Natural Variation in the AAT1 Promoter Is Responsible for the Disparity in Ester Aroma Between Actinidia chinensis and Actinidia eriantha


Abstract
1. Introduction
2. Materials and Methods
2.1. Material Collection
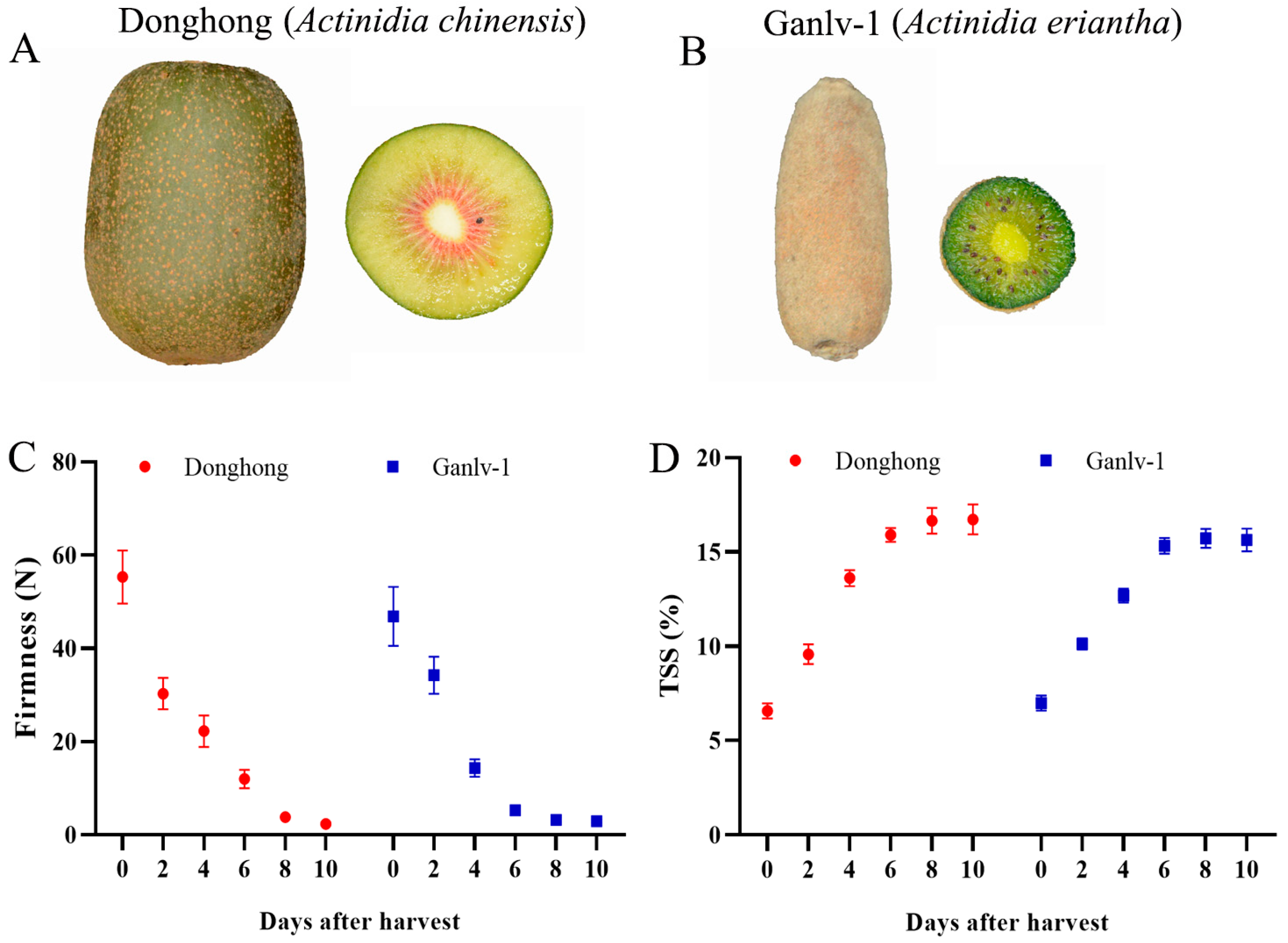
2.2. Fruit Firmness and the TSS
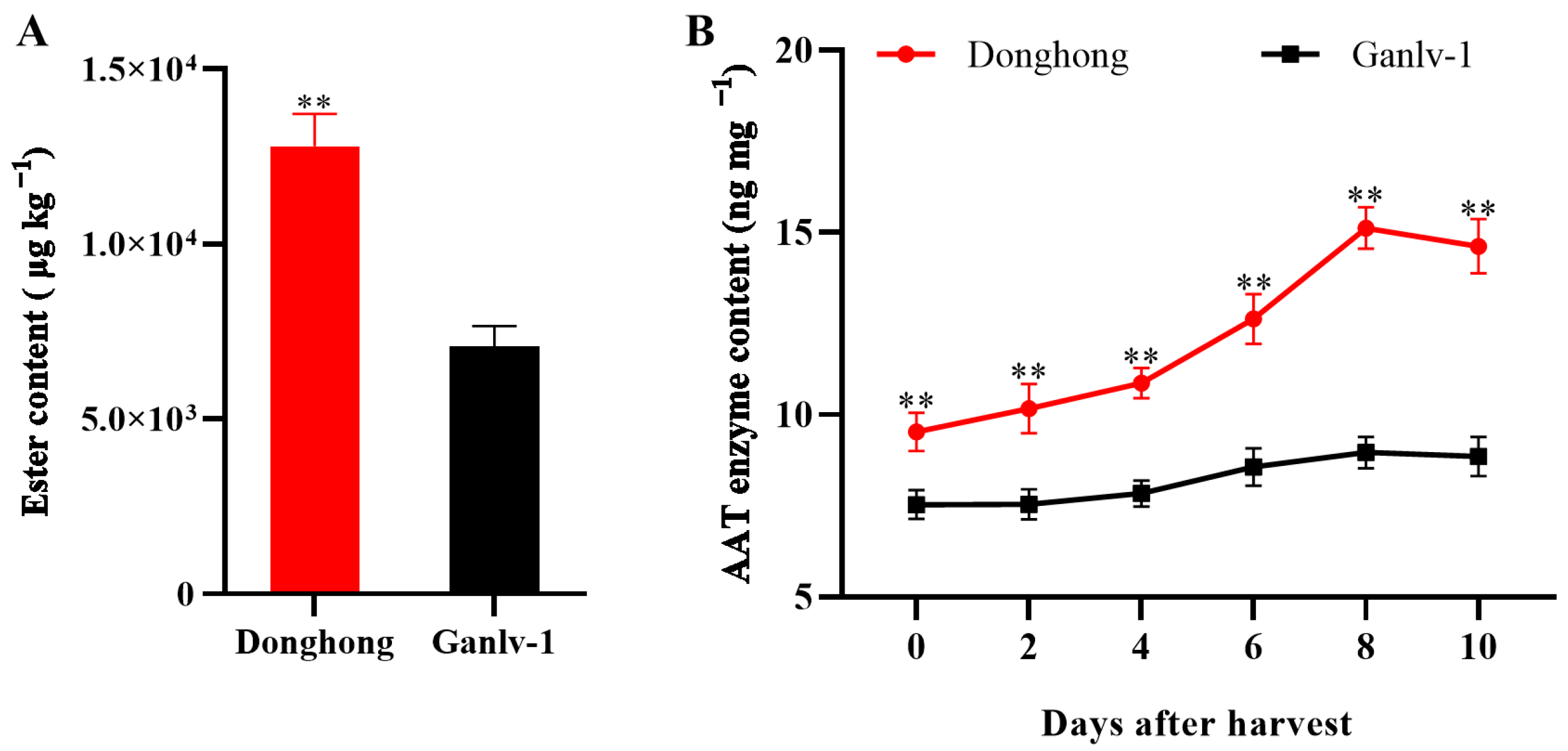
2.3. Extraction and Determination of Esters
2.4. Determination of AAT Content
2.5. RNA Extraction and Gene Expression Analysis
2.6. AAT1 Transient Overexpression in Kiwifruit
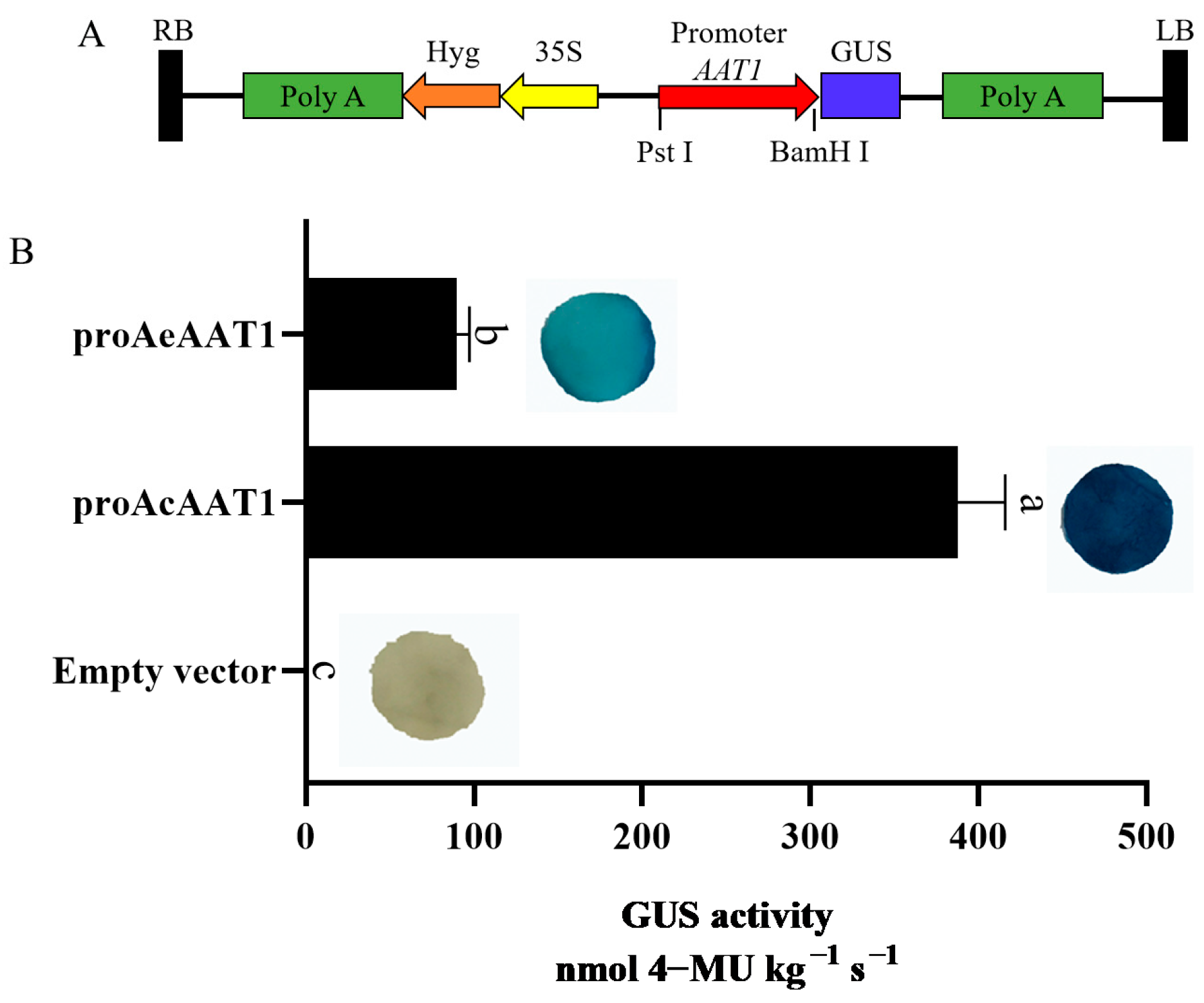
2.7. Analysis of AAT1 Promoter Activity
2.8. Statistical Analysis
3. Results
3.1. Volatile Esters in Ripe Actinidia chinensis and Actinidia eriantha
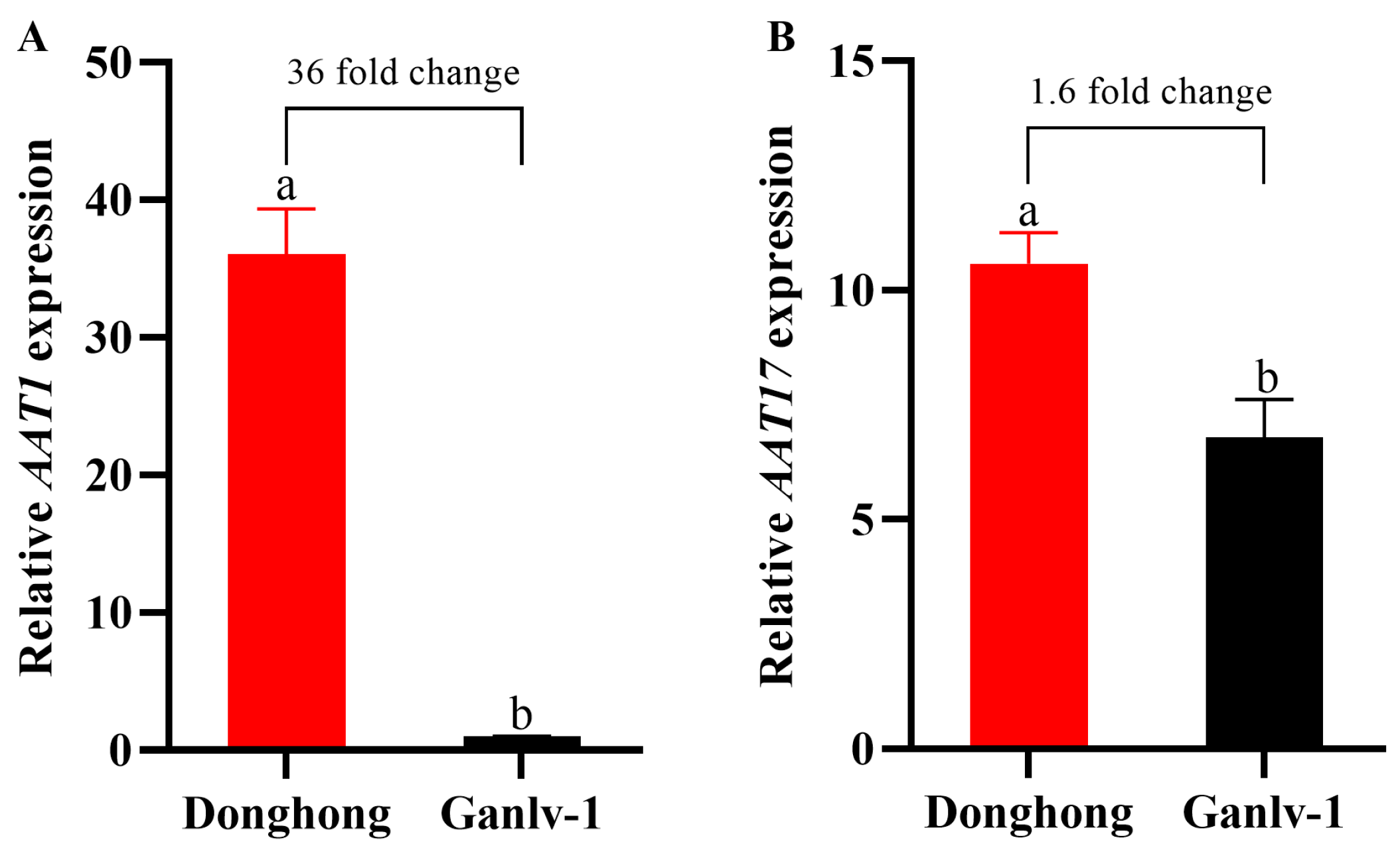
3.2. Changes in the Expression of AATs in Ripe Actinidia chinensis and Actinidia eriantha
3.3. Overexpression of the AAT1 Gene Alters Volatile Ester Content
3.4. Analysis of AAT1 Promoter Activity Differences Between Actinidia chinensis and Actinidia eriantha
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fan, J.B.; Zhong, C.H. Fruit scientific research in New China in the past 70 years: Kiwifruit. J. Fruit Sci. 2019, 36, 1352–1359. [Google Scholar] [CrossRef]
- Li, X.W.; Li, J.Q.; Soejarto, D.D. New synonyms in Actinidiaceae from China. Acta Phytotax Sin. 2007, 45, 633–660. [Google Scholar] [CrossRef]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef] [PubMed]
- El Hadi, M.A.M.; Zhang, F.J.; Wu, F.F.; Zhou, C.H.; Tao, J. Advances in fruit aroma volatile research. Molecules 2013, 18, 8200–8229. [Google Scholar] [CrossRef]
- Ye, Y.H.; Zheng, S.Y.; Wang, Y.X. Analysis of aroma components changes in Gannan navel orange at different growth stages by HS-SPME-GC-MS, OAV, and multivariate analysis. Food Res. Int. 2024, 175, 113622. [Google Scholar] [CrossRef]
