Abstract
Recombinases are in part responsible for homologous recombination and genome integrity during DNA repair. DMC1 has a typical RecA domain, and belongs to the recombinase superfamily. The reactive oxygen species (ROS) as a potent DNA damage agent is produced during seed germination under stress conditions. DNA repair should be initiated immediately to allow for subsequent seedling development. In this study, we attempted to characterize the underlying mechanism of BrDMC1 responsiveness to salinity stress using the RNA interference approach in Brassica rapa (B. rapa). Bioinformatics and expression pattern analysis revealed that BrDMC1 only retained BrDMC1.A01 after the whole genome triplication (WGT) event and was primarily transcribed in flowers and seeds. BrDMC1 had high activity in the promoter region during germination, according to histochemical GUS staining. The data showed that salt treatment reduced the germination rate, weakened seed vigor and decreased antioxidant enzyme activity, but increased oxidative damage in BrDMC1-RNAi seeds. Furthermore, the expression of stress-responsive genes and damage repair genes was significantly different in transgenic lines exposed to salt stress. Therefore, BrDMC1 may respond to salt stress by controlling seed germination and the expression of stress-related and damage repair genes in B. rapa.
1. Introduction
Abiotic stressors such as high temperatures, high salinities, toxins, and ionizing radiation can severely damage plants [1,2,3,4,5]. Plants can be divided into halophytes and glycophytes according to their tolerance of salt. Halophytes are salt-tolerant plants, but the majority of crops are glycophytes, which means that salt negatively inhibits their growth and development [6,7]. Salt stress is an unavoidable environmental factor that can impede plant growth and development by affecting seed germination and photosynthesis inhibition [8]. It causes an increase in reactive oxygen species (ROS; e.g., O2•−, H2O2, OH• and 1O2) [9]. If unchecked, the elevated level of ROS can result in DNA base damage, single-strand breaks (SSBs) and even double-strand breaks (DSBs) [10]. In plants, ROS detoxifying proteins (e.g., ascorbate peroxidase (APX), glutathione peroxidase (GPX), superoxide dismutase (SOD) and catalase (CAT)), antioxidants, for instance ascorbic acid and glutathione (GSH), and osmoprotectants such as proline and soluble sugars are used to remove ROS and improve tolerance to salinity and oxidative stress [11,12,13]. So, plants need some complex regulatory mechanisms to survive in high salinity environments.
Seed germination and lifetime are significantly influenced by genome maintenance [14,15]. Successful seed germination is a critical developmental stage in the plant life cycle, during which the plant is exposed to extremely high levels of genotoxic stress [16]. When seeds are exposed to unfavorable environmental conditions, ROS can accumulate, seed germination potential can be reduced, seed vigor is lost and, eventually, viability is lost [17,18]. Dehydration and rehydration during seed development and germination are linked to oxidative stress caused by ROS, which causes organelle disruption and protein, RNA and DNA damage (e.g., SSBs and DSBs), thus exacerbating genomic damage [10,19]. Double-strand breaks (DSBs) are considered to be the most important DNA damage because they can lead to chromosome fragmentation and rearrangement and loss of genetic information [20]. The integrity of the genome is an important determinant of seed quality [21]. Thus, seed germination and viability are dependent on the ability to resist or repair genomic damage [10].
Brassica rapa (B. rapa) has been widely cultivated and extensively studied as an important economic and nutritional horticultural species. Although B. rapa and Arabidopsis thaliana (A. thaliana) share a common ancestor, the whole genome triplication (WGT) event distinguishes their genomes, making B. rapa an exemplary model for studying polyploidy events [22,23]. DMC1 (disruption Meiotic cDNA 1), discovered in Saccharomyces cerevisiae, is a homolog of the RecA protein, a DNA-dependent ATPase that participates in nucleation, homology search and chain exchange [24,25]. Many studies of DMC1 in various species have concentrated on its role in meiosis [24,26,27,28,29]. The model plant A. thaliana has a single copy of the DMC1 gene, which is expressed in young buds during meiosis as well as vegetative cells, with low expression in leaves [30]. OsDMC1 has two copies in rice and has an expression pattern similar to A. thaliana, but it is not expressed in mature leaves [31]. Three homologous copies of TaDMC1 were expressed in meiosis tissue and vegetative tissue in bread wheat [24]. It has also been reported that the expression patterns of the DMC1 gene under salt and temperature stresses have also been examined [32]. These results suggest that DMC1 may perform different functions in different species. RecA family proteins are also involved in a variety of abiotic stresses, not just reproductive development [33,34,35]. Nevertheless, the exact information on response to stress in recombinase DMC1 remains lacking. It is unknown whether the known-function DMC1 gene is conserved in B. rapa and whether other special types of the RecA domain participate in stress responses, particularly in B. rapa.
In this study, we preliminarily verified the functional mechanism of BrDMC1 in B. rapa, which is involved in the stress response under salt treatment, and provided a reference for the genetic varieties of crop resistance.
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
B. rapa, A. thaliana (Col-0) and Nicotiana benthamiana (N. benthamiana) were grown in a growth room with 16 h/22° in light, 8 h/16° in darkness and about 60% relative humidity.
2.2. Evolutionary Analysis and Molecular Evolutionary Rate Analysis
The cruciferae species genomes, CDS sequences and protein sequences were obtained from the Ensembl plant database (http://plants.ensembl.org/index.html, accessed on 8 June 2022) [36]. The phylogenetic tree was built using MEGA7 software and the maximum likelihood (ML) method. The number of DMC1 was calculated via evolutionary retention. The synonymous mutation rate (Ks) and non-synonymous mutation rate (Ka) of coding sequences were calculated using the NG method [37]. We used the online tool GSDS (http://gsds.cbi.pku.edu.cn/, accessed on 8 June 2022) to analyze the exons and introns of the DMC1 gene, MEME (http://meme-suite.org/, accessed on 8 June 2022), and analyzed the conserved motifs of the DMC1 gene; conservative functional domain information is available for download from the NCBI database (https://www.ncbi.nlm.nih.gov/cdd, accessed on 8 June 2022).
2.3. Cis-Acting Elements and Co-Expression Network Analysis
We downloaded the upstream 2000 bp promoter region of the DMC1 coding region of the cruciferae species from the Ensembl plant database and analyzed cis-acting regulatory elements of the DMC1 gene of the cruciferae species using the PlantCARE online website (http://bioinformatics.psb.ugent.be/webtools/PlantCARE/htmL, accessed on 8 June 2022). The results were visualized using TBtools software. The DMC1 interaction network was examined and reconstructed using Cytoscape software via the STRING online website (https://cn.string-db.org/, accessed on 8 June 2022).
