
Effect of Nitrogen Fertilization and Inoculation of Durum Wheat with Fusarium pseudograminearum on Yield, Technological Quality and Gluten Protein Composition



Abstract
1. Introduction
2. Materials and Methods
2.1. Location and Soil Characteristics
2.2. Experimental Treatments and Growing Conditions
2.3. Grain and Semolina Analyses
2.4. Gluten Protein Composition Analysis of Harvested Grain from 2021 Trial
2.5. Statistical Analysis
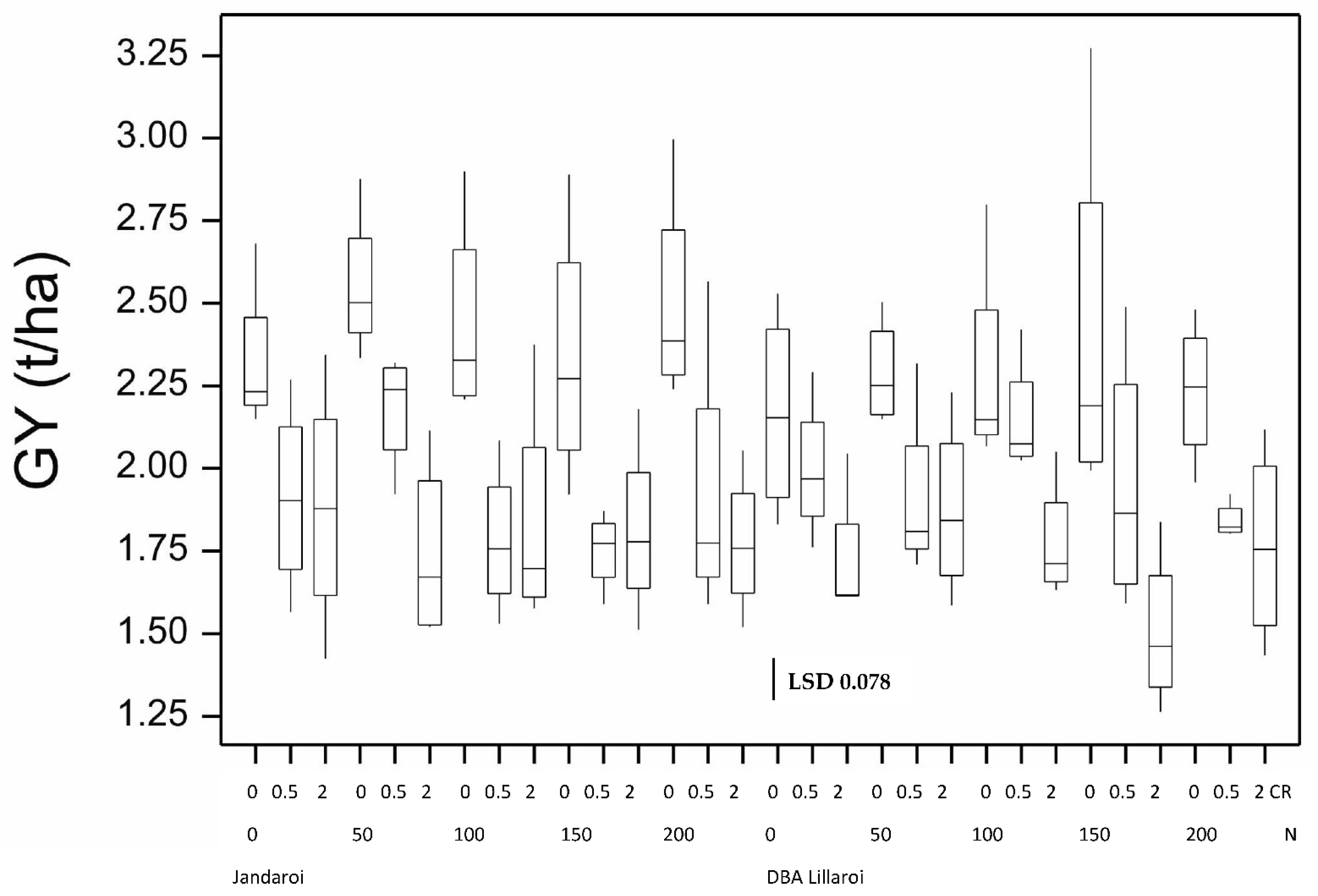
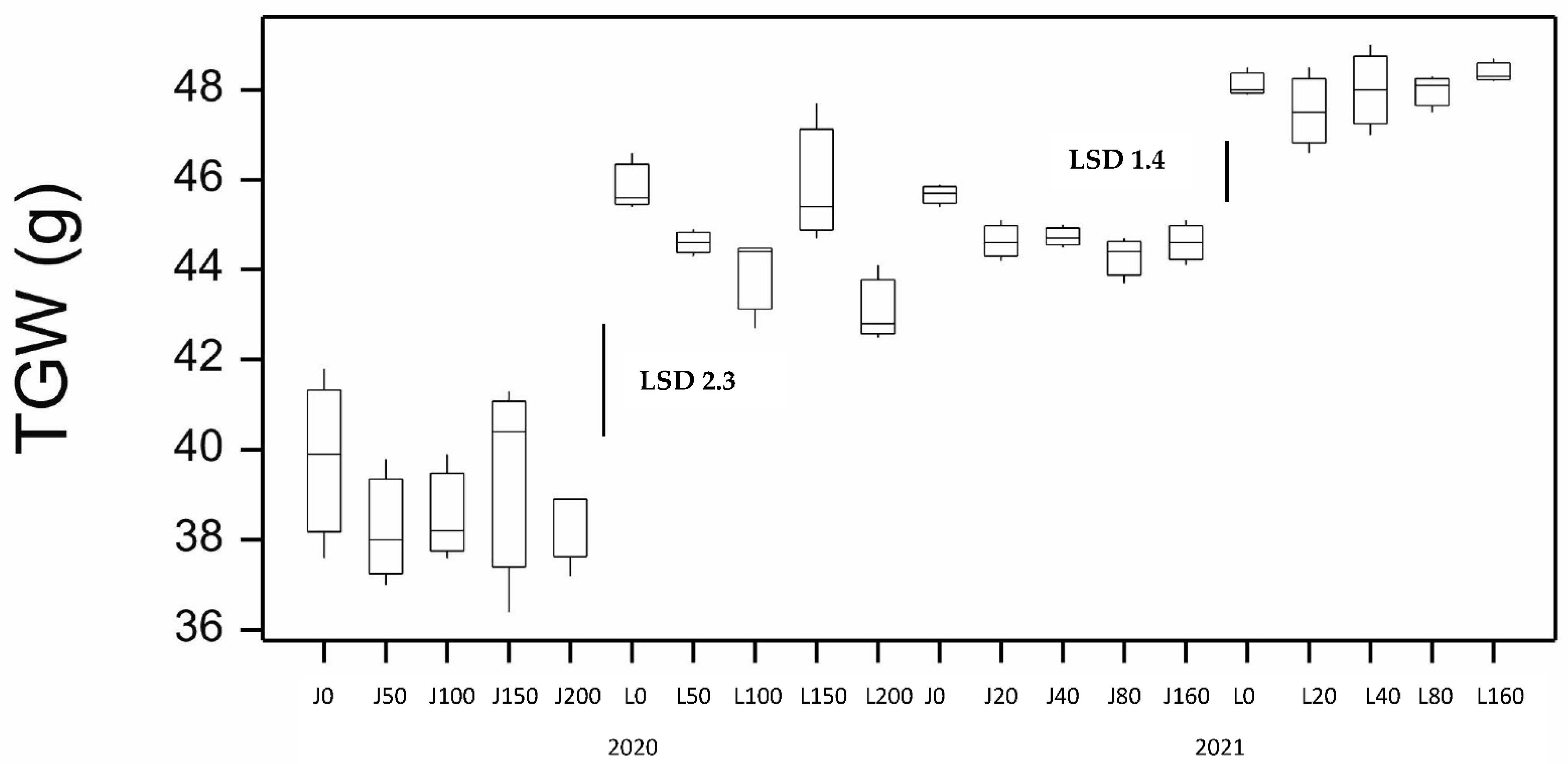
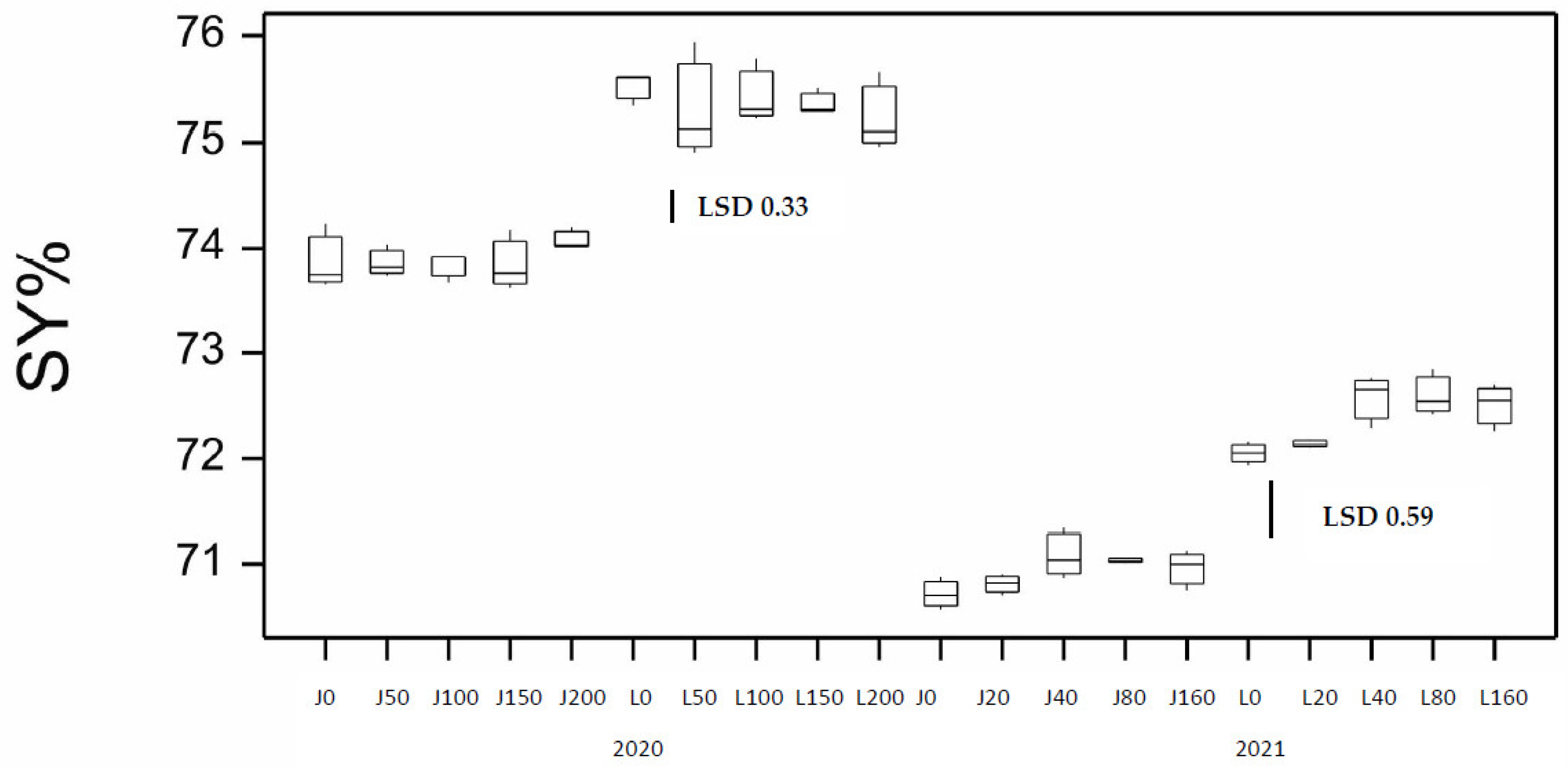
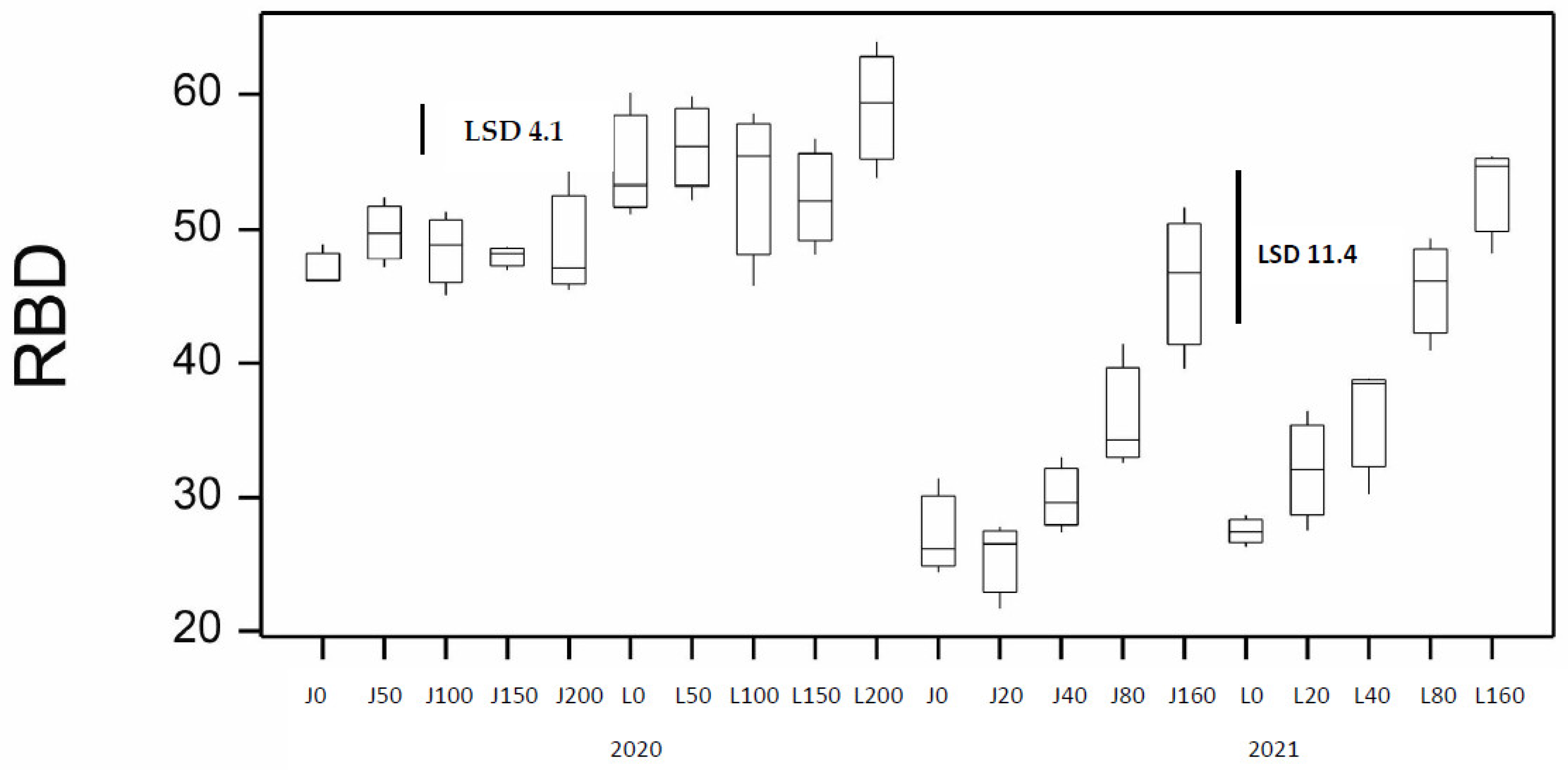
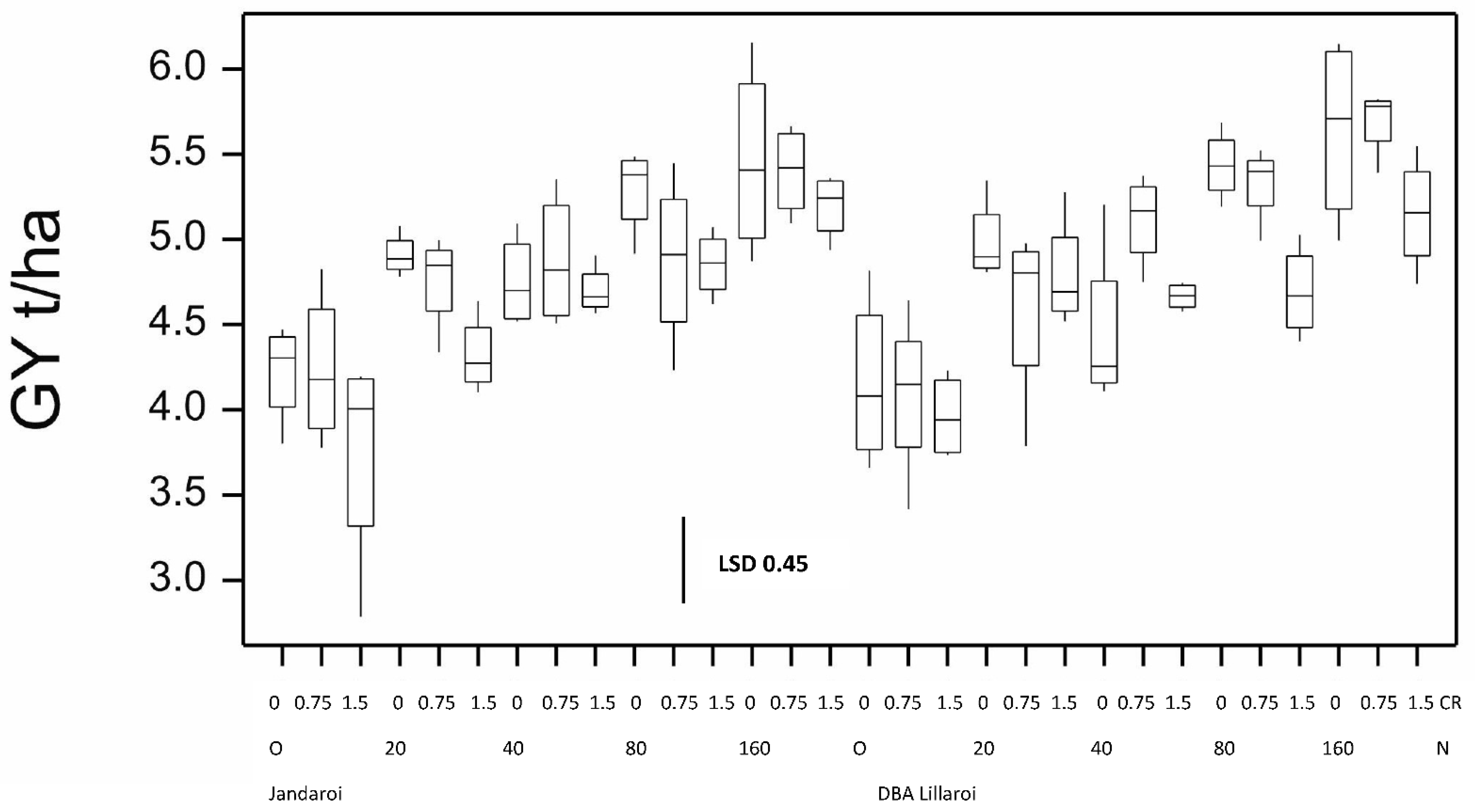
3. Results
