

_Qin.png)
Synergistic Effects of Subsoil Calcium in Conjunction with Nitrogen on the Root Growth and Yields of Maize and Soybeans in a Tropical Cropping System


Abstract
1. Introduction
2. Materials and Methods
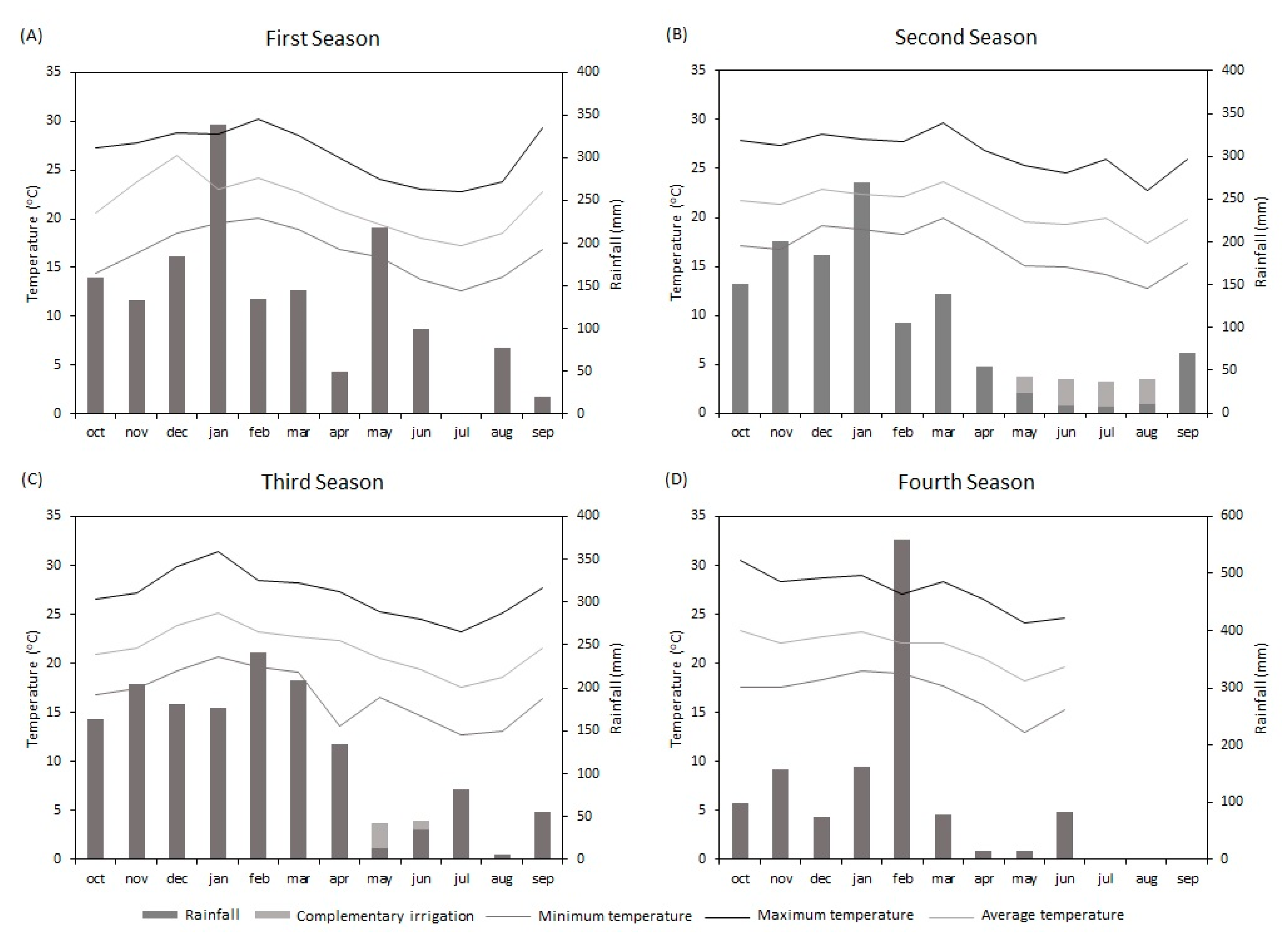
2.1. Site Description
2.2. Experimental Design and Treatments
2.3. Experiment Management
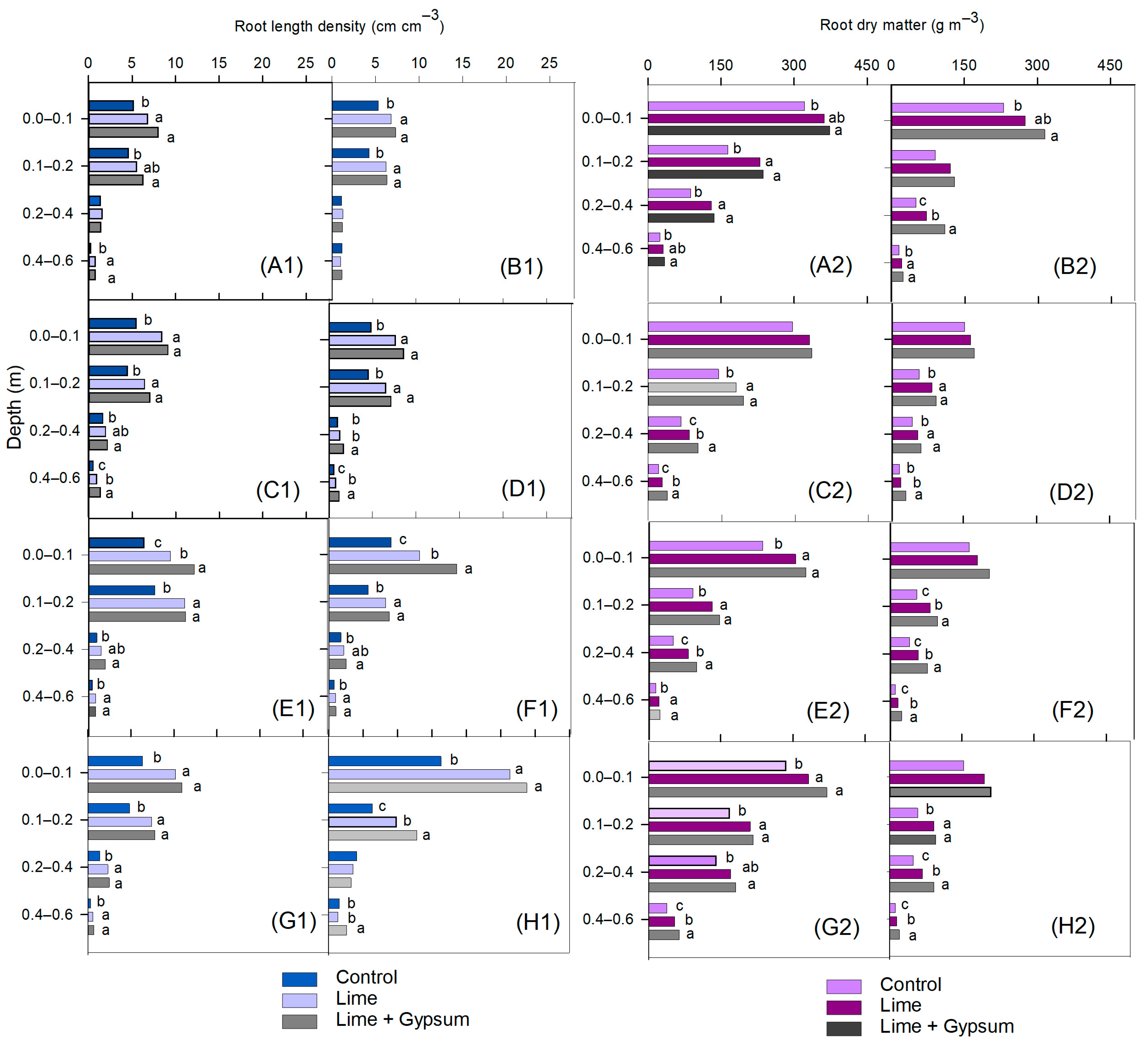
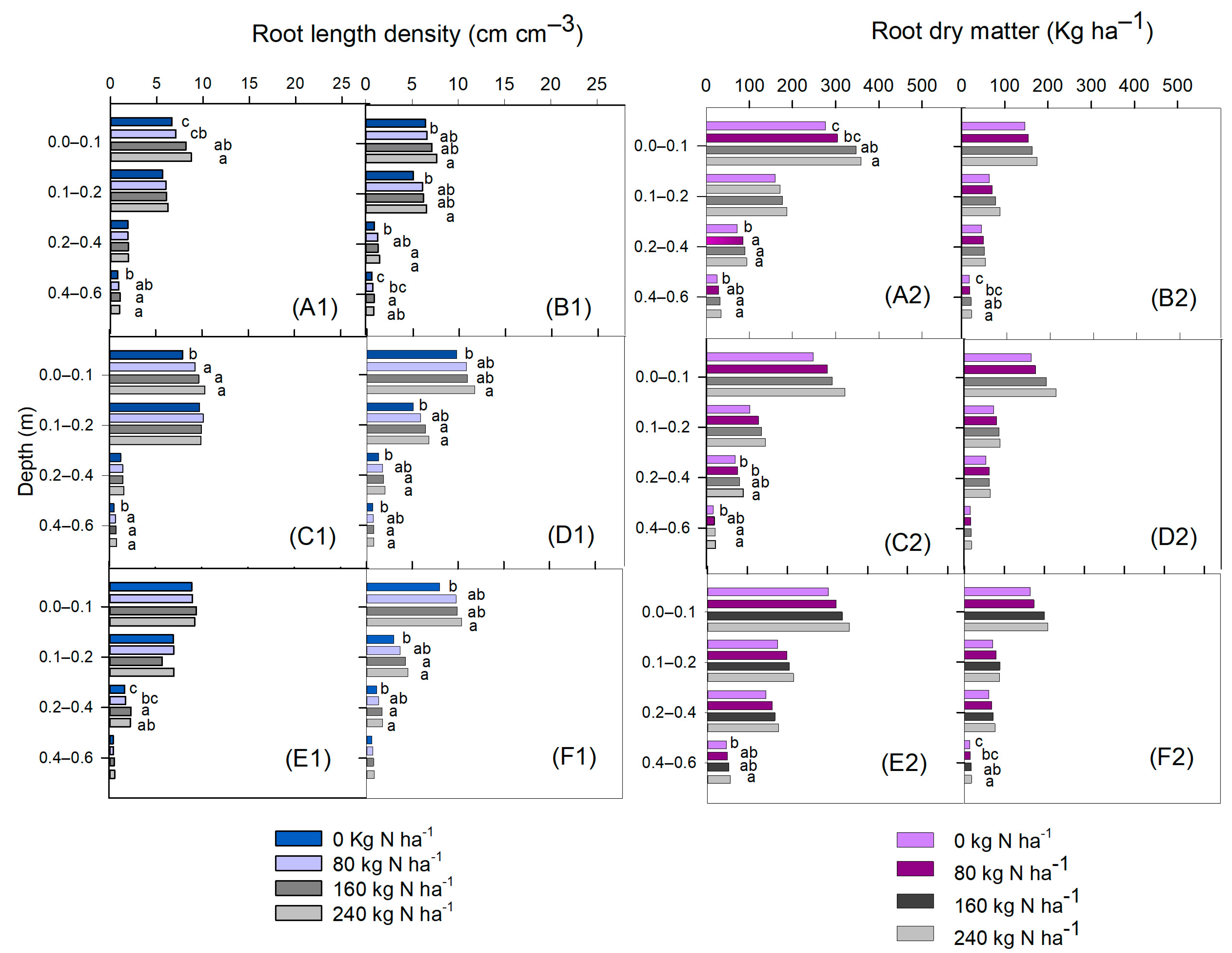
2.4. Root Sampling and Dry Matter Determination
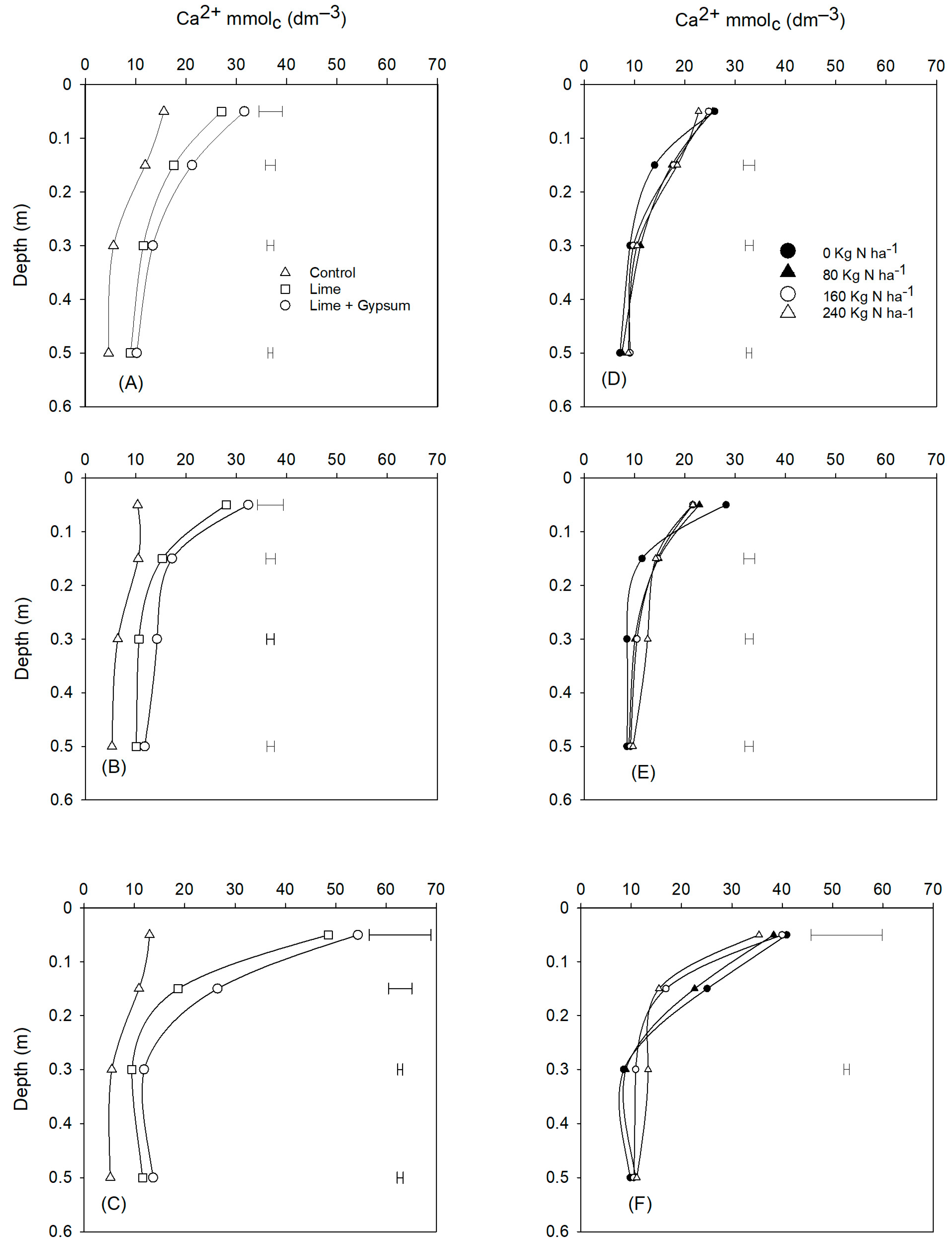
2.5. Soil Sampling and Chemical Analysis
2.6. Statistical Analysis
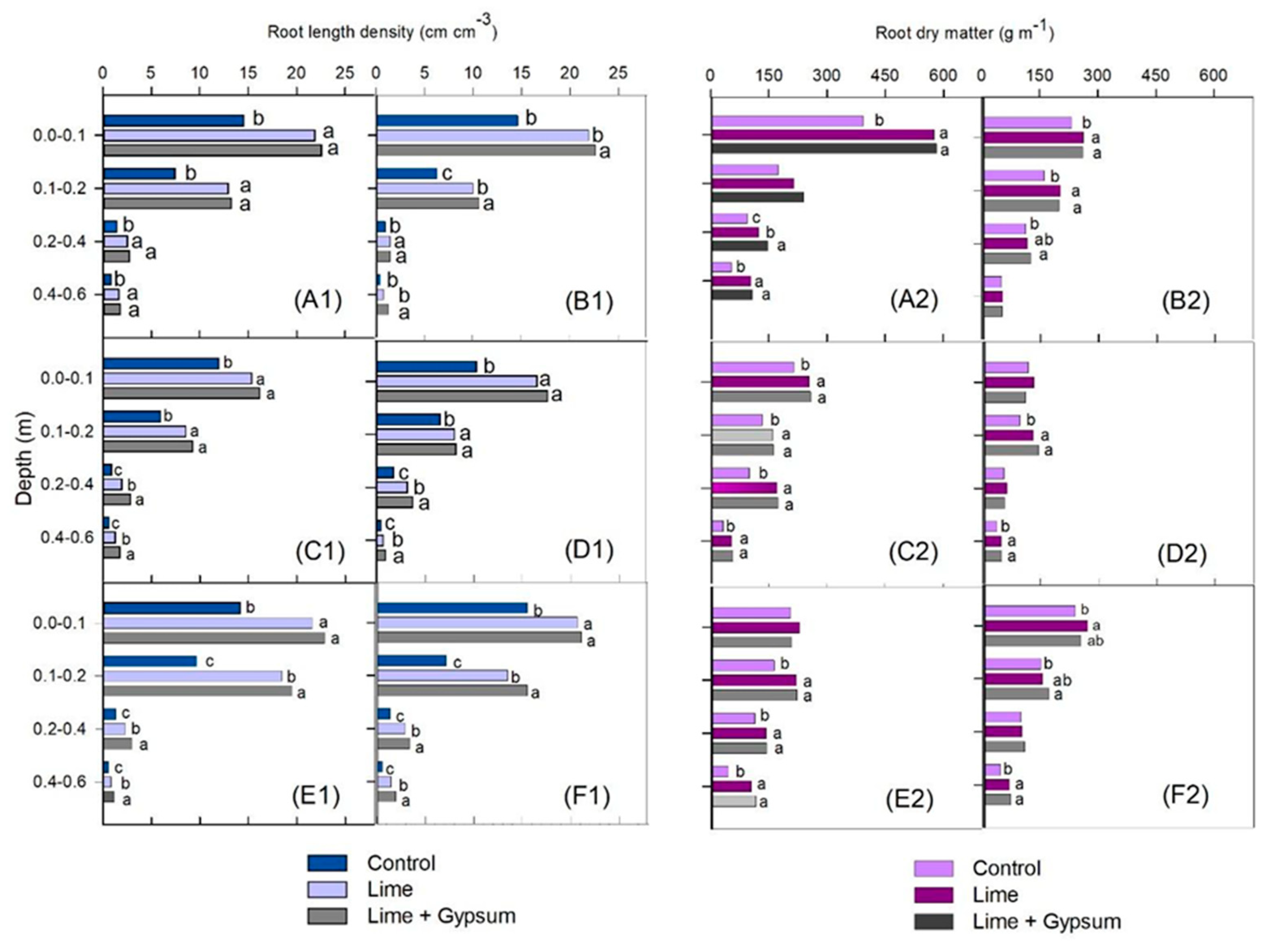
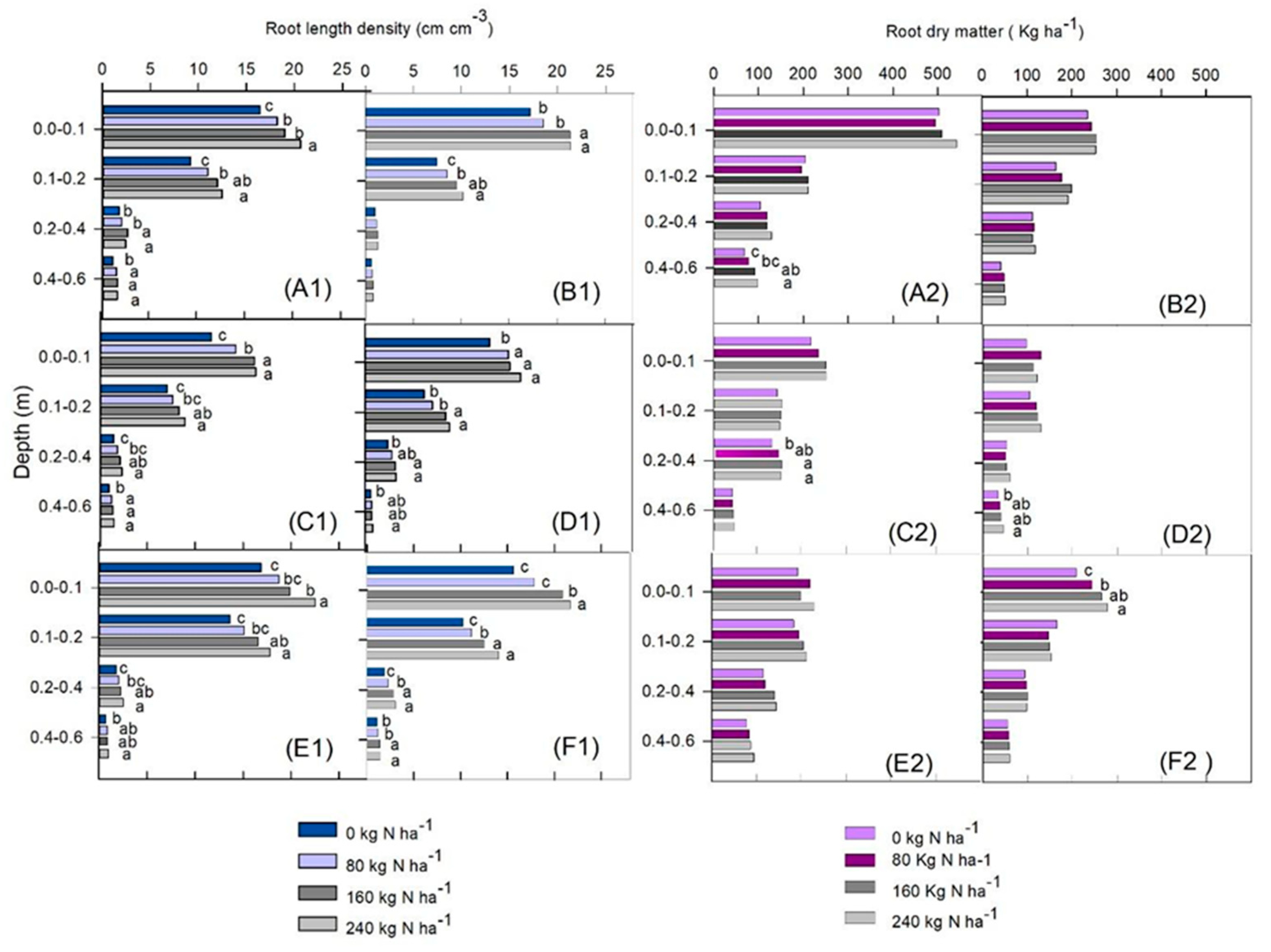
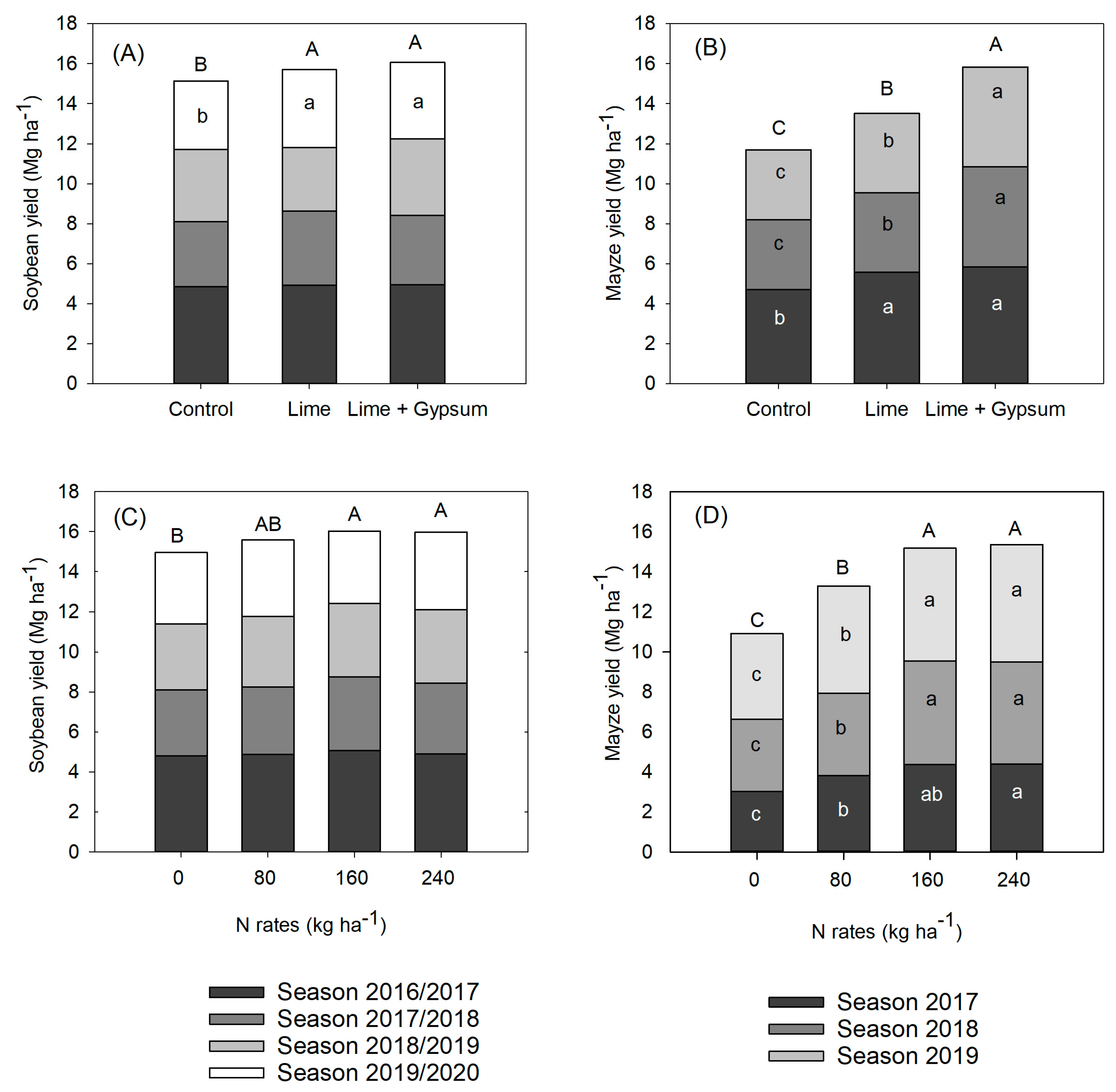
3. Results
4. Discussion
Grain Yield
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jones, D.L.; Ryan, P.R. Aluminum Toxicity. In Encyclopedia of Applied Plant Sciences; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar] [CrossRef]
- Ritchey, D.K.; Feldhake, C.M.; Clark, R.B.; de Sousa, D.M.G. Improved Water and Nutrient Uptake from Subsurface Layers of Gypsum-Amended Soils. In Agricultural Utilization of Urban and Industrial By-Products; Karlen, D., Wright, R., Kemper, W., Eds.; ACS Special Publication: Washington, DC, USA, 1995. [Google Scholar] [CrossRef]
- Rosolem, C.A.; Ritz, K.; Cantarella, H.; Galdos, M.V.; Hawkesford, M.J.; Whalley, W.R.; Mooney, S.J. Enhanced Plant Rooting and Crop System Management for Improved N Use Efficiency. Adv. Agron. 2017, 146, 205–239. [Google Scholar] [CrossRef]
- Caires, E.F.; Garbuio, F.J.; Alleoni, L.R.F.; Cambri, M.A. Calagem superficial e cobertura de aveia preta antecedendo os cultivos de milho e soja em sistema plantio direto. Rev. Bras. Ciência Solo 2006, 30, 87–98. [Google Scholar] [CrossRef]