2.4. Subcellular Localization Analysis in N.benthamiana
To understand the subcellular localization of BrDMC1, recombinant gene expression vectors were constructed, and transient transformation was performed [38]. The full-length BrDMC1 coding sequence was first cloned into the pEASY-T1 vector for amplification before being subcloned into the pCAMBIA2300-GFP vector using BamHI and XbaI restriction sites. They were then transformed in Agrobacterium tumefaciens (A. tumefaciens) (EHA105) and infiltrated in N. benthamiana with 1 mL syringes. An Olympus fluorescence microscope (DX53, Tokoyo, Japan) was used to detect the fluorescence signal of the GFP fusion protein in leaves 48 h after transformation.
2.5. Histochemical Estimation of GUS Activity and ROS Activity Determination of BrDMC1
To create the ProBrDMC1::GUS vector, we cloned the BrDMC1 upstream sequence into the pEASY-T1 vector for amplification before subcloning it into the pCAMBIA3301 vector with XbaI and NcoI restriction sites. They were then transferred to A. tumefaciens (EHA105), and the vector was successfully built. To determine the degree of stress response of the BrDMC1 promoter, the constructed vector was infiltrated into N. benthamiana leaves, and external hormone and stress treatments were applied 48 h later, with the results being observed 24 h later. The ProBrDMC1::GUS transgenic A. thaliana was first seeded on MS solid medium for 7 days, and then transferred to Hoagland nutrient solution for hydroculture to maturity. The flowers, leaves, stems, pods and seeds of the transgenic A. thaliana were incubated in X-Gluc for 12–24 h to detect GUS activity, and were then decolorized using ethanol to observe the expression patterns [39]. To determine ROS activity, embryos of germinating seeds were incubated in 10 mM potassium phosphate buffer (pH 7.8) containing 0.5 mg·mL−1 nitroblue tetrazolium (NBT) for 30 min at room temperature in the dark. Following that, the seeds were stored in 70% glycerol. An Axiocam HRc Digital Camera (Carl Zeiss) was used to photograph GUS and ROS staining.
2.6. Analysis of Proline, Malondialdehyde (MDA) and Soluble Sugar Content
We designed the 406 bp interference fragment using the CDS sequence of BrDMC1, connected it to the PHELLSGATE12 vector via BP recombination and transferred it to A. tumefaciens GV3101 after correct sequencing. Seeds were harvested via the floral dip method [40]. Seeds were initially screened on resistant medium and then identified via PDK sequence amplification and BrDMC1 expression changes. Finally, we selected three independent BrDMC1-RNAi transgenic lines (#1, #3 and #6) and, compared with the wild type, the expression of DMC1 in the BrDMC1-RNAi plants was reduced. The method previously described was used to determine proline [41]. First, 0.1 g of tissue was weighed; then, 1 mL of extract was added for ice bath homogenization before being transferred to a 1.5 mL EP tube, oscillated in a water bath at 90° for 10 min and centrifuged at 12,000 rpm for 10 min at 25°, and supernatant was collected for cooling. After removing 300 μL of supernatant, 300 μL of acetic acid and 600 μL of reagent I were added, and the mixture was heated for 30 min in a 95° water bath. After cooling to room temperature, the solution was analyzed at 520 nm. Malondialdehyde (MDA) content analysis was performed as follows: 0.1 g tissue was weighed, 1 mL extract was added for ice bath homogenization and centrifuged at 12,000 rpm at 4° for 10 min, 400 μL supernatant was taken and 600 μL working liquid was added. After mixing, the tissue was immersed in 95° water for 30 min before being cooled and centrifuged at room temperature at 12,000 rpm for 10 min. They were examined at 532 and 600 nm, respectively [42]. The soluble sugar content was determined according to Grace’s instructions. In short, about 0.1 g of materials was collected, 0.8 mL of 80% ethanol was added and then the sample was ground in an ice bath and incubated at 95° for 20 min. Then, it was cooled and centrifuged at room temperature at 12,000 rpm for 10 min. The 50 μL supernatant was taken out and added to 150 μL distilled water, 60 μL working fluid and 500 μL concentrated sulfuric acid, and then incubated at 95° for 10 min and analyzed via a spectrophotometer at 620 nm.
2.7. Assays of Antioxidative Enzyme Activity
Seed in the quantity of 0.5 g was thoroughly ground in 10 mL of 50 mM sodium phosphate buffer (pH 7.8) containing 1% (w/v) polyvinylpyrrolidone using a pre-chilled mortar and pestle (PVP). The homogenate was centrifuged for 10 min at 4° at 15,000 g. The supernatant was collected and placed in an ice bath as a crude enzyme extract [43]. Superoxide dismutase (SOD; EC 1.15.1.1) activity was determined by taking 60 μL of supernatant and adding reagents 1, 2, 3 and 4 in turn. After thoroughly mixing, the supernatant was allowed to stand for 30 min at room temperature and away from light before being analyzed at 450 nm. Peroxidase (POD; EC 1.11.1.7) activity determination was as follows: take 40 μL of supernatant, add reagents 1, 2 and 3 in order, mix and immediately analyze at 470 nm; then, read again 1 min later. Catalase (CAT; EC 1.11.1.6) activity analysis was as follows: First, 10 μL of supernatant was taken and reagents 1, 2 and 3 were added sequentially. After mixing, 10 μL of the mixture was immediately taken and reagents 1 and 4 were added. After mixing, the reaction was performed at room temperature for 5 min before being analyzed at 510 nm.
2.8. Quantitative Real-Time PCR Analysis
Total RNA was extracted from B. rapa using the Plant Total RNA Isolation Kit Plus (Foregene, Chengdu, China), and cDNA was synthesized using the HiFiScript cDNA Synthesis Kit (CWBio, Beijing, China). For qRT-PCR analysis, the qPCR Master Mix (NEB, Beijing, China) and Light Cycler 480 systems were used. The following PCR program was used as described: 94° for 30 s, 40 cycles at 94° for 10 s and 58° for 30 s, and the melting curve was then used to determine the specificity of the amplification. The 2−∆∆Ct method was used to calculate relative expression levels [44], and β-actin served as an internal control. Three biological replicates were performed using various cDNA preparations. All of the primers used are listed in Table A1 (Appendix A).
2.9. Statistical Analyses
At least three independent biological and technical replicates were used in the experiment. GraphPad Prism 5 was used to perform the statistical analyses and data graphing. A one-way ANOVA and a t test were used to assess the significance of differences between the control and experimental groups. *** p < 0.001, ** p < 0.01 and * p < 0.05 indicate significant differences from the control.
