
Phytoplasma-Induced Leaf Reddening as a Monitoring Symptom of Apple Proliferation Disease with Regard to the Development of Remote Sensing Strategies



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Samples
2.2. Experimental Induction of Reddening
2.3. PCR Detection
2.4. Quantitative Real-Time PCR
2.5. AP Phytoplasma Subtype Characterisation by PCR-RFLP
2.6. Statistical Analyses
3. Results
3.1. Correlation of Typical Symptoms of AP with PCR Detection of ‘Ca. P. mali’
3.2. Correlation of Leaf Reddening with Typical Symptoms of AP
3.3. PCR Detection of ‘Ca. P. mali’ in Red-Leafed Trees without Typical Symptoms of AP
3.4. AP-Correlated Leaf Reddening in Different Cultivars
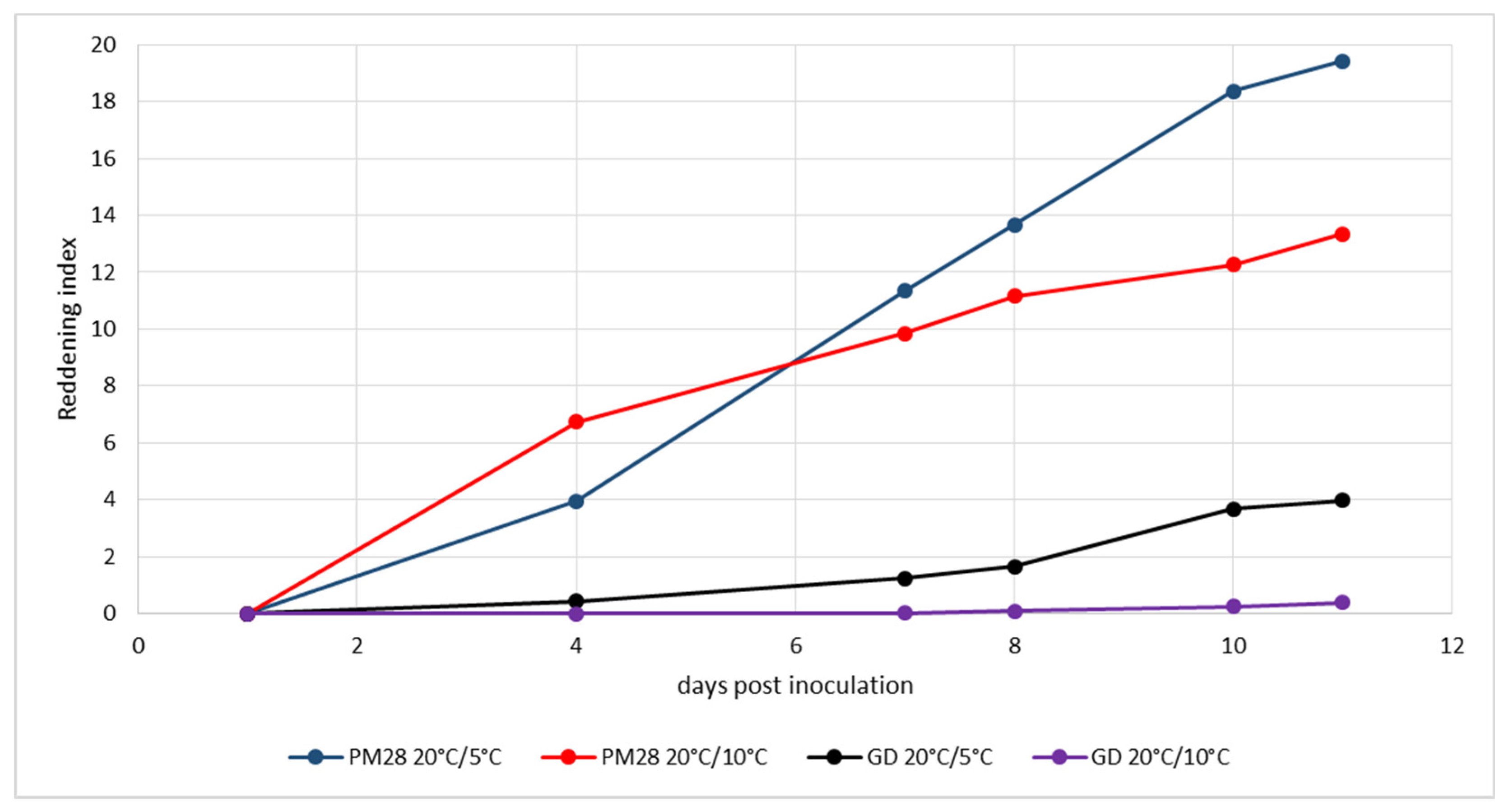
3.5. Experimental Induction of Phytoplasma-Induced Leaf Reddening
3.6. Correlation with Field Observations
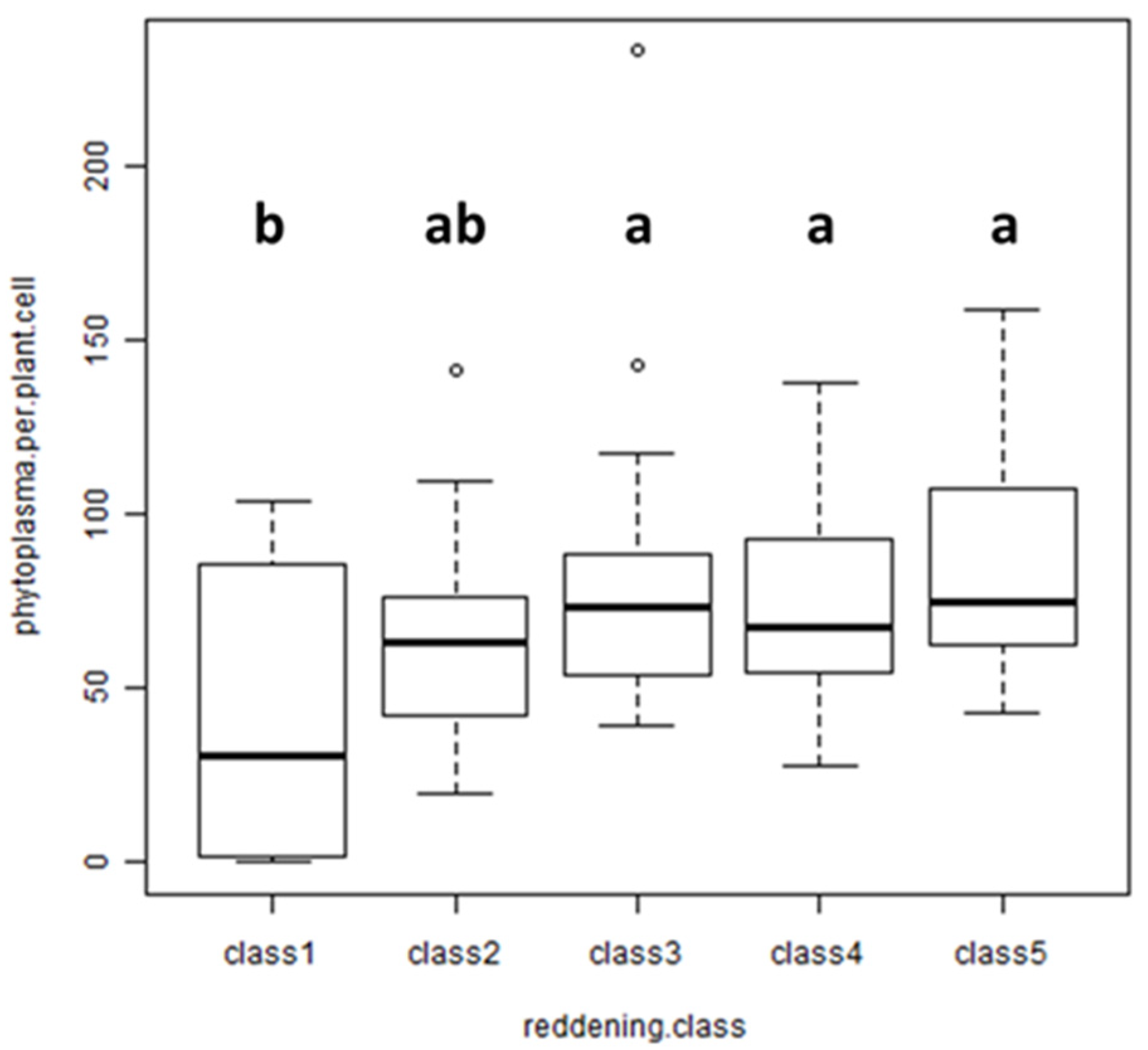
3.7. Correlation of Leaf Reddening with Phytoplasma Titer
3.8. Correlation of Leaf Reddening with ‘Ca. Phytoplasma mali’ Subtype
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seemüller, E.; Carraro, L.; Jarausch, W.; Schneider, B. Apple Proliferation Phytoplasma. In Virus and Virus-like Diseases of Pome and Stone Fruits; Hadidi, A., Barba, M., Candresse, T., Jelkmann, W., Eds.; APS Press: Saint Paul, MN, USA, 2011; pp. 67–73. ISBN 978-0-89054-396-2. [Google Scholar]
- Strauss, E. Microbiology. Phytoplasma research begins to bloom. Science 2009, 325, 388–390. [Google Scholar] [CrossRef] [PubMed]
- Seemüller, E.; Schneider, B. ‘Candidatus Phytoplasma mali’, ‘Candidatus Phytoplasma pyri’ and ‘Candidatus Phytoplasma prunorum’, the causal agents of apple proliferation, pear decline and European stone fruit yellows, respectively. Int. J. Syst. Evol. Microbiol. 2004, 54, 1217–1226. [Google Scholar] [CrossRef] [PubMed]
- Jarausch, B.; Tedeschi, R.; Sauvion, N.; Gross, J.; Jarausch, W. Psyllid vectors. In Phytoplasmas: Plant Pathogenic Bacteria—II. Transmission and Management of Phytoplasma—Associated Diseases; Bertaccini, A., Weintraub, P.G., Rao, G.P., Mori, N., Eds.; Springer: Singapore, 2019; pp. 53–78. [Google Scholar] [CrossRef]
- Barthel, D.; Fischnaller, S.; Letschka, T.; Janik, J.; Mittelberger, C.; Öttl, S.; Panassiti, B.; Angeli, G.; Baldessari, M.; Bianchedi, P.L.; et al. Scopazzi del Melo: Stato Attuale della Ricerca—Apfeltriebsucht: Aktueller Stand der Forschung; Janik, K., Barthel, D., Oppedisano, T., Anfora, G., Eds.; Fondazione Edmund Mach: San Michele all’Adige, Italy; Centro di Sperimentazione Laimburg: Ora, Italy, 2020; pp. 1–153. ISBN 9788878430532. [Google Scholar]
