Abstract
The Golden2-like (GLK) transcription factors belong to the GARP family of transcription factors and play significant roles in plant growth, development, and responses to both abiotic and biotic stresses. This study employed bioinformatics and expression analyses to investigate the regulatory roles of wheat GLK proteins under various stress conditions, including abscisic acid (ABA) treatment, osmotic stress, and infection by Fusarium graminearum. The study identified 125 TaGLK proteins and revealed that TaGLKs play a significant role in wheat’s development and response to adverse environmental conditions. The results indicate that TaGLKs may serve as potential transcriptional regulators capable of integrating multiple cellular signals to coordinate various developmental and physiological processes. Evolutionary analysis classified the TaGLK proteins into six subgroups, which shared similar conserved domains and motifs. Protein–protein interaction network analysis revealed that TaGLKs are involved in photoreceptor activity, cell cycle progression, and protein regulation. Gene expression analysis of TaGLKs discovered that they play key functions in wheat development, as well as regulation of biotic and abiotic stress conditions. RT-qPCR analysis showed that TaGLKs regulate earlier and late effects of osmotic stress, F. graminearum infections, and ABA treatment in wheat. These findings provide knowledge for future studies of the functions of TaGLK TFs in wheat stress tolerance and development, which could have significant implications for enhancing wheat tolerance to various environmental stressors.
1. Introduction
Plants have evolved complex strategies to cope with various environmental challenges that affect their growth and survival [1]. Transcription factors (TFs) are an essential part of these strategies because they play a major role in controlling stress-related behaviors and developmental processes. They act as vital links in signal transduction, control gene expression, and impact multiple facets of plant growth [2,3]. Throughout their lifespan, plants face a variety of abiotic challenges, such as extreme temperatures, exposure to herbicides, heavy metal contamination, drought, saline conditions, cold weather, and attacks from pests and diseases [4]. In response to these challenges, TFs modulate transcription by binding to DNA promoters via their DNA-binding domains (DBDs) [5,6]. Stress conditions can modify the characteristics of TFs, influencing their location within the cell, as well as their stability, functionality, and interactions with other proteins, which in turn affect the expression of target genes. Among the various families of transcription factors, the MYB-like group is particularly important, as they play a significant function in the plant’s response to a broad spectrum of stressors [7].
The Golden2-like (GLK) TFs belong to the MYB-related GARP family of regulatory proteins in plants. A GCT box located at the C-terminus of these proteins and a MYB-type DNA-binding domain (DBD) are their two defining characteristics [8,9]. GLK family members are important in plants for various defense mechanisms and stress responses to abiotic and biotic stressors [10,11]. In the model organism Arabidopsis thaliana, the closely related GLK TFs, AtGLK1 and AtGLK2, work together to oversee chloroplast development and photosynthetic processes [12]. Overexpressing AtGLK1 boosts resistance to F. graminearum but makes the plant more vulnerable to Hyaloperonospora arabidopsidis (Hpa). Conversely, the Arabidopsis glk1 glk2 double mutant shows increased resistance to Hpa compared to wild-type plants [13]. Beyond their roles in chloroplast function and pathogen resistance, AtGLKs also influence the expression of Cucumber mosaic virus resistance genes in Arabidopsis. The OsGLK1 has been linked to resistance against certain pathogens [14]. Genetic analyses indicate that Arabidopsis GLKs may act as negative regulators of immunity to the bacterial pathogen Pseudomonas syringae [15]. Furthermore, GLKs have been implicated in enhancing resistance to viral infections, such as Cucumber mosaic virus, by affecting genes related to antioxidant and defense expression levels, as well as key phytohormones like jasmonic acid and salicylic acid [10].
GLK TFs in Arabidopsis are associated with various responses to abiotic stress. A recent study revealed that enhanced tolerance to the air pollutant ozone was linked to a closed-stomata phenotype, resulting from the suppression of AtGLK1 or AtGLK2 using the SRDX transcriptional repression domain [16]. In contrast, overexpressing either AtGLK1 or AtGLK2 led to an open-stomata phenotype, increasing sensitivity to ozone. Research has also shown that GLK TFs can negatively regulate tolerance to salt and osmotic stress in both Arabidopsis and Salicornia europaea [17,18]. Conversely, GLK1 has been found to enhance drought recovery in both cotton and Arabidopsis [19,20]. The Arabidopsis glk1 glk2 double mutant exhibited improved survival after drought stress and rewatering, highlighting the role of GLKs in ABA-mediated signaling pathways that control stress-related gene expression [21]. In Brassica napus, GLKs are regulated by C-Repeat Binding Factor (CBF) TFs, which are crucial for cold acclimation. Additionally, the cotton GLK1 has been shown to enhance cold tolerance when expressed in Arabidopsis [20,22]. Moreover, short-term ABA treatment has been demonstrated to upregulate the expression of the Arabidopsis GLK1, while the light signaling component CONSTITUTIVE PHOTOMORPHOGENIC 1 (COP1) increases in response to ABA and light, facilitating the degradation of GLK1 [18,23].
Wheat (Triticum aestivum L.) is an important cereal crop that supports a significant portion of the global population. As the most widely cultivated cereal grain in the world, it plays a vital role in agriculture and provides food for 36% of people, being grown in 70% of the world’s arable land [24,25]. However, wheat is vulnerable to various biotic and abiotic stresses, necessitating the establishment of genetically controlled defense mechanisms. Research has indicated that TFs are important in regulating wheat’s responses to different stress conditions, highlighting their potential roles in enhancing resistance to abiotic stresses and diseases in this crop [26]. Previous studies have reported that TFs such as WRKY, NAC, MYB, and GRF are associated with various processes, including seed development, leaf senescence, plant proliferation, expansion, and responses to biotic and abiotic stresses [27]. However, the regulatory function of GLK TFs in plant stress responses remains poorly understood. Although some research has indicated that GLKs play a role in regulating stress-responsive pathways in specific plant species, the precise mechanisms through which they contribute to stress tolerance in wheat are still unclear. This study investigated the regulatory roles of TaGLK TFs under various stresses, including osmotic stress, ABA treatment, and F. graminearumi infection, using bioinformatics, gene expression data, and RT-qPCR analyses to better understand the potential regulatory role of TaGLKs in wheat during abiotic and biotic stress conditions. The findings offer essential insights for further exploration of the roles of TaGLKs and their possible application in the molecular breeding of stress-tolerant wheat cultivars.
2. Materials and Methods
2.1. Identification of the GLK Gene Family in Wheat
G2-like protein sequences from tobacco (LOC107817975), Arabidopsis (AT2G20570.2 and AT5G44190.1), maize (LOC542493), and rice (LOC4326363) were retrieved from the NCBI (https://www.ncbi.nlm.nih.gov/, accessed on 29 November 2023) database and utilized to search for G2-like sequences against the Triticum aestivum cv. Chinese Spring v2.1 wheat genome in the Phytozome v13 (https://phytozome-next.jgi.doe.gov/ (accessed on 29 November 2023)) database using the default parameters. The retrieved TaGLK proteins were subsequently refined using SMART (http://smart.embl.de/ (accessed on 11 December 2023)), along with the NCBI conserved domains CDDv3.21-62456 PSSMs (https://www.ncbi.nlm.nih.gov/cdd/, accessed on 11 December 2023) using an e-value cutoff of 1 × 10−5, and InterProScan (www.ebi.ac.uk/interpro/about/Interproscan/, accessed on 11 December 2023) databases to eliminate unnecessary, unmatched, and incomplete TaGLK proteins.
2.2. Physicochemical Properties and Chromosome Mapping Analyses of TaGLKs
The physicochemical characteristics of the TaGLK proteins, such as number of amino acids, isoelectric point, molecular weight, grand average of hydropathicity, and instability index, were assessed using the ExPASy-ProtParam program (https://www.expasy.org/resources/protparam (accessed on 5 January 2024)). Using the wheat genome GFF3 file obtained from the Phytozome v13 database, the distribution of the TaGLKs across the wheat chromosomes was visualized using MapInspect v1.0. Subcellular location of the TaGLK proteins was analyzed using the Plant-mPLoc software v2.0 (Plant-mPLoc server (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/, accessed on 21 January 2024)) [28]. The SignalP 6.0 program (SignalP 6.0-DTU Health Tech-Bioinformatic Services/ (https://services.healthtech.dtu.dk/services/SignalP-6.0/, accessed on 8 January 2024)) was used to analyze signal peptides of the TaGLK proteins employing the slow model mode with default settings [29]. The TMHMM-2.0 software (TMHMM 2.0-DTU Health Tech-Bioinformatic Services (https://services.healthtech.dtu.dk/services/TMHMM-2.0/, accessed on 8 January 2024)) was employed to analyze the transmembrane helices in the TaGLK proteins.
2.3. Phylogenetic Analysis of TaGLK Proteins
The protein sequences of the GLKs of Oryza sativa and Arabidopsis thaliana were sourced from the Phytozome v13 database. An evolutionary analysis of 125 TaGLKs, 46 rice, and 10 Arabidopsis GLK proteins was conducted using ClustalW in the MEGA 11 program with default parameters [30]. The Neighbor-Joining Method (NJ), pairwise deletion, Poisson model, uniform rates, and a bootstrap value of 1000 repetitions were used to build the phylogenetic tree. The iTOL v6 software (https://itol.embl.de/, accessed on 24 January 2024) [31] was utilized to visualize and annotate the evolutionary tree of the TaGLK, OsGLK, and AtGLK proteins.
2.4. Gene Structure, Cis-Acting Elements, and Conserved Motif Analyses of TaGLKs
The gene structures of the TaGLKs were analyzed using the wheat genome GFF3 file with GSDS v2.0 software (http://gsds.cbi.pku.edu.cn/, accessed on 10 January 2024) [32]. The promoter sequence, extending 1 kb upstream of the TaGLKs, was retrieved using the Ensembl Plants database (http://plants.ensembl.org/, accessed on 29 January 2024). The PlantCARE database was used to analyze the TaGLK promoter sequences [33]. To predict the conserved structural domains of the TaGLK proteins, the MEME Suite v5.5.5 online software (https://meme-suite.org/meme/tools/meme, accessed on 11 January 2024) was used. The motif number was set to 20 and the site distribution to One Occurrence Per Sequence (oops) while keeping all other parameters at their default settings [34]. The conserved motifs of the TaGLKs were displayed using TBtools software v2.119 [28].
2.5. Gene Ontology (GO) Annotation and Protein–Protein Interaction of TaGLKs
All TaGLK protein sequences were subjected to Gene Ontology (GO) annotation using the OmicsBox/Blast2GO software (https://www.blast2go.com/, accessed on 17 February 2024) with default parameters. The TaGLKs were categorized into three main categories: Biological Process (BP), Molecular Function (MF), and Cellular Component (CC), and the number of transcripts annotated to each category was counted. GO functional enrichment analysis of the TaGLK TFs was conducted using the DAVID Bioinformatics Functional Annotation Tools (https://davidbioinformatics.nih.gov/, accessed on 17 February 2024) [35], which further categorized the enriched GO terms into BP, CC, and MF. All TaGLK protein sequences were submitted to the STRING v12.0 database (http://string-db.org), selecting Triticum aestivum as the reference organism. Following a BLAST analysis, orthologous STRING proteins with 100% identity were utilized to construct the TaGLKs protein interaction (PPI) network, while proteins without interactions were excluded [36]. The PPI network was then visualized and modified using Cytoscape 3.10.2 software [37].
2.6. Expression Data of Wheat TaGLKs Under Abiotic, Biotic, and Developmental Conditions
Gene expression data for the TaGLKs in wheat under various stress conditions, including biotic stresses (stripe rust, powdery mildew, and Fusarium head blight), abiotic stresses (PEG6000, salt stress, drought/heat), and developmental stages (early spike and wheat grain), were obtained from the WheatOmics 1.0 database (Wheat Gene Expression Database - sdau.edu.cn (http://wheatomics.sdau.edu.cn/expression/wheat.html, accessed on 26 February 2024)) [38]. TaGLKs with TPM values greater than 1 in at least one treatment (biotic, abiotic, and developmental stages) condition were selected for further analysis. The expression of the TaGLK TFs was estimated as log2TPM values. A gene expression heatmap for the expressed TaGLKs based on the estimated log2TPM values was generated using the Omicstudio program (https://www.omicstudio.cn, accessed on 1 March 2024) [39]. Based on the expression profiles, 8 expressed TaGLKs (TaGLK4, TaGLK28, TaGLK32, TaGLK36, TaGLK55, TaGLK59, TaGLK98, and TaGLK117) in biotic and abiotic stress conditions were chosen for RT-qPCR gene expression study under abiotic and biotic stress treatments.