- Li, R.; Yan, D.; Tan, C.Y.; Li, C.; Song, M.J.; Zhao, Q.Q.; Yang, Y.M.; Yin, W.J.; Liu, Z.D.; Ren, X.L.; et al. Transcriptome and metabolomics integrated analysis reveals MdMYB94 associated with esters biosynthesis in apple (Malus domestica). J. Agric. Food Chem. 2023, 71, 7904–7920. [Google Scholar] [CrossRef]
- Liu, Y.; Wen, H.; Yang, X.P.; Wu, C.Y.; Ming, J.Q.; Zhang, H.Y.; Chen, J.J.; Wang, J.B.; Xu, J. Metabolome and transcriptome profiling revealed the enhanced synthesis of volatile esters in Korla pear. BMC Plant Biol. 2023, 23, 264. [Google Scholar] [CrossRef]
- Garcia, C.V.; Stevenson, R.J.; Atkinson, R.G.; Winz, R.A.; Quek, S.Y. Changes in the bound aroma profiles of ‘Hayward’ and ‘Hort16A’ kiwifruit (Actinidia spp.) during ripening and GC-olfactometry analysis. Food Chem. 2013, 137, 45–54. [Google Scholar] [CrossRef]
- Xiao, Z.B.; Chen, L.N.; Niu, Y.W.; Zhu, J.C.; Zhang, J.; Deng, J.M. Evaluation of the interaction between esters and sulfur compounds in pineapple using feller’s additive model, OAV, and odor activity coefficient. Food Anal. Method 2021, 14, 1714–1729. [Google Scholar] [CrossRef]
- Xu, S.J.; He, W.Y.; Yan, J.T.; Zhang, R.G.; Wang, P.; Tian, H.L.; Zhan, P. Volatomics-assisted characterization of aroma and off-flavor contributors in fresh and thermally treated kiwifruit juice. Food Res. Int. 2023, 167, 112656. [Google Scholar] [CrossRef]
- Ye, L.Q.; Yang, C.X.; Li, W.D.; Hao, J.B.; Sun, M.; Zhang, J.R.; Zhang, Z.S. Evaluation of volatile compounds from Chinese dwarf cherry (Cerasus humilis (Bge.) Sok.) germplasms by headspace solid-phase microextraction and gas chromatography-mass spectrometry. Food Chem. 2017, 217, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.P.; Xie, Y.X.; Liu, C.H.; Chen, S.L.; Hu, S.S.; Xie, Z.Z.; Deng, X.X.; Xu, J. Comprehensive comparative analysis of volatile compounds in citrus fruits of different species. Food Chem. 2017, 230, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Young, H.; Paterson, V.J. Characterisation of bound flavour components in kiwifruit. J. Sci. Food Agric. 2010, 68, 257–260. [Google Scholar] [CrossRef]
- Chai, J.X.; Liao, B.A.; Li, R.; Liu, Z.D. Changes in taste and volatile compounds and ethylene production determined the eating window of ‘Xuxiang’ and ‘Cuixiang’ kiwifruit cultivars. Postharvest Biol. Technol. 2022, 194, 112093. [Google Scholar] [CrossRef]
- Nieuwenhuizen, N.J.; Chen, X.Y.; Wang, M.Y.; Matich, A.J.; Perez, R.L.; Allan, A.C.; Green, S.A.; Atkinson, R.G. Natural variation in monoterpene synthesis in kiwifruit: Transcriptional regulation of terpene synthases by NAC and ETHYLENE-INSENSITIVE3-like transcription factors. Plant Physiol. 2015, 167, 1243–1258. [Google Scholar] [CrossRef]
- Yang, H.Y.; Li, J.Z.; Li, X.H.; Wu, R.; Zhang, X.L.; Fan, X.G.; Li, G.T.; Gong, H.S.; Yin, X.R.; Zhang, A.D. The mechanism of gibberellins treatment suppressing kiwifruit postharvest ripening processes by transcriptome analysis. Postharvest. Biol. Technol. 2023, 198, 112223. [Google Scholar] [CrossRef]
- Stolterfoht, H.; Rinnofner, C.; Winkler, M.; Pichler, H. Recombinant lipoxygenases (LOX) and hydroperoxide lyases (HPL) for the synthesis of green leaf volatiles. J. Agric. Food Chem. 2019, 2019, 13367–13392. [Google Scholar] [CrossRef]
- Qin, G.H.; Qi, X.X.; Qi, Y.J.; Gao, Z.H.; Yi, X.K.; Pan, H.F.; Xu, Y.L. Identification and expression patterns of alcohol dehydrogenase genes involving in ester volatile biosynthesis in pear fruit. J. Integr. Agric. 2017, 16, 1742–1750. [Google Scholar] [CrossRef]