- Jarausch, W. Apfeltriebsucht. 2007. Available online: https://www.apfeltriebsucht.de (accessed on 4 September 2023).
- Mattedi, L.; Forno, F.; Branz, A.; Bragagna, P.; Battocletti, I.; Gualandri, V.; Pedrazzoli, F.; Bianchedi, P.; Deromedi, M.; Filippi, M.; et al. Come riconoscere la malattia in campo: Novità sulla sintomatologia. In Scopazzi del Melo—Apple Proliferation; Ioriatti, C., Jarausch, W., Eds.; Fondazione Edmund Mach: San Michele all’Adige, Italy, 2008; pp. 41–50. [Google Scholar]
- Barthel, D.; Dordevic, N.; Fischnaller, S.; Kerschbamer, C.; Messner, M.; Eisenstecken, D.; Robatscher, P.; Janik, K. Detection of apple proliferation disease in Malus × domestica by near infrared reflectance analysis of leaves. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2021, 263, 120178. [Google Scholar] [CrossRef]
- Berni, J.A.J.; Zarco-Tejada, P.J.; Suárez, L.; González-Dugo, V.; Fereres, E. Remote sensing of vegetation from UAV platforms using lightweight multispectral and thermal imaging sensors. Int. Arch. Photogramm. Remote Sens. Spatial Inform. Sci. 2009, 38, 6. [Google Scholar]
- Jones, H.G.; Robin, A.V. Remote Sensing of Vegetation: Principles, Techniques, and Applications, 1st ed.; Oxford University Press: New York, NY, USA, 2010. [Google Scholar]
- Boochs, F.; Kupfer, G.; Dockter, K.; Kühbauch, W. Shape of the red edge as vitality indicator for plants. Int. J. Remote Sens. 1990, 11, 1741–1753. [Google Scholar] [CrossRef]
- Sharifi, A. Remotely sensed vegetation indices for crop nutrition mapping. J. Sci. Food Agric. 2020, 100, 5191–5196. [Google Scholar] [CrossRef] [PubMed]
- Delalieux, S.; Somers, B.; Verstraeten, W.W.; van Aardt, J.A.N.; Keulemans, W.; Coppin, P. Hyperspectral indices to diagnose leaf biotic stress of apple plants, considering leaf phenology. Int. J. Remote Sens. 2009, 30, 1887–1912. [Google Scholar] [CrossRef]
- Kim, Y.J.; Glenn, D.M.; Park, J.; Ngugi, H.K.; Lehman, B.L. Hyperspectral image analysis for water stress detection of apple trees. Comput. Electron. Agric. 2011, 77, 155–160. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, D. Improving forest aboveground biomass estimation using seasonal Landsat NDVI time-series. ISPRS J. Photogramm. Remote Sens. 2015, 102, 222–231. [Google Scholar] [CrossRef]
