
Interspecific Competition between Invasive Spodoptera frugiperda and Indigenous Helicoverpa armigera in Maize Fields of China


Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Maize Plants
2.2. Predation Assays
2.3. Predation Functional Responses
2.4. Field Competition and Population Dynamics
2.4.1. Interspecific Competition between FAW and CBW on Maize Plants
2.4.2. Population Density Survey of FAW and CBW in Local Maize Fields
2.5. Data Analyses
3. Results
3.1. Predation Rates between FAW and CBW
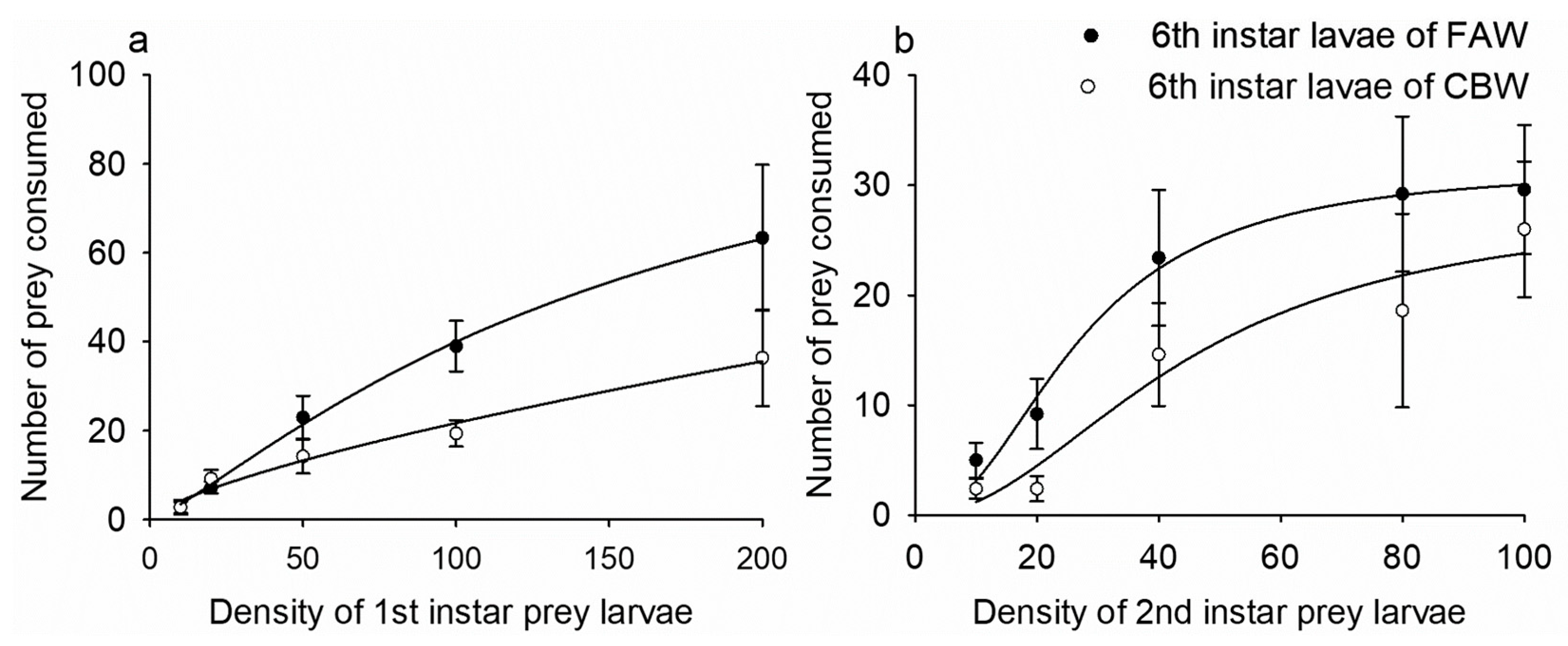
3.2. Predation Functional Responses between FAW and CBW
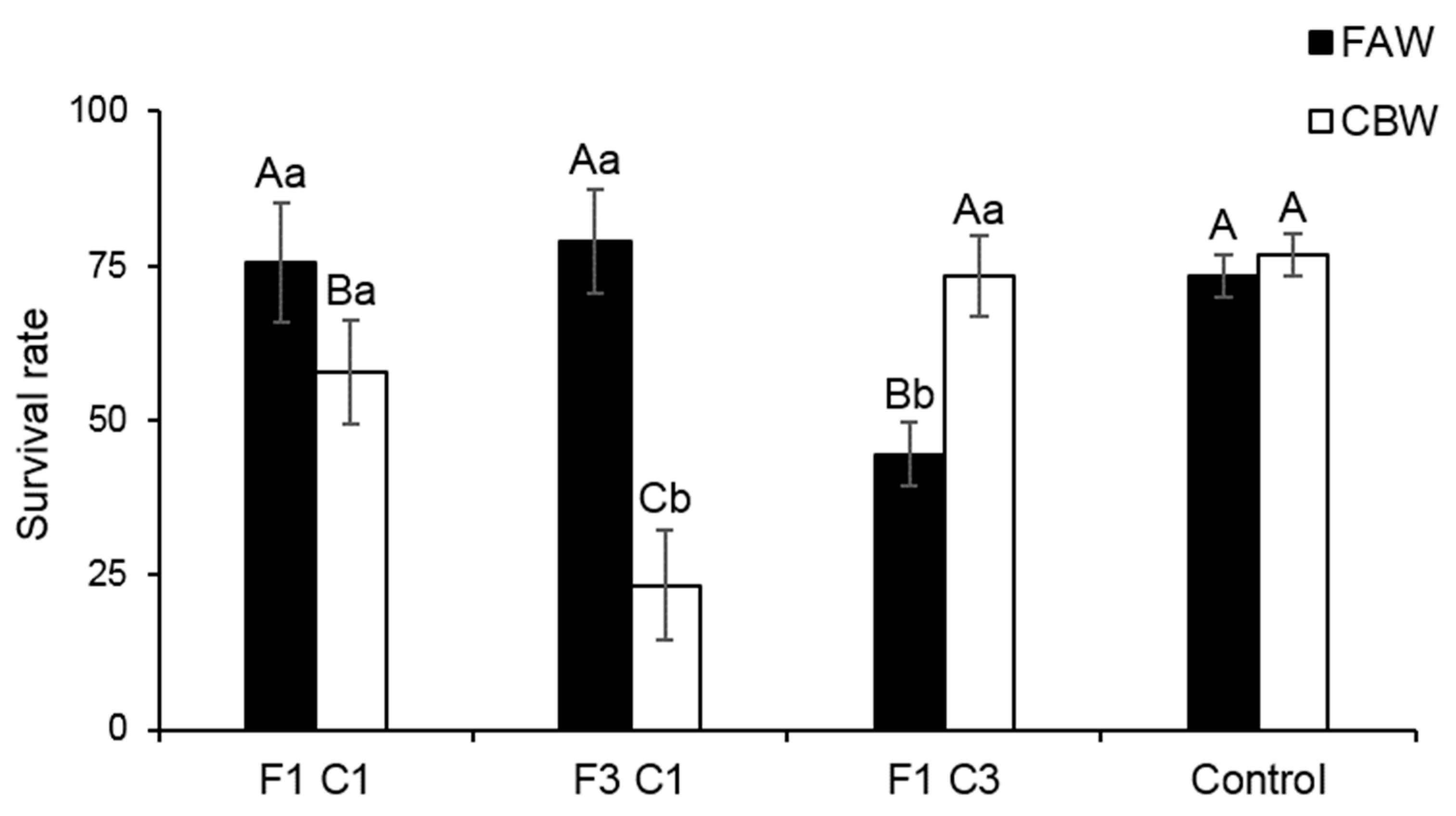
3.3. Interspecific Competition between FAW and CBW on Maize Plants
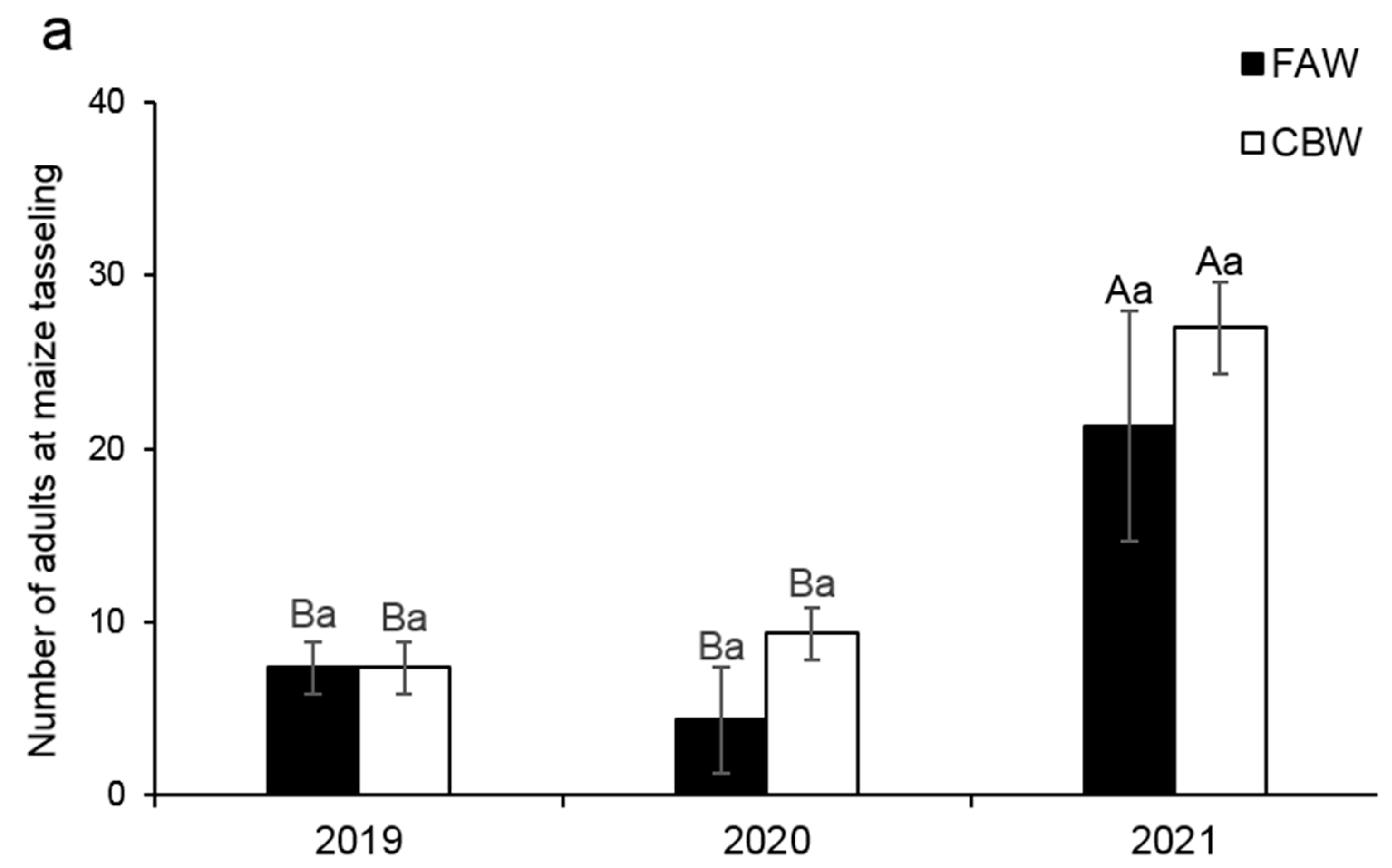
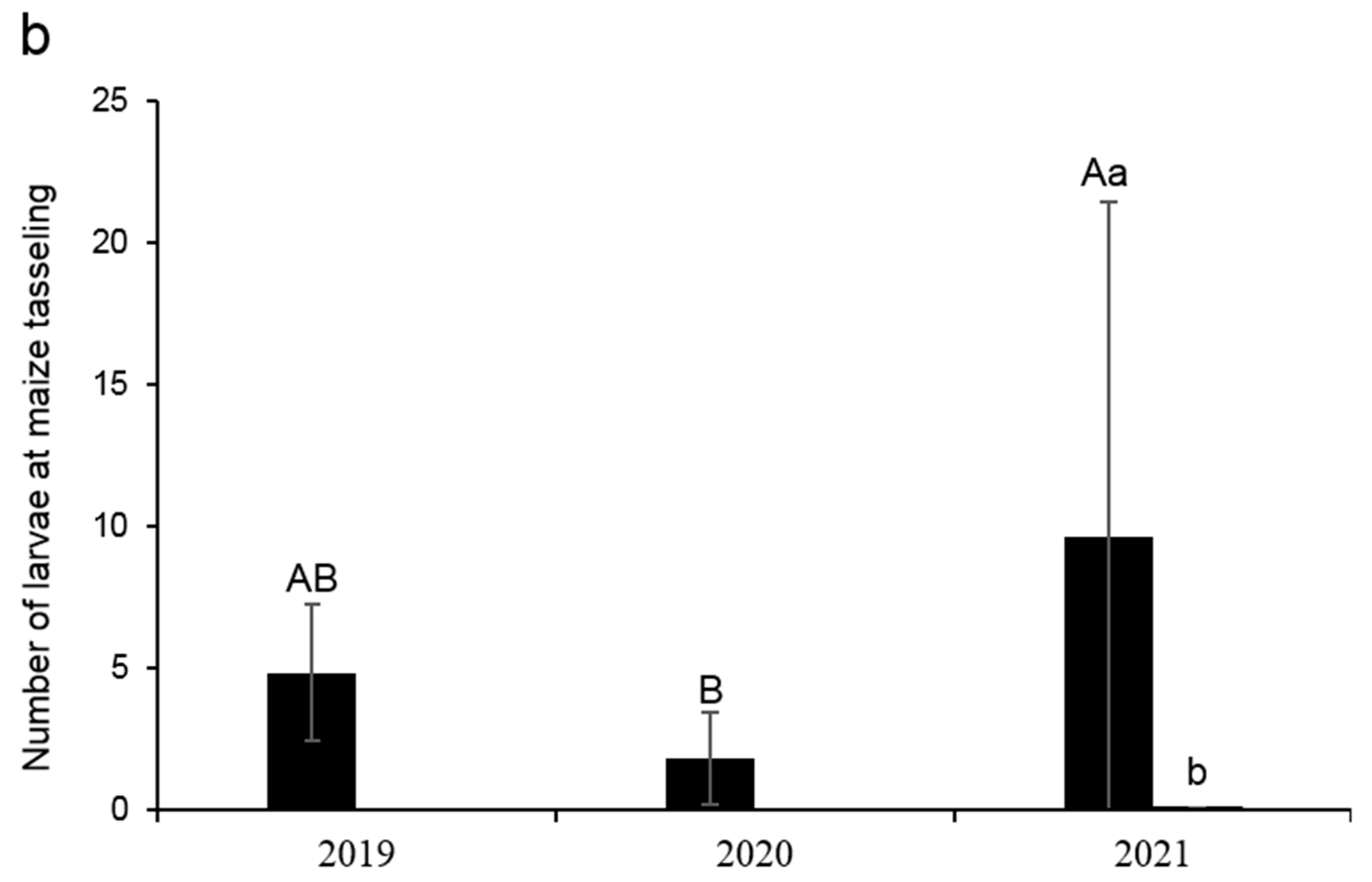
3.4. Population Abundances of FAW and CBW in Local Maize Fields
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Levine, J.M.; D’Antonio, C.M. Forecasting biological invasions with increasing international trade. Conserv. Biol. 2003, 17, 322–326. [Google Scholar] [CrossRef]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Wan, F.H.; Yang, N.W. Invasion and management of agricultural alien insects in China. Annu. Rev. Entomol. 2016, 61, 77–98. [Google Scholar] [CrossRef] [PubMed]