- Schwab, E.; Davidovich-Rikanati, R.; Lewinsohn, E. Biosynthesis of plant-derived flavor compounds. Plant J. 2008, 54, 712–732. [Google Scholar] [CrossRef]
- Beekwilder, J. Functional characterization of enzymes forming volatile esters from strawberry and banana. Plant Physiol. 2004, 135, 1865–1878. [Google Scholar] [CrossRef]
- Liu, C.; Qiao, X.; Li, Q.; Zeng, W.; Wei, S.; Wang, X.; Chen, Y.; Wu, X.; Wu, J.; Yin, H.; et al. Genome-wide comparative analysis of the BAHD superfamily in seven Rosaceae species and expression analysis in pear (Pyrus bretschneideri). BMC Plant Biol. 2020, 20, 14. [Google Scholar] [CrossRef] [PubMed]
- Cumplido-Laso, G.; Medina-Puche, L.; Moyano, E.; Hoffmann, T.; Sinz, Q.; Ring, L.; Studart-Wittkowski, C.; Luis Caballero, J.; Schwab, W.; Munoz-Blanco, J.; et al. The fruit ripening-related gene FaAAT2 encodes an acyl transferase involved in strawberry aroma biogenesis. J. Exp. Bot. 2012, 63, 4275–4290. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, M.; Gaete-Eastman, C.; Valdenegro, M.; Figueroa, C.R.; Fuentes, L.; Herrera, R.; Alejandra Moya-Leon, M. Aroma development during ripening of Fragaria chiloensis fruit and participation of an alcohol acyltransferase (FcAAT1) gene. J. Agric. Food Chem. 2009, 57, 9123–9132. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Shen, J.; Wu, T.; Xu, Y.; Zong, X.; Li, D.; Shu, H. Overexpression of the apple alcohol acyltransferase gene alters the profile of volatile blends in transgenic tobacco leaves. Physiol. Plantarum. 2008, 134, 394–402. [Google Scholar] [CrossRef]