- Liu, Z.; Guo, P.; Liu, H.; Fan, P.; Zeng, P.; Liu, X.; Feng, C.; Wang, W.; Yang, F. Gradient boosting estimation of the leaf area index of apple orchards in UAV remote sensing. Remote Sens. 2021, 13, 3263. [Google Scholar] [CrossRef]
- Li, C.; Zhu, X.; Wei, Y.; Cao, S.; Guo, X.; Yu, X.; Chang, C. Estimating apple tree canopy chlorophyll content based on Sentinel-2A remote sensing imaging. Sci Rep. 2018, 8, 3756. [Google Scholar] [CrossRef]
- Zhu, Y.; Yang, G.; Yang, H.; Wu, J.; Lei, L.; Zhao, F.; Fan, L.; Zhao, C. Identification of apple orchard planting year based on spatiotemporally fused satellite images and clustering analysis of foliage phenophase. Remote Sens. 2020, 12, 1199. [Google Scholar] [CrossRef]
- Zhang, C.; Valente, J.; Kooistra, L.; Guo, L.; Wang, W. Orchard management with small unmanned aerial vehicles: A survey of sensing and analysis approaches. Precis. Agric. 2021, 22, 2007–2052. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Y.; Jiang, D.; Zhang, Z.; Chang, Q. Quantitative assessment of apple mosaic disease severity based on hyperspectral images and chlorophyll content. Remote Sens. 2023, 15, 2202. [Google Scholar] [CrossRef]
- Pieczywek, P.M.; Cybulska, J.; Szymańska-Chargot, M.; Siedliska, A.; Zdunek, A.; Nosalewicz, A.; Baranowski, P.; Kurenda, A. Early detection of fungal infection of stored apple fruit with optical sensors–Comparison of biospeckle, hyperspectral imaging and chlorophyll fluorescence. Food Control 2018, 85, 327–338. [Google Scholar] [CrossRef]
- Genangeli, A.; Allasia, G.; Bindi, M.; Cantini, C.; Cavaliere, A.; Genesio, L.; Giannotta, G.; Miglietta, F.; Gioli, B. A Novel hyperspectral method to detect moldy core in apple fruits. Sensors 2022, 22, 4479. [Google Scholar] [CrossRef] [PubMed]
- Bertamini, M.; Grando, M.S.; Nedunchezhian, N. Effects of Phytoplasma Infection on Pigments, Chlorophyll-Protein Complex and Photosynthetic Activities in Field Grown Apple Leaves. Biol. Plant 2003, 47, 237–242. [Google Scholar] [CrossRef]