- Wan, F.H.; Jiang, M.X.; Zhan, A. Biological invasions and its management in China. In Invading Nature-Springer Series in Invasion Ecology; Daniel, S., Ed.; Springer: Berlin, Germany, 2017; Volume 1, pp. 125–145. [Google Scholar]
- De Bach, P. The competitive displacement and coexistence principles. Annu. Rev. Entomol. 1966, 11, 183–212. [Google Scholar] [CrossRef]
- Giga, D.P.; Canheo, S.J. Competition between Prostephanus truncatus (Horn) and Sitophilus zeamais (Motsch.) in maize at two temperatures. J. Stored Prod. Res. 1993, 29, 63–70. [Google Scholar] [CrossRef]
- Durrett, R.; Levin, S. Spatial aspects of interspecific competition. Theor. Popul. Biol. 1998, 53, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Ekesi, S.; Billah, M.K.; Nderitu, P.W. Evidence for competitive displacement of Ceratitis cosyra by the invasive fruit fly Bactrocera invadens (Diptera: Tephritidae) on mango and mechanisms contributing to the displacement. J. Econ. Entomol. 2009, 102, 981–991. [Google Scholar] [CrossRef]
- Kfir, R. Competitive displacement of Busseola fusca (Lepidoptera: Noctuidae) by Chilo partellus (Lepidoptera: Pyralidae). Ann. Entomol. Soc. Am. 1997, 90, 619–624. [Google Scholar] [CrossRef]
- Liu, S.S.; De Barro, P.J.; Xu, J.; Luan, J.B.; Zang, L.S.; Ruan, Y.M.; Wan, F.H. Asymmetric mating interactions drive widespread invasion and displacement in a whitefly. Science 2007, 318, 1769–1772. [Google Scholar] [CrossRef] [PubMed]