- Souleyre, E.J.F.; Chagne, D.; Chen, X.; Tomes, S.; Turner, R.M.; Wang, M.Y.; Maddumage, R.; Hunt, M.B.; Winz, R.A.; Wiedow, C.; et al. The AAT1 locus is critical for the biosynthesis of esters contributing to “ripe apple” flavour in “Royal Gala” and “Granny Smith” apples. Plant J. 2014, 78, 903–915. [Google Scholar] [CrossRef]
- Cao, X.; Wei, C.; Duan, W.; Gao, Y.; Kuang, J.; Liu, M.; Chen, K.; Klee, H.; Zhang, B. Transcriptional and epigenetic analysis reveals that NAC transcription factors regulate fruit flavor ester biosynthesis. Plant J. 2021, 106, 785–800. [Google Scholar] [CrossRef]
- Crowhurst, R.N.; Gleave, A.P.; MacRae, E.A.; Ampomah-Dwamena, C.; Atkinson, R.G.; Beuning, L.L.; Bulley, S.M.; Chagne, D.; Marsh, K.B.; Matich, A.J.; et al. Analysis of expressed sequence tags from Actinidia: Applications of a cross species EST database for gene discovery in the areas of flavor, health, color and ripening. BMC Genom. 2008, 9, 351. [Google Scholar] [CrossRef]
- Kim, J.; Lee, J.G.; Lim, S.; Lee, E.J. A comparison of physicochemical and ripening characteristics of golden-fleshed ‘Haegeum’ and green-fleshed ‘Hayward’ kiwifruit during storage at 0 °C and ripening at 25 °C. Postharvest Biol. Technol. 2023, 196, 112166. [Google Scholar] [CrossRef]
- Gan, Z.; Shan, N.; Fei, L.; Wan, C.; Chen, J. Isolation of the 9-cis-epoxycarotenoid dioxygenase (NCED) gene from kiwifruit and its effects on postharvest softening and ripening. Sci. Hortic. 2020, 261, 109020. [Google Scholar] [CrossRef]
- Gan, Z.; Yuan, X.; Shan, N.; Wan, C.; Chen, C.; Zhu, L.; Xu, Y.; Kai, W.; Zhai, X.; Chen, J. AcERF1B and AcERF073 positively regulate indole-3-acetic acid degradation by activating AcGH3.1 transcription during postharvest kiwifruit ripening. J. Agric. Food Chem. 2021, 69, 13859–13870. [Google Scholar] [CrossRef]