2.7. Growth and Stress Treatments of Wheat Seedlings
The wheat cultivar Jingshuang 16, which is moderately susceptible to powdery mildew and stripe rust, was obtained from the College of Agriculture, Yangtze University, Hubei, China, and used as the experimental material. After sterilizing the seeds with a 1% hypochlorous acid solution, they were carefully washed three times using distilled water. For 3 days, the sterilized seeds were grown in a temperature-controlled growth room with an average humidity of 60% on wet filter paper in a culture dish [40]. A day/night cycle of 25 °C/27 °C and a light/dark cycle of 16 h were used as the growing conditions. After being moved to a hydroponic germination dish, the sprouting seedlings were given a half-strength Hoagland nutrient solution [41]. At the two-leaf stage, the seedlings were subjected to osmotic stress by treatment with 20% PEG6000 and hormone treatment with 100 μM abscisic acid (ABA). Following the treatments, three biological replicates of leaf samples were taken at 0, 2, 6, 12, 24, and 48 h. The collected samples were immediately frozen in liquid nitrogen and kept at −80 °C. Wild-type F. graminearum strain PH-1 obtained from the College of Agriculture, Yangtze University, Hubei, China, was cultured on potato dextrose agar (PDA). Preparation of fungal inoculum was performed based on the previously described protocol [42]. At 16 °C, pot soil was used to germinate wheat seeds in a temperature-controlled growth chamber with a 16-h light/eight-h dark cycle. At the two-leaf stage, the leaves were inoculated with the wild-type PH-1 strain of F. graminearum and kept in moist, dark conditions for 24 h to promote infection. Following inoculation, the leaf samples were taken 0 days, 1 day, 3 days, and 5 days later, with three biological replicates. The samples were promptly frozen at −80 °C and kept for later examination.
2.8. RT-qPCR Expression Analysis of TaGLKs
The TRIzol reagent (Invitrogen) was used to extract total RNA from the wheat leaves in accordance with the manufacturer’s instructions. Electrophoresis on a 1.5% agarose gel was conducted to assess the extracted RNA’s integrity, and a NanoDrop 2000 spectrophotometer was used to measure the RNA’s concentration and purity. cDNA was synthesized using the HiScript Reverse Transcriptase (Vazyme, Nanjing, China) in accordance with the manufacturer’s instructions. Quantitative real-time PCR (RT-qPCR) was performed using the ChamQ SYBR qPCR Master Mix (Vazyme Biotech Co., Ltd., Nanjing, China) in a 20 μL reaction volume. Gene-specific primers for TaGLKs used for RT-qPCR analysis (Table S1) were designed using the Primer3Plus software, and the ADP-ribosylation factor Ta2291 was used as an internal reference gene. The thermal cycling profile consisted of an initial denaturation step at 95 °C for 3 min, followed by 40 cycles of denaturation at 95 °C for 10 s and annealing/extension at 60 °C for 30 s. Lastly, a temperature range of 95 °C for 15 s, 60 °C for 1 min, and 95 °C for 15 s was used to conduct a melt curve analysis. For every sample, three biological replicates were analyzed, and the 2−ΔΔCt method was utilized to determine the relative expression of the TaGLKs [43]. GraphPad Prism was used to conduct statistical analysis to assess the significant differences between the treatment samples at different time points at p < 0.05.
3. Results
3.1. Identification and Physicochemical Properties of TaGLKs
A comprehensive bioinformatics analysis was performed to identify and characterize the TaGLK TFs using the Phytozome v13 database. A total of 253 TaGLKs were identified from the initial search and screening of the proteins using the SMART, NCBI-CDD, and InterProScan databases, leading to the identification of 125 TaGLKs. Subsequently, the basic physicochemical properties of the TaGLK proteins were then predicted and analyzed. The amino acid sequence lengths of the TaGLK proteins ranged from 225 to 534 residues. The molecular weights of the TaGLK proteins ranged from 24,233.07 to 58,472.61, suggesting that TaGLKs may have different functions in wheat, enabling them to participate in various biological processes. The isoelectric points were in the range of 4.84 to 9.92. The instability index, which provides an estimate of the stability of the proteins, ranged from 28 to 75.74 for the TaGLKs. Seven proteins (TaGLK13, TaGLK19, TaGLK24, TaGLK78, TaGLK86, TaGLK88, and TaGLK94) exhibited instability indices below 40, suggesting that these proteins are likely to be stable enough to persist in cells and maintain their functions over time. This stability is important for effective regulation of biological processes. The grand average of hydropathicity (GRAVY) values, which indicate the overall hydrophobicity of the proteins, ranged from −0.945 to −0.084 for the TaGLK proteins. Subcellular localization analysis revealed the diverse intracellular distribution of the TaGLK proteins. Three TaGLKs (TaGLK55, TaGLK63, and TaGLK70) were found to be localized in the chloroplast. Additionally, three TaGLKs (TaGLK11, TaGLK44, and TaGLK111) were localized in both the chloroplast and the nucleus. TaGLK23 was found in the cell membrane, chloroplast, and nucleus. The majority of the TaGLKs (118 out of 125) were localized solely in the nucleus (Table S2), suggesting that they may function primarily as transcriptional regulators, influencing gene expression in response to various signals. The localization of TaGLKs in different cellular compartments suggests that these TFs not only function in transcriptional regulation but may also be potentially involved in metabolic regulation.
3.2. Evolutionary Relationships, Chromosome Mapping, and Domains of TaGLK Proteins
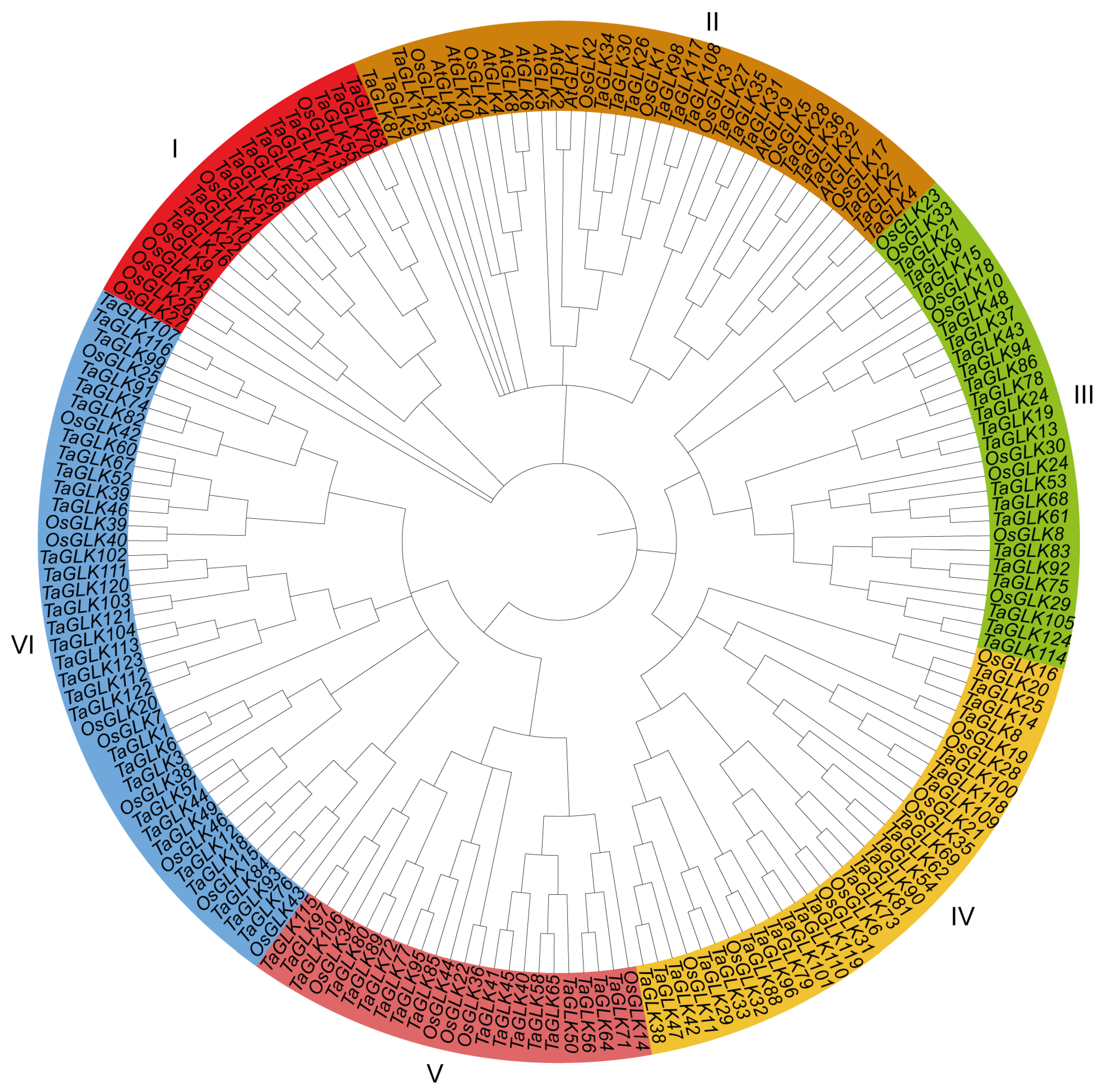
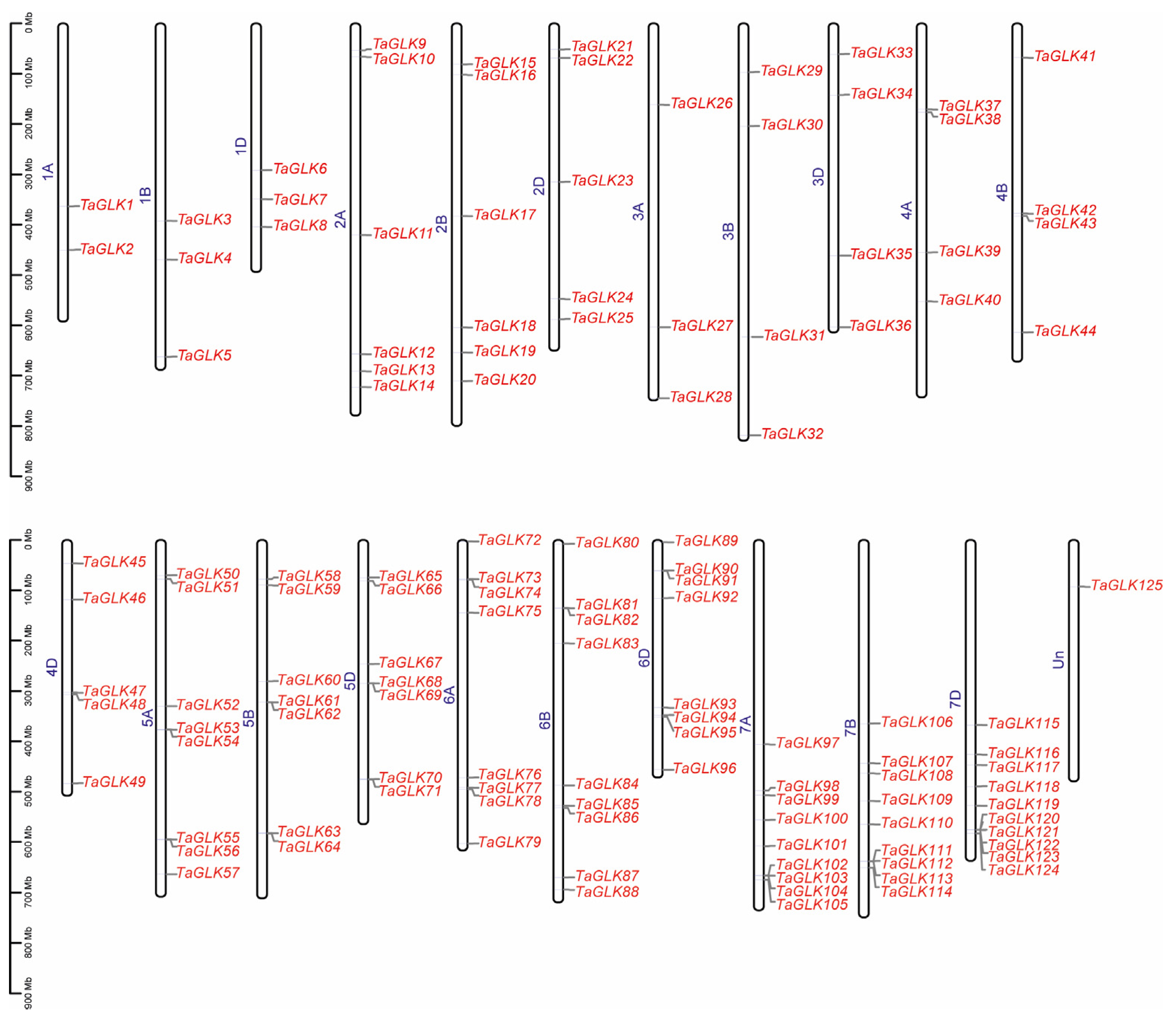