- Caires, E.F.; Joris, H.A.W.; Churka, S. Long-term effects of lime and gypsum additions on no-till corn and soybean yield and soil chemical properties in southern Brazil. Soil Use Manag. 2011, 27, 45–53. [Google Scholar] [CrossRef]
- Costa, A.; Rosolem, C.A. Liming in the transition to no-till under a wheat–soybean rotation. Soil Tillage Res. 2007, 97, 207–217. [Google Scholar] [CrossRef]
- Raij, B.V. Reações de Gesso em Solos Ácidos. In Seminário Sobre o Uso do Gesso na Agricultura; Ibrafos: Uberaba, Brazil, 1992; pp. 105–120. [Google Scholar]
- Pivetta, L.A.; Castoldi, G.; Pivetta, L.G.; Maia, S.C.M.; Rosolem, C.A. Gypsum application, soil fertility and cotton root growth. Bragantia 2019, 78, 264–273. [Google Scholar] [CrossRef]
- Galdos, M.V.; Brown, E.; Rosolem, C.A.; Pires, L.F.; Hallett, P.D.; Mooney, S.J. Brachiaria species influence nitrate transport in soil by modifying soil structure with their root system. Sci. Rep. 2020, 10, 5072. [Google Scholar] [CrossRef] [PubMed]
- Maeght, J.-L.; Rewald, B.; Pierret, A. How to study deep roots—And why it matters. Front. Plant Sci. 2013, 4, 299. [Google Scholar] [CrossRef]
- Jing, J.; Rui, Y.; Zhang, F.; Rengel, Z.; Shen, J. Localized application of phosphorus and ammonium improves growth of maize seedlings by stimulating root proliferation and rhizosphere acidification. Field Crops Res. 2010, 119, 355–364. [Google Scholar] [CrossRef]
- Shu, L.; Shen, J.; Rengel, Z.; Tang, C.; Zhang, F. Cluster Root Formation by Lupinus Albus is Modified by Stratified Application of Phosphorus in a Split-Root System. J. Plant Nutr. 2007, 30, 271–288. [Google Scholar] [CrossRef]