- Sparks, A.N. A review of the biology of the fall armyworm. Fla. Entomol. 1979, 62, 82–87. [Google Scholar] [CrossRef]
- Sharanabasappa, D.; Kalleshwaraswamy, C.M.; Asokan, R.; Swamy, H.M.M.; Maruthi, M.S.; Pavithra, H.B.; Hegde, K.; Navi, S.; Prabhu, S.T.; Goergen, G. First report of the fall armyworm, Spodoptera frugiperda (J E Smith) (Lepidoptera: Noctuidae), an alien invasive pest on maize in India. Pest Manag. Hortic. Ecosyst. 2018, 24, 23–29. [Google Scholar]
- Sun, X.X.; Hu, C.X.; Jia, H.R.; Wu, Q.L.; Shen, X.J.; Zhao, S.Y.; Jiang, Y.Y.; Wu, K.M. Case study on the first immigration of fall armyworm Spodoptera frugiperda invading into China. J. Integr. Agric. 2019, 18, 2–10. [Google Scholar] [CrossRef]
- Li, X.J.; Wu, M.F.; Ma, J.; Gao, B.Y.; Wu, Q.L.; Chen, A.D.; Liu, J.; Jiang, Y.Y.; Zhai, B.P.; Early, R.; et al. Prediction of migratory routes of the invasive fall armyworm in eastern China using a trajectory analytical approach. Pest Manag. Sci. 2020, 76, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Hang, D.T.; Liem, N.V.; Lam, P.V.; Wyckhuys, W.A.G.; Thuy, N.T.; Hang, T.T.T.; Trong, P.D.; Viêt, N.D. First record of fall armyworm Spodoptera frugiperda (J.E. Smith), (Lepidoptera: Noctuidae) on maize in Viet Nam. Zootaxa 2020, 4772, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.L.; Jiang, Y.Y.; Wu, K.M. Analysis of migration routes of fall armyworm Spodoptera frugiperda (J. E. Smith) from Myanmar to China. Plant Prot. 2019, 45, 1–6. [Google Scholar] [CrossRef]
