magorokosho, C.; Zarco-Tejada, P.J.; Hornero, A.; Albà, A.H.; Das, B.; Craufurd, P.; et al. Unmanned aerial platform-based multi-spectral imaging for field phenotyping of maize. Plant Methods 2015, 11, 35. [Google Scholar] [CrossRef] [PubMed]
- Hämmerle, M.; Höfle, B. Direct derivation of maize plant and crop height from low-cost time-of-flight camera measurements. Plant Methods 2016, 12, 50. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Wang, K.; Huang, C.; Zhang, X.; Yan, J.; Yang, W. A high-throughput maize kernel traits scorer based online-scan imaging. Measurement 2016, 90, 453–460. [Google Scholar] [CrossRef]
- Tesfaye, K.; Sonder, K.; Caims, J.; Magorokosho, C.; Tarekegn, A.; Kassie, G.T.; Getaneh, F.; Abdoulaye, T.; Abate, T.; Erenstein, O. Targeting drought-tolerant maize varieties in southern Africa: A geospatial crop modeling approach using big data. Int. Food Agribus. Manag. Rev. 2016, 19, 75–92. [Google Scholar]
- Shi, J.; Gao, H.; Wang, H.; Lafitte, H.R.; Archibald, R.L.; Yang, M.; Hakimi, S.M.; Mo, H.; Habben, J.E. ARGOS8 variants generated by CRISPR-Cas9 improve maize grain yield under field drought stress conditions. Plant Biotechnol. 2017, 15, 207–216. [Google Scholar] [CrossRef]
- Acharjee, A.; Kloosterman, B.; Visser, R.G.; Maliepaard, C. Integration of multi-omics data for prediction of phenotypic traits using random forest. BMC Bioinform. 2016, 17 (Suppl. S5), 180. [Google Scholar] [CrossRef]
- Pauli, D.; Andrade-Sanchez, P.; Carmo-Silva, A.E.; Gazave, E.; French, A.N.; Heun, J.; Hunsaker, D.J.; Lipka, A.E.; Setter, T.L.; Strand, R.J.; et al. Field-based high-throughput plant phenotyping reveals the temporal patterns of quantitative trait loci associated with stress-responsive traits in cotton. G3 Genes Genomes Genet. 2016, 36, 865–879. [Google Scholar] [CrossRef]
- Zsögön, A.; Čermák, T.; Naves, E.R.; Notini, M.M.; Edel, K.H.; Weinl, S.; Freschi, L.; Voytas, D.F.; Kudla, J.; Peres, L.E.P. De novo domestication of wild tomato using genome editing. Nat. Biotechnol. 2018, 36, 1211–1216. [Google Scholar] [CrossRef]
- Li, T.; Yang, X.; Yu, Y.; Si, X.; Zhai, X.; Zhang, H.; Dong, W.; Gao, C.; Xu, C. Domestication of wild tomato is accelerated by genome editing. Nat. Biotechnol. 2018, 36, 1160–1163. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Leal, D.; Lemmon, Z.H.; Man, J.; Bartlett, M.E.; Lippman, Z.B. Engineering quantitative trait variation for crop improvement by genome editing. Cell 2017, 171, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, Z.H.; Reem, N.T.; Dalrymple, J.; Soyk, S.; Swartwood, K.E.; Rodriguez-Leal, D.; Van Eck, J.; Lippman, Z.B. Rapid improvement of domestication traits in an orphan crop by genome editing. Nat. Plants 2018, 4, 766–770. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Zhou, H.; Bi, H.; Fromm, M.; Yang, B.; Weeks, D.P. Demonstration of CRISPR/Cas9/sgRNA-mediated targeted gene modification in Arabidopsis, tobacco, sorghum and rice. Nucleic Acids Res. 2013, 41, e188. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Jacobs, T.B.; Xue, L.-J.; Harding, S.A.; Tsai, C.-J. Exploiting SNPs for biallelic CRISPR mutations in the outcrossing woody perennial Populus reveals 4-coumarate:CoA ligase specificity and redundancy. New Phytol. 2015, 208, 298–301. [Google Scholar] [CrossRef]