3. Results
3.1. Analysis of DMC1 Genes following a Whole-Genome Triplication Event in B. rapa
Related genes were lost or retained after the Brassica species underwent WGT replication [45]. We analyzed the phylogenetic relationships and retained a copy number of the DMC1 gene in A. thaliana, Brassica napus (B. napus), B. rapa and Brassica oleracea (B. oleracea) (Figure 1A). The results showed that the DMC1 gene retained redundancy after the WGT event, with two copies in B. napus, B. rapa and B. oleracea. The conserved domain analysis of DMC1 showed several domains, such as RecA, Rad51_DMC1_RecA, Rad51 and recomb_DMC1 (Figure 1C). Except for four genes (BoDMC1.C01, AtDMC1, BrDMC1.A03 and BoDMC1.A05), gene structure analysis revealed that the majority of DMC1 genes had fourteen exons. Motifs 1–6 were six conserved motifs. Motif 1–Motif 6 RecA protein domains were found in BnaDMC1, BnaDMC1.A01, BrDMC1.A01, BoDMC1.C01 and AtDMC1, while Motif 5 and Motif 6 were found in BrDMC1.A03; BoDMC1.C05 contained Motif 1. BrDMC1.A03 and BoDMC1.C05 contained only a portion of the RecA recombinase functional domain, which was predicted to be the fragment DMC1 generated after genome replication (Figure 1C).
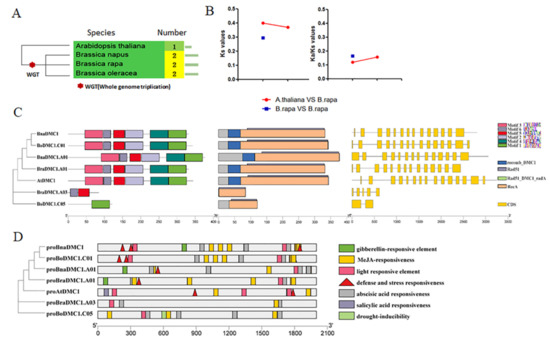
Figure 1.
DMC1 genes following WGT in Brassica species. (A) The number of DMC1 gene copies that Arabidopsis thaliana (A. thaliana) and Brassica species have retained. Phylogenetic tree of DMC1 was constructed in whole protein sequences. (B) The Ka/Ks between BrDMC1 and AtDMC1. (C) DMC1 gene structures and conserved motifs. An unrooted phylogenetic tree was derived from the full-length amino acid alignment of all DMC1 proteins. MEGA7 was used to build the tree, and maximum likelihood (ML) and bootstrap values were calculated with 1000 replications. (D) DMC1 promoter cis-acting element analysis.
There are two copies of the DMC1 gene in B. rapa, BrDMC1.A01 (ID: Bra023796) and BrDMC1.A03 (ID: Bra001890), which is an ortholog of AtDMC1 (ID: AT3G22880). We calculated the Ka/Ks value with reference to AtDMC1 (Figure 1B). The Ks values calculated between BrDMC1 and AtDMC1 were 0.4 and 0.37, and the corresponding differentiation times were 13.26 million years ago and 12.31 million years ago. The time when the WGT replication event occurred in B. rapa was about 13 million years ago, the calculated Ks value between BrDMC1 was 0.29 and the corresponding divergence time was 9.83 million years ago, indicating that the two copies of DMC1 are generated in B. rapa by WGT events. By calculation, the ratio of Ka/Ks between BrDMC1 and AtDMC1 is between 0.12 and 0.16, and the ratio of Ka/Ks < 1 is generally considered to have a lower selection pressure during biological evolution and tends to be conserved in function. It was further proved that the DMC1 of B. rapa did not differentiate significantly in function.
3.2. Interaction Networks of Potential DNA-Repair-related BrDMC1 in B. rapa
To predict the interaction relationship between potential DNA-repair-related DMC1 genes and other genes in B. rapa, we constructed interaction networks based on orthologous genes with known functions in A. thaliana (Figure 2). Through the prediction and analysis of the interaction relationship, we can see that BrDMC1 (Bra023796) has the highest correlation with BRCA2 (Bra037060) and XRCC3 (Bra3037372), and BRCA2 (Bra037060) is a homologous recombinant repair protein, XRCC3 (Bra3037372) is a DNA damage repair protein, which also has a large correlation with DNA repair proteins, RecA (Bra008393 and Bra003623), meiotic nuclear division protein MND1 (Bra024159), ASY1 (Bra004222), etc. BrDMC1 (Bra001890) has the highest correlation with XRCC3 (Bra3037372 and Bra006820) and Rad51 (Bra035691). The above results show that BrDMC1 is likely to participate in DNA damage repair.
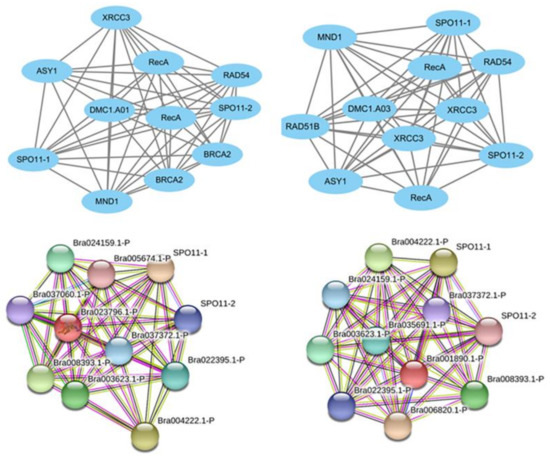
Figure 2.
BrDMC1 interaction network analysis was obtained from the STRING database and built using Cytoscape software.
3.3. GUS Staining Analysis of Putative Promoter Regions of BrDMC1
The upstream 2000 bp genome sequence of the BrDMC1 gene was extracted in order to identify cis-regulatory elements and learn more about the potential function and transcriptional regulation of stress-related BrDMC1 (Figure 1D). It was found that BnaDMC1, BnaDMC1.A01, BrDMC1.A01, BoDMC1 and AtDMC1 gene promoters all contain stress response elements, ABA response elements, light response elements and MeJA response elements. There are few response elements in BrDMC1.A03 and BoDMC1.C05 promoters. The prediction results showed that the promoter region of the DMC1 gene contained more stress response elements. Our results suggest that proBrDMC1.A03 does not have a promoter function, and proBrDMC1.A01 is a normal promoter segment. GUS signal could not be detected in A. thaliana pods, stems, or leaves, but could be detected in flowers and germinating seeds (Figure 3A). The expression pattern of the BrDMC1.A01 gene was preliminarily verified by applying exogenous hormones and stress treatments to N. benthamiana leaves (Figure 3B). The results showed that GUS staining deepened significantly after hormone and salt treatments. It was sensitive to ABA, GA and other hormones, but after SA treatment there was not a significant difference. Semi-quantitative PCR also confirmed the level of GUS expression (Figure 3C).
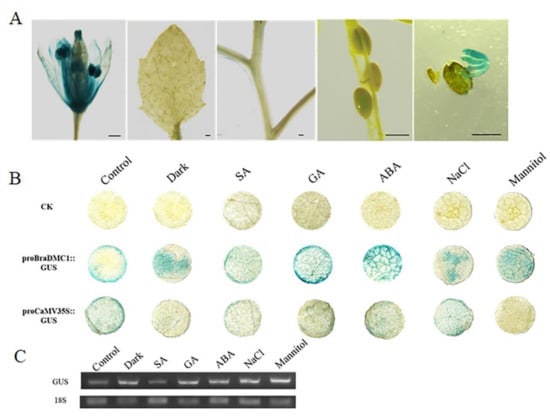
Figure 3.