To investigate the evolutionary relationships within the TaGLK family, a phylogenetic analysis was conducted, incorporating the 125 TaGLK proteins, 46 OsGLKs, and 10 AtGLKs, including the two Arabidopsis query proteins AT2G20570.1 (TaGLK1) and AT5G44190.1 (TaGLK2), involved in the positive regulation of DNA-templated transcription. The analysis categorized the TaGLKs, AtGLKs, and OsGLKs into six distinct subgroups, labeled I through VI, indicating potential functional conservation among GLK proteins across different species. This suggests that GLK proteins have diversified in response to varying environmental stresses and functional demands across species. The distribution of TaGLK proteins in each subgroup was as follows: 12 in subgroup I, 18 in II, 21 in III, 24 in IV, 18 in V, and 32 in VI. The OsGLK proteins were distributed across the subgroups as 7, 7, 8, 9, 5, and 10, respectively. The Arabidopsis GLK proteins were categorized into subgroup II. A distinct clustering pattern was observed in the phylogeny. For instance, all 10 AtGLK proteins were classified into subgroup II. Additionally, TaGLK8 and OsGLK19 (Os05g0491500), which are associated with DNA-binding transcription factor activity, were clustered together (Figure 1). This suggests that certain members of the GLK protein family from different species may have similar functions in critical biological processes such as stress response, development, and metabolic regulations. The 125 TaGLKs were discovered to be sporadically dispersed across the 22 wheat genome chromosomes, indicating potential historical gene duplications and chromosomal rearrangements that may contribute to the functional diversity and regulatory mechanisms of TaGLKs in wheat’s response to various stress conditions. The number of TaGLKs per chromosome ranged from 1 to 10, with chromosome 7D containing the most (10 TaGLKs) and chromosomes 7A and 7B having nine TaGLKs each. One gene (TaGLK125) was mapped to the chromosome Un, highlighting the complex nature of the wheat genome and the need for further genomic investigation. The GO term for TaGLK125 was related to the regulation of DNA-templated transcription (GO:0006355), suggesting its potential role in modulating signaling pathways to facilitate the timely expression of stress-responsive genes, which is essential for wheat survival under stress conditions. The TaGLKs were renamed based on the common wheat abbreviation (“Ta”), followed by “GLK” and their physical positions (from lowest to highest) on the chromosomes, ranging from 1A to Un (Figure 2). Conserved domain analysis found that most TaGLK proteins have the Myb-like DNA-binding and Myb-CC-LHEQLE domains, indicating the potential of the TaGLKs’ involvement in regulating gene expression in wheat response to stress. Furthermore, the phylogenetic analysis showed that all six subgroups of the TaGLKs shared the Myb-like DNA-binding and SANT domains, suggesting a conserved evolutionary function related to transcriptional regulation. Interestingly, members of subgroup II, TaGLK5 and TaGLK125, were found to contain the REC domain, while TaGLK87 contained the AtoC domains (Supplementary Figure S1), indicating potential involvement in specific signaling pathways and functional specialization within the TaGLK family. These results suggest that TaGLK TFs may integrate multiple signals to coordinate complex responses to various stresses in wheat.
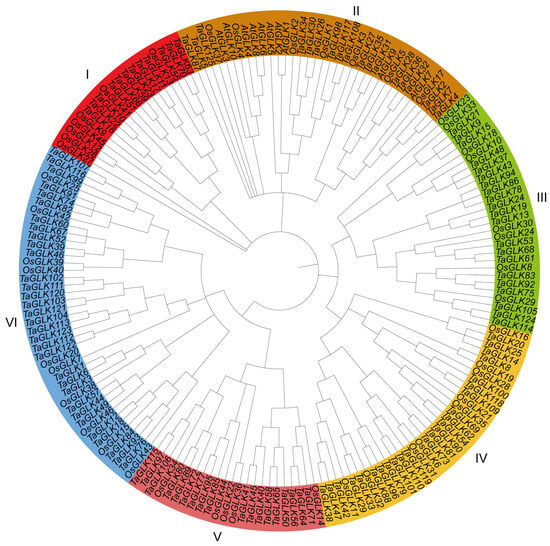
Figure 1.
Phylogenetic tree of GLK proteins in wheat and other species. The nodes were tested through bootstrap analysis with 1000 replicates, the Poisson model, uniform rates, and pairwise deletion. The phylogenetic relationships of the TaGLKs, AtGLKs, and OsGLKs were classified into 6 subgroups (I, II, III, IV, V, and VI).
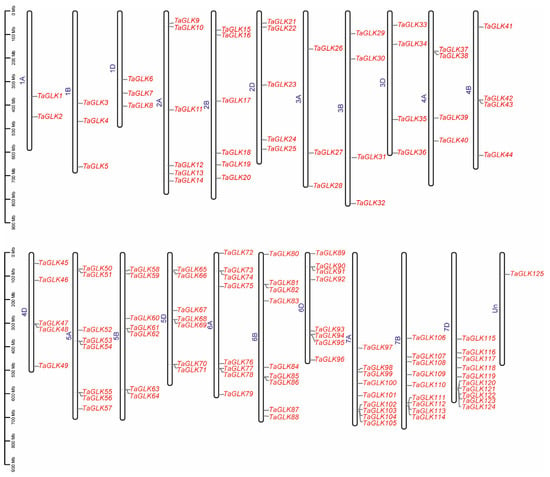
Figure 2.
Distribution of TaGLKs on chromosomes. The names of each chromosome (1A to Un) are shown in color blue at the left position. The TaGLK names are indicated at the right position of each chromosome with color red. The chromosome lengths are shown in Mb (millions of bases).
3.3. Conserved Motifs, Gene Structures, and Cis-Acting Elements of TaGLKs
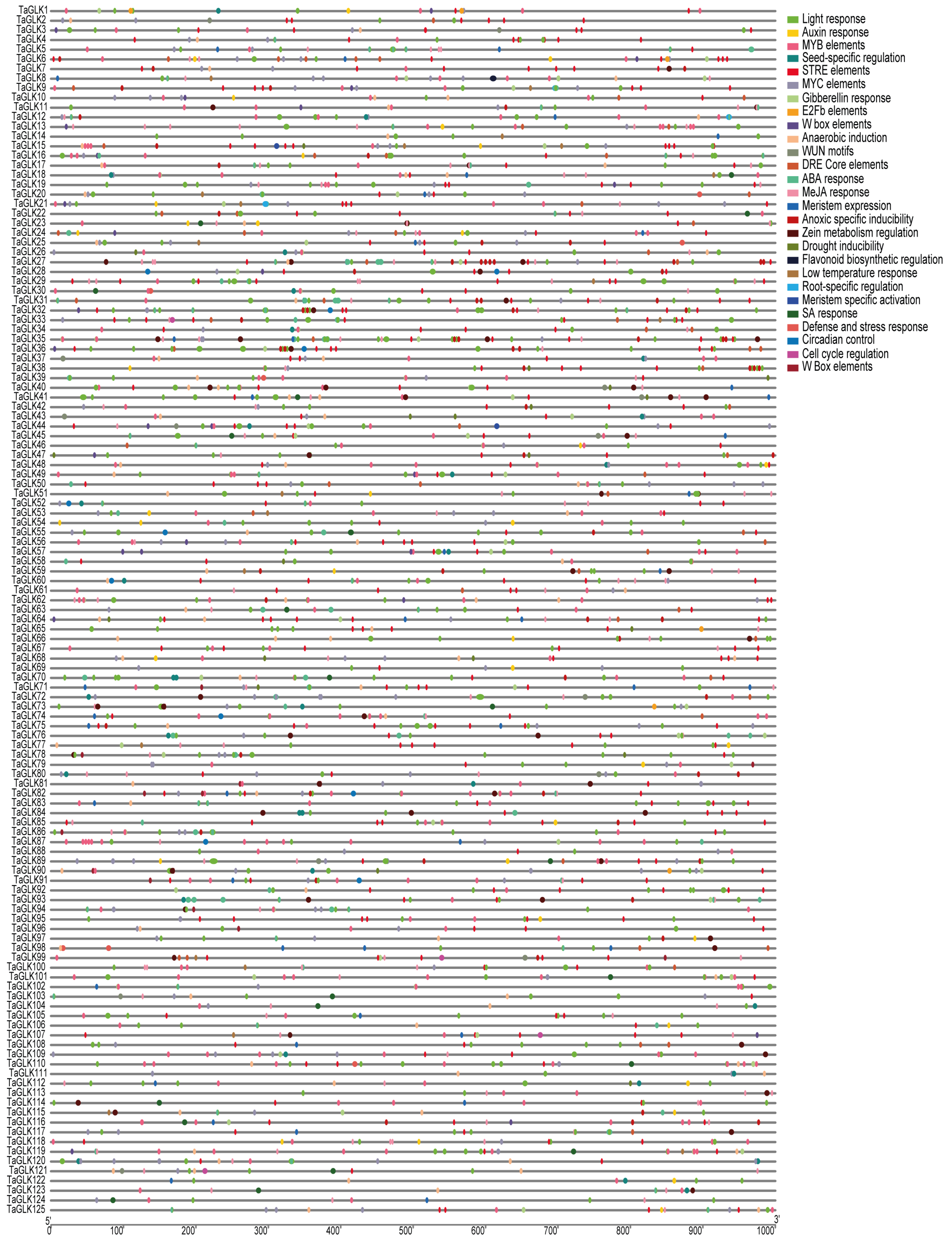
To investigate the structural characteristics of the TaGLKs, the MEME Suite was used to identify conserved motifs within the TaGLK proteins. The analysis revealed a total of 20 putative motifs, ranging in length from 11 to 50 amino acids, with each motif present only once (Figure 3A), highlighting significant structural diversity among TaGLK proteins. The TaGLK proteins contained 2 to 10 different motifs, with the members of subgroup II having five motifs each. This motif composition was highly conserved among TaGLK proteins within the same phylogenetic subgroup, suggesting potential similarities in regulatory functions and biological processes across different subgroups. To explore the evolution of the TaGLKs, the intron–exon structures were analyzed. The results indicated variability in the number of exons, UTRs, and introns among the TaGLK. Specifically, the exon numbers were 4 to 8 in total, with most TaGLKs containing 5 to 7 exons, indicating an evolutionary difference among the TaGLKs. This variability among the TaGLKs may potentially influence gene expression and functional specialization, suggesting that different TaGLK proteins may perform specific functions in wheat developmental processes. Fourteen members, including TaGLK5, lacked any UTRs, while four members, including TaGLK3, contained only 5’-terminal UTRs, and three members, including TaGLK51, had only 3’-terminal UTRs, implying potential differences in regulatory mechanisms. These structural differences may suggest potential functional divergence within the TaGLK protein family (Figure 3B). Cis-acting regulatory element analysis of the TaGLKs promoters identified several stress-responsive elements related to biotic and abiotic stress responses, such as low-temperature response, ABA response, anoxic-specific inducibility, MeJA response, anaerobic induction, gibberellin response, MYB elements, DRE core elements, auxin response, drought inducibility, and salicylic acid (SA) response. Additionally, various elements associated with plant development and growth and light-responsive elements were found, including meristem-specific activation, zein metabolism regulation, seed-specific regulation, cell cycle control, and W box elements (Figure 4). This suggests that TaGLK proteins may potentially play significant functions in integrating diverse signaling pathways related to developmental processes in wheat.