- Mittelberger, C.; Pichler, C.; Yalcinkaya, H.; Erhart, T.; Gasser, J.; Schumacher, S.; Janik, K.; Robatscher, P.; Kräutler, B.; Oberhuber, M. Pathogen-Induced Leaf Chlorosis: Products of Chlorophyll Breakdown Found in Degreened Leaves of Phytoplasma-Infected Apple (Malus × domestica Borkh.) and Apricot (Prunus armeniaca L.) Trees Relate to the Pheophorbide a Oxygenase/Phyllobilin Pathway. J. Agric. Food Chem. 2017, 65, 2651–2660. [Google Scholar] [CrossRef] [PubMed]
- Zorer, R.; Bianchedi, P.L.; Mattedi, L.; Branz, A. Arrossamento fogliare autunnale. Relazioni con la presenza del fitoplasma e possibili utilizzi nel monitoraggio prossimale o remoto. In Scopazzi del Melo—Apple Proliferation; Ioriatti, C., Jarausch, W., Eds.; Fondazione Edmund Mach: San Michele all’Adige, Italy, 2008; pp. 51–62. [Google Scholar]
- Barthel, D.; Cullinan, C.; Mejia-Aguilar, A.; Chuprikova, E.; McLeod, B.A.; Kerschbamer, C.; Trenti, M.; Monsorno, R.; Prechsl, U.E.; Janik, K. Identification of spectral ranges that contribute to phytoplasma detection in apple trees—A step towards an on-site method. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2023, 303, 123246. [Google Scholar] [CrossRef] [PubMed]
- Albetis, J.; Duthoit, S.; Guttler, F.; Jacquin, A.; Goulard, M.; Poilvé, H.; Féret, J.-B.; Dedieu, G. Detection of Flavescence dorée grapevine disease using unmanned aerial vehicle (UAV) multispectral imagery. Remote Sens. 2017, 9, 308. [Google Scholar] [CrossRef]
- Albetis, J.; Jacquin, A.; Goulard, M.; Poilvé, H.; Rousseau, J.; Clenet, H.; Dedieu, G.; Duthoit, S. On the potentiality of UAV multispectral imagery to detect Flavescence dorée and grapevine trunk diseases. Remote Sens. 2018, 11, 23. [Google Scholar] [CrossRef]
- Jarausch, W.; Menz, P.; Al Masri, A.; Runne, R.; Thielert, B.; Kohler, K.; Warnemünde, S.; Kilias, D.; Jarausch, B.; Knauer, U. Digital Phytoplasmology: Remote sensing of fruit tree phytoplasma diseases. Phytopath. Mollicutes 2023, 13, 135–136. [Google Scholar] [CrossRef]
- Musetti, R.; Paolacci, A.; Ciaffi, M.; Tanzarella, O.A.; Polizzotto, R.; Tubaro, F.; Mizzau, M.; Ermacora, P.; Badiani, M.; Osler, R. Phloem cytochemical modification and gene expression following the recovery of apple plants from apple proliferation disease. Phytopathology 2010, 100, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Jarausch, W.; Saillard, C.; Helliot, B.; Garnier, M.; Dosba, F. Genetic variability of apple proliferation phytoplasmas as determined by PCR-RFLP and sequencing of a non-ribosomal fragment. Mol. Cell. Probes 2000, 14, 17–24. [Google Scholar] [CrossRef]