- Yang, X.M.; Song, Y.F.; Sun, X.X.; Shen, X.J.; Wu, Q.L.; Zhang, H.W.; Zhang, D.D.; Zhao, S.Y.; Liang, G.M.; Wu, K.M. Population occurrence of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), in the winter season of China. J. Integr. Agric. 2021, 20, 772–782. [Google Scholar] [CrossRef]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in west and central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [PubMed]
- Montezano, D.G.; Specht, A.; Sosa–Gòmez, D.R.; Roque–Specht, V.F.; Sousa–Silva, J.C.; Paula–Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–301. [Google Scholar] [CrossRef]
- Wu, K.M. Management strategies of fall armyworm (Spodoptera frugiperda) in China. Plant Prot. 2020, 46, 1–5. [Google Scholar] [CrossRef]
- Zhou, Y.; Wu, Q.L.; Zhang, H.W.; Wu, K.M. Spread of invasive migratory pest Spodoptera frugiperda and management practices throughout China. J. Integr. Agric. 2021, 20, 637–645. [Google Scholar] [CrossRef]
- Guo, Y.Y. Research on Cotton Bollworm; China Agricultural Press: Beijing, China, 1998; pp. 1–407. [Google Scholar]
- Zalucki, M.P.; Daglish, G.; Firempong, S.; Twine, P. The biology and ecology of Heliothis armigera (Hübner) and Heliothis punctigera Wallengren (Lepidoptera, Noctuidae) in Australia–What do we know. Aust. J. Zool. 1986, 34, 779–814. [Google Scholar] [CrossRef]