Figure 3.
Conserved motifs and gene structure analysis of TaGLKs. (A) Conserved motifs of TaGLKs. (B) Structural dynamics of TaGLKs: the color yellow represents CDSs, the color green indicates UTRs, and black lines represent introns.
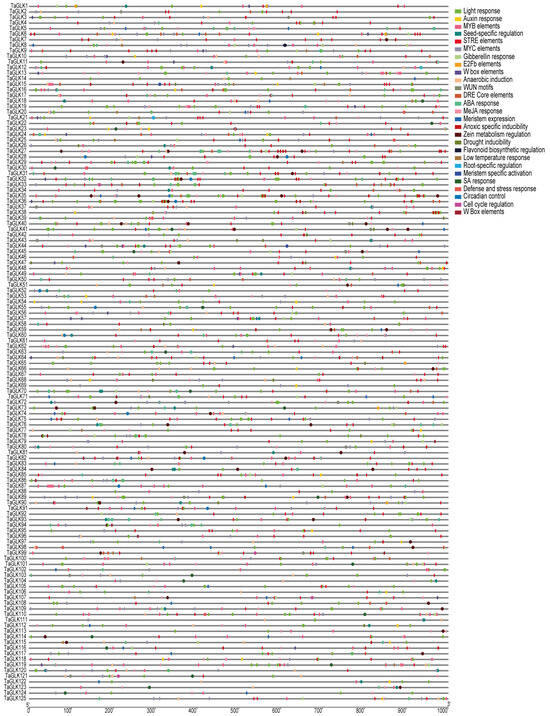
Figure 4.
Cis-acting element analyses of a 1 kb promoter region of the TaGLKs.
3.4. GO Functional Annotation and Protein–Protein Interaction of TaGLKs
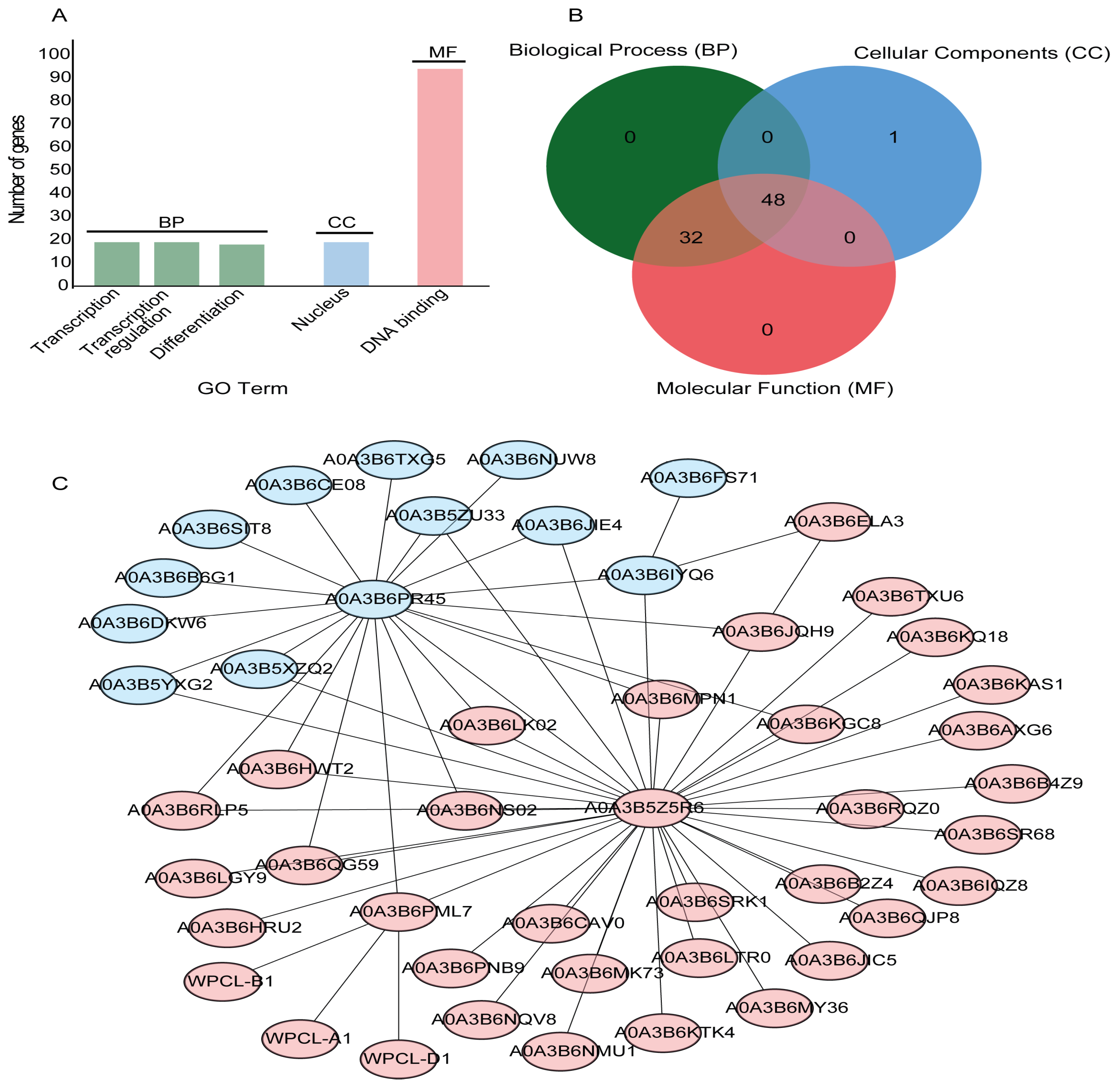
To explore the regulatory roles of the TaGLKs, Gene Ontology (GO) term functional and annotation enrichment analyses were performed using the DAVID Bioinformatics and OmicsBox/Blast2Go databases. The analyses revealed that 19 TaGLKs were enriched in transcription, 19 in transcription regulation, and 18 in differentiation within the Biological Process (BP) category. For Cellular Component (CC), 19 TaGLKs were found to be enriched in the nucleus, while 94 were enriched in DNA binding within the Molecular Function (MF) category (Figure 5A). The GO annotation enrichment analysis identified a total of 81 TaGLKs with known annotations. Most of these TaGLKs exhibited at least two annotation functions, with the exception of TaGLK87, which was only associated with the nucleus. These results indicate significant multifunctionality among the majority of TaGLK TFs, highlighting their involvement in various cellular processes essential for wheat stress regulation. Notably, 80 annotated TaGLKs were related to at least two or more GO terms, with 48 TaGLKs associated with all three GO categories. Additionally, 32 TaGLKs were linked to both MF and BP, while only one TaGLK was related exclusively to the CC category (Figure 5B). Our findings suggest that TaGLKs have diverse roles in cellular activities and are significantly involved in regulating various metabolic and developmental processes. The most highly annotated functional category was BP, followed by MF and CC. Within the BP category, several GO terms related to plant development and stress regulation were identified. For instance, 80 TaGLKs were associated with the positive regulation of DNA-templated transcription (GO:0003700), 12 were linked to the regulation of the chlorophyll biosynthetic process (GO:1900056), and others were involved in the ethylene-activated signaling pathway (GO:0009736) and the regulation of stomatal movement (GO:2000280). Furthermore, 18 TaGLKs were associated with the cytokinin-activated signaling pathway (GO:0010150) and the regulation of root meristem growth (GO:0009873). Six TaGLKs were linked to the response to water deprivation (GO:0090506), while 33 TaGLKs were associated with positive regulation of gene expression (GO:0010089) and 18 TaGLKs were related to the regulation of seed development (GO:0003700) (Table S3). These findings indicate that TaGLKs play significant roles in wheat development processes, particularly through their involvement in various biological functions, including photosynthesis, transpiration, and regulation of seed development. In terms of MF, 80 TaGLKs were enriched in terms related to DNA-binding transcription factor activity (GO:0006355), 18 were associated with phosphorelay response regulator activity (GO:0010628) and heterocyclic compound binding (GO:0003700), and nine TaGLKs were linked to transcription cis-regulatory region binding (GO:0000976). This suggests that TaGLKs are involved in signal transduction pathways and modulate the transcription of stress-responsive genes, which are essential for wheat’s response to stress signals. Additionally, 52 TaGLKs were associated with the nucleus (GO:0005634) and nine were related to the nucleus in the enriched CC GO terms, suggesting the important role of TaGLKs in nuclear functions and gene expression regulation.
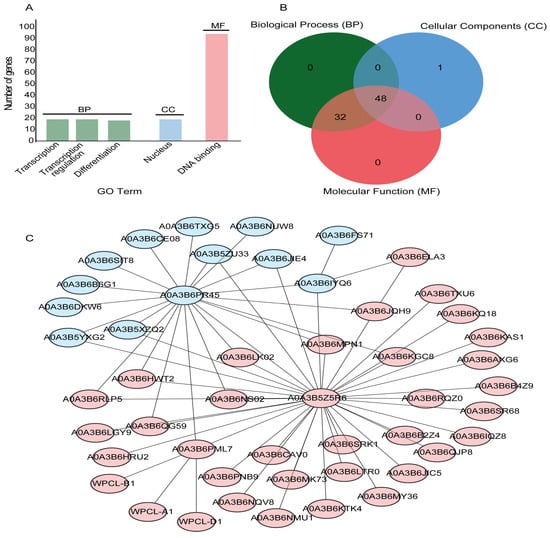
Figure 5.
GO annotation and protein interaction network of TaGLKs. (A) GO-term functional enrichment terms of TaGLKs. (B) GO annotation functions of TaGLK proteins. (C) Protein–protein interaction network of TaGLK proteins: Blue-colored TaGLKs are related to the Myb-like DNA-binding domain and CCT motif; red-colored TaGLKs are related to the detection of light stimulus and photoreceptor activity.
To investigate the coordinated functions of the TaGLKs in response to plant stress, the STRING database was used to conduct a protein interaction analysis. We aligned the protein sequences of the TaGLKs against those from Triticum aestivum and selected orthologous proteins with 100% identity to construct a protein–protein interaction (PPI) network for the TaGLKs and removed those without any interaction. A total of 49 proteins were identified as part of the main network, as shown in the STRING database for Triticum aestivum (Figure 5C). The HTH myb-type domain-containing protein A0A3B5Z5R6 (TaGLK5) and an uncharacterized protein A0A3B6PR45 (TaGLK87) were highly clustered within this network. The pathways significantly enriched in the TaGLKs protein network included cutin, suberine, and wax biosynthesis (map00073), indicating the important role of TaGLKs in forming protective barriers in plants. This function is significant for enhancing plant resistance against environmental stresses, such as water loss and pathogen invasion. GO enrichment analysis of the 49 proteins in the TaGLKs network revealed significant enrichment in terms related to integument development (GO:0080060), ubiquitin ligase activator activity (GO:1990757), and the anaphase-promoting complex (GO:0005680). These findings highlight the roles of TaGLKs in plant development, protein regulation, and cell cycle progression during stress responses. To explore the interconnected functions of these proteins, we employed k-means clustering to group the 49 proteins into two clusters based on their shared functions. The results indicated that 14 proteins, colored blue, were involved in the detection of light stimuli and photoreceptor activity, with GO-term functions related to rhythmic processes (GO:0048511), DNA-binding transcription factor activity (GO:0003700), and cellular anatomical entities (GO:0110165) (Figure 5C). This suggests the importance of these TaGLKs in light-mediated plant responses. Furthermore, the remaining 35 proteins, colored orange, were associated with the Myb-like DNA-binding domain and the CCT motif, exhibiting GO-term functions related to integument development (GO:0080060), ubiquitin ligase activator activity (GO:1990757), and the anaphase-promoting complex (GO:0005680). This indicates their involvement in developmental processes and protein regulation during stress responses in wheat.