- Jarausch, B.; Schwind, N.; Fuchs, A.; Jarausch, W. Characteristics of the spread of apple proliferation by its vector Cacopsylla picta. Phytopathology 2011, 101, 1471–1480. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, K.H.; Schneider, B.; Ahrens, U.; Seemüller, E. Detection of the apple proliferation and pear decline phytoplasmas by PCR amplification of ribosomal and nonribosomal DNA. Phytopathology 1995, 85, 771–776. [Google Scholar] [CrossRef]
- Schneider, B.; Seemüller, E.; Smart, C.D.; Kirkpatrick, B.C. Phylogenetic classification of plant pathogenic mycoplasma-like organism or phytoplasmas. In Molecular and Diagnostic Procedures in Mycoplasmology; Razin, S., Tully, J.G., Eds.; Academic Press: San Diego, CA, USA, 1995; pp. 369–380. [Google Scholar]
- Ahrens, U.; Seemüller, E. Detection of DNA of plant pathogenic mycoplasmalike organisms by a polymerase chain-reaction that amplifies a sequence of the 16S RNA gene. Phytopathology 1992, 82, 828–832. [Google Scholar] [CrossRef]
- Jarausch, W.; Peccerella, T.; Schwind, N.; Jarausch, B.; Krczal, G. Establishment of a quantitative real-time PCR assay for the quantification of apple proliferation phytoplasmas in plants and insects. Acta Hort. 2004, 657, 415–420. [Google Scholar] [CrossRef]
- Liebenberg, A. Influence of Latent Apple Viruses on Malus sieboldii-Derived Apple Proliferation Resistant Rootstocks. Ph.D. Thesis, Heidelberg University, Heidelberg, Germany, 2013; p. 165. [Google Scholar]
- Han, Y.; Gasic, K.; Sun, F.; Xu, M.; Korban, S.S. A gene encoding starch branching enzyme I (SBEI) in apple (Malus × domestica, Rosaceae) and its phylogenetic relationship to Sbe genes from other angiosperms. Mol. Phylogenetic Evol. 2007, 43, 852–863. [Google Scholar] [CrossRef]
- Jarausch, W.; Saillard, C.; Dosba, F.; Bové, J.M. Differentiation of mycoplasmalike organisms (MLOs) in European fruit trees by PCR using specific primers derived from the sequence of a chromosomal fragment of the apple proliferation MLO. Appl. Environ. Microbiol. 1994, 60, 2916–2923. [Google Scholar] [CrossRef]
- Seemüller, E.; Moll, E.; Schneider, B. Apple proliferation resistance of Malus sieboldii-based rootstocks in comparison to rootstocks derived from other Malus species. Eur. J. Plant Pathol. 2008, 121, 109–119. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org/ (accessed on 17 November 2023).