- Zhang, A.D.; Zhang, Q.Y.; Li, J.H.; Gong, H.S.; Fan, X.G.; Yang, Y.Q.; Liu, X.F.; Yin, X.R. Transcriptome co-expression network analysis identifies key genes and regulators of ripening kiwifruit ester biosynthesis. BMC Plant Biol. 2020, 20, 103. [Google Scholar] [CrossRef] [PubMed]
- Souleyre, E.J.F.; Nieuwenhuizen, N.J.; Wang, M.Y.; Winz, R.A.; Matich, A.J.; Ileperuma, N.R.; Tang, H.; Baldwin, S.J.; Wang, T.C.; List, B.W.; et al. Alcohol acyl transferase genes at a high-flavor intensity locus contribute to ester biosynthesis in kiwifruit. Plant Physiol. 2022, 190, 1100–1116. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.Y.; Zheng, H.; Fan, J.T.; Liu, F.X.; Li, J.T.; Zhong, C.H.; Zhang, Q. Comparative study on physicochemical and nutritional qualities of kiwifruit varieties. Foods 2023, 12, 108. [Google Scholar] [CrossRef]
- Shi, Y.L.; Wang, M.Q.; Dong, Z.B.; Zhu, Y.; Shi, J.; Ma, W.J.; Lin, Z.; Lv, H.P. Volatile components and key odorants of Chinese yellow tea (Camellia sinensis). LWT 2021, 146, 111512. [Google Scholar] [CrossRef]
- Wang, Q.; An, X.X.; Xiang, M.L.; Chen, X.; Luo, Z.Y.; Fu, Y.Q.; Chen, M.; Chen, J.Y. Effects of 1-MCP on the physiological attributes, volatile components and ester-biosynthesis-related gene expression during storage of ‘jinyan’ kiwifruit. Horticulturae 2021, 7, 381. [Google Scholar] [CrossRef]
- Du, D.D.; Xu, M.; Wang, J.; Gu, S.; Zhu, L.Y.; Hong, X.Z. Tracing internal quality and aroma of a red-fleshed kiwifruit during ripening by means of GC-MS and E-nose. RSC Adv. 2019, 9, 21164–21174. [Google Scholar] [CrossRef]
- Li, D.P.; Xu, Y.F.; Xu, G.M.; Gu, L.K.; Li, D.Q.; Shu, H.R. Molecular cloning and expression of a gene encoding alcohol acyltransferase (MdAAT2) from apple (cv. Golden Delicious). Phytochemistry 2006, 67, 658–667. [Google Scholar] [CrossRef]