3.1. Results from the 2020 Season
3.2. Results from the 2021 Season and Comparison to the 2020 Season
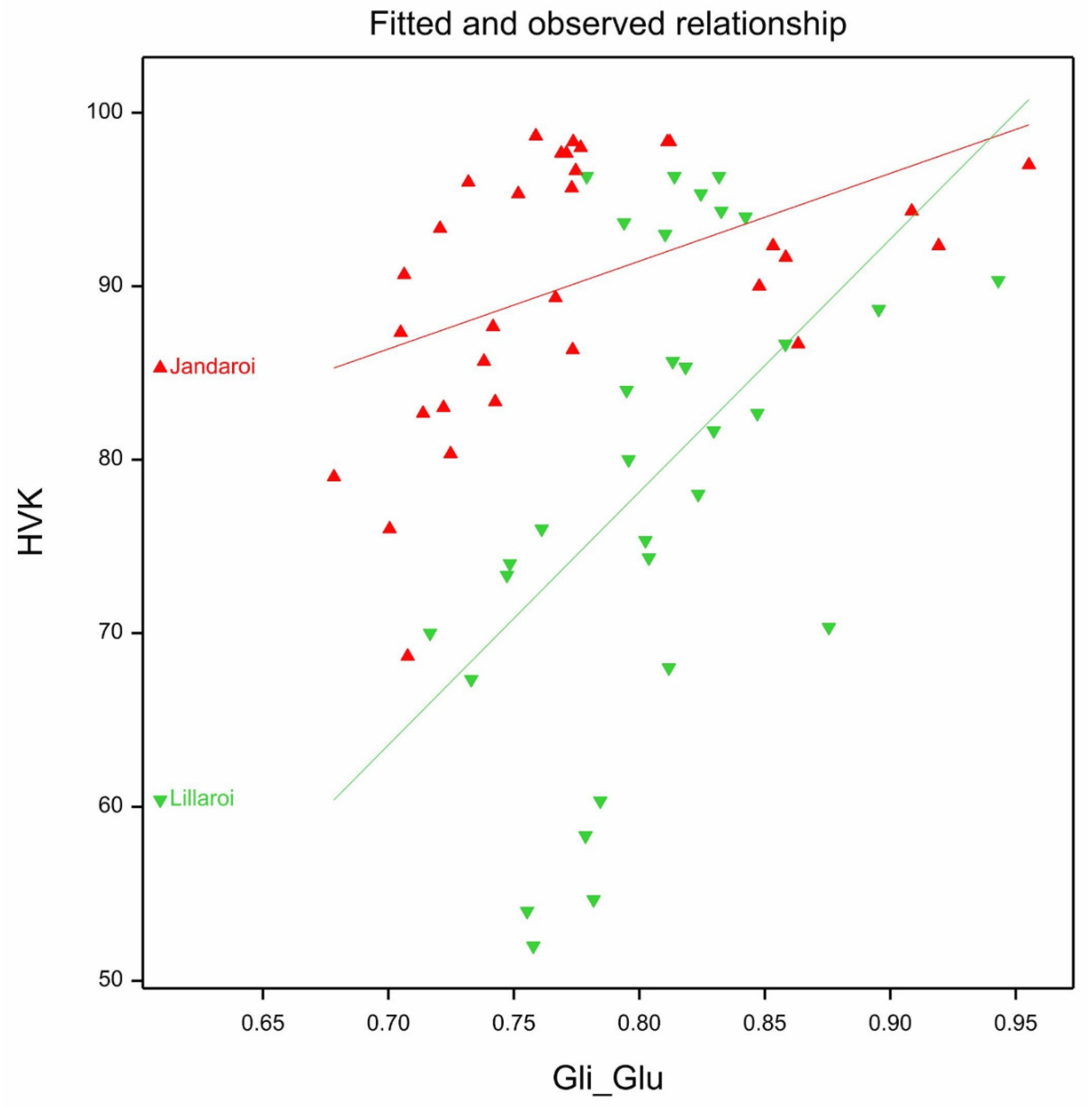
3.3. Protein Compositional Changes (2021 Season)
4. Discussion
Comparison of Seasonal Impact on Yield and Quality
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Richards, R.A.; Rebetzke, G.J.; Condon, A.G.; van Herwaarden, A.F. Breeding opportunities for increasing the efficiency of water use and crop yield in temperate cereals. Crop Sci. 2002, 42, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.X.; Wang, K.; Dupuis, B.; Taylor, D.; Nam, S. Kernel vitreousness and protein content: Relationship, interaction and synergistic effects on durum wheat quality. J. Cereal Sci. 2018, 79, 210–217. [Google Scholar] [CrossRef]
- Anonymous. Grain Trade Australia Wheat Standards 2020/21. 2020. Available online: http://www.graintrade.org.au/sites/default/files/WHEAT%20STANDARDS%202020-2021%20All%20Grades%20updated%20061120.pdf (accessed on 22 November 2022).