- Seemüller, E. Apple proliferation. In Compendium of Apple and Pear Diseases; Jones, A.L., Aldwinkle, H.S., Eds.; APS Press: St. Paul, MN, USA, 1990; pp. 67–68. [Google Scholar]
- Bertamini, M.; Muthuchelian, K.; Grando, M.S.; Nedunchezhian, N. Effects of phytoplasma infection on growth and photosynthesis in leaves of field grown apple. Photosynthetica 2002, 40, 157–160. [Google Scholar] [CrossRef]
- Aldaghi, M.; Bertaccini, A.; Lepoivre, P. cDNA-AFLP analysis of gene expression changes in apple trees induced by phytoplasma infection during compatible interaction. Eur. J. Plant Pathol. 2012, 134, 117–130. [Google Scholar] [CrossRef]
- Öttl, S.; Baric, S.; Dalla Via, J. Teilweise Rotfärbung weist nicht auf Apfeltriebsucht hin. Obstbau Weinbau 2008, 2, 58–59. [Google Scholar]
- Schaper, U.; Seemüller, E. Condition of the phloem and the persistence of mycoplasmalike organisms associated with apple proliferation and pear decline. Phytopathology 1982, 72, 736–742. [Google Scholar] [CrossRef]
- Jarausch, W.; Schwind, N.; Jarausch, B.; Krczal, G. Analysis of the distribution of apple proliferation phytoplasma subtypes in a local fruit growing region in Southwest Germany. Acta Hort. 2004, 657, 421–424. [Google Scholar] [CrossRef]
- Seemüller, E.; Schneider, B. Differences in Virulence and Genomic Features of Strains of ‘Candidatus Phytoplasma mali’, the Apple Proliferation Agent. Phytopathology 2007, 97, 964–970. [Google Scholar] [CrossRef] [PubMed]
- Cieślińska, M.; Hennig, E.; Kruczyńska, D.; Bertaccini, A. Genetic diversity of ‘Candidatus Phytoplasma mali’ strains in Poland. Phytopathol. Mediterr. 2015, 54, 477–487. [Google Scholar]
- Martini, M.; Ermacora, P.; Falginella, L.; Loi, N.; Carraro, L. Molecular differentiation of ‘Candidatus Phytoplasma mali’ and its spreading in Friuli Venezia Giulia Region (north-east Italy). Acta Hort. 2008, 781, 395–402. [Google Scholar] [CrossRef]
- Casati, P.; Quaglino, F.; Tedeschi, R.; Spiga, M.F.; Alma, A.; Spadone, P. Identification and molecular characterization of ‘Candidatus Phytoplasma mali’ isolates in Northwestern Italy. J. Phytopath. 2010, 158, 81–87. [Google Scholar] [CrossRef]
- Cainelli, C.; Bisognin, C.; Vindimian, M.E.; Grando, M.S. Genetic variability of AP phytoplasmas detected in the apple growing area of Trentino (Nord Italy). Acta Hort. 2004, 657, 425–430. [Google Scholar] [CrossRef]
- Baric, S.; Berger, J.; Cainelli, C.; Kerschbamer, C.; Dalla Via, J. Molecular typing of ‘Candidatus Phytoplasma mali’ and epidemic history tracing by a combined T-RFLP/VNTR analysis approach. Eur. J. Plant Pathol. 2011, 131, 573–584. [Google Scholar] [CrossRef]
- Mayer, C.J.; Jarausch, B.; Jarausch, W.; Jelkmann, W.; Vilcinskas, A.; Gross, J. Cacopsylla melanoneura has no relevance as vector of apple proliferation in Germany. Phytopathology 2009, 99, 729–738. [Google Scholar] [CrossRef]
- Malagnini, V.; Pedrazzoli, F.; Papetti, C.; Cainelli, C.; Zasso, R.; Gualandri, V.; Pozzebon, A.; Ioriatti, C. Ecological and Genetic Differences between Cacopsylla melanoneura (Hemiptera, Psyllidae) Populations Reveal Species Host Plant Preference. PLoS ONE 2013, 8, e69663. [Google Scholar] [CrossRef] [PubMed][Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Local Fruit-Growing Region | No. Orchards Monitored | No. Trees Monitored | No. Trees with Partial Reddening | No. Trees with Entire Reddening | % Trees with Reddening in Total |
|---|---|---|---|---|---|---|