- Williamson, L.C.; Ribrioux, S.P.C.P.; Fitter, A.H.; Leyser, H.M.O. Phosphate Availability Regulates Root System Architecture in Arabidopsis. Plant Physiol. 2001, 126, 87–882. [Google Scholar] [CrossRef]
- Souza, E.A.; Ferreira-Eloy, N.R.; Grassmann, C.S.; Rosolem, C.A.; White, P.J. Ammonium improves corn phosphorus acquisition through changes in the rhizosphere processes and root morphology. Pedosphere 2019, 29, 534–539. [Google Scholar] [CrossRef]
- Farina, M.P.W.; Channon, P.; Thibaud, G.R. A Comparison of Strategies for Ameliorating Subsoil Acidity I. Long-Term Growth Effects. Soil Sci. Soc. Am. J. 2000, 64, 646–651. [Google Scholar] [CrossRef]
- Sumner, M.E. Amelioration of Subsoil Acidity with Minimum Disturbance. In Advances in Soil Science: Subsoil Management Techniques; Jayawardane, N.S., Stewart, B.A., Eds.; Lewis Publishers: Boca Raton, FL, USA, 1995; pp. 147–185. [Google Scholar]
- Nora, D.D.; Amado, T.J.; Nicoloso, R.D.; Gruhn, E.M. Modern High-Yielding Maize, Wheat and Soybean Cultivars in Response to Gypsum and Lime Application on No-Till Oxisol. Rev. Bras. Ciência Solo 2017, 41, 1–21. [Google Scholar] [CrossRef]
- Tiecher, T.; Pias, O.H.; Bayer, C.; Martins, A.P.; Denardin, L.G.; Anghinoni, I. Crop Response to Gypsum Application to Subtropical Soils Under No-Till in Brazil: A Systematic Review. Rev. Bras. Ciência Solo 2018, 42, 1–17. [Google Scholar] [CrossRef]
- Rocha, K.F.; de Souza, M.; Almeida, D.S.; Chadwick, D.R.; Jones, D.L.; Mooney, S.J.; Rosolem, C.A. Cover crops affect the partial nitrogen balance in a maize-forage cropping system. Geoderma 2020, 360, 114000. [Google Scholar] [CrossRef]