- Okuzaki, A.; Ogawa, T.; Koizuka, C.; Kaneko, K.; Inaba, M.; Imamura, J.; Koizuka, N. CRISPR/Cas9-mediated genome editing of the fatty acid desaturase 2 gene in Brassica napus. Plant Physiol. Biochem. 2018, 131, 63–69. [Google Scholar] [CrossRef]
- Huang, H.; Cui, T.; Zhang, L.; Yang, Q.; Yang, Y.; Xie, K.; Fan, C.; Zhou, Y. Modifications of fatty acid profile through targeted mutation at BnaFAD2 gene with CRISPR/Cas9-mediated gene editing in Brassica napus. Theor. Appl. Genet. 2020, 133, 2401–2411. [Google Scholar] [CrossRef]
- Jiang, W.Z.; Henry, I.M.; Lynagh, P.G.; Comai, L.; Cahoon, E.B.; Weeks, D.P. Significant enhancement of fatty acid composition in seeds of the allohexaploid, Camelina sativa, using CRISPR/Cas9 gene editing. Plant Biotechnol. J. 2017, 15, 648–657. [Google Scholar] [CrossRef]
- Morineau, C.; Bellec, Y.; Tellier, F.; Gissot, L.; Kelemen, Z.; Nogué, F.; Faure, J.D. Selective gene dosage by CRISPR-Cas9 genome editing in hexaploid Camelina sativa. Plant Biotechnol. J. 2017, 15, 729–739. [Google Scholar] [CrossRef]
- Ozseyhan, M.E.; Kang, J.; Mu, X.; Lu, C. Mutagenesis of the FAE1 genes significantly changes fatty acid composition in seeds of Camelina sativa. Plant Physiol. Biochem. 2018, 123, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Jähne, F.; Hahn, V.; Würschum, T.; Leiser, W.L. Speed breeding short-day crops by LED-controlled light schemes. Theor. Appl. Genet. 2020, 133, 2335–2342. [Google Scholar] [CrossRef]
- Yadav, D.K.; Hossain, F.; Choudhary, P.R.; Kumar, D.; Singh, A.K.; Mohapatra, T. Crop Cultivars Developed Through Molecular Breeding; Indian Council of Agricultural Research: New Delhi, India, 2020; p. 52. Available online: https://icar.org.in/sites/default/files/Circulars/Crop-Cultivars-2nd-Edition.pdf (accessed on 1 March 2024).
- Carpentieri-Pipolo, V. UEL 175: A novel lipoxygenase-free soybean cultivar with kunitz trypsin inhibitor absence. Crop Breed. Appl. Biotechnol. 2015, 15, 191–192. [Google Scholar] [CrossRef]
- Croser, J.S.; Pazos-Navarro, M.; Bennett, R.G.; Tschirren, S.; Edwards, K.; Erskine, W.; Creasy, R.; Ribalta, F.M. Time to flowering of temperate pulses in vivo and generation turnover in vivo–in vitro of narrow-leaf lupin accelerated by low red to far-red ratio and high intensity in the far-red region. Plant Cell Tissue Organ Cult. PCTOC 2016, 127, 591–599. [Google Scholar] [CrossRef]
- Razzaq, M.K.; Aleem, M.; Mansoor, S.; Khan, M.A.; Rauf, S.; Iqbal, S.; Siddique, K.H. Omics and CRISPR-Cas9 approaches for molecular insight, functional gene analysis, and stress tolerance development in crops. Int. J. Mol. Sci. 2021, 22, 1292. [Google Scholar] [CrossRef] [PubMed]
- Muthamilarasan, M.; Singh, N.K.; Prasad, M. Multi-omics approaches for strategic improvement of stress tolerance in underutilized crop species: A climate change perspective. Adv. Genet. 2019, 103, 1–8. [Google Scholar]