3.5. Analysis of TaGLKs’ Expression Under Wheat Abiotic, Biotic, and Developmental Conditions
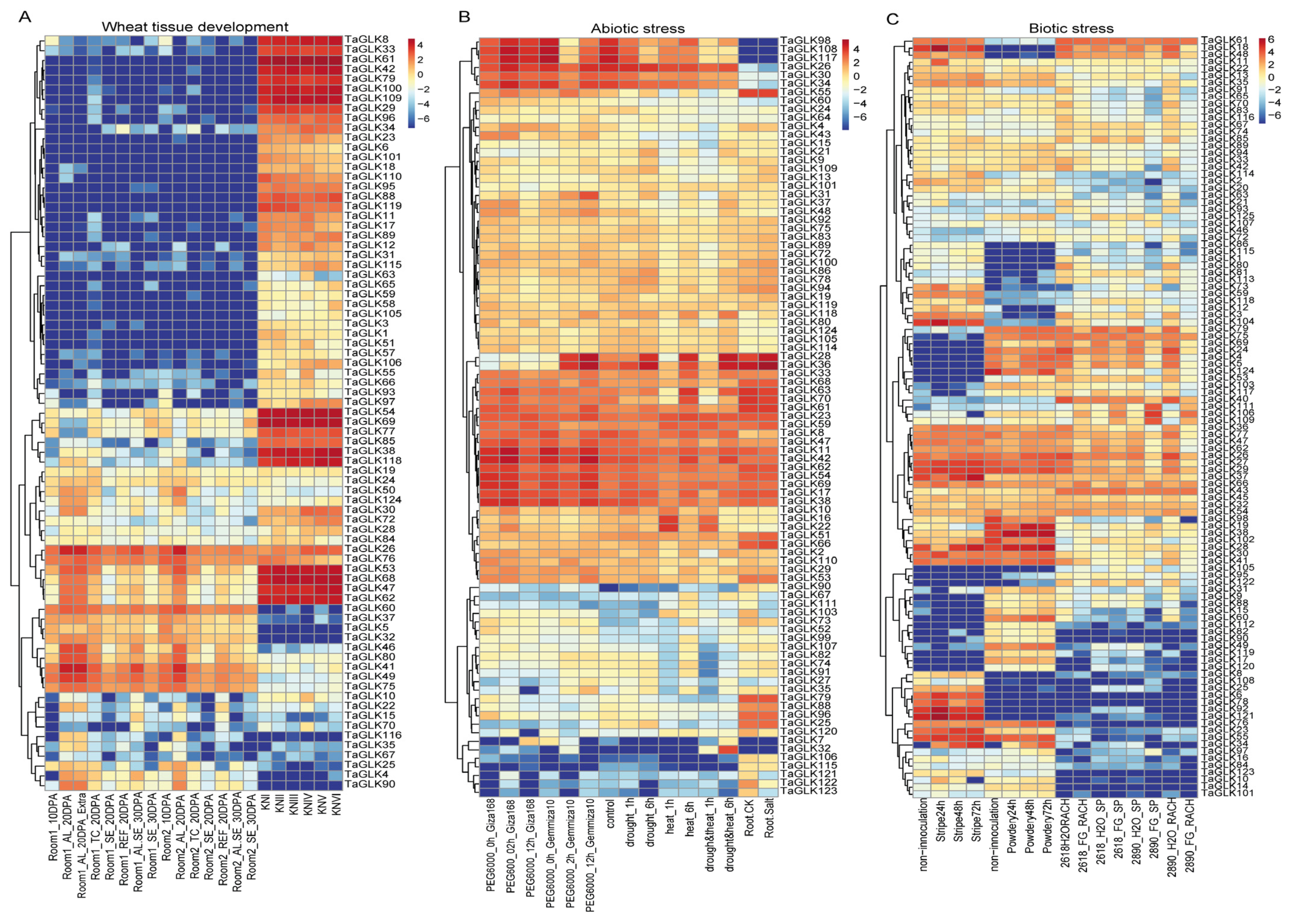
To investigate the functional roles of the TaGLKs, we retrieved their expression patterns during tissue development and under abiotic and biotic stresses from the WheatOmics 1.0 database. The expression analysis revealed varied differences in the levels of certain TaGLKs across various wheat tissues (Figure 6A). Specifically, TaGLK62, TaGLK29, TaGLK47, TaGLK54, and TaGLK69, all belonging to the TaGLK subgroup IV, were highly expressed during early wheat spike development. Additionally, TaGLK26, TaGLK60, and TaGLK76 showed high expression levels in developing wheat grains. This suggests that TaGLKs may play important roles in regulating developmental processes, which could be important for optimizing wheat yield and enhancing stress responses during critical growth phases. Further analysis of the expression data under abiotic stress conditions, specifically PEG6000, drought/heat, and salt stress, demonstrated distinct expression patterns for various TaGLK TFs (Figure 6B). The results indicated that several TaGLKs, including TaGLK42, TaGLK11, TaGLK36, and TaGLK23, exhibited significantly higher expression compared to the control in all stress treatments. Conversely, some TaGLKs, such as TaGLK98, TaGLK108, TaGLK117, and TaGLK32, were induced in specific treatments while showing lower expression in others. Most TaGLKs in subgroup V had reduced expression under different abiotic stress conditions. These findings suggest a complex regulatory network in which different TaGLKs may interact to manage stress signaling and physiological responses. The temporal expression changes imply that TaGLKs could be involved in early stress detection and subsequent adaptive responses, helping to balance wheat growth and stress tolerance. The expression patterns of the TaGLKs also differ under different biotic treatment conditions (Figure 6C). Most TaGLKs were highly expressed across various treatment time points compared to the control. However, some TaGLKs exhibited higher expression in certain treatments while showing lower levels in others. For instance, TaGLK34 and TaGLK14 had increased expression during stripe rust and powdery mildew infections but lower levels during Fusarium head blight (FHB) infection. Conversely, TaGLK48 and TaGLK18 were expressed at higher levels under stripe rust and FHB but showed reduced expression in response to powdery mildew. These results indicate that TaGLKs play a vital role in wheat response to pathogens by modulating their expression in response to specific pathogens, thereby enhancing the wheat’s ability to mount effective defense mechanisms during disease-induced stress.
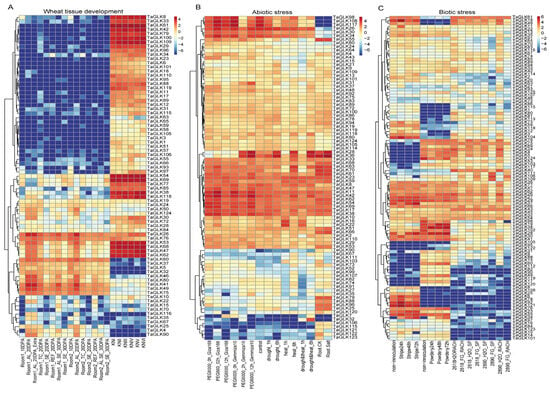
Figure 6.
The expression of TaGLKs under different tissue development, biotic, and abiotic stress conditions. (A) The expression levels of TaGLKs in various tissue development stages. (B) Expression of TaGLKs under different abiotic stresses. (C) Expression of TaGLKs under different biotic stresses.
3.6. RT-qPCR Analysis of Wheat Biotic and Abiotic Stress Responses
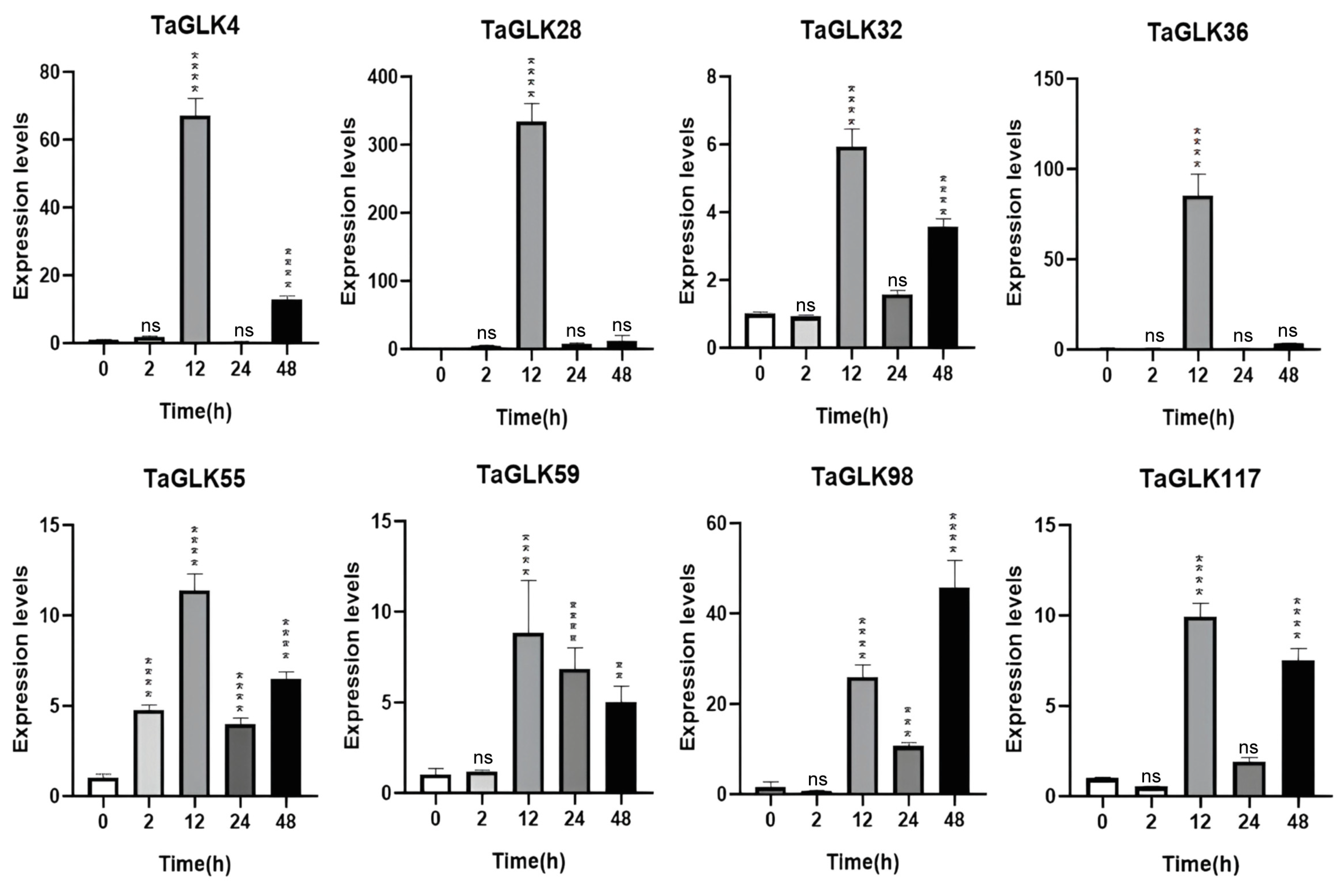
To investigate the potential roles of the TaGLKs in response to biotic and abiotic stresses, the expression levels of TaGLK4, TaGLK28, TaGLK32, TaGLK36, TaGLK55, TaGLK59, TaGLK98, and TaGLK117 were quantified using RT-qPCR under FHB infection, ABA treatment, and osmotic stress. Under PEG-induced osmotic stress progression, most TaGLKs were significantly upregulated at the 12 h time point. During the early stages of the PEG treatment (2 h), expression of these TaGLKs was generally low, except for TaGLK55, which showed significant induction. This indicates that TaGLK55 is potentially involved in wheat osmotic stress rapid regulatory function. By 24 h, expression of all genes had decreased, with TaGLK4 and TaGLK36 downregulated, while the other TaGLKs remained upregulated compared to the control. After 48 h, most TaGLKs exhibited a significant increase in expression again, except for TaGLK59, suggesting that these TaGLK TFs may be important for later stages of the osmotic stress response, while TaGLK59 may either not be essential during later stages or could be regulated differently. The highest expression at 48 h was observed for TaGLK98, which was markedly higher than the 12 h time point. In contrast, TaGLK98 expression remained low during the initial 0–2 h period before a substantial induction peak at 48 h (Figure 7). This suggests that TaGLK98 may play an important function in wheat’s longer-term physiological or developmental adaptations under osmotic stress. The subsequent decrease in expression of TaGLK4 and TaGLK36 at 24 h, alongside the overall modulation of other TaGLKs, points to a complex regulatory network adjusting to prolonged stress.