- Fitt, G.P. The ecology of Heliothis species in relation to agroecosystems. Annu. Rev. Entomol. 1989, 34, 17–53. [Google Scholar] [CrossRef]
- Wu, K.M.; Guo, Y.Y. The evolution of cotton pest management practices in China. Annu. Rev. Entomol. 2005, 50, 31–52. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Wang, X.M. Current status management strategies for corn pests and diseases in China. Plant Prot. 2019, 45, 1–11. [Google Scholar] [CrossRef]
- Chen, Q.; Duan, Y.; Hou, Y.H.; Chen, L.; Fan, Z.Y. Morphological characteristics of Spodoptera frugiperda in comparison with three other common Noctuidae species at maize filling stage. Plant Prot. 2020, 46, 34–41. [Google Scholar] [CrossRef]
- Song, Y.F.; Yang, X.M.; Zhang, H.W.; Zhang, D.D.; Wu, K.M. Interference competition and predation between invasive and native herbivores in maize. J. Pest Sci. 2021, 94, 1053–1063. [Google Scholar] [CrossRef]
- Greene, G.L.; Leppla, N.C.; Dickerson, W.A. Velvetbean caterpillar: A rearing procedure and artificial medium. J. Econ. Entomol. 1976, 69, 487–488. [Google Scholar] [CrossRef]
- Nielsen, R.L. Determining Corn Leaf Stages. Purdue University. 2019. Available online: https://www.agry.purdue.edu/ext/corn/news/index.html (accessed on 3 March 2022).