- Hungria, M.; Franchini, J.C.; Campo, R.J.; Crispino, C.C.; Moraes, J.Z.; Sibaldelli, R.N.R.; Mendes, I.C.; Arihara, J. Nitrogen nutrition of soybean in Brazil: Contributions of biological N2 fixation and N fertilizer to grain yield. Can. J. Plant Sci. 2006, 86, 927–939. [Google Scholar] [CrossRef]
- Salvagiotti, F.; Cassman, K.G.; Specht, J.E.; Walters, D.T.; Weiss, A.; Dobermann, A. Nitrogen uptake, fixation and response to fertilizer N in soybeans: A review. Field Crops Res. 2008, 108, 1–13. [Google Scholar] [CrossRef]
- USDA. Keys to Soil Taxonomy, 12th ed.; Soil Survey Staff: Washington, DC, USA, 2014. [Google Scholar]
- Van Raij, B.; Quaggio, J.A.; Da Silva, N.M. Extraction of phosphorus, potassium, calcium and magnesium from soils by an ion-exchange resin procedure. Commun. Soil Sci. Plant Anal. 1986, 17, 544–566. [Google Scholar] [CrossRef]
- Duarte, A.P.; Cantarella, H.; Quaggio, J.A. Milho (Zea Mays). In Boletim 100: Recomendações de adubação e Calagem para o Estado de São Paulo; Cantarella, H., Quaggio, J.A., Mattos, D., Jr., Boaretto, R.M., Van Raij, B., Eds.; Instituto Agronomico (IAC): Campinas, SP, Brazil, 2022; pp. 199–205. [Google Scholar]
- Fehr, W.R.; Caviness, C.E.; Burmood, D.T.; Pennington, J.S. Stage of Development Descriptions for Soybeans, Glycine Max (L.) Merrill 1. Crop Sci. 1971, 11, 929–931. [Google Scholar] [CrossRef]
- Tennant, D. A Test of a Modified Line Intersect Method of Estimating Root Length. J. Ecol. 1975, 63, 995–1001. [Google Scholar] [CrossRef]
- Rosolem, C.A.; Pace, L.; Crusciol, C.A.C. Nitrogen management in maize cover crop rotations. Plant Soil 2004, 264, 261–271. [Google Scholar] [CrossRef]
- Rosolem, C.A.; Foloni, J.S.; Oliveira, R.H. Dinâmica do nitrogênio no solo em razão da calagem e adubação nitrogenada, com palha na superfície. Pesqui. Agropecuária Bras. 2003, 38, 301–309. [Google Scholar] [CrossRef]