- Li, L.X.; Fang, Y.; Li, D.; Zhu, Z.H.; Zhang, Y.; Tang, Z.Y.; Li, T.; Chen, X.S.; Feng, S.Q. Transcription factors MdMYC2 and MdMYB85 interact with ester aroma synthesis gene MdAAT1 in apple. Plant Physiol. 2023, 193, 2442–2458. [Google Scholar] [CrossRef]
- Balbontin, C.; Gaete-Eastman, C.; Fuentes, L.; Figueroa, C.R.; Herrera, R.; Manriquez, D.; Latche, A.; Pech, J.C.; Alejandra Moya-Leon, M. VpAAT1, a gene encoding an alcohol acyltransferase, is involved in ester biosynthesis during ripening of mountain papaya fruit. J. Agric. Food Chem. 2010, 58, 5114–5121. [Google Scholar] [CrossRef]
- Goulet, C.; Kamiyoshihara, Y.; Lam, N.B.; Richard, T.; Taylor, M.G.; Tieman, D.M.; Klee, H.J. Divergence in the enzymatic activities of a tomato and solanum pennellii alcohol acyltransferase impacts fruit volatile ester composition. Mol. Plant 2014, 8, 153–162. [Google Scholar] [CrossRef]
- Zheng, X.; Li, Y.; Ma, C.; Chen, B.; Sun, Z.; Tian, Y.; Wang, C. A mutation in the promoter of the arabinogalactan protein 7-like gene PcAGP7-1 affects cell morphogenesis and brassinolide content in pear (Pyrus communis L.) stems. Plant J. 2022, 109, 47–63. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Donghong (μg kg−1) 1 | Ganlv-1 (μg kg−1) |
|---|---|---|
| Benzoic acid, methyl ester | 4136.01 ± 406.39 a | 4262.66 ± 544.86 a |
| 2-Butenoic acid, 3-methyl-, methyl ester | 1740.43 ± 318.42 a | 421.64 ± 63.74 b |
| Hexanoic acid, methyl ester | 63.02 ± 5.08 a | 23.68 ± 0.42 b |
| Butanoic acid, 2-pentenyl ester, (Z)- | 749.87 ± 83.03 a | 514.14 ± 47.57 b |
| Methyl salicylate | 417.93 ± 62.92 b | 1170.67 ± 88.61 a |
| Octanoic acid, methyl ester | 10.65 ± 2.91 a | 2.86 ± 0.41 b |