- Feng, H.Q.; Wu, X.F.; Wu, B.; Wu, K.M. Seasonal migration of Helicoverpa armigera (Lepidoptera: Noctuidae) over the Bohai Sea. J. Econ. Entomol. 2009, 102, 95–104. [Google Scholar] [CrossRef]
- Juliano, S.A. Nonlinear Curve Fitting: Predation and Functional Response Curves; Oxford University Press: Oxford, UK, 2001; pp. 178–196. [Google Scholar]
- De Clercq, P.; Mohaghegh, J.; Tirry, L. Effect of host plant on the functional response of the predator Podisus nigrispinus (Heteroptera: Pentatomidae). Biol. Control. 2000, 18, 65–70. [Google Scholar] [CrossRef]
- Chapman, J.W.; Williams, T.; Martínez, A.M.; Cisneros, J.; Caballero, P.; Cave, R.D.; Goulson, D. Does cannibalism in Spodoptera frugiperda (Lepidoptera: Noctuidae) reduce the risk of predation? Behav. Ecol. Sociobiol. 2000, 48, 321–327. [Google Scholar] [CrossRef]
- Andow, D.A.; Farias, J.R.; Horikoshi, R.J.; Bernardi, D.; Nascimento, A.R.; Omoto, C. Dynamics of cannibalism in equal–aged cohorts of Spodoptera frugiperda. Ecol. Entomol. 2015, 40, 229–236. [Google Scholar] [CrossRef]
- Tsuyoshi, K.; Fujisaki, K.; Takahisa, M. Egg laying preference, larval dispersion, and cannibalism in Helicoverpa armigera (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 2001, 6, 793–798. [Google Scholar] [CrossRef]
- Sigsgaard, L.; Greenstone, M.H.; Duffield, S.J. Egg cannibalism in Helicoverpa armigera on sorghum and pigeon pea. BioControl 2002, 47, 151–165. [Google Scholar] [CrossRef]
- Zanuncio, J.C.; Silva, C.A.D.D.; Lima, E.R.D.; Pereira, F.F.; Ramalho, F.D.S.; Serrao, J.E. Predation rate of Spodoptera frugiperda (Lepidoptera: Noctuidae) larvae with and without defense by Podisus nigrispinus (Heteroptera: Pentatomidae). Braz. Arch. Biol. Technol. 2008, 51, 121–125. [Google Scholar] [CrossRef]
- Silva, R.B.D.; Corrêa, A.S.; Della Lucia, T.M.C.; Pereira, A.I.A.; Cruz, I.; Zanuncio, J.C. Does the aggressiveness of the prey modify the attack behavior of the predator Supputius cincticeps (Stal) (Hemiptera, Pentatomidae)? Rev. Bras. Entomol. 2012, 56, 244–248. [Google Scholar] [CrossRef]
- Li, H.; Jiang, S.S.; Zhang, H.W.; Geng, T.; Kris, A.G.W.; Wu, K.M. Two–way predation between immature stages of the hoverfly Eupeodes corollae and the invasive fall armyworm (Spodoptera frugiperda J. E. Smith). J. Integr. Agric. 2021, 20, 829–839. [Google Scholar] [CrossRef]
- Li, H.; Wu, K.M. Bidirectional predation between larvae of the hoverfly Episyrphus balteatus (Diptera: Syrphidae) and the fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Econ. Entomol. 2022, 115, 545–555. [Google Scholar] [CrossRef]
- Gao, Y.L.; Reitz, S.R. Emerging themes in our understanding of species displacements. Annu. Rev. Entomol. 2017, 62, 165–183. [Google Scholar] [CrossRef]
- Bentivenha, J.P.F.; Montezano, D.G.; Hunt, T.E.; Baldin, E.L.L.; Peterson, J.A.; Victor, V.S.; Pannuti, L.E.R.; Vélez, A.M. Intraguild interactions and behavior of Spodoptera frugiperda and Helicoverpa spp. on maize. Pest Manag. Sci. 2017, 73, 2244–2251. [Google Scholar] [CrossRef]
- Bentivenha, J.P.F.; Baldin, E.L.L.; Hunt, T.E.; Paula–Moraes, S.V.; Blankenship, E.E. Intraguild competition of three noctuid maize pests. Environ. Entomol. 2016, 45, 999–1008. [Google Scholar] [CrossRef]
- Bentivenha, J.P.F.; Baldin, E.L.L.; Montezano, D.G.; Hunt, T.E.; Paula–Moraes, S.V. Attack and defense movements involved in the interaction of Spodoptera frugiperda and Helicoverpa zea (Lepidoptera: Noctuidae). J. Pest Sci. 2017, 90, 433–445. [Google Scholar] [CrossRef]
- Zhao, J.; Hoffmann, A.; Jiang, Y.P.; Xiao, L.B.; Tan, Y.G.; Zhou, C.Y.; Bai, L.X. Competitive interactions of a new invader (Spodoptera frugiperda) and indigenous species (Ostrinia furnacalis) on maize in China. J. Pest Sci. 2021, 95, 159–168. [Google Scholar] [CrossRef]