- Pearson, R.W.; Abruna, F.; Vicente-Chandler, J. Effect of lime and nitrogen applications on downward movement of calcium and magnesium in two humid tropical soils of puerto rico. Soil Sci. 1962, 93, 77–82. [Google Scholar] [CrossRef]
- Hanson, J.B. The functions of calcium in plant nutrition. Adv. Plant Nutr. 1984, 149–208. [Google Scholar]
- Ritchey, K.D.; Silva, J.R.; Costa, U.F. Calcium deficiency in clayey B horizons of savanna oxisols. Soil Sci. 1982, 133, 378–382. [Google Scholar] [CrossRef]
- Rosolem, C.A.; Bicudo, S.J.; Marubayashi, O.M. Soybean Yield and Root Growth as Affected by Lime Rate and Quality. In Plant-Soil Interactions at Low pH: Principles and Management; Date, R.A.A., Grundon, N.J., Rayment, G.E., Probert, M.E., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995; pp. 543–548. [Google Scholar]
- Rosolem, C.A.; Vale, L.S.R.; Grassi Filho, H.; Moraes, N.H. Sistema radicular e nutrição do milho em função da calagem e da compactação do solo. Rev. Bras. Ciência Solo 1994, 18, 491–497. [Google Scholar]
- Chen, Y.H.; Kao, C.H. Calcium is involved in nitric oxide- and auxin-induced lateral root formation in rice. Protoplasma 2012, 249, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Hasenstein, K.-H.; Evans, M.L. Calcium Dependence of Rapid Auxin Action in Maize Roots. Plant Physiol. 1986, 81, 439–443. [Google Scholar] [CrossRef]
- Sun, C.-H.; Yu, J.-Q.; Hu, D.-G. Nitrate: A Crucial Signal during Lateral Roots Development. Front. Plant Sci. 2017, 8, 485. [Google Scholar] [CrossRef]
