Stimulative Effects of Lupinus sp. and Melilotus albus Underseed on the Photosynthetic Performance of Maize (Zea mays) in Two Intercropping Systems

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Experiment Design
- (1).
- Monoculture A: maize (Zea mays), the treatment abbreviated SM
- (2).
- Mixed maize and white sweetclover (Melilotus albus), the treatment abbreviated MM
- (3).
- Mixed maize and white lupin (Lupinus sp.), the treatment abbreviated ML
2.2. Soil Characteristics
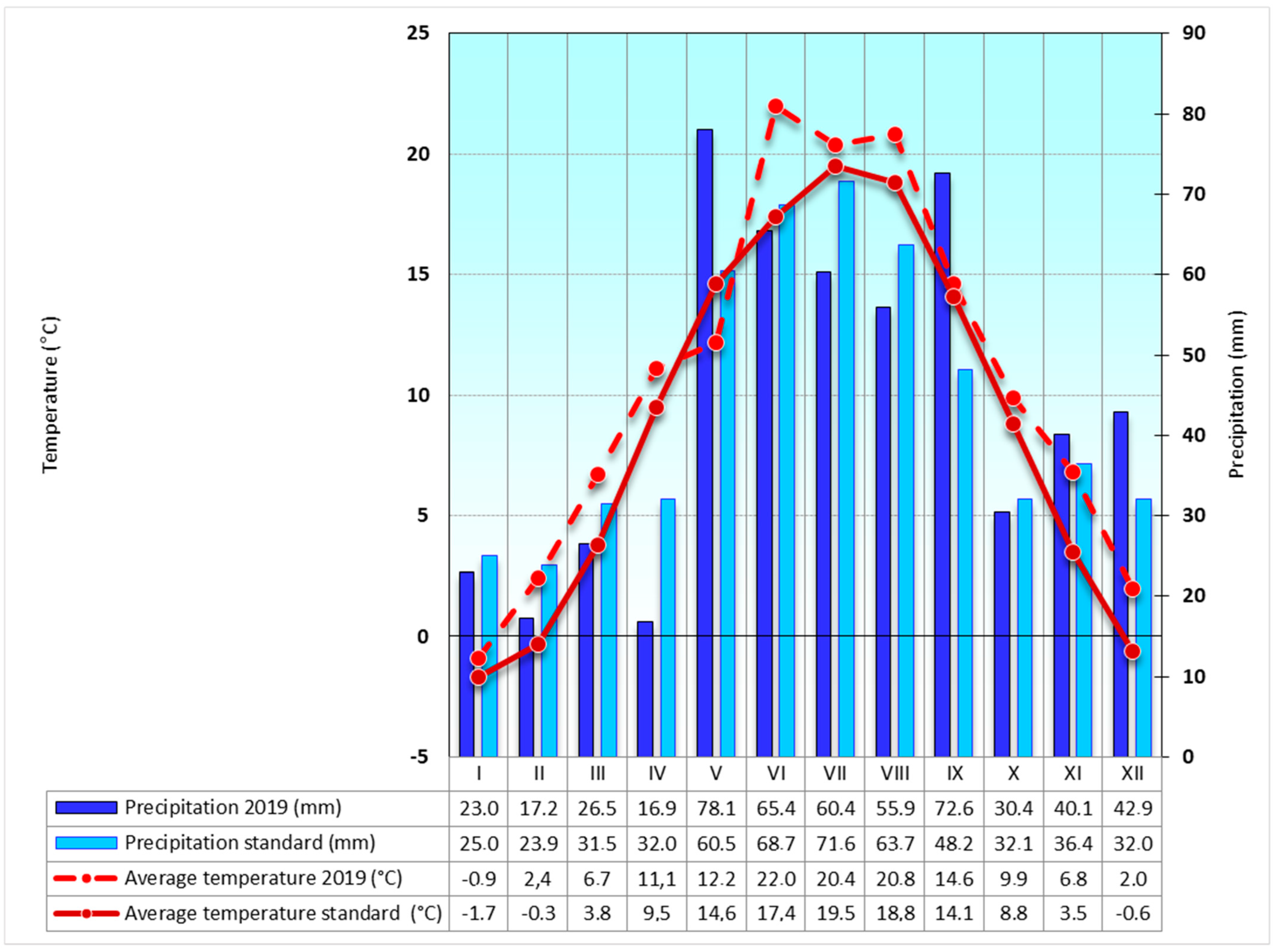
2.3. Local Climate
2.4. Measurements of Microclimate