- Harwood, J.D.; Obrycki, J.J. Web–Construction behavior of linyphiid spiders (Araneae, Linyphiidae): Competition and co–existence within a generalist predator guild. J. Insect Behav. 2005, 18, 593–607. [Google Scholar] [CrossRef]
- Chapman, J.W.; Williams, T.; Escribano, A.; Caballero, P.; Cave, R.D.; Goulson, D. Age-related cannibalism and horizontal transmission of a nuclear polyhedrosis virus in larval Spodoptera frugiperda. Ecol. Entomol. 1999, 24, 268–275. [Google Scholar] [CrossRef]
- Deng, P.; Ma, W.H.; Li, G.Q. Age–and nutrition–related cannibalism in larvae of the cotton bollworm, Helicoverpa armigera (Lepidoptera: Noctuidae). Acta Entomol. Sin. 2015, 58, 175–180. [Google Scholar] [CrossRef]
- Wang, Z.J.; Li, D.M.; Xie, B.Y. Determination and assessment for risk areas of the Helicoverpa armigera Hübner. Acta Ecol. Sin. 2003, 12, 2642–2652. [Google Scholar]
- Luo, S.Q. Huge occurrence of cotton bollworm in wheat fields in Changning County, Yunnan Province. Plant Prot. 1992, 5, 44. [Google Scholar]
- Zhang, L. Pomegranate pests in Mengzi, Yunnan and their comprehensive control. China Fruits 2002, 5, 32–41. [Google Scholar] [CrossRef]
- An, T.X.; Dai, P.; Wu, Z.B.; Niu, H.L.; Dai, L.K. Control efficacy of intercropping of sweet maize and vegetables on main pest and disease. J. Yunnan Agric. Univ. 2011, 26, 449–453. [Google Scholar] [CrossRef]
- Bu, M.; Li, A.G. Regional change characteristics of interannual temperature and precipitation in Yunnan during 1960–2012. Guizhou Agric. Sci. 2017, 45, 130–136. [Google Scholar]
- Hailu, G.; Niassy, S.; Bässler, T.; Ochatum, N.; Studer, C.; Salifu, D.; Agbodzavu, M.K.; Khan, Z.R.; Midega, C.; Subramanian, S. Could fall armyworm, Spodoptera frugiperda (J. E. Smith) invasion in Africa contribute to the displacement of cereal stemborers in maize and sorghum cropping systems. Int. J. Trop. Insect Sci. 2021, 41, 1753–1762. [Google Scholar] [CrossRef]
- Mutua, J.M.; Mutyambai, D.M.; Asudi, G.O.; Khamis, F.; Niassy, S.; Jalloh, A.A.; Salifu, D.; Magara, H.J.O.; Calatayud, P.A. Competitive plant–mediated and intraguild predation interactions of the invasive Spodoptera frugiperda and resident stem borers Busseola fusca and Chilo partellus in maize cropping systems in Kenya. Insects 2022, 13, 790. [Google Scholar] [CrossRef]
- Yang, X.M.; Wyckhuys, K.; Jia, X.P.; Nie, F.Y.; Wu, K.M. Fall armyworm invasion heightens pesticide expenditure among Chinese smallholder farmers. J. Environ. Manag. 2021, 282, 111949. [Google Scholar] [CrossRef]
- Wu, K.M.; Lu, Y.H.; Feng, H.Q.; Jiang, Y.Y.; Zhao, Z.J. Suppression of cotton bollworm in multiple crops in China in areas with Bt toxin–containing cotton. Science 2008, 321, 1676–1678. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.M.; Zhao, S.Y.; Liu, B.; Gao, Y.; Hu, C.X.; Li, W.J.; Yang, Y.Z.; Li, G.P.; Wang, L.L.; Yang, X.Q.; et al. Bt maize can provide non–chemical pest control and enhance food safety in China. Plant Biotechnol. J. 2022, 21, 391–404. [Google Scholar] [CrossRef]
- Lu, Y.H.; Wu, K.M.; Jiang, Y.Y.; Guo, Y.Y.; Desneux, N. Widespread adoption of Bt cotton and insecticide decrease promotes biocontrol services. Nature 2012, 487, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.H.; Jiang, Y.Y.; Liu, J.; Zeng, J.; Yang, X.M.; Wu, K.M. Adjustment of cropping structure increases the risk of cotton bollworm outbreaks in China. Chin. J. Appl. Entomol. 2018, 55, 19–24. [Google Scholar] [CrossRef]
- Liu, J.; Liu, W.C.; Jiang, Y.Y. Analysis of the occurrence characteristics and causes of major diseases and pests of maize in 2017. Plant Prot. 2019, 45, 152–157. [Google Scholar] [CrossRef]