- Muday, G.K.; Haworth, P. Tomato root growth, gravitropism, and lateral development: Correlation with auxin transport. Plant Physiol. Biochem. PPB 1994, 32, 193–203. [Google Scholar]
- Vidal, E.A.; Gutiérrez, R.A. A systems view of nitrogen nutrient and metabolite responses in Arabidopsis. Curr. Opin. Plant Biol. 2008, 11, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Wang, R.; Crawford, N.M. The Arabidopsis dual-affinity nitrate transporter gene AtNRT1.1 (CHL1) is regulated by auxin in both shoots and roots. J. Exp. Bot. 2002, 53, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Walch-Liu, P.; Forde, B.G. Nitrate signalling mediated by the NRT1.1 nitrate transporter antagonises l-glutamate-induced changes in root architecture. Plant J. 2008, 54, 820–828. [Google Scholar] [CrossRef] [PubMed]
- Rocha, K.F.; Mariano, E.; Grassmann, C.S.; Trivelin, P.C.O.; Rosolem, C.A. Fate of 15N fertilizer applied to maize in rotation with tropical forage grasses. Field Crops Res. 2019, 238, 35–44. [Google Scholar] [CrossRef]
- Calonego, J.C.; Rosolem, C.A. Soybean root growth and yield in rotation with cover crops under chiseling and no-till. Eur. J. Agron. 2010, 33, 242–249. [Google Scholar] [CrossRef]
- Williams, S.M.; Weil, R.R. Crop Cover Root Channels May Alleviate Soil Compaction Effects on Soybean Crop. Soil Sci. Soc. Am. J. 2004, 68, 148–153. [Google Scholar] [CrossRef]
- Hernandez-Ramirez, G.; Lawrence-Smith, E.J.; Sinton, S.M.; Tabley, F.; Schwen, A.; Beare, M.H.; Brown, H.E. Root responses to alterations in microporosity and penetrability in a silt loam soil. Soil Sci. Soc. Am. J. 2014, 78, 1392–1403. [Google Scholar] [CrossRef]
- Oliveira, E.L.; Pavan, M.A. Control of soil acidity in no-tillage system for soybean production. Soil Tillage Res. 1996, 38, 47–57. [Google Scholar] [CrossRef]