| 2019 | Meckenheim | 6 | 3819 | 589 | 95 | 17.9% |
| Ilbesheim | 8 | 1730 | 230 | 58 | 16.6% | |
| 2020 | Erpolzheim–Weisenheim | 12 | 615 | 30 | 47 | 12.5% |
| Meckenheim | 5 | 2501 | 548 | 200 | 29.9% | |
| Ilbesheim | 14 | 2802 | 271 | 33 | 10.8% | |
| Winden | 10 | 1895 | 311 | 158 | 24.7% | |
| 2021 | Erpolzheim–Weisenheim | 21 | 2494 | 403 | 170 | 23.0% |
| Meckenheim | 8 | 2629 | 367 | 197 | 21.5% | |
| Ilbesheim | 9 | 693 | 88 | 28 | 16.7% | |
| Winden | 4 | 561 | 158 | 32 | 33.9% | |
| 2022 | Meckenheim | 2 | 849 | 237 | 115 | 41.5% |
| Ilbesheim | 3 | 336 | 81 | 53 | 39.9% |
| Symptom | 2019 | 2020 | 2021 | 2022 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. Orchards | No. Positive/ Total Samples | % Correlation | No. Orchards | No. Positive/ Total Samples | % Correlation | No. Orchards | No. Positive/ Total Samples | % Correlation | No. Orchards | No. Positive/ Total Samples | % Correlation | |
| Witches’ broom Enlarged stipule Stunted branch Small-sized fruit Leaf reddening alone | 6 | 36/36 | 100 | 5 | 18/18 | 100 | 2 | 8/8 | 100 | 2 | 10/10 | 100 |
| 10 | 76/77 | 98.70 | 8 | 71/72 | 98.61 | 2 | 18/18 | 100 | 1 | 12/12 | 100 | |
| 1 | 2/2 | 100 | 2 | 5/5 | 100 | 2 | 5/5 | 100 | 1 | 2/2 | 100 | |
| 7 | 15/15 | 100 | 6 | 28/28 | 100 | 2 | 4/4 | 100 | 1 | 3/3 | 100 | |
| 12 | 125/142 | 88.03 | 27 | 123/146 | 84.25 | 22 | 98/119 | 82.25 | 5 | 72/101 | 71.29 | |
| Partial leaf reddening Leaf reddening of entire crown | 11 | 90/106 | 84.91 | 21 | 66/78 | 84.62 | 13 | 45/52 | 86.54 | 5 | 25/35 | 71.43 |
| 11 | 35/36 | 97.22 | 19 | 57/68 | 83.82 | 18 | 53/67 | 79.10 | 4 | 47/66 | 71.21 |
| Symptom | 2019 | 2020 | 2021 | 2022 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. Orchards | No. Trees Reddened/ Total | % Correlation | No. Orchards | No. Trees Reddened/ Total | % Correlation | No. Orchards | No. Trees Reddened/ Total | % Correlation | No. Orchards | No. Trees Reddened/ Total | % Corre-lation | |
| Witches’ broom Enlarged stipule Small-sized fruit Stunted branch | 12 | 102/106 | 96.23 | 27 | 222/236 | 94.07 | 31 | 328/333 | 98.50 | 3 | 54/56 | 96.43 |
| 12 | 417/432 | 96.53 | 30 | 826/871 | 94.83 | 38 | 693/713 | 97.19 | 5 | 46/50 | 92.00 | |
| 10 | 34/42 | 80.95 | 9 | 42/59 | 71.19 | 22 | 150/154 | 97.40 | 4 | 44/46 | 95.65 | |
| 3 | 5/5 | 100 | 4 | 4/6 | 66.67 | 32 | 305/306 | 99.67 | 4 | 10/13 | 76.92 |
| Cultivar | No. Orchards Monitored | No. Trees with Reddening per Total No. Monitored | No. Infected Trees per Total No. Reddening Trees 1 | No. PCR-Positive Trees per No. Reddening Trees without Symptoms 2 |
|---|---|---|---|---|
| Axam | 2 | 74/121 (61.16%) | 23/74 (31.08%) | 4/5 (80.00%) |
| Berlepsch | 2 | 80/236 (33.90%) | 52/80 (65.00%) | 6/7 (85.71%) |
| Boskoop | 3 | 112/259 (43.24%) | 62/112 (56.25%) | 16/16 (100%) |
| Braeburn | 3 | 119/620 (19.19%) | 88/119 (73.95%) | 8/8 (100%) |
| Celest | 1 | 16/57 (28.07%) | 15/16 (93.75%) | 10/10 (100%) |
| Delbarestivale | 3 | 115/271 (42.43%) | 41/115 (35.65%) | 16/29 (55.17%) |
| Falstaff | 1 | 17/46 (36.96%) | 11/17 (64.71%) | 2/2 (100%) |
| Fuji | 2 | 13/130 (10.00%) | 10/13 (76.92%) | 2/2 (100%) |
| Gala | 6 | 230/824 (27.91%) | 174/230 (75.65%) | 25/28 (89.29%) |
| Golden Delicious | 5 | 489/1303 (37.53%) | 373/489 (76.28%) | 39/39 (100%) |
| Idared | 3 | 42/275 (15.27%) | 38/42 (90.48%) | 1/1 (100%) |
| Jonagold | 6 | 158/622 (25.40%) | 94/158 (59.49%) | 13/14 (92.86%) |
| Melrose | 2 | 22/212 (10.38%) | 15/22 (68.18%) | 8/8 (100%) |