- Herbert, A. An international benchmarking comparison of Australian crop production and profitability. In GRDC Updates; GRDC, Ed.; GRDC: Perth, Australia, 2017. [Google Scholar]
- Chakraborty, S.; Liu, C.L.; Mitter, V.; Scott, J.B.; Akinsanmi, O.A.; Ali, S.; Dill-Macky, R.; Backhouse, D.; Simpfendorfer, S. Pathogen population structure and epidemiology are keys to wheat crown rot and Fusarium head blight management. Australas. Plant Pathol. 2006, 35, 643–655. [Google Scholar] [CrossRef]
- Simpfendorfer, S.; McKay, A.; Ophel-Keller, K. New approaches to crop disease management in conservation agriculture. In Australian Agriculture in 2020: From Conservation to Automation; Pratley, J., Kirkegaard, J., Eds.; Agronomy Australia and Charles Sturt University: Wagga Wagga, Australia, 2019; pp. 173–188. [Google Scholar]
- Alahmad, S.; Simpfendorfer, S.; Bentley, A.R.; Hickey, L.T. Crown rot of wheat in Australia: Fusarium pseudograminearum taxonomy, population biology and disease management. Australas. Plant Pathol. 2018, 47, 285–299. [Google Scholar] [CrossRef]
- Murray, G.M.; Brennan, J.P. Estimating disease losses to the Australian wheat industry. Australas. Plant Pathol. 2009, 38, 558–570. [Google Scholar] [CrossRef]
- Petronaitis, T.; Simpfendorfer, S.; Hüberli, D. Importance of Fusarium spp. in Wheat to Food Security: A Global Perspective. In Plant Diseases and Food Security in the 21st Century; Scott, P., Strange, R., Korsten, L., Gullino, M.L., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 127–159. [Google Scholar]
- Evans, M.L.; Hollaway, G.J.; Dennis, J.L.; Correll, R.; Wallwork, H. Crop sequence as a tool for managing populations of Fusarium pseudograminearum and F. culmorum in south-eastern Australia. Australas. Plant Pathol. 2010, 39, 376–382. [Google Scholar] [CrossRef]
- Kirkegaard, J.A.; Simpfendorfer, S.; Holland, J.; Bambach, R.; Moore, K.J.; Rebetzke, G.J. Effect of previous crops on crown rot and yield of durum and bread wheat in northern NSW. Aust. J. Agric. Res. 2004, 55, 321–334. [Google Scholar] [CrossRef]
- Verrell, A.G.; Simpfendorfer, S.; Moore, K.J. Effect of row placement, stubble management and ground engaging tool on crown rot and grain yield in a no-till continuous wheat sequence. Soil Tillage Res. 2017, 165, 16–22. [Google Scholar] [CrossRef]
- Forknall, C.R.; Simpfendorfer, S.; Kelly, A.M. Using yield response curves to measure variation in the tolerance and resistance of wheat cultivars to Fusarium crown rot. Phytopathology 2019, 109, 932–941. [Google Scholar] [CrossRef]
- Davis, R.A.; Huggins, D.R.; Cook, J.R.; Paulitz, T.C. Nitrogen and crop rotation effects on fusarium crown rot in no-till spring wheat. Can. J. Plant Pathol. 2009, 31, 456–467. [Google Scholar] [CrossRef]
- Buster, M.; Simpfendorfer, S.; Guppy, C.; Sissons, M.; Flavel, R.J. Interactions of Fusarium crown rot of wheat with nitrogen. Plants 2023, 12, 533. [Google Scholar] [CrossRef]
- Hughes, N.; Galeano, D.; Hatfield-Dodds, S. The Effects of Drought and Climate Variability on Australian Farms; Australian Bureau of Agricultural and Resource Economics and Sciences: Canberra, Australia, 2019. [CrossRef]
- Zörb, C.; Ludewig, U.; Hawkesford, M.J. Perspective on wheat yield and quality with reduced nitrogen supply. Trends Plant Sci. 2018, 23, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Ophel-Keller, K.; McKay, A.; Hartley, D.; Herdina; Curran, J. Development of a routine DNA-based testing service for soilborne diseases in Australia. Australas. Plant Pathol. 2008, 37, 243–253. [Google Scholar] [CrossRef]
- Sissons, M.; Ovenden, B.; Adorada, D.; Milgate, A. Durum wheat quality in high input irrigation systems in south-eastern Australia. Crop Pasture Sci. 2014, 65, 411–422. [Google Scholar] [CrossRef]
- Sissons, M.J.; Batey, I.L. Protein and starch properties of some tetraploid wheats. Cereal Chem. 2003, 80, 468–475. [Google Scholar] [CrossRef]
- Hollaway, G.J.; Evans, M.L.; Wallwork, H.; Dyson, C.B.; McKay, A.C. Yield loss in cereals, caused by Fusarium culmorum and F. pseudograminearum, is related to fungal DNA in soil prior to planting, rainfall, and cereal type. Plant Dis. 2013, 97, 977–982. [Google Scholar] [CrossRef]