- Yang, Y.; Saand, M.A.; Huang, L.; Abdelaal, W.B.; Zhang, J.; Wu, Y.; Li, J.; Sirohi, M.H.; Wang, F. Applications of multi-omics technologies for crop improvement. Front. Plant Sci. 2021, 12, 563953. [Google Scholar] [CrossRef]
- Niu, Y.; Wu, L.; Li, Y.; Huang, H.; Qian, M.; Sun, W.; Zhu, H.; Xu, Y.; Fan, Y.; Mahmood, U.; et al. Deciphering the transcriptional regulatory networks that control size, color, and oil content in Brassica rapa seeds. Biotechnol. Biofuels 2020, 13, 90. [Google Scholar] [CrossRef]
- Coutinho, I.D.; Henning, L.M.M.; Döpp, S.A.; Nepomuceno, A.; Moraes, L.A.C.; Marcolino-Gomes, J.; Richter, C.; Schwalbe, H.; Colnago, L.A. Flooded soybean metabolomic analysis reveals important primary and secondary metabolites involved in the hypoxia stress response and tolerance. Environ. Exp. Bot. 2018, 153, 176–187. [Google Scholar] [CrossRef]
- Templer, S.E.; Ammon, A.; Pscheidt, D.; Ciobotea, O.; Schuy, C.; McCollum, C.; Sonnewald, U.; Hanemann, A.; Förster, J.; Ordon, F.; et al. Metabolite profiling of barley flag leaves under drought and combined heat and drought stress reveals metabolic QTLs for metabolites associated with antioxidant defense. J. Exp. Bot. 2017, 68, 1697–1713. [Google Scholar] [CrossRef]
- Sauvage, C.; Segura, V.; Bauchet, G.; Stevens, R.; Do, P.T.; Nikoloski, Z.; Fernie, A.R.; Causse, M. Genome-wide association in tomato reveals 44 candidate loci for fruit metabolic traits. Plant Physiol. 2014, 165, 1120–1132. [Google Scholar] [CrossRef] [PubMed]
- Weckwerth, W. Green systems biology—From single genomes, proteomes and metabolomes to ecosystems research and biotechnology. J. Proteom. 2011, 75, 284–305. [Google Scholar] [CrossRef]
- Luo, J. Metabolite-based genome-wide association studies in plants. Curr. Opin. Plant Biol. 2015, 24, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, F.; Nakabayashi, R.; Yang, Z.; Okazaki, Y.; Yonemaru, J.I.; Ebana, K.; Yano, M.; Saito, K. Metabolome-genome-wide association study dissects genetic architecture for generating natural variation in rice secondary metabolism. Plant J. 2015, 81, 13–23. [Google Scholar] [CrossRef]
- Talukdar, D.; Sinjushin, A. Cytogenomics and mutagenomics in plant functional biology and breeding. In PlantOmics: The Omics of Plant Science; Barh, D., Khan, M., Davies, E., Eds.; Springer: New Dehli, India, 2015; pp. 113–156. [Google Scholar] [CrossRef]
- Weckwerth, W.; Ghatak, A.; Bellaire, A.; Chaturvedi, P.; Varshney, R.K. PANOMICS meets germplasm. Plant Biotechnol. J. 2020, 18, 1507–1525. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.D.; Holzinger, E.R.; Li, R.; Pendergrass, S.A.; Kim, D. Methods of integrating data to uncover genotype–phenotype interactions. Nat. Rev. Genet. 2015, 16, 85–97. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Crop Species | Trait | Genes/Locus | Approach Used | Improved Variety Developed | References |
|---|---|---|---|---|---|
| Cereals and multiple (biotic/or abiotic) stress tolerance with quality traits | |||||
| Oryza sativa (Rice) | Bacterial blight disease resistance | Xa21, Xa13, and Xa5 | MAS | Improved ‘Samba Mahsuri’ (ISM) | [28] |