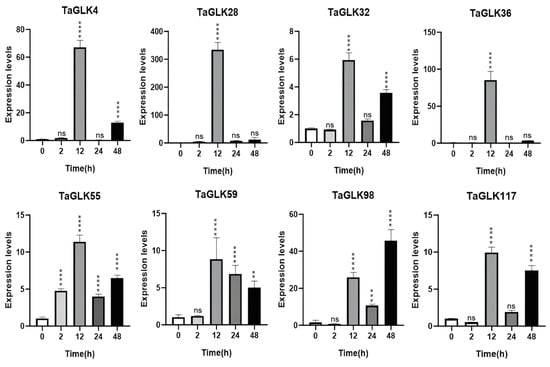
Figure 7.
Expression levels of TaGLKs under drought stress. Significant differences between three biological replicates are denoted by asterisks, indicating statistical difference at p < 0.05. The levels of significance are indicated as p < 0.0001 (****), p < 0.0005 (***), p < 0.001 (**), and non-significant (ns) between treatment and control groups.
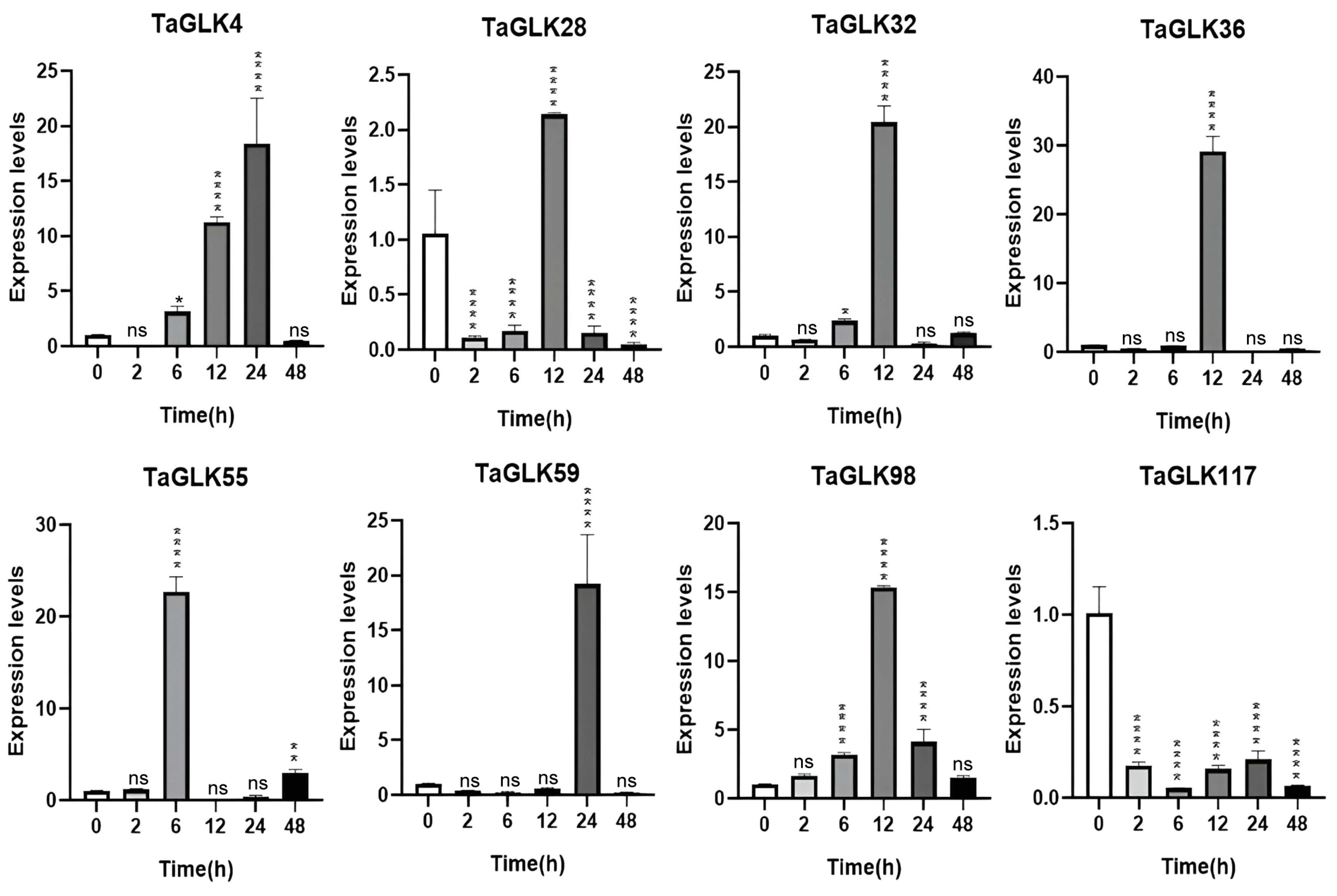
The TaGLKs exhibited distinct expression patterns in response to ABA treatment. TaGLK4 expression was downregulated at 2 h, gradually increased to a peak at 24 h, and then decreased again by 48 h, suggesting its involvement in delayed response mechanisms related to later stages of stress response or recovery in wheat (Figure 8). TaGLK28 and TaGLK36 were downregulated at 2 and 6 h, strongly induced at 12 h, and then decreased by 24–48 h, indicating their early response to ABA and a potential tightly regulated feedback mechanism to prevent over-activation. TaGLK32 increased gradually, peaking at 12 h, then decreased at 24 h, before being upregulated again by 48 h. TaGLK55 remained stable at 2 h, peaked at 6 h, decreased at 12 h, then gradually increased to be upregulated at 48 h. TaGLK59 was downregulated from 2–12 h, strongly upregulated at 24 h, then decreased by 48 h. TaGLK98 was consistently upregulated, peaking at 12 h and decreasing at 24–48 h, indicating that TaGLK98 may be involved in an important function in wheat ABA response by coordinating stress-related signaling pathways. TaGLK117, on the other hand, was downregulated at all time points (Figure 8), suggesting that it may not participate in the ABA response or could function to suppress certain stress pathways.
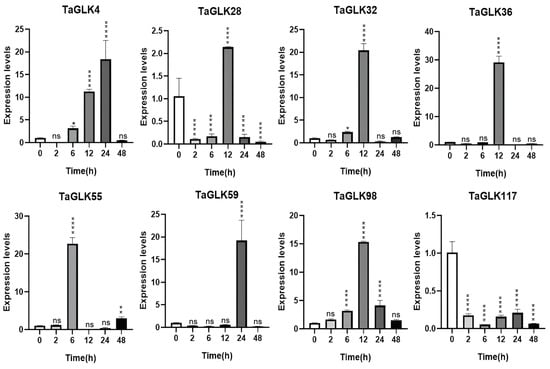
Figure 8.
Expression levels of TaGLKs under abscisic acid (ABA) treatment. Significant differences between three biological replicates are denoted by asterisks, indicating statistical difference at p < 0.05. The levels of significance are indicated as p < 0.0001 (****), p < 0.001 (**), p < 0.05 (*), and non-significant (ns) between treatment and control groups.
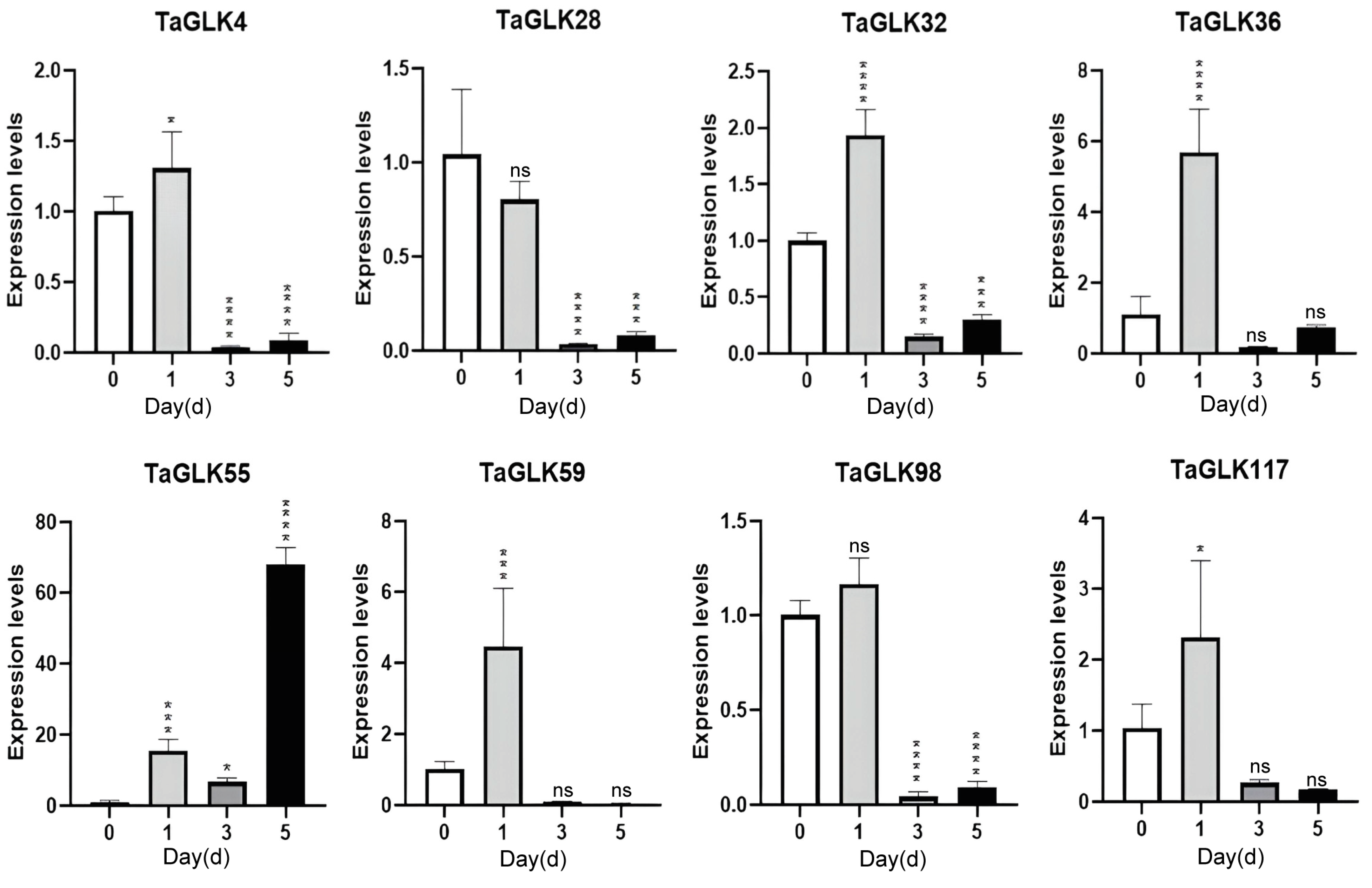
In response to F. graminearum infection, the TaGLKs showed a similar overall expression pattern. With the exception of TaGLK28, all TaGLKs were upregulated after 1 h of treatment, suggesting that these TaGLKs may be involved in wheat’s early defense mechanisms in response to FHB diseases. TaGLK4, TaGLK28, TaGLK32, TaGLK36, and TaGLK98 were subsequently downregulated at 3 h but increased again at 5 h, although their levels remained below those of the control. This may indicate a complex and time-dependent regulation of the TaGLKs in response to FHB infection. TaGLK55 decreased at 3 h, then increased at 5 h compared to 1 h. TaGLK59 and TaGLK117 gradually decreased from 3 to 5 h. TaGLK28 was downregulated throughout the time course (Figure 9), indicating a potential negative regulatory role in wheat’s regulation of FHB infection. The results revealed dynamic transcriptional responses of the TaGLKs to biotic and abiotic stresses in wheat, indicating their important roles in the wheat stress response network.
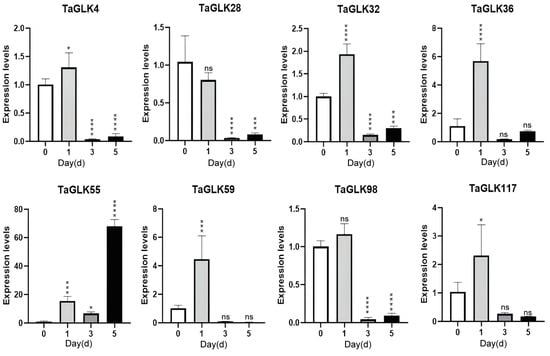
Figure 9.