- Liu, J.; Li, T.J.; Jiang, Y.Y.; Zeng, J.; Wang, Y.P. Characteristics of major diseases and pests of maize in China in 2020. China Plant Prot. 2021, 45, 30–35. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predator Species | Larval Instar | Larval Instar of Prey (H. armigera/S. frugiperda) | |||||
|---|---|---|---|---|---|---|---|
| 1st | 2nd | 3rd | 4th | 5th | 6th | ||
| S. frugiperda | 1st | 0 | 0 | 0 | 0 | 0 | 0 |
| 2nd | 53.3 ± 9.1 Abc | 10.0 ± 5.5 Bc | 0 | 0 | 0 | 0 | |
| 3rd | 76.7 ± 7.7 Ab | 50.0 ± 9.1 Bb | 16.7 ± 6.8 Cd | 0 | 0 | 0 | |
| 4th | 93.3 ± 4.6 Aab | 96.7 ± 3.3 Aa | 50.0 ± 9.1 Bc | 30.0 ± 8.4 Cb | 3.3 ± 3.3 Dc | 0 | |
| 5th | 96.7 ± 3.3 Aa | 96.7 ± 3.3 Aa | 86.7 ± 6.2 Abb | 73.3 ± 8.1 Ba | 20.0 ± 7.3 Cb | 3.3 ± 3.3 Da | |
| 6th | 96.7 ± 3.3 Aa | 100.0 ± 0.0 Aa | 100.0 ± 0.0 Aa | 76.7 ± 7.7 Ba | 76.7 ± 7.7 Ba | 16.7 ± 6.8 Ca | |
| H. armigera | 1st | 0 | 0 | 0 | 0 | 0 | 0 |
| 2nd | 36.7 ± 8.8 Ab | 10.0 ± 5.5 Bc | 0 | 0 | 0 | 0 | |
| 3rd | 93.3 ± 4.6 Aa | 66.7 ± 8.6 Bb | 10.0 ± 5.5 Cc | 0 | 0 | 0 | |
| 4th | 100.0 ± 0.0 Aa | 90.0 ± 5.5 Aa | 50.0 ± 9.1 Bb | 3.3 ± 3.3 Cb | 0 | 0 | |
| 5th | 100.0 ± 0.0 Aa | 96.7 ± 3.3 Aa | 63.3 ± 8.8 Bb | 66.7 ± 8.6 Ba | 10.0 ± 5.5 Cb | 0 | |
| 6th | 100.0 ± 0.0 Aa | 100.0 ± 0.0 Aa | 100.0 ± 0.0 Aa | 86.7 ± 6.2 Ba | 56.7 ± 9.1 Ca | 6.7 ± 4.6 D | |
| Predator Species | Larval Stage of Prey | Parameter | Estimated Value | SE | T-Value | Pr(>|z|) |
|---|---|---|---|---|---|---|
| S. frugiperda | 1st-instar of | P0 | –0.1382 | 0.2568 | –5.3820 | 0.0000 |
| H. armigera | P1 | 0.0896 | 0.0332 | 2.6950 | 0.0122 | |
| P2 | –0.0025 | 0.0011 | –2.3580 | 0.0262 | ||
| P3 | 0.0000 | 0.0000 | 2.1670 | 0.0396 | ||
| 2nd-instar of | P0 | –1.9691 | 0.7408 | –2.6580 | 0.0133 | |
| H. armigera | P1 | 0.3781 | 0.1703 | 2.2200 | 0.0354 | |
| P2 | –0.0215 | 0.0104 | –2.0630 | 0.0492 | ||
| P3 | 0.0003 | 0.0002 | 1.8880 | 0.0702 | ||
| H. armigera | 1st-instar of | P0 | –0.1370 | 0.3875 | –3.5360 | 0.0020 |
| S. frugiperda | P1 | 0.1856 | 0.0828 | 2.2420 | 0.0359 | |
| P2 | –0.0012 | 0.0047 | –2.6260 | 0.0157 | ||
| P3 | 0.0001 | 0.0000 | 2.6730 | 0.0143 | ||
| 2nd-instar of | P0 | –1.7780 | 0.2440 | –7.2860 | 0.0000 | |
| S. frugiperda | P1 | 0.1036 | 0.0479 | 2.1640 | 0.0406 | |
| P2 | –0.0027 | 0.0021 | –1.2830 | 0.2116 | ||
| P3 | 0.0000 | 0.0000 | 1.1180 | 0.2748 |
| Predator | Stage of Prey Species | Model | a | Th (d) | Nm | R2 |
|---|---|---|---|---|---|---|
| FAW | 1st-instar of CBW | III | b1N0 | 0.014 ± 0.001 | 71.43 | 0.871 |
| 2nd-instar of CBW | III | b2N0 | 0.031 ± 0.002 | 32.26 | 0.818 | |
| CBW | 1st-instar of FAW | III | b3N0 | 0.026 ± 0.003 | 38.46 | 0.732 |
| 2nd-instar of FAW | III | b4N0 | 0.035 ± 0.005 | 28.57 | 0.795 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.; Li, H.; He, L.; Zhang, H.; Zhao, S.; Yang, X.; Wu, K. Interspecific Competition between Invasive Spodoptera frugiperda and Indigenous Helicoverpa armigera in Maize Fields of China. Agronomy 2023, 13, 911. https://doi.org/10.3390/agronomy13030911
Song Y, Li H, He L, Zhang H, Zhao S, Yang X, Wu K. Interspecific Competition between Invasive Spodoptera frugiperda and Indigenous Helicoverpa armigera in Maize Fields of China. Agronomy. 2023; 13(3):911. https://doi.org/10.3390/agronomy13030911
Chicago/Turabian StyleSong, Yifei, Hui Li, Limei He, Haowen Zhang, Shengyuan Zhao, Xianming Yang, and Kongming Wu. 2023. "Interspecific Competition between Invasive Spodoptera frugiperda and Indigenous Helicoverpa armigera in Maize Fields of China" Agronomy 13, no. 3: 911. https://doi.org/10.3390/agronomy13030911
APA StyleSong, Y., Li, H., He, L., Zhang, H., Zhao, S., Yang, X., & Wu, K. (2023). Interspecific Competition between Invasive Spodoptera frugiperda and Indigenous Helicoverpa armigera in Maize Fields of China. Agronomy, 13(3), 911. https://doi.org/10.3390/agronomy13030911