| Blast disease resistance | Blast resistance genes (Pi-2 and Pi-54); BB (Xa38) | Gene stacking | Improved ‘Samba Mahsuri’ (ISM) | [20,29] | |
| Blast disease resistance | Pi2, Pi5, Pi54, Pi1, Pita | Gene stacking | Pusa Basmati 1 | [30] | |
| Blast and bacterial blight diseases | Pi2 and Pi54 | MABC | Pusa Basmati 1121 | [31] | |
| Xa13 and Xa21 | MABC | Pusa Basmati 6 | [31] | ||
| Submergence tolerance | Sub1 | MAS | Swarna, ‘Samba Mahsuri’ (BPT 5204), ‘CR 1009’, ‘Thadokkham 1’ (TDK1) OM1490/IR64-Sub1cross (in Vietnam); ‘BR 11’ | [32,33,34] | |
| Salt tolerance | Saltol | MAS | ‘Pusa Basmati 1121’, ‘Pusa Basmati 6’, BRRI-Dhan 49’ ‘AS 996’, ‘BT 7’, ‘Q5DB ‘Bacthom 7’ | [35] | |
| Drought tolerance and high yield | MAS | ‘Sabitri’ | [36] | ||
| Blast and gall midge disease resistance submergence and salinity stress tolerance | Pi2, Pi9, Gm1, Gm4, Xa4, Xa5, Xa13, Xa2 | Gene pyramiding | Improved Tapaswini | [37] | |
| Improved fragrance and intermediate amylose content | badh2 and Wx | Transfer of variant allele from basmati into ‘Manawthukha’ | Improved ‘Manawthukha’ (from Myanmar) | [21] | |
| Triticum aestivum (Wheat) | Stress response and quality traits | Yr40/Lr57 and Lr58 | MAS | ‘Jagger’ and ‘Overley’ (hard red winter wheat (HRWW)) cultivars | [38] |
| Stress response and quality traits | Lr34 | MAS | Spring wheat cultivar ‘HUW510’ | [39] | |
| Improving protein content | Gpc-B1 | MAS | High GPC wheat cultivars Farnum’, ‘Lassik’, ‘Westmore’, and ‘Desert King-High Protein’, (‘Lillian ‘Somerset’, ‘Burnside’), ‘VR 1128’ Wyalkatchem’, ‘Gladius’ | [24] | |
| (Pennisetum glaucum (L.) R. Br.) (Pearl millet) | Downy mildew resistance, drought tolerance, and fodder quality | Downy mildew-resistant genes/locus (QRsg.icp-4.1/4.2; Qgydt.icp-2.1/6.1; 863B) | MAS | HHB 67-improved | [40] |
| Hordeum vulgare (Barley) | Eyespot (Rhizoctonia cerealis) resistance | resistance gene Pch1 | MAS | DD 248/12, KBP 15.2, and STH 4431 | [41] |
| Barley yellow mosaic viruses’ resistance | rym4/rym5 | MAS | Improved winter barley | [42] | |
| Barley powdery mildew disease resistance | mlo | Gene silencing | Improved resistance in barley | [43] | |
| Zea mays (Maize) | Nutritional traits (QPM, pro-vitamin A) | opaque2 | Transferred in elite hybrids (MAS) | Pusa Vivek QPM-9, Pusa Vivek Hybrid-27, Pusa HQPM5, Pusa HQPM7, | [44,45] |
| Legumes and multiple (biotic/or abiotic) stress tolerance with quality traits | |||||
| Glycine max (Soybean) | Multiple soybean cyst nematode (Heterodera glycines) races (2, 3, 5, and 14) | Multiple disease resistance and high-yielding genes | Gene stacking/ pyramiding | ‘JTN 5503’, ‘JTN 5303’, ‘DS 880’, and ‘JTN 5109’ | [46,47,48] |
| Arachis hypogaea (Groundnut) | Rust (Puccinia arachidis) resistance | Increased yield and rust resistance genes | MAS | ‘ICGV 91114’, ‘JL 24’, and ‘TAG 24’ (introgression lines) | [49] |
| Abiotic/biotic stress | Rust and late leaf spot resistance, stress adaptation-related traits | MAS | Resistant peanut | [22,23,50,51,52,53] | |
| Improved oil quality | Oil quality | MAS | Improved quality peanut | [54] | |