- Fuertes-Mendizabal, T.A.; Aizpurua, M.B.; Gonzalez-Moro, J.M.; Estavillo, J.M. Improving wheat breadmaking quality by splitting the N fertilizer rate. Euro. J. Agron. 2010, 33, 52–61. [Google Scholar] [CrossRef]
- Johansson, V.A.; Mattern, P.J. Wheat, rye, and triticale. In Nutritional Quality of Cereal Grains; Olson, R.A., Frey, K.J., Eds.; ASA: Branchburg, NJ, USA; CSSA: Little Rock, ML, USA; SSSA: Madison, WI, USA, 1987; Volume 28, pp. 133–182. [Google Scholar]
- Reznick, J.P.K.; Barth, G.; Kaschuk, G.; Pauletti, V. Nitrogen and cultivars as field strategies to improve the nutritional status of wheat grain and flour. J. Cereal Sci. 2021, 102, 103290. [Google Scholar] [CrossRef]
- Giuliani, M.M.; Giuzio, L.; De Caro, A.; Flagella, Z. Relationships between nitrogen utilization and grain technological quality in durum wheat: II. Grain yield and quality. Agron. J. 2011, 103, 1668–1675. [Google Scholar] [CrossRef]
- Ehdaie, B.; Waines, J.G. Sowing date and nitrogen rate effects on dry matter and nitrogen partitioning in bread and durum wheat. Field Crops Res. 2001, 73, 47–61. [Google Scholar] [CrossRef]
- Martre, P.; Porter, J.R.; Jamieson, P.D.; Triboi, E. Modelling grain nitrogen accumulation and protein composition to understand the sink/source regulation of nitrogen remobilization for wheat. Plant Phys. 2003, 133, 1959–1967. [Google Scholar] [CrossRef] [PubMed]
- Clarke, J.M.; Clarke, F.R.; Pozniak, C.J. Forty-six years of genetic improvement in Canadian durum wheat cultivars. Can. J. Plant Sci. 2010, 90, 791–801. [Google Scholar] [CrossRef]
- DePauw, R.M.; Knox, R.E.; Clarke, F.R.; Wang, H.; Fernandez, M.R.; Clarke, J.M.; McCaig, T.N. Shifting undesirable correlations. Euphytica 2007, 157, 409–415. [Google Scholar] [CrossRef]
- Motzo, R.; Fois, S.; Giunta, F. Relationship between grain yield and quality of durum wheats from different eras of breeding. Euphytica 2004, 140, 147–154. [Google Scholar] [CrossRef]
- Sissons, M.; Kadkol, G.; Taylor, J. Genotype by environment effects on durum wheat quality and yield implications for breeding. Crop Breed. Genet. Genomics. 2020, 2, e200018. [Google Scholar]
- Kadkol, G.; Sissons, M.; Lambert, N.; Lisle, C. Genetic improvement in grain yield and quality of Australian durum wheat over six decades of breeding. Cereal Chem. 2022, 100, 109–130. [Google Scholar] [CrossRef]
- Dexter, J.E.; D’Egidio, M.G. Grading factors impacting durum wheat processing quality. In Durum Wheat Chemistry and Technology, 2nd ed.; Sissons, M.S., Abecassis, J., Marchylo, B., Carcea, M., Eds.; AACC International Press: St. Paul, MN, USA, 2012; pp. 235–250. [Google Scholar]
- Dexter, J.E.; Williams, P.C.; Edwards, N.M.; Martin, D.G. The relationships between durum wheat vitreousness, kernel hardness and processing quality. J. Cereal Sci. 1988, 7, 169–181. [Google Scholar] [CrossRef]
- Samson, M.-F.; Mabille, F.; Cheret, R.; Abecassis, J.; Morel, M.H. Mechanical and physicochemical characterization of vitreous and mealy durum wheat endosperm. Cereal Chem. 2005, 82, 81–87. [Google Scholar] [CrossRef]
- Boukef, S.; Karmous, C.; Trifa, Y.; Rezgui, S. Durum wheat grain quality traits as affected by nitrogen fertilization sources under mediterranean rainfed conditions. J. Agric. Sustain. 2013, 4, 99–114. [Google Scholar]
- Sieber, A.N.; Würschum, T.; Longin, C.G.H. Vitreosity, its stability and relationship to protein content in durum wheat. J. Cereal Sci. 2015, 61, 71–77. [Google Scholar] [CrossRef]
- Wang, K.; Taylor, D.; Chen, Y.; Suchy, J.; Fu, B.X. Effect of kernel size and its potential interaction with genotype on key quality traits of durum wheat. Foods 2021, 10, 2992. [Google Scholar] [CrossRef] [PubMed]
- Sissons, M.; Abecassis, J.; Cubadda, R.; Marchylo, B. Methods used to assess and predict quality of durum wheat, semolina and pasta. In Durum Wheat Chemistry and Technology, 2nd ed.; Sissons, M.S., Abecassis, J., Marchylo, B., Carcea, M., Eds.; AACC International Press: St. Paul, MN, USA, 2012; pp. 213–234. [Google Scholar]
- Ficco, D.B.M.; Mastrangelo, A.M.; Trono, D.; Borrelli, G.M.; Vita, P.D.; Fares, C.; Beleggia, R.; Platani, C.; Papa, R. The colours of durum wheat: A review. Crop Pasture Sci. 2014, 65, 1–15. [Google Scholar] [CrossRef]