GUS staining analysis of the BrDMC1 promoter. (A) GUS staining analysis of BrDMC1 promoter in flowers, leaves, stems, pods and seeds of transgenic A. thaliana. Bar = 400 µm. (B) GUS staining of Nicotiana benthamiana (N. benthamiana) leaves after different treatments. (C) The expression of GUS was determined by semi-quantitative PCR.
3.4. Expression Pattern, Salt Stress and Subcellular Localization of BrDMC1
The localization of the BrDMC1 protein has not been determined previously. We examined the subcellular localization of BrDMC1 via the transient expression of 35S::BrDMC1-GFP in N. benthamiana leaves, and the results revealed that BrDMC1 is a nuclear protein (Figure 4A), as the green fluorescence signal was predominately detected in the nucleus. BrDMC1 was localized in the nucleus, indicating that it is involved in DNA repair.
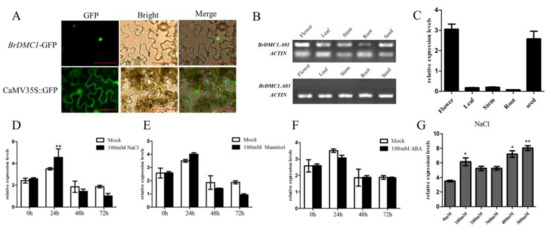
Figure 4.
Expression pattern and subcellular localization of BrDMC1. (A) Subcellular localization of the fused 35S::BrDMC1-GFP in N. benthamiana leaf cells. Bar = 50 µm. (B) Expression pattern of BrDMC1 in different tissues. (C) qRT-PCR analysis expression levels of BrDMC1 in different tissues. (D–G) BrDMC1 expression levels in different treatments and different times. (** p < 0.01, * p < 0.05.)
The level of BrDMC1 expression in different tissues of B. rapa showed that the BrDMC1.A03 (ID: Bra001890) did not have transcriptional activity (Figure 4B). BrDMC1.A01 (ID: Bra023796) is expressed in different plant tissues, the lowest in the roots, the highest in the flowers and the second in the seeds (Figure 4C), which could be crucial in seed development.
In order to characterize the expression mode of BrDMC1 under salt stress, the expression level of BrDMC1 under different treatments (NaCl, mannitol and ABA) was different via qRT-PCR (Figure 4D–F). The finding revealed that BrDMC1 was responsive to salt stress, and no significant difference was observed in ABA and mannitol treatment. BrDMC1 expression peaked 24 h after various stress treatments, and then the expression of BrDMC1 gradually decreased and tended to be gentle. The peak value of expression appeared on the first day after NaCl treatment, about twice the initial level. We designed salt treatments with different concentration gradients to investigate the response of BrDMC1 to different levels of stress (Figure 4G). The results showed that the expression of BrDMC1 was high after 100 mM NaCl, 400 mM NaCl and 500 mM NaCl treatments.
3.5. BrDMC1-RNAi Reduced Salt Tolerance in B. rapa Seed Germination
Seed germination is vulnerable to salt stress. We successfully obtained BrDMC1-RNAi transgenic lines (#1, #3 and #6), which was consistent with RT-PCR results, indicating the successful selection of BrDMC1-RNAi transgenic plants (Figure 5A). We selected WT and BrDMC1-RNAi line seeds to carry out the germination test under salt stress. We observed the seeds treated for 0 h, 24 h, 48 h, 72 h and 96 h and counted the germination rate and morphology. The findings revealed that the germination rate of BrDMC1-RNAi plants significantly decreased after salt stress treatment for 24 h (Figure 5B). In the control with water treatment, there was no difference in the germination rates of WT and BrDMC1-RNAi seeds, but after salt stress treatment, the germination rate of BrDMC1-RNAi seeds decreased significantly, to only about 20%, much lower than the control (Figure 5D–E). The results showed that BrDMC1 expression affected the seed germination rate under salt stress.
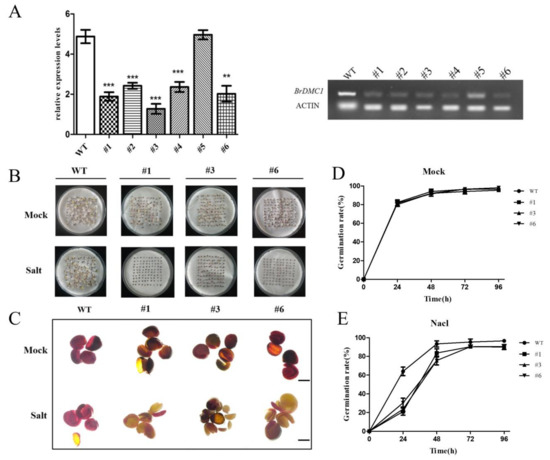
Figure 5.
BrDMC1-RNAi decreased salt tolerance during seed germination. (A) Relative expression level confirmation of the BrDMC1 gene in BrDMC1-RNAi determined via qRT-PCR. (B) Phenotypes of WT and BrDMC1-RNAi seeds in 100 mM NaCl treatment. (C) TTC staining analysis seed vigor. (D) Statistical analysis of germination rate in mock. (E) Statistical analysis of germination rate in 100 mM NaCl. (** p < 0.01 and *** p < 0.001.)
The seed vigor is closely related to the DNA repair ability. The size of seed vigor can reflect the damage and repair ability of seed to a certain extent. The TTC staining method was used to analyze the seeds that germinated for 24 h (Figure 5C). It was found that, under salt stress, the seeds stained shallowly and their vigor weakened. Among them, BrDMC1-RNAi seeds stained the least, suggesting that the repair ability of seeds might be seriously damaged.
3.6. BrDMC1-RNAi Decreased Antioxidant Enzyme Activity under Salt Stress
The major components of ROS active oxygen are O2•− and H2O2. The content of O2•− in germinated seeds was determined for the WT and BrDMC1-RNAi line treated with salt through dyeing depth, according to the dyeing depth of nitroblue tetrazolium (NBT) reduced to determine the O2•− level in the seeds, and excessive reactive oxygen radicals cause damage to the DNA structure. The results showed that after salt treatment, the seeds accumulated a large amount of active oxygen. Compared with the control treatment, BrDMC1-RNAi plants were stained deeper, and the O2•− and H2O2 contents were significantly increased (Figure 6A,B). This implies that BrDMC1-RNAi plants may suffer more severe oxidative damage in germinated seeds.
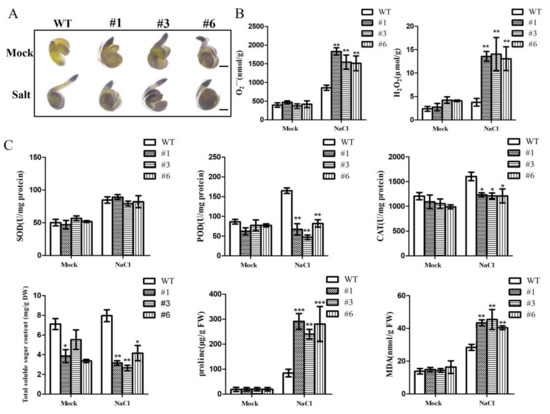
Figure 6.