| 2(3H)-Furanone, 5-butyldihydro- | 291.12 ± 36.68 a | 188.24 ± 17.53 b |
| Butanoic acid, ethyl ester | 3137.28 ± 132.80 a | 13.72 ± 1.95 b |
| Formic acid, 2-phenylethyl ester | 482.59 ± 85.48 a | 80.68 ± 9.47 b |
| Hexanoic acid, butyl ester | 80.44 ± 13.85 a | 70.44 ± 1.68 a |
| Hexanoic acid, ethyl ester | 692.09 ± 88.16 a | 25.78 ± 0.91 b |
| 1-Ethylpropyl acetate | 575.46 ± 20.33 a | 1.38 ± 0.51 b |
| Benzeneacetic acid, methyl ester | 128.66 ± 24.76 a | 22.86 ± 1.36 b |
| Octanoic acid, ethyl ester | 97.54 ± 15.46 a | 46.31 ± 1.14 b |
| Butanoic acid, propyl ester | 49.84 ± 6.00 b | 70.47 ± 7.80 a |
| Pentanoic acid, ethyl ester | 96.69 ± 10.94 a | 0.48 ± 0.09 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Q.; Huang, Z.; Chen, J.; Gan, Z. Natural Variation in the AAT1 Promoter Is Responsible for the Disparity in Ester Aroma Between Actinidia chinensis and Actinidia eriantha. Agronomy 2024, 14, 2965. https://doi.org/10.3390/agronomy14122965
Cao Q, Huang Z, Chen J, Gan Z. Natural Variation in the AAT1 Promoter Is Responsible for the Disparity in Ester Aroma Between Actinidia chinensis and Actinidia eriantha. Agronomy. 2024; 14(12):2965. https://doi.org/10.3390/agronomy14122965
Chicago/Turabian StyleCao, Qing, Zhenyu Huang, Jinyin Chen, and Zengyu Gan. 2024. "Natural Variation in the AAT1 Promoter Is Responsible for the Disparity in Ester Aroma Between Actinidia chinensis and Actinidia eriantha" Agronomy 14, no. 12: 2965. https://doi.org/10.3390/agronomy14122965
APA StyleCao, Q., Huang, Z., Chen, J., & Gan, Z. (2024). Natural Variation in the AAT1 Promoter Is Responsible for the Disparity in Ester Aroma Between Actinidia chinensis and Actinidia eriantha. Agronomy, 14(12), 2965. https://doi.org/10.3390/agronomy14122965