| High oleic acid content and nematode resistance | Tifguard high O/L’ | MAS/gene pyramiding | Reducing breeding cycle | [55] | |
| Cicer arietinum (Chickpea) | Wilt and blight disease resistance | foc1 locus, ABQTL-I, and ABQTL-II | MABC | C 214 | [56] |
| Biotic stresses | Ascochyta blight resistance and botrytis gray mold resistance genes | MAS | Biotic stress-resistant chickpea | [57,58] | |
| Drought tolerance traits | Introgression into Pusa 372 | QTL hotspot by MAS | ‘Pusa 10216’ | [59] | |
| Abiotic stresses | Terminal drought stress | QTL | Improved grain yield in Cicer arietinum | [60] | |
| Salt stress | QTL | Improved seed yield and their components in ICCV 2 × JG 62 Progenies | [61] | ||
| Quality traits | Seed weight-regulating gene | CNMS, genome mapping, and transcript profiling | Improved seed weight in chickpea | [60,62,63,64,65,66,67] | |
| Crop Species | Trait Improvement | Gene Targeted | Approach Used | References |
|---|---|---|---|---|
| Oryza sativa (Rice) | Grain yield and quality | Genetic gain-related genes | Large-scale whole-genome resequencing (WGRS) + haplo-pheno analysis | [90] |
| Yield component | Grain yield | High-throughput phenotyping | [91] | |
| Disease resistance | OsMPK5 | Gene editing | [92] | |
| Solanum lycopersicum (Tomato) | Disease resistance | Plant disease and pest recognition traits | Deep learning-based detectors | [93] |
| Vitis vinifera (Grapevine) | Yield component | Berry number/cluster | High-throughput phenotyping | [94] |
| Cajanus cajan (Pigeon pea) | Drought tolerance | Plant weight and relative water content | Haplotype breeding | [95] |
| Triticum aestivum (Wheat) | Yield-related traits | TaGW2-A | Haplotype breeding | [96] |
| Physiological and morphological traits | Nitrogen per unit leaf area, leaf dry mass per area, Rubisco activity, and electron transport rate | High-throughput phenotyping | [97] | |
| Yield component | Grain yield | High-throughput phenotyping | [98] | |
| Yield component | Yield and sowing date | Microsatellite data | [99] | |
| Powdery mildew diseases | TaMLO-A1 | Gene editing | [100] | |
| Australian high-protein milling variety (DS Faraday) | Tolerance to preharvest sprouting and leaf, stem rust, stripe | SB | [101] | |
| Zea mays (Maize) | All plant phenotypic traits (morphological, physiological), biotic and abiotic stress-related traits | Plant/canopy height, plant density, fruit size, grain number and size, diseases, senescence, chlorophyll content | High-throughput phenotyping | [102] |
| Qualitative/quantitative traits | Ear, cob, kernel | High-throughput phenotyping | [103] | |
| Fertilization | Nitrogen fertilization | High-throughput phenotyping | [104] | |
| Fertilization and abiotic stress | Nitrogen fertilization and heat stress | High-throughput phenotyping | [105] | |
| Physiological and biochemical traits in leaf | Chlorophyll content, nitrogen content, sucrose content, leaf area, photosynthesis rate, leaf oxygen radical, rate of phosphophenol pyruvate carboxylation | High-throughput phenotyping | [106] | |
| Stress resilience | Leaf/canopy reflectance for low-N stress | High-throughput phenotyping | [107] | |