ROS activity analysis during seed germination. (A) NBT staining analysis content of O2•−. (B) O2•− and H2O2 content analysis. (C) Activities of SOD, POD, CAT, soluble sugar content, proline and MDA. The values represent the means ± SE of three independent experiments. (*** p < 0.001, ** p < 0.01 and * p < 0.05.)
When plants are stressed, they activate the antioxidative stress system, which allows them to remove ROS by increasing the activity of antioxidant enzymes such as SOD, POD and CAT. Proline and soluble sugar accumulate quickly as osmotic protective substances to keep plants stable during stress. The presence of malondialdehyde (MDA) in the membrane can indicate the degree of lipid peroxidation. Therefore, the seeds of WT and BrDMC1-RNAi plants were treated with salt stress during germination for 24 h, and the changes in related antioxidant and osmotic protective substances were measured (Figure 6C). The results showed that, compared with WT, SOD activity was not significantly different, and POD and CAT enzyme activities and soluble sugar content were significantly decreased in BrDMC1-RNAi. The contents of proline and malondialdehyde (MDA) increased obviously. Malondialdehyde (MDA) content rose, indicating an increase in membrane lipid peroxidation. Therefore, we concluded that BrDMC1-RNAi seedlings accumulated too much ROS in seeds under salt stress, and BrDMC1-RNAi seeds suffered serious oxidative damage and eventually had a reduced germination rate.
3.7. BrDMC1 Differently Regulates the Expression of Stress-Related Genes and Damage Repair Genes under Salt Stress
The expression of stress-related genes is considered to be an important indicator of the plant stress response. We chose genes associated with stress and damage repair (SOD, POD, P5C, RAD51, TOP6B, NCED3, XRCC3 and BRCA2) and used qRT-PCR to compare the expression levels of BrDMC1-RNAi and WT seeds under salt stress (Figure 7). The melting curves of the experimental results are shown in Appendix B. There was no significant difference in the expression of the POD gene or the NCED3 gene in BrDMC1-RNAi seeds after salt treatment compared to the WT, but the expression levels of the stress-related TOP6B gene, the DNA damage repair genes XRCC3 and BRCA2 and the key proline synthesis gene P5C were significantly up-regulated. The expressions of the homologous recombinant protein RAD51 gene and the antioxidant SOD gene were both significantly down-regulated. These findings imply that the BrDMC1 gene responds to salt stress via a synergistic action of multiple genes.
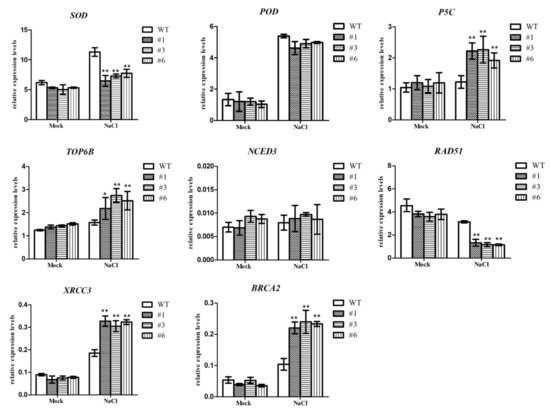
Figure 7.
Expression levels of stress-related genes (SOD, POD, P5C, TOP6B, NCED3, RAD51, XRCC3 and BRCA2) in control plants and BrDMC1-RNAi. The values were means ± SE of three independent experiments. (** p < 0.01 and * p < 0.05.)
4. Discussion
4.1. BrDMC1 Evolved to Retain a Single Copy Function in B.rapa
Following the WGT event, the Brassica genome replicates, and many duplicated genes undergo subfunctionalization or neofunctionalization. There is a theory that the evolution of genes following genome replication is biased, namely the biased retention of Brassica, and this biased retention of genes would further promote the diversity in Brassica morphological types [46,47]. B. napus, B. rapa and B. oleracea are the brassica plants with recent WGT incidents. To investigate the impact of WGT events on DMC1 gene retention, we predicted the copy number and conserved functional domain of the DMC1 gene (Figure 1). The findings revealed that the DMC1 gene had two copies and contained the RecA recombinase functional domain, which could carry out chain exchange reactions and catalyze DNA recombination repair [48]. The expression analysis of two copies of BrDMC1 revealed that BrDMC1.A03 was not transcribed, indicating that BrDMC1 was lost in evolution after the WGT event, leaving only a single copy of BrDMC1.A01 to function (Figure 4). Genes involved in DNA repair are preferentially lost after genome replication [49], implying that gene redundancy after replication may affect normal function. At the same time, most genes are lost, and the remaining copies may acquire new functions, allowing for gene changes [50]. The loss or retention of replicative genes are important aspects of the long-term evolution of polyploids. B. rapa is a common Brassica crop that can be used to study evolution.
4.2. BrDMC1 May Play Essential Roles in Response to Stress
Many classical studies have confirmed that the DNA recombinase DMC1 has the function of repairing double-strand breaks, participates in the recombination process of chromosomal associations and has a significant impact on the fertility of pollen, as can be seen in studies of A. thaliana, rice and barley, but its role in abiotic stress has been less studied [29,51,52,53]. In this study, we used BrDMC1 transgenic plants to investigate their roles in plant growth and development under stress conditions. The expression pattern analysis of the BrDMC1 in different tissues revealed that the expression level of the BrDMC1 was highest in flowers (Figure 4), which was consistent with its function of recombination and repair [52], while the expression level in seeds was second, indicating that the BrDMC1 may play an important role in the process of seed development. We examined the expression of BrDMC1 in plants treated with NaCl, ABA and mannitol (Figure 4). The results showed that BrDMC1 was sensitive to salt stress and that there was no significant difference between ABA and mannitol treatment, which was consistent with the previous study on the DMC1 gene.
RecA mutants in Acinetobacter baumannii were found to be more sensitive to oxidants such as sodium nitroprusside, H2O2 and menadione [54], implying that RecA is involved in homologous recombination and DNA repair, as well as oxidant-induced cell protection. In this study, we discovered that under salt stress, O2•− and H2O2 levels in BrDMC1-RNAi plants were significantly higher than in the WT (Figure 6), indicating that BrDMC1-RNAi plants suffered more severe oxidative damage. Furthermore, we discovered that under salt stress, the ROS detoxification protein content of BrDMC1-RNAi plants decreased while the contents of proline and malondialdehyde significantly increased (Figure 6), indicating that BrDMC1 may also have an oxidation-induced function.