- Caires, E.F.; Alleoni, L.R.F.; Cambri, M.A.; Barth, G. Surface Application of Lime for Crop Grain Production Under a No-Till System. Agron. J. 2005, 97, 791–798. [Google Scholar] [CrossRef]
- Franchini, J.C.; Miyazawa, M.; Pavan, M.A.; Malavolta, E. Dinâmica de íons em solo ácido lixiviado com extratos de resíduos de adubos verdes e soluções puras de ácidos orgânicos. Pesqui. Agropecuária Bras. 1999, 34, 2267–2276. [Google Scholar] [CrossRef]
- Pariz, C.M.; Andreotti, M.; Buzetti, S.; Bergamaschine, A.F.; Ulian, N.D.; Furlan, L.C.; Meirelles, P.R.; Cavasano, F.A. Straw decomposition of nitrogen-fertilized grasses intercropped with irrigated maize in an integrated crop-livestock system. Rev. Bras. Ciência Solo 2011, 35, 1–17. [Google Scholar] [CrossRef]
- Penariol, F.G.; Fornasieri Filho, D.; Coicev, L.; Bordin, L.; Farinelli, R. Comportamento de Cultivares de Milho Semeadas em Diferentes Espaçamentos entre Linhas e Densidades Populacionais, na Safrinha. Rev. Bras. Milho E Sorgo 2003, 2, 52–60. [Google Scholar] [CrossRef]
- Souza, E.F.C.; Soratto, R.P. Efeito de Fontes e Doses de Nitrogênio em Cobertura, no Milho Safrinha, em Plantio Direto. Rev. Bras. Milho E Sorgo 2006, 5, 395–405. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Depth | pH # | OM † | P | S | K | Ca | Mg | Al | H + Al | CEC ‡ | BS § |
|---|---|---|---|---|---|---|---|---|---|---|---|
| m | CaCl2 | g dm−3 | mg dm−3 | --------------- mmolc dm−3 --------------- | % | ||||||
| 0.00–0.10 | 4.4 | 18 | 13 | 5 | 2.1 | 12 | 12 | 9 | 52 | 78 | 32 |
| 0.10–0.20 | 4.3 | 14 | 10 | 5 | 1.1 | 9 | 8 | 9 | 48 | 66 | 27 |
| 0.20–0.40 | 4.1 | 11 | 6 | 22 | 0.5 | 4 | 4 | 12 | 55 | 63 | 13 |