| Yield component | Plant height | High-throughput phenotyping | [108] | |
| Yield component | Kernel trait | High-throughput phenotyping | [109] | |
| Drought tolerance | Drought tolerance-related genes | Big data | [110] | |
| Grain Yield trait under drought stress | AGROS8 | Gene editing | [111] | |
| Solanum tuberosum (Potato) | Tuber quality traits (phenotypic, physiological, biochemical, and molecular) | Tuber flesh color, DSC onset, tuber shape, and enzymatic discoloration | Integration of the multi-omics (transcriptomics, metabolomics, proteomics) data with the genetic mapping | [112] |
| Anthonomus grandis (Cotton) | Stress resilience | Stress-responsive traits | High-throughput phenotyping | [113] |
| Solanum pimpinellifolium (Wild tomato) | Quality and nutritional traits (self-pruning, fruit weight, lycopene beta cyclase) | SP, SP5G, SlCLV3, Sl SlWUS, | Genome editing for de novo domestication | [114,115,116] |
| Physalis pruinosa (Groundcherry) | Quality and nutritional traits SELF-PRUNING, SELF-PRUNING 5G, CLAVATA WUSCHEL | SP, SP5G, and CLV | Genome editing | [117] |
| Arabidopsis thaliana, Nicotiana tabacum, Sorghum bicolor, Oryza sativa (Arabidopsis, Tobacco, Sorghum, and Rice) | Bacterial blight | OsSWEET14 and OsSWEET11 | Gene editing | [118] |
| Populus tremula x alba | Lignin and flavonoid biosynthesis | 4-coumarate: CoA ligase (4CL) genes, 4CL1, 4CL2, and 4CL5 | Gene editing | [119] |
| Brassica napus (Rapeseed) | High oleic acid | FAD2 | Gene editing | [120,121] |
| Camelina sativa (Camelina/false flax) | High oleic acid | FAD2, FAE1 | Gene editing | [122,123,124] |
| Glycine max (Soybean) | 10 h photoperiod + blue light | Improve generation turnover. | Speed breeding | [125] |
| Kuntiz-trypsin inhibitor (KTI) free | NRC21, UEL175 | Molecular breeding and gene editing | [126,127] | |
| Light quality/intensity | Photo-morphogenic response | Speed breeding | [128] | |
| Oryza sativa and Amaranthus spp. (Rice and Amaranth) | Light quality/intensity | Improve generation turnover | Speed breeding | [125] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tyagi, A.; Mir, Z.A.; Almalki, M.A.; Deshmukh, R.; Ali, S. Genomics-Assisted Breeding: A Powerful Breeding Approach for Improving Plant Growth and Stress Resilience. Agronomy 2024, 14, 1128. https://doi.org/10.3390/agronomy14061128
Tyagi A, Mir ZA, Almalki MA, Deshmukh R, Ali S. Genomics-Assisted Breeding: A Powerful Breeding Approach for Improving Plant Growth and Stress Resilience. Agronomy. 2024; 14(6):1128. https://doi.org/10.3390/agronomy14061128
Chicago/Turabian StyleTyagi, Anshika, Zahoor Ahmad Mir, Mohammed A. Almalki, Rupesh Deshmukh, and Sajad Ali. 2024. "Genomics-Assisted Breeding: A Powerful Breeding Approach for Improving Plant Growth and Stress Resilience" Agronomy 14, no. 6: 1128. https://doi.org/10.3390/agronomy14061128
APA StyleTyagi, A., Mir, Z. A., Almalki, M. A., Deshmukh, R., & Ali, S. (2024). Genomics-Assisted Breeding: A Powerful Breeding Approach for Improving Plant Growth and Stress Resilience. Agronomy, 14(6), 1128. https://doi.org/10.3390/agronomy14061128