The overexpression of the ZmRAD51A gene in rice increased plant disease resistance, resulting in the increased expression of defense-related genes [35]. In this study, it was discovered that in BrDMC1-RNAi plants, the expression levels of the stress-related TOP6B gene, DNA damage repair genes XRCC3 and BRCA2 and key proline synthesis gene P5C were significantly increased compared to WT, while the expression levels of the homologous recombinant protein RAD51 and antioxidant SOD were significantly down-regulated (Figure 7). These findings imply that the BrDMC1 responds to salt stress via the synergistic action of several genes.
5. Conclusions
In this study, the effects of BrDMC1 on the growth process of B. rapa under salt stress were investigated by measuring physiological and biochemical indexes as well as molecular expression levels. The findings revealed that the silent transgenic plants were susceptible to salt stress. These findings suggest that, in addition to its functions of homologous recombination and damage repair, BrDMC1 is involved in abiotic stress. These findings are important for furthering the study of stress response in plants, and it is necessary to investigate and clarify the molecular mechanism of DMC1 involved in stress response.
Author Contributions
Conceptualization, X.W. (Xulin Wang); data curation, Z.T., B.T. and X.W. (Xiaochun Wei); formal analysis, Z.T., W.C., X.W. (Xiaochun Wei) and L.Z.; funding acquisition, F.W.; investigation, S.W.; methodology, X.W. (Xulin Wang) and F.W.; project administration, F.W.; validation, Z.X. and G.S.; visualization, G.S., G.C. and L.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Henan Provincial Natural Science Foundation of China (No. 202300410366), the Youth Innovation Project of Key Discipline of Zhengzhou University (No. XKZDQN202002) and the Fostering Project for Basic Research of Zhengzhou University (No. JC21310015).
Data Availability Statement
All data generated in this study are included in this article.
Acknowledgments
We would like to express our thanks to the anonymous reviewers for their useful comments.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

Table A1.
Information of primer sequences.
Table A1.
Information of primer sequences.
| Primer Name | Primer Sequence (5′ to 3′) |
|---|---|
| qBrDMC1.A01-F | GGAGGTGGGATTGAGACTGC |
| qBrDMC1.A01-R | GGACAATCCTATCAGGGCGG |
| qBrDMC1.A03-F | TCACTACTGGTTGCCAAGCTC |
| qBrDMC1.A03-R | TAAGCCACTTTTCCGTTCCCT |
| β-antin-F | ATCAACTACCAGCCTCCAAC |
| β-antin-R | CTGCTGTGTTGTTGCTGATC |
| ProBrDMC1.A01-F | AAGCAAATGTGTGCTGTGAAC |
| ProBrDMC1.A01-R | GAAGCGAGCAGAGATCGAAG |
| ProBrDMC1.A03-F | ATATGCACACCAAGTGAGACAG |
| ProBrDMC1.A03-R | CATCAAGAGCTTGGCAACCAG |
| GUS-F | TACCGACGAAAACGGCAAGA |
| GUS-R | ATGCCATGTTCATCTGCCCA |
| BrDMC1.A01-F | ATGCTTTCTGCTCTCAAATC |
| BrDMC1.A03-R | TTAAAAGGATATAGCTTCGC |
| BrDMC1(RNAi)-BP-F | TGTAATGGTCTCATGATGCATACCAAAAAGAACCTCACTGGAATC |
| BrDMC1(RNAi)-BP-R | CAGTGTCAATGTAAGCCACTTTCCCATTCCCACCTTTCATGCTTG |
| PDK(RNAi)-F | GACGAAGAAGATAAAAGTTGAGAGT |
| PDK(RNAi)-R | ACCTTGTTTATTCATGTTCGACTAA |
| P5C-F | GAACCGTACCTTCTCAAACT |
| P5C-R | CTCCATCGGTATCTGATGTT |
| TOP6B-F | CTCGTGGAAAGTTTGGTTTA |
| TOP6B-R | CTGTATTTCGGCTCCATGCC |
| NCED3-F | TCACCACTTCTTCGACGGAG |
| NCED3-R | GGTCGACTAAACCGGCTGCG |
| RAD51-F | GCAGCTTCAAAGCTGGTTCC |
| RAD51-R | CAGCTCCATTTAGTCCAAAC |
| XRCC3-F | GCTCTCGCGATCGTTTCATC |
| XRCC3-R | CACAGTTAGCCCAAGCCAATC |
| BRCA2-F | TCGCAATGATATAGCTCATG |
| BRCA2-R | CGTAGCTCTTGGACGCATTC |
Appendix B

Figure A1.
The melting curves of related genes (SOD, POD, P5C, TOP6B, NCED3, RAD51, XRCC3 and BRCA2) were analyzed via qRT-PCR.
Figure A1.
The melting curves of related genes (SOD, POD, P5C, TOP6B, NCED3, RAD51, XRCC3 and BRCA2) were analyzed via qRT-PCR.

References
- Wang, L.; Ma, R.; Yin, Y.; Jiao, Z. Role of Carbon Ion Beams Irradiation in Mitigating Cold Stress in Arabidopsis Thaliana. Ecotoxicol. Environ. Saf. 2018, 162, 341–347. [Google Scholar] [CrossRef]
- Jafari, A.; Kamarehie, B.; Ghaderpoori, M.; Khoshnamvand, N.; Birjandi, M. The Concentration Data of Heavy Metals in Iranian Grown and Imported Rice and Human Health Hazard Assessment. Data Brief 2018, 16, 453–459. [Google Scholar] [CrossRef]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.Y.; Li, J.; Wang, P.Y.; Qin, F.; et al. Plant Abiotic Stress Response and Nutrient Use Efficiency. Sci. China Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef]
- Morton, M.J.L.; Awlia, M.; Al-Tamimi, N.; Saade, S.; Pailles, Y.; Negrão, S.; Tester, M. Salt Stress under the Scalpel—Dissecting the Genetics of Salt Tolerance. Plant J. 2019, 97, 148–163. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Su, T.; Li, X.; Yang, M.; Shao, Q.; Zhao, Y.; Ma, C.; Wang, P. Autophagy: An Intracellular Degradation Pathway Regulating Plant Survival and Stress Response. Front. Plant Sci. 2020, 11, 164. [Google Scholar] [CrossRef]
- Horie, T.; Karahara, I.; Katsuhara, M. Salinity Tolerance Mechanisms in Glycophytes: An Overview with the Central Focus on Rice Plants. Rice 2012, 5, 11. [Google Scholar] [CrossRef]