Expression levels of TaGLKs under Fusarium graminearum infection. Significant differences between three biological replicates are denoted by asterisks, indicating statistical difference at p < 0.05. The levels of significance are indicated as p < 0.0001 (****), p < 0.0005 (***), p < 0.05 (*), and non-significant (ns) between treatment and control groups.
4. Discussion
GLK transcription factors are part of the Myb family within the GARP superfamily of transcription factors. These GLK proteins are present in several plants, such as tobacco, maize, Arabidopsis, and cotton, where they play roles in various stress responses as well as in plant growth and development [44]. In the present study, we investigated the motifs, physicochemical properties, structure, and expression levels of the GLK protein family in wheat under abiotic and biotic stresses. The findings suggest that TaGLKs are activated in response to these stresses, indicating their important role in regulating wheat’s response to challenging environmental conditions.
Previous research has identified 27 GLK TFs in sweet orange, 59 in foxtail millet, and 86 in tobacco [45,46,47]. An analysis of the physicochemical properties of GLK proteins from these species showed significant variation in their amino acid lengths, isoelectric points, and molecular weights. Additionally, all identified GLKs contained the conserved Myb-DNA binding domain, along with others, including the REC and Myb CC LHEQLE domains. In the present study, we identified 125 GLK proteins in the wheat genome. Similar to GLK proteins from other plant species, all TaGLKs have the Myb-DNA binding domain along with additional domains, including Myb CC LHEQLE, SANT, and REC. This indicates that TaGLK proteins may act as transcriptional regulators that integrate multiple cellular signals and coordinate various developmental and physiological processes in wheat. Analysis of the physicochemical characteristics of TaGLK proteins found considerable variation in sequence length, molecular weight, and isoelectric point, suggesting that this diversity may facilitate specialized functions, adaptability, and complex transcriptional regulation in wheat.
Previous studies on foxtail millet (Setaria italica L.) and cotton (Gossypium hirsutum) have found that the GLKs in these plant species are primarily localized in the nucleus and chloroplast [46,48]. The current study’s subcellular localization analysis discovered that, with the exception of TaGLK70, TaGLK63, and TaGLK55, which were located only in the chloroplast, all the other TaGLKs were localized in the nucleus. Furthermore, three TaGLKs (TaGLK111, TaGLK44, and TaGLK11) were found in both the chloroplast and the nucleus, while TaGLK23 was localized in the cell membrane, chloroplast, and nucleus. The analysis also found that seven of the TaGLKs (TaGLK13, TaGLK19, TaGLK24, TaGLK78, TaGLK86, TaGLK88, and TaGLK94) localized in the nucleus are likely to be stable proteins. These findings suggest a complex regulatory network that promotes functional specialization and subcellular partitioning of the TaGLKs, which may contribute to the adaptability and resilience of wheat.
Previous research on GLK proteins in tomato and maize has identified seven distinct groups [49]. In our study, evolutionary analysis of the TaGLK proteins also revealed six subgroups, each characterized by similar conserved motifs and domains. Notably, rice GLK (OsGLK32) associated with phosphate starvation response regulatory factors was classified into subgroup IV. In contrast, two TaGLKs containing the REC domain were categorized in subgroup II. The REC domain is primarily linked to B-type response regulators, which form part of a two-component signal transduction system that can either activate or inhibit target gene transcription based on environmental conditions [50]. Furthermore, the AtGLK1 and AtGLK2, which belong to subgroup II, are known to positively influence DNA-templated transcription and chloroplast development in Arabidopsis. Phylogenetic analysis indicated that all six subgroups of the TaGLK contain members that possess the conserved Myb-like DNA binding domain and the SANT domain. This suggests that these TaGLK subgroups may share similar functional roles in DNA binding and the regulation of gene expression. Additionally, the subgroup II members TaGLK5 and TaGLK125 were found to contain the REC superfamily, indicating a complex evolutionary history and functional diversification of GLK TFs in wheat. This diversification likely enhances their ability to regulate various physiological and developmental processes. Interestingly, TaGLK87 was noted to possess both the REC superfamily and AtoC domains, suggesting it has the functionalities of both a sensor kinase and a response regulator, allowing it to participate in two-component regulatory systems. Chromosome mapping of the TaGLKs revealed that the largest cluster, consisting of 10 TaGLKs, is located on chromosome 7D, while chromosomes 7A and 7B each harbor nine TaGLKs. In contrast, chromosomes 1A, 3A, and Un each contain only TaGLK125. This distribution suggests gene duplication, functional specialization, and chromosomal rearrangements during the evolutionary history of wheat GLKs.
Research conducted on tomato [51] and G. hirsutum [48] indicated that the exon–intron structures of GLK subfamilies are quite similar, with the majority of members having between 4 and 7 exons. Supporting these prior findings, the current study demonstrated variations in the number of exons, UTRs, and introns among the TaGLKs, with exon counts ranging from 4 to 8 and most TaGLKs containing 5 to 7 exons. This indicates functional specialization and evolutionary adaptation within the TaGLKs. The analysis of conserved motifs revealed that each TaGLK protein possesses between 2 and 10 unique motifs, with subgroup III TaGLKs each having five motifs. These findings reinforce the concept of sub-functionalization and divergent evolution within the TaGLK proteins, which may enhance the overall transcriptional complexity and regulatory flexibility of these TFs in wheat. A previous study on soybeans identified cis-acting elements in GmGLKs promoters that play regulatory roles in abscisic acid, methyl jasmonate, auxin signaling, low temperatures, and various biotic and abiotic stresses [52]. In line with these earlier studies, the current analysis of TaGLK promoters revealed a range of stress-responsive elements related to both biotic and abiotic stress responses, as well as elements linked to plant growth and development. This indicates that TaGLKs may have intricate regulatory roles in mediating stress responses and developmental processes in wheat.
Gene expression data from the WheatOmics database indicated that the TaGLKs were significantly altered in response to biotic and abiotic stresses, as well as during various stages of wheat tissue development. This suggests that TaGLKs have specific roles in regulating stress responses in wheat. Under biotic stress conditions, the expression levels of TaGLKs from different subgroups varied markedly across developmental stages and in response to pathogen infections. These observations highlight the involvement of these TaGLK subgroups in the disease defense mechanisms of wheat. The results indicate that TaGLKs display diverse and complex expression patterns in reaction to various biotic and abiotic conditions, suggesting their role in enhancing wheat’s adaptability and survival under different environmental conditions. KEGG pathway enrichment analysis revealed that the TaGLKs were particularly enriched in pathways related to cutin, suberin, and wax biosynthesis. These lipid polymers are crucial in plant cell walls, playing key roles in regulating responses to biotic and abiotic stresses. Cutin and suberin create protective barriers against pathogens and reduce water loss, while waxes improve water repellency. The biosynthesis of these compounds is often upregulated under stress conditions, underscoring their vital role in the plant’s adaptive strategies for surviving diverse environmental challenges [53].
Previous research has shown that GLK TFs play significant regulatory roles in plant stress responses. Experiments involving overexpression and knockdown have demonstrated that rice GLK1 is vital for resistance to the rice black-streaked dwarf virus and is associated with a significant quantitative trait locus (QTL) [54]. Another study identified a potential regulatory function for ZmGLKs in the ABA signaling pathway, revealing that GLKs can directly activate the expression of WRKY40 and its complex, which negatively regulates ABA signaling in Arabidopsis [22]. Additionally, recent findings by Li et al. [28] showed that constitutive expression of maize GLK in rice can enhance stomatal conductance and photosynthetic efficiency under field conditions, leading to improved drought tolerance. This enhanced drought tolerance in rice overexpressing ZmGLK1 was linked to the rapid closure of stomata mediated by ABA. The expression of the tomato GLK45 was found to be induced by the plant hormones auxin (IAA) and jasmonic acid (JA), while being repressed by abiotic stresses such as cold, salt, and drought, as well as by the hormones ABA and gibberellic acid (GA) [5]. In line with these previous findings, the current study observed dynamic and complex regulation of TaGLKs in response to osmotic stress, ABA treatment, and F. graminearum infection, suggesting their potential role in wheat’s stress response mechanisms.
Under progressive osmotic stress, the majority of the TaGLKs were upregulated after 12 h of treatment. However, expression levels varied at different time points, with some TaGLKs experiencing downregulation while others remained upregulated, suggesting their potentially diverse roles in wheat’s response to osmotic stress. The TaGLKs also displayed distinct expression patterns in response to ABA treatment. For example, TaGLK117 was consistently downregulated throughout the treatment, while others, such as TaGLK98, showed consistent upregulation. This indicates that TaGLKs may have varying functions in ABA-mediated stress responses in wheat. In the context of the wheat–Fusarium pathosystem, all TaGLKs, except for TaGLK55, were initially upregulated in response to Fusarium head blight (FHB) infection but exhibited decreased expression at later time points. This indicates that TaGLKs play a key role in wheat defense mechanisms against this significant fungal disease.
5. Conclusions
This study identified and thoroughly analyzed the GLK proteins in wheat. The study discovered 125 TaGLK proteins, and comprehensive analyses, including examination of sequence structure, chromosome localization, cis-regulatory elements, and expression analysis, were performed on the identified TaGLKs. The expression levels of the TaGLKs under abiotic and biotic stress conditions were analyzed. The study found that TaGLKs are potential transcriptional regulators with the capacity to integrate multiple cellular signals and coordinate various developmental and physiological processes. Phylogenetic analyses categorized the TaGLKs into six subgroups and revealed a complex evolutionary history and functional diversification of the TaGLK protein family. PPI network reveals that TaGLKs are involved in photoreceptor activity, cell cycle progression, and protein regulation in wheat. Gene expression analysis showed that TaGLKs are involved in wheat developmental processes and biotic and abiotic stress responses. RT-qPCR analysis discovered that TaGLKs are involved in osmotic stress, ABA stress, and F. graminearum disease regulatory functions in wheat. The study provides fundamental knowledge for future studies of the roles of TaGLKs in wheat stress responses and development. Future efforts to improve wheat tolerance to abiotic and biotic stress conditions can benefit from the findings of this study.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy14123070/s1, Table S1: All primer sequences in this study, Table S2: Prediction and analysis of basic physicochemical properties of 125 wheat TaGLK in this study, Table S3: GO-term and pathway annotation enrichment of TaGLK proteins, Figure S1: Conserved domains of TaGLK proteins.
Author Contributions
All authors contributed to the study conception and design, developed the experimental design, conceived and performed all experiments, analyzed the results and corresponding data, and wrote the manuscript draft. Project administration and funding acquisition, D.M.; Conceptualization and methodology, Y.L.; Software, formal analysis, and writing—original draft preparation, J.X. and P.L.; Formal analysis, writing—review and editing, Y.W. and D.B.; Validation and resources, Y.W. and J.S.; Visualization and supervision, Y.L.; Investigation, X.L.; Data curation, S.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Natural Science Funds of Hubei Province of China (2024AFB1015).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors have no conflicts of interest to declare.