| Pilot | 2 | 19/83 (22.89%) | 12/19 (63.16%) | 5/6 (83.33%) |
| Pink Lady | 2 | 21/76 (27.63%) | 18/21 (85.71%) | 5/5 (100%) |
| Pinova | 8 | 166/965 (17.20%) | 126/166 (75.90%) | 13/16 (81.25%) |
| Royal Gala | 2 | 98/171 (57.03%) | 92/98 (93.88%) | 10/11 (90.90%) |
| Rubinette | 5 | 152/602 (25.25%) | 107/152 (70.39%) | 34/40 (85.00%) |
| Rubinola | 2 | 54/350 (15.43%) | 27/54 (50.00%) | 13/19 (68.42%) |
| Topaz | 2 | 96/334 (28.74%) | 63/96 (65.63%) | 26/33 (78.79%) |
| Total | 62 | 2093/7557 (27.70%) | 1441/2093 (68.85%) | 256/299 (85.62%) |
| 2019 | 2020 | 2021 | 2022 | |
|---|---|---|---|---|
| Night | 18 September–21 September | 19 September–21 September | 21 September–25 September | 21 September–24 September |
| mean Tmin | 5.15 °C | 7.3 °C | 7.34 °C | 4.05 °C |
| Day | 19 September–23 September | 19 September–24 September | 22 September–27 September | 21 September–24 September |
| mean Tmax | 21.80 °C | 24.97 °C | 23.42 °C | 20.05 °C |
| Period of Induction | No. Plants | Temperature Regime (12 h Light/12 h dark) | Phytoplasma Titer (Phytoplasma/Plant Cell) | Reddening Index | Correlation |
|---|---|---|---|---|---|
| 11 August 2020–3 September 2020 | 2 | 20 °C/5 °C | 7.03 n.s. 1 | 0.76 4 | −0.24269 |
| 11 August 2020–3 September 2020 | 2 | 22 °C/15 °C | 5.30 n.s. | 0.10 | 0.15467 |
| 5 September 2019–20 September 2019 | 3 | 20 °C/5 °C | 39.91 n.s. 2 | 1.70 5 | 0.05243 |
| 5 September 2019–20 September 2019 | 3 | 22 °C/15 °C | 46.48 n.s. | 0.30 | 0.36145 |
| 21 September 2020–2 October 2020 | 3 | 20 °C/5 °C | 6.15 3 | 1.89 6 | 0.06469 |
| 21 September 2020–2 October 2020 | 4 | 22 °C/15 °C | 3.31 | 1.28 | −0.49252 |
| Origin of Samples | Correlation with Different Symptoms | |||||||
|---|---|---|---|---|---|---|---|---|
| ‘Ca. P. mali’ Subtype | No. Regions | No. Orchards | Total No. Analyzed | Witches’ Broom | Enlarged Stipules | Partial Reddening | Entire Reddening | Mean CDI 1 |
| AT-1 | 7 | 20 | 89 | 15/89 (16.85%) | 31/89 (34.83%) | 49/89 (55.06%) | 32/89 (35.96%) | 1.80 n.s. 2 |
| AT-2 | 5 | 13 | 33 | 1/33 (3.03%) | 17/33 (51.52%) | 22/33 (66.67%) | 7/33 (21.21%) | 1.67 n.s. |
| AP | 7 | 25 | 116 | 28/116 (24.14%) | 56/116 (48.28%) | 73/116 (62.93%) | 25/116 (21.55%) | 2.10 n.s. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jarausch, W.; Runne, M.; Schwind, N.; Jarausch, B.; Knauer, U. Phytoplasma-Induced Leaf Reddening as a Monitoring Symptom of Apple Proliferation Disease with Regard to the Development of Remote Sensing Strategies. Agronomy 2024, 14, 376. https://doi.org/10.3390/agronomy14020376
Jarausch W, Runne M, Schwind N, Jarausch B, Knauer U. Phytoplasma-Induced Leaf Reddening as a Monitoring Symptom of Apple Proliferation Disease with Regard to the Development of Remote Sensing Strategies. Agronomy. 2024; 14(2):376. https://doi.org/10.3390/agronomy14020376
Chicago/Turabian StyleJarausch, Wolfgang, Miriam Runne, Nora Schwind, Barbara Jarausch, and Uwe Knauer. 2024. "Phytoplasma-Induced Leaf Reddening as a Monitoring Symptom of Apple Proliferation Disease with Regard to the Development of Remote Sensing Strategies" Agronomy 14, no. 2: 376. https://doi.org/10.3390/agronomy14020376
APA StyleJarausch, W., Runne, M., Schwind, N., Jarausch, B., & Knauer, U. (2024). Phytoplasma-Induced Leaf Reddening as a Monitoring Symptom of Apple Proliferation Disease with Regard to the Development of Remote Sensing Strategies. Agronomy, 14(2), 376. https://doi.org/10.3390/agronomy14020376