- Quan, R.; Lin, H.; Mendoza, I.; Zhang, Y.; Cao, W.; Yang, Y.; Shang, M.; Chen, S.; Pardo, J.M.; Guo, Y. SCABP8/CBL10, a Putative Calcium Sensor, Interacts with the Protein Kinase SOS2 to Protect Arabidopsis Shoots from Salt Stress. Plant Cell 2007, 19, 1415–1431. [Google Scholar] [CrossRef]
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef]
- Waterworth, W.M.; Bray, C.M.; West, C.E. The Importance of Safeguarding Genome Integrity in Germination and Seed Longevity. J. Exp. Bot. 2015, 66, 3549–3558. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive Oxygen Species, Abiotic Stress and Stress Combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of Plant Responses to Salt Stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef]
- Khan, T.A.; Fariduddin, Q.; Yusuf, M. Low-Temperature Stress: Is Phytohormones Application a Remedy? Environ. Sci. Pollut. Res. 2017, 24, 21574–21590. [Google Scholar] [CrossRef]
- Waterworth, W.M.; Footitt, S.; Bray, C.M.; Finch-Savage, W.E.; West, C.E. DNA Damage Checkpoint Kinase ATM Regulates Germination and Maintains Genome Stability in Seeds. Proc. Natl. Acad. Sci. USA 2016, 113, 9647–9652. [Google Scholar] [CrossRef]
- Waterworth, W.M.; Masnavi, G.; Bhardwaj, R.M.; Jiang, Q.; Bray, C.M.; West, C.E. A Plant DNA Ligase Is an Important Determinant of Seed Longevity. Plant J. 2010, 63, 848–860. [Google Scholar] [CrossRef]
- Waterworth, W.M.; Drury, G.E.; Bray, C.M.; West, C.E. Repairing Breaks in the Plant Genome: The Importance of Keeping It Together. New Phytol. 2011, 192, 805–822. [Google Scholar] [CrossRef]
- Powell, A.; Matthews, S. Association of Official Seed Analysts Society of Commercial Seed Technologists (SCST) Seed Aging/Repair Hypothesis Leads to New Testing Methods. Seed Technol. 2012, 3, 2. [Google Scholar]
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Bally, J.; Job, C.; Job, D. Seed Germination and Vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef]
- Dandoy, E.; Schnys, R.; Deltour, R.; Verly, W.G. Appearance and Repair of Apurinic/Apyrimidinic Sites in DNA during Early Germination of Zea Mays. Mutat. Res. Fundam. Mol. Mech. Mutagen. 1987, 181, 57–60. [Google Scholar] [CrossRef]
- Waterworth, W.M.; Latham, R.; Wang, D.; Alsharif, M.; West, C.E. Seed DNA Damage Responses Promote Germination and Growth in Arabidopsis Thaliana. Proc. Natl. Acad. Sci. USA 2022, 119, 1–8. [Google Scholar] [CrossRef]
- Navashin, M.; Shkvarnikov, P. Process of Mutation in Resting Seeds Accelerated by Increased Temperature. Nature 1933, 132, 482–483. [Google Scholar] [CrossRef]
- Sequencing, G.; Consortium, P. The Genome of the Mesopolyploid Crop Species Brassica Rapa. Nat. Genet. 2011, 43, 1035–1040. [Google Scholar] [CrossRef]
- Lyu, T.; Liu, W.; Hu, Z.; Xiang, X.; Liu, T.; Xiong, X.; Cao, J. Molecular Characterization and Expression Analysis Reveal the Roles of Cys2/His2 Zinc-Finger Transcription Factors during Flower Development of Brassica Rapa Subsp. Chinensis. Plant Mol. Biol. 2020, 102, 123–141. [Google Scholar] [CrossRef]
- Devisetty, U.K.; Mayes, K.; Mayes, S. The RAD51 and DMC1 Homoeologous Genes of Bread Wheat: Cloning, Molecular Characterization and Expression Analysis. BMC Res. Notes 2010, 3, 245. [Google Scholar] [CrossRef]
- Prentiss, M.; Prévost, C.; Danilowicz, C.; Prentiss, M.; Prévost, C.; Structure, C.D.; Danilowicz, C.; Prentiss, M.; Pre, C. Structure/Function Relationships in RecA Protein- Mediated Homology Recognition and Strand Exchange Recognition and Strand Exchange. Crit. Rev. Biochem. Mol. Biol. 2015, 50, 453–476. [Google Scholar] [CrossRef]
- Steinfeld, J.B.; Belán, O.; Kwon, Y.; Terakawa, T.; Al-zain, A.; Smith, M.J.; Crickard, J.B.; Qi, Z.; Zhao, W.; Rothstein, R.; et al. Defining the Influence of Rad51 and Dmc1 Lineage-Specific Amino Acids on Genetic Recombination. Genes Dev. 2019, 33, 1191–1207. [Google Scholar] [CrossRef]
- Lan, W.; Lin, S.; Kao, C.; Chang, W.; Yeh, H.; Chang, H. Rad51 Facilitates Filament Assembly of Meiosis-Specific. Proc. Natl. Acad. Sci. USA 2020, 117, 11257–11264. [Google Scholar] [CrossRef]
- Li, T.; Shi, Q.; Davies, B.; Donnelly, P.; Hinch, A.G.; Becker, P.W.; Li, T.; Moralli, D.; Zhang, G.; Bycroft, C.; et al. Article The Configuration of RPA, RAD51, and DMC1 Binding in Meiosis Reveals the Nature of Critical Recombination Intermediates Ll The Configuration of RPA, RAD51, and DMC1 Binding in Meiosis Reveals the Nature of Critical Recombination Intermediates. Mol. Cell 2020, 79, 689–701.e10. [Google Scholar] [CrossRef]
- Szurman-Zubrzycka, M.; Baran, B.; Stolarek-Januszkiewicz, M.; Kwaśniewska, J.; Szarejko, I.; Gruszka, D. The Dmc1 Mutant Allows an Insight into the DNA Double-Strand Break Repair during Meiosis in Barley (Hordeum Vulgare L.). Front. Plant Sci. 2019, 10, 761. [Google Scholar] [CrossRef]
- Couteau, F.; Belzile, F.; Horlow, C.; Grandjean, O.; Vezon, D.; Doutriaux, M.P. Random Chromosome Segregation without Meiotic Arrest in Both Male and Female Meiocytes of a Dmc1 Mutant of Arabidopsis. Plant Cell 1999, 11, 1623–1634. [Google Scholar] [CrossRef]
- Akada, I.S. Characterization of a DMC1 Homologue, RiLIM15, in Meiotic Panicles, Mitotic Cultured Cells and Mature Leaves of Rice (Oryza Sativa L.). Theor. Appl. Genet. 2001, 102, 1159–1163. [Google Scholar]
- Draeger, T.; Martin, A.C.; Kader, A.; Ali, A.; María, P.; Rey, D.; Shaw, P.; Moore, G. Dmc1 Is a Candidate for Temperature Tolerance during Wheat Meiosis. Theor. Appl. Genet. 2020, 133, 809–828. [Google Scholar] [CrossRef]