References
- Bohnert, H.J.; Gong, Q.; Li, P.; Ma, S. Unraveling abiotic stress tolerance mechanisms–getting genomics going. Curr. Opin. Plant Biol. 2006, 9, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Zaikina, E.A.; Rumyantsev, S.D.; Sarvarova, E.; Kuluev, B. Transcription factor genes involved in plant response to abiotic stress factors. Ecol. Genet. 2019, 17, 47–58. [Google Scholar] [CrossRef]
- Song, A.; Wu, D.; Fan, Q.; Tian, C.; Chen, S.; Guan, Z.; Xin, J.; Zhao, K.; Chen, F. Transcriptome-wide identification and expression profiling analysis of chrysanthemum trihelix transcription factors. Int. J. Mol. Sci. 2016, 17, 198. [Google Scholar] [CrossRef]
- Nguyen, H.C.; Lin, K.H.; Ho, S.L.; Chiang, C.M.; Chi, M.Y. Enhancing the abiotic stress tolerance of plants: From chemical treatment to biotechnological approaches. Physiol. Plant. 2018, 164, 452–466. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zheng, C.; Shao, X.; Hu, Z.; Li, J.; Wang, P.; Wang, A.; Yu, J.; Shi, K. Transcriptomic and genetic approaches reveal an essential role of the NAC transcription factor SlNAP1 in the growth and defense response of tomato. Hortic. Res. 2020, 7, 11. [Google Scholar] [CrossRef]
- Kou, X.; Wang, S.; Wu, M.; Guo, R.; Xue, Z.; Meng, N.; Tao, X.; Chen, M.; Zhang, Y. Molecular Characterization and Expression Analysis of NAC Family Transcription Factors in Tomato. Plant Mol. Biol. Rep. 2014, 32, 501–516. [Google Scholar] [CrossRef]
- Li, M.; Lee, K.P.; Liu, T.; Dogra, V.; Duan, J.; Li, M.; Xing, W.; Kim, C. Antagonistic modules regulate photosynthesis-associated nuclear genes via GOLDEN2-LIKE transcription factors. Plant Physiol. 2021, 188, 2308–2324. [Google Scholar] [CrossRef]
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R.; et al. Arabidopsis transcription factors: Genome-wide comparative analysis among eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar] [CrossRef] [PubMed]
- Rossini, L.; Cribb, L.; Martin, D.J.; Langdale, J.A. The maize golden2 gene defines a novel class of transcriptional regulators in plants. Plant Cell 2001, 13, 1231–1244. [Google Scholar] [CrossRef] [PubMed]
- Han, X.Y.; Li, P.X.; Zou, L.J.; Tan, W.R.; Zheng, T.; Zhang, D.W.; Lin, H.H. GOLDEN2-LIKE transcription factors coordinate the tolerance to Cucumber mosaic virus in Arabidopsis. Biochem. Biophys. Res. Commun. 2016, 477, 626–632. [Google Scholar] [CrossRef]
- Wu, R.; Guo, L.; Wang, R.; Zhang, Q.; Yao, H. Genome-Wide Identification and Characterization of G2-Like Transcription Factor Genes in Moso Bamboo (Phyllostachys edulis). Molecules 2022, 27, 5491. [Google Scholar] [CrossRef] [PubMed]
- Yasumura, Y.; Moylan, E.C.; Langdale, J.A. A conserved transcription factor mediates nuclear control of organelle biogenesis in anciently diverged land plants. Plant Cell 2005, 17, 1894–1907. [Google Scholar] [CrossRef]
- Murmu, J.; Wilton, M.; Allard, G.; Pandeya, R.; Desveaux, D.; Singh, J.; Subramaniam, R. Arabidopsis GOLDEN2-LIKE (GLK) transcription factors activate jasmonic acid (JA)-dependent disease susceptibility to the biotrophic pathogen Hyaloperonospora arabidopsidis, as well as JA-independent plant immunity against the necrotrophic pathogen Botrytis cinerea. Mol. Plant Pathol. 2014, 15, 174–184. [Google Scholar]
- Chen, M.; Ji, M.; Wen, B.; Liu, L.; Li, S.; Chen, X.; Gao, D.; Li, L. GOLDEN 2-LIKE transcription factors of plants. Front. Plant Sci. 2016, 7, 1509. [Google Scholar] [CrossRef]
- Wang, H.; Seo, J.K.; Gao, S.; Cui, X.; Jin, H. Silencing of AtRAP, a target gene of a bacteria-induced small RNA, triggers antibacterial defense responses through activation of LSU2 and down-regulation of GLK1. New Phytol. 2017, 215, 1144–1155. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Choi, H.; Ti, T.; Gwak, D.; Ha, S.H. Pleiotropic properties of GOLDEN2-LIKE transcription factors for crop improvement. Appl. Biol. Chem. 2023, 66, 81. [Google Scholar] [CrossRef]
- Fan, P.; Nie, L.; Jiang, P.; Feng, J.; Lv, S.; Chen, X.; Bao, H.; Guo, J.; Tai, F.; Wang, J.; et al. Transcriptome analysis of Salicornia europaea under saline conditions revealed the adaptive primary metabolic pathways as early events to facilitate salt adaptation. PLoS ONE 2013, 8, e80595-18. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Choi, B.; Yun, A.; Son, N.; Ahn, G.; Cha, J.Y.; Kim, W.Y.; Hwang, I. Long-term ABA promotes GLK1 degradation through COP1 in a light intensity-dependent manner to suppress chloroplast development. Plant Cell Environ. 2021, 44, 3034–3048. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, L.; Zhang, B.; Zeng, L.; Li, L. AhHDA1-mediated AhGLK1 promoted chlorophyll synthesis and photosynthesis regulates recovery growth of peanut leaves after water stress. Plant Sci. 2020, 294, 110461. [Google Scholar] [CrossRef]
- Liu, J.; Mehari, T.G.; Xu, Y.; Umer, M.J.; Hou, Y.; Wang, Y.; Peng, R.; Wang, K.; Cai, X.; Zhou, Z.; et al. GhGLK1 a key candidate gene from GARP family enhances cold and drought stress tolerance in cotton. Front. Plant Sci. 2021, 12, 759312. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; Liu, Y.; Wang, T.J.; Meng, Q.; Yin, H.; Wang, X.; Wu, Y.; Nan, N.; Liu, B.; Xu, Z.Y. GOLDEN 2-LIKE transcription factors regulate WRKY40 expression in response to abscisic acid. Plant Physiol. 2019, 179, 1844–1860. [Google Scholar] [CrossRef] [PubMed]
- Savitch, L.V.; Allard, G.; Seki, M.; Robert, L.S.; Tinker, N.A.; Huner, N.P.; Shinozaki, K.; Singh, J. The effect of overexpression of two Brassica CBF/DREB1-like transcription factors on photosynthetic capacity and freezing tolerance in Brassica napus. Plant Cell Physiol. 2005, 46, 1525–1539. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Lee, J.; Choi, B.; Park, Y.; Sim, H.J.; Kim, H.; Hwang, I. Physiological and molecular processes associated with long duration of ABA treatment. Front. Plant Sci. 2018, 9, 176. [Google Scholar] [CrossRef]
- Khalid, A.; Hameed, A.; Tahir, M.F. Wheat quality: A review on chemical composition, nutritional attributes, grain anatomy, types, classification, and function of seed storage proteins in bread making quality. Front. Nutr. 2023, 10, 1053196. [Google Scholar] [CrossRef] [PubMed]
- Khalid, A.; Hameed, A.; Shamim, S.; Ahmad, J. Divergence in single kernel characteristics and grain nutritional profiles of wheat genetic resource and association among traits. Front. Nutr. 2022, 8, 805446. [Google Scholar] [CrossRef]
- Wattoo, F.M.; Rana, R.M.; Fiaz, S. Transcriptional Factors’ Response Under Biotic Stress in Wheat. In Transcription Factors for Biotic Stress Tolerance in Plants; Springer International Publishing: Cham, Switzerland, 2022; pp. 129–141. [Google Scholar]
- Liu, H.; Yang, Y.; Zhang, L. Zinc Finger-Homeodomain Transcriptional Factors (ZF-HDs) in Wheat (Triticum aestivum L.): Identification, Evolution, Expression Analysis and Response to Abiotic Stresses. Plants 2021, 10, 593. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, J.; Wei, S.; Gao, Y.; Pei, H.; Geng, R.; Lu, Z.; Wang, P.; Zhou, W. Maize GOLDEN2-LIKE proteins enhance drought tolerance in rice by promoting stomatal closure. Plant Physiol. 2024, 194, 774–786. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 60 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v6: Recent updates to the phylogenetic tree display and annotation tool. Nucleic Acids Res. 2024, 52, W78–W82. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 8, 1296–1297. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a plant cis-acting regulatory element database. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S.; et al. The STRING database in 2023: Protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2023, 51, D638–D646. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Ma, S.; Wang, M.; Wu, J.; Guo, W.; Chen, Y.; Li, G.; Wang, Y.; Shi, W.; Xia, G.; Fu, D.; et al. WheatOmics: A platform combining multiple omics data to accelerate functional genomics studies in wheat. Mol. Plant 2021, 14, 1965–1968. [Google Scholar] [CrossRef]
- Lyu, F.; Han, F.; Ge, C.; Mao, W.; Chen, L.; Hu, H.; Chen, G.; Lang, Q.; Fang, C. OmicStudio: A composable bioinformatics cloud platform with real-time feedback that can generate high-quality graphs for publication. Imeta 2023, 2, e85. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Geng, Y.; Liu, Y.; Chen, S.; Cao, S.; Li, W.; Chen, H.; Ma, D.; Yin, J. Genome-wide identification and characterization of SRO gene family in wheat: Molecular evolution and expression profiles during different stresses. Plant Physiol. Biochem. PPB 2020, 154, 590–611. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Su, C.; Wang, Y.; Xu, X.; Li, Y.; Ma, D. Genome-wide identification of Glutathione peroxidase (GPX) family genes and silencing TaGPX3.2A reduced disease resistance in wheat. Plant Physiol. Biochem. PPB 2023, 204, 108139. [Google Scholar] [CrossRef]
- Jiang, C.; Cao, S.; Wang, Z.; Xu, H.; Liang, J.; Liu, H.; Wang, G.; Ding, M.; Wang, Q.; Gong, C.; et al. An expanded subfamily of G-protein-coupled receptor genes in Fusarium graminearum required for wheat infection. Nat. Microbiol. 2019, 4, 1582–1591. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Verdeja, T.; Lundgren, M.R. GOLDEN2-LIKE transcription factors: A golden ticket to improve crops? Plants People Planet 2023, 6, 79–93. [Google Scholar] [CrossRef]
- Xiong, B.; Gong, Y.; Li, Q.; Li, L.; Mao, H.; Liao, L.; Wang, X.; Deng, H.; Zhang, M.; Wang, Z. Genome-Wide Analysis of the GLK Gene Family and the Expression under Different Growth Stages and Dark Stress in Sweet Orange (Citrus sinensis). Horticulturae 2022, 8, 1076. [Google Scholar] [CrossRef]
- Chen, H.; Qin, L.; Wang, X. Identification and Evolutionary Analysis of the GOLDEN 2-LIKE Gene Family in Foxtail Millet. Trop. Plant Biol. 2022, 15, 301–318. [Google Scholar] [CrossRef]
- Qin, M.; Zhang, B.; Gu, G.; Yuan, J.; Yang, X.; Yang, J.; Xie, X. Genome-Wide Analysis of the G2-Like Transcription Factor Genes and Their Expression in Different Senescence Stages of Tobacco (Nicotiana tabacum L.). Front. Genet. 2021, 12, 787. [Google Scholar] [CrossRef]
- Zhao, Z.; Shuang, J.; Li, Z.; Xiao, H.; Liu, Y.; Wang, T.; Wei, Y.; Hu, S.; Wan, S.; Peng, R. Identification of the Golden-2-like transcription factors gene family in Gossypium hirsutum. PeerJ 2021, 9, e12484. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Xu, Y.; Han, G.; Zhou, L.; Ali, A.; Zhu, S.; Li, X. Molecular Evolution and Genetic Variation of G2-Like Transcription Factor Genes in Maize. PLoS ONE 2016, 11, e0161763. [Google Scholar] [CrossRef]
- Ni, J.; Bai, S.; Gao, L.; Qian, M.; Zhong, L.; Teng, Y. Identification, classification, and transcription profiles of the B-type response regulator family in pear. PLoS ONE 2017, 12, e0171523. [Google Scholar] [CrossRef]
- Liu, J.F. Bioinformatics Analysis of Tomato G2-Like Transcription Factor Family and Identification of Resistance-Related Genes. Doctoral Thesis, College of Horticulture & Landscape Architecture, West Lafayette, IN, USA, 2018. [Google Scholar]
- Alam, I.; Manghwar, H.; Zhang, H.; Yu, Q.; Ge, L. Identification of GOLDEN2-like transcription factor genes in soybeans and their role in regulating plant development and metal ion stresses. Front. Plant Sci. 2022, 13, 1052659. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, M.; Keyl, A.; Feussner, I. Wax biosynthesis in response to danger: Its regulation upon abiotic and biotic stress. New Phytologist. 2020, 227, 698–713. [Google Scholar] [CrossRef]
- Li, X.; Lin, F.; Li, C.; Du, L.; Liu, Z.; Shi, W.; Lv, J.; Cao, X.; Lan, Y.; Fan, Y. Golden 2-like transcription factor contributes to the major QTL against rice black-streaked dwarf virus disease. Theor. Appl. Genet. 2022, 135, 4233–4243. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).