- Feillet, P.; Autran, J.C.; Icard-Verniere, C. Pasta brownness: An assessment. J. Cereal Sci. 2000, 32, 215–233. [Google Scholar] [CrossRef]
- Dalla Marta, A.D.; Grifoni, M.; Mancini, G.; Zipoli, S.; Orlandini, S. The influence of climate on durum wheat quality in Tuscany, central Italy. Intern. J. Biometeorol. 2011, 55, 87–96. [Google Scholar] [CrossRef]
- Fois, S.; Schlichting, L.; Marchylo, B.; Dexter, J.; Motzoa, R.; Giuntaa, F. Environmental conditions affect semolina quality in durum wheat (Triticum turgidum ssp. durum L.) cultivars with different gluten strength and gluten protein composition. J. Sci. Food Agric. 2011, 91, 2664–2673. [Google Scholar] [CrossRef]
- Saint Pierre, C.; Peterson, C.J.; Ross, A.S.; Ohm, J.B.; Verhoeven, M.C.; Larson, M.; Hoefer, B. Winter wheat cultivars under different levels of nitrogen and water stress: Changes in grain protein composition. J. Cereal Sci. 2008, 47, 407–416. [Google Scholar] [CrossRef]
- Barak, S.; Mudgil, D.; Khatkar, B.S. Influence of gliadin and glutenin fractions on rheological, pasting, and textural properties of dough. Intern. J. Food Prop. 2014, 17, 1428–1438. [Google Scholar] [CrossRef]
- Tang, J.W.; Liu, J.J.; Zhang, P.P.; Zhang, Y.; Xiao, Y.G.; Qu, Y.Y.; Zhang, Y.; He, Z.H. Effects of gluten protein fractions on dough property and products quality in common wheat. Sci. Agric. Sin. 2008, 41, 2937–2946. [Google Scholar]
- AbuHammad, W.A.; Elias, E.M.; Manthey, F.A.; Alamri, M.S.; Mergoum, M. A comparison of methods for assessing dough and gluten strength of durum wheat and their relationship to pasta cooking quality. Intern. J. Food Sci. Tech. 2012, 47, 2561–2673. [Google Scholar] [CrossRef]
- Horvat, D.; Šimić, G.; Dvojković, K.; Ivić, M.; Plavšin, I.; Novoselović, D. Gluten protein compositional changes in response to nitrogen application rate. Agronomy 2021, 11, 325. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metric | January | February | March | April | May | June | July | August | September | October | November | December | Annual |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rainfall 2020 | 97.4 | 202.2 | 83.4 | 49.0 | 39.2 | 29.4 | 37.0 | 29.2 | 26.8 | 73.6 | 3.0 | 223.6 | 893.8 |
| Rainfall 2021 | 23.6 | 108.2 | 162.0 | 22.6 | 24.6 | 125.0 | 65.4 | 48.4 | 36.8 | 52.0 | 191.0 | 70.6 | 930.2 |
| Rainfall LTA | 61.2 | 73.9 | 57.2 | 25.3 | 30.4 | 53.4 | 40.9 | 39.5 | 46.3 | 58.3 | 82.3 | 80.4 | 638.5 |
| Min T 2020 | 21.4 | 19.0 | 14.4 | 9.8 | 5.1 | 3.7 | 3.5 | 2.9 | 6.6 | 10.3 | 13.3 | 17.0 | |
| Min T 2021 | 15.3 | 15.7 | 14.4 | 7.0 | 4.1 | 3.7 | 2.9 | 3.4 | 4.1 | 8.7 | 14.0 | 14.7 | |
| Min T LTA | 17.7 | 16.9 | 14.5 | 10.0 | 6.0 | 3.6 | 2.3 | 2.8 | 5.8 | 9.7 | 13.3 | 15.7 | |
| Max T 2020 | 36.3 | 29.7 | 27.5 | 24.1 | 18.9 | 17.5 | 16.9 | 17.6 | 23.3 | 27.1 | 31.8 | 30.4 | |
| Max T 2021 | 30.8 | 29.9 | 27.4 | 23.9 | 20.5 | 16.8 | 16.6 | 19.9 | 22.1 | 25.4 | 25.9 | 29.9 | |
| Max T LTA | 33.0 | 31.5 | 29.2 | 25.5 | 20.8 | 17.0 | 16.5 | 18.5 | 22.0 | 25.6 | 28.6 | 30.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sissons, M.; Egan, N.; Simpfendorfer, S. Effect of Nitrogen Fertilization and Inoculation of Durum Wheat with Fusarium pseudograminearum on Yield, Technological Quality and Gluten Protein Composition. Agronomy 2023, 13, 1658. https://doi.org/10.3390/agronomy13061658
Sissons M, Egan N, Simpfendorfer S. Effect of Nitrogen Fertilization and Inoculation of Durum Wheat with Fusarium pseudograminearum on Yield, Technological Quality and Gluten Protein Composition. Agronomy. 2023; 13(6):1658. https://doi.org/10.3390/agronomy13061658
Chicago/Turabian StyleSissons, Mike, Narelle Egan, and Steven Simpfendorfer. 2023. "Effect of Nitrogen Fertilization and Inoculation of Durum Wheat with Fusarium pseudograminearum on Yield, Technological Quality and Gluten Protein Composition" Agronomy 13, no. 6: 1658. https://doi.org/10.3390/agronomy13061658
APA StyleSissons, M., Egan, N., & Simpfendorfer, S. (2023). Effect of Nitrogen Fertilization and Inoculation of Durum Wheat with Fusarium pseudograminearum on Yield, Technological Quality and Gluten Protein Composition. Agronomy, 13(6), 1658. https://doi.org/10.3390/agronomy13061658