2.5. Measurements of OJIPs and OJIP-Derived Parameters
2.6. Light Response Curve Analysis
2.7. Biomass Determination
2.8. Statistical Analysis
3. Results
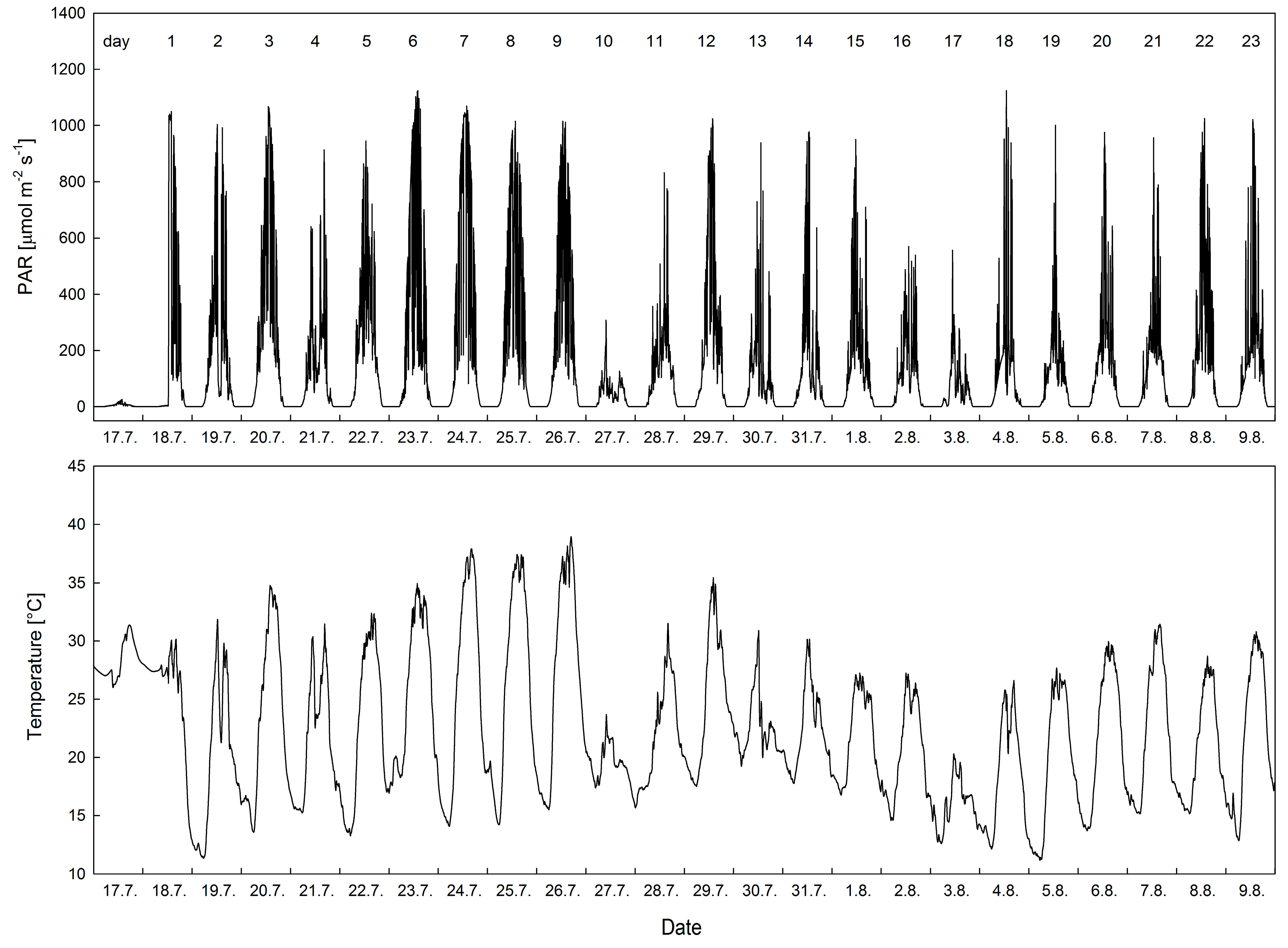
3.1. Local Microclimate
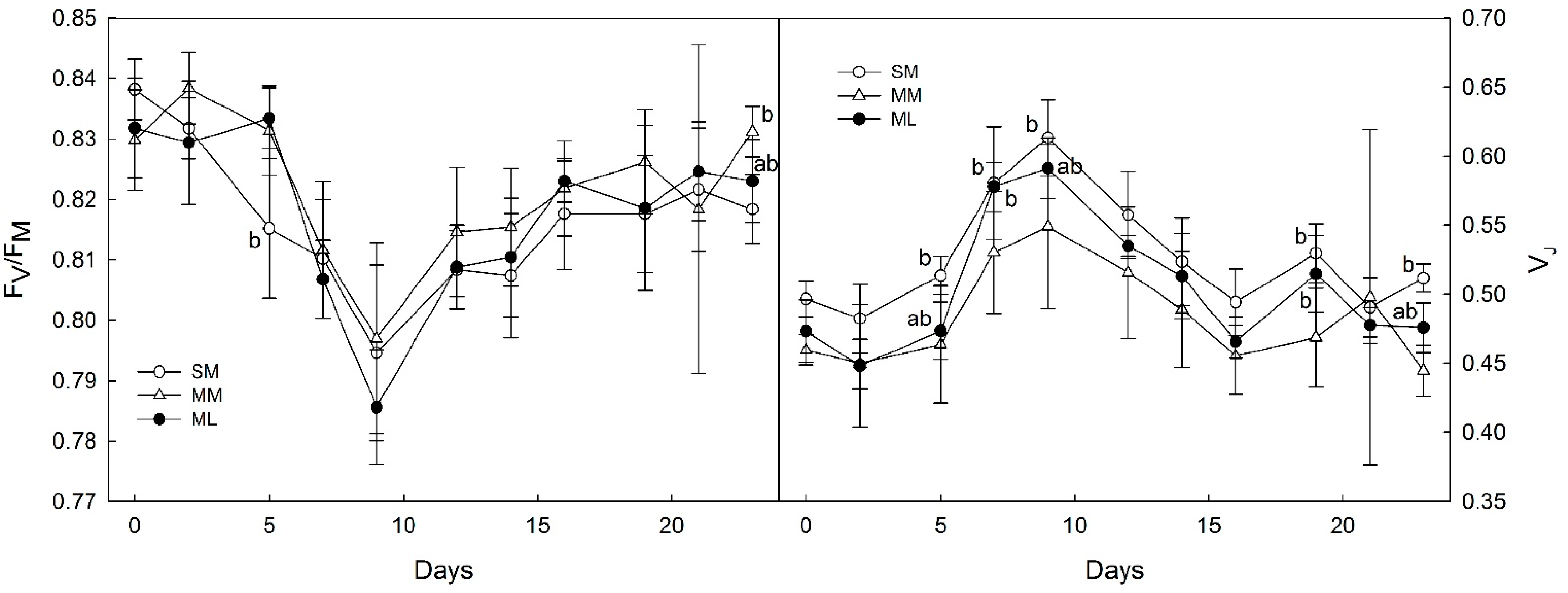
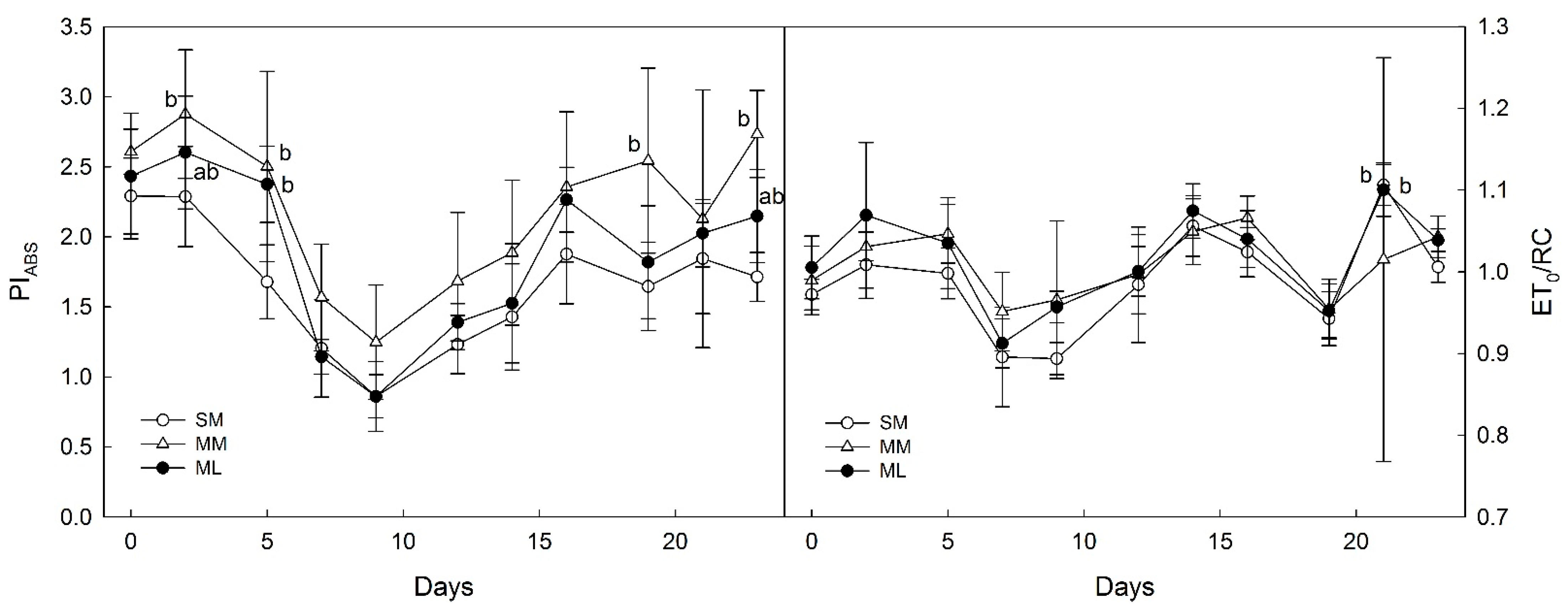