| 0.40–0.60 | 4.0 | 11 | 6 | 21 | 0.4 | 3 | 4 | 20 | 90 | 97 | 8 |
| Depth (m) | 2017/2018 | 2018/2019 | 2019/2020 | |||
|---|---|---|---|---|---|---|
| Row | Inter-Row | Row | Inter-Row | Row | Inter-Row | |
| 0–0.10 | 0.641 ** | 0.588 ** | 0.733 ** | 0.686 ** | 0.493 ** | 0.597 ** |
| 0.10–0.20 | 0.481 ** | 0.630 ** | 0.319 * | 0.544 ** | 0.407 ** | 0.661 ** |
| 0.20–0.40 | 0.292 * | 0.269 ns | 0.433 ** | 0.302 * | 0.565 ** | −0.066 ns |
| 0.40–0.60 | 0.699 ** | 0.732 ** | 0.735 ** | 0.448 ** | 0.387 ** | 0.462 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Souza, M.; Barcelos, J.P.d.Q.; Rosolem, C.A. Synergistic Effects of Subsoil Calcium in Conjunction with Nitrogen on the Root Growth and Yields of Maize and Soybeans in a Tropical Cropping System. Agronomy 2023, 13, 1547. https://doi.org/10.3390/agronomy13061547
De Souza M, Barcelos JPdQ, Rosolem CA. Synergistic Effects of Subsoil Calcium in Conjunction with Nitrogen on the Root Growth and Yields of Maize and Soybeans in a Tropical Cropping System. Agronomy. 2023; 13(6):1547. https://doi.org/10.3390/agronomy13061547
Chicago/Turabian StyleDe Souza, Murilo, Jéssica Pigatto de Queiroz Barcelos, and Ciro A. Rosolem. 2023. "Synergistic Effects of Subsoil Calcium in Conjunction with Nitrogen on the Root Growth and Yields of Maize and Soybeans in a Tropical Cropping System" Agronomy 13, no. 6: 1547. https://doi.org/10.3390/agronomy13061547
APA StyleDe Souza, M., Barcelos, J. P. d. Q., & Rosolem, C. A. (2023). Synergistic Effects of Subsoil Calcium in Conjunction with Nitrogen on the Root Growth and Yields of Maize and Soybeans in a Tropical Cropping System. Agronomy, 13(6), 1547. https://doi.org/10.3390/agronomy13061547