- Jeon, H.; Jin, Y.; Choi, M.H.; Lee, H.; Kim, M. Chloroplast-Targeted Bacterial RecA Proteins Confer Tolerance to Chloroplast DNA Damage by Methyl Viologen or UV-C Radiation in Tobacco (Nicotiana Tabacum) Plants. Physiol. Plant. 2013, 147, 218–233. [Google Scholar] [CrossRef]
- Muris, D.F.R.; Vreeken, K.; Carr, A.M.; Broughton, B.C.; Lehmann, A.R.; Lohman, P.H.M.; Pastink, A. Cloning the RAD51 Homologue of Schizosaccharomyces Pombe. Nucleic Acids Res. 1993, 21, 4586–4591. [Google Scholar] [CrossRef]
- Liu, F.; Xu, Y.; Zhou, L.; Ali, A.; Jiang, H.; Zhu, S.; Li, X. DNA Repair Gene ZmRAD51A Improves Rice and Arabidopsis Resistance to Disease. Int. J. Mol. Sci. 2019, 20, 807. [Google Scholar] [CrossRef]
- Cao, G.; Jiang, W.; Shi, G.; Tian, Z.; Shang, J.; Xie, Z.; Chen, W.; Tian, B.; Wei, X.; Wei, F.; et al. Br PARP1, a Poly (ADP-Ribose) Polymerase Gene, Is Involved in Root Development in Brassica Rapa under Drought Stress. Horticulturae 2022, 8, 78. [Google Scholar] [CrossRef]
- Cao, G.; Gu, H.; Jiang, W.; Tian, Z.; Shi, G.; Chen, W.; Tian, B.; Wei, X.; Zhang, L.; Wei, F.; et al. BrDHC1, a Novel Putative DEAD-Box Helicase Gene, Confers Drought Tolerance in Transgenic Brassica Rapa. Horticulturae 2022, 8, 707. [Google Scholar] [CrossRef]
- Sparkes, I.A.; Runions, J.; Kearns, A.; Hawes, C. Rapid, Transient Expression of Fluorescent Fusion Proteins in Tobacco Plants and Generation of Stably Transformed Plants. Nat. Protoc. 2006, 1, 2019–2025. [Google Scholar] [CrossRef]
- Wang, C.Q.; Guthrie, C.; Sarmast, M.K.; Dehesh, K. BBX19 Interacts with CONSTANS to Repress FLOWERING LOCUS T Transcription, Defining a Flowering Time Checkpoint in Arabidopsis. Plant Cell 2014, 26, 3589–3602. [Google Scholar] [CrossRef]
- Zhang, X.; Henriques, R.; Lin, S.-S.; Niu, Q.-W.; Chua, N.-H. Agrobacterium-Mediated Transformation of Arabidopsis Thaliana Using the Floral Dip Method. Nat. Protoc. 2006, 1, 641–646. [Google Scholar] [CrossRef]
- Leclercq, J.; Martin, F.; Sanier, C.; Clément-Vidal, A.; Fabre, D.; Oliver, G.; Lardet, L.; Ayar, A.; Peyramard, M.; Montoro, P. Over-Expression of a Cytosolic Isoform of the HbCuZnSOD Gene in Hevea Brasiliensis Changes Its Response to a Water Deficit. Plant Mol. Biol. 2012, 80, 255–272. [Google Scholar] [CrossRef]
- Kong, W.; Liu, F.; Zhang, C.; Zhang, J.; Feng, H. Non-Destructive Determination of Malondialdehyde (MDA) Distribution in Oilseed Rape Leaves by Laboratory Scale NIR Hyperspectral Imaging. Sci. Rep. 2016, 6, 35393. [Google Scholar] [CrossRef]
- Ahmad, H.; Hayat, S.; Ali, M.; Liu, T.; Cheng, Z. The Combination of Arbuscular Mycorrhizal Fungi Inoculation (Glomus Versiforme) and 28-Homobrassinolide Spraying Intervals Improves Growth by Enhancing Photosynthesis, Nutrient Absorption, and Antioxidant System in Cucumber (Cucumis Sativus L.) under Sali. Ecol. Evol. 2018, 8, 5724–5740. [Google Scholar] [CrossRef]
- Michaelidou, K.; Tzovaras, A.; Missitzis, I.; Ardavanis, A.; Scorilas, A. The Expression of the CEACAM19 Gene, a Novel Member of the CEA Family, Is Associated with Breast Cancer Progression. Int. J. Oncol. 2013, 42, 1770–1777. [Google Scholar] [CrossRef]
- Freeling, M. Bias in Plant Gene Content Following Different Sorts of Duplication: Tandem, Whole-Genome, Segmental, or by Transposition. Annu. Rev. Plant Biol. 2009, 60, 433–453. [Google Scholar] [CrossRef]
- Cheng, F.; Wu, J.; Wang, X. Genome Triplication Drove the Diversification of Brassica Plants. Hortic. Res. 2014, 1, 14024. [Google Scholar] [CrossRef]
- Wu, P.; Wang, W.; Duan, W.; Li, Y.; Hou, X. Comprehensive Analysis of the CDPK-SnRK Superfamily Genes in Chinese Cabbage and Its Evolutionary Implications in Plants. Front. Plant Sci. 2017, 8, 162. [Google Scholar] [CrossRef]
- McGrew, D.A.; Knight, K.L. Molecular Design and Functional Organization of the RecA Protein. Crit. Rev. Biochem. Mol. Biol. 2003, 38, 385–432. [Google Scholar] [CrossRef]
- Blanc, G.; Wolfe, K.H. Functional Divergence of Duplicated Genes Formed by Polyploidy during Arabidopsis Evolution. Plant Cell 2004, 16, 1679–1691. [Google Scholar] [CrossRef]
- Rensing, S.A. Gene Duplication as a Driver of Plant Morphogenetic Evolution. Curr. Opin. Plant Biol. 2014, 17, 43–48. [Google Scholar] [CrossRef]
- Wang, H.; Hu, Q.; Tang, D.; Liu, X.; Du, G.; Shen, Y.; Li, Y.; Cheng, Z. OsDMC1 Is Not Required for Homologous Pairing in Rice Meiosis. Plant Physiol. 2016, 171, 230–241. [Google Scholar] [CrossRef]
- Kurzbauer, M.T.; Uanschou, C.; Chen, D.; Schlögelhofer, P. The Recombinases DMC1 and RAD51 Are Functionally and Spatially Separated during Meiosis in Arabidopsis. Plant Cell 2012, 24, 2058–2070. [Google Scholar] [CrossRef]
- Kobayashi, W.; Liu, E.; Ishii, H.; Matsunaga, S.; Schlögelhofer, P.; Kurumizaka, H. Homologous Pairing Activities of Arabidopsis Thaliana RAD51 and DMC1. J. Biochem. 2019, 165, 289–295. [Google Scholar] [CrossRef]
- Aranda, J.; Bardina, C.; Beceiro, A.; Rumbo, S.; Cabral, M.P.; Barbé, J.; Bou, G. Acinetobacter Baumannii Reca Protein in Repair of DNA Damage, Antimicrobial Resistance, General Stress Response, and Virulence. J. Bacteriol. 2011, 193, 3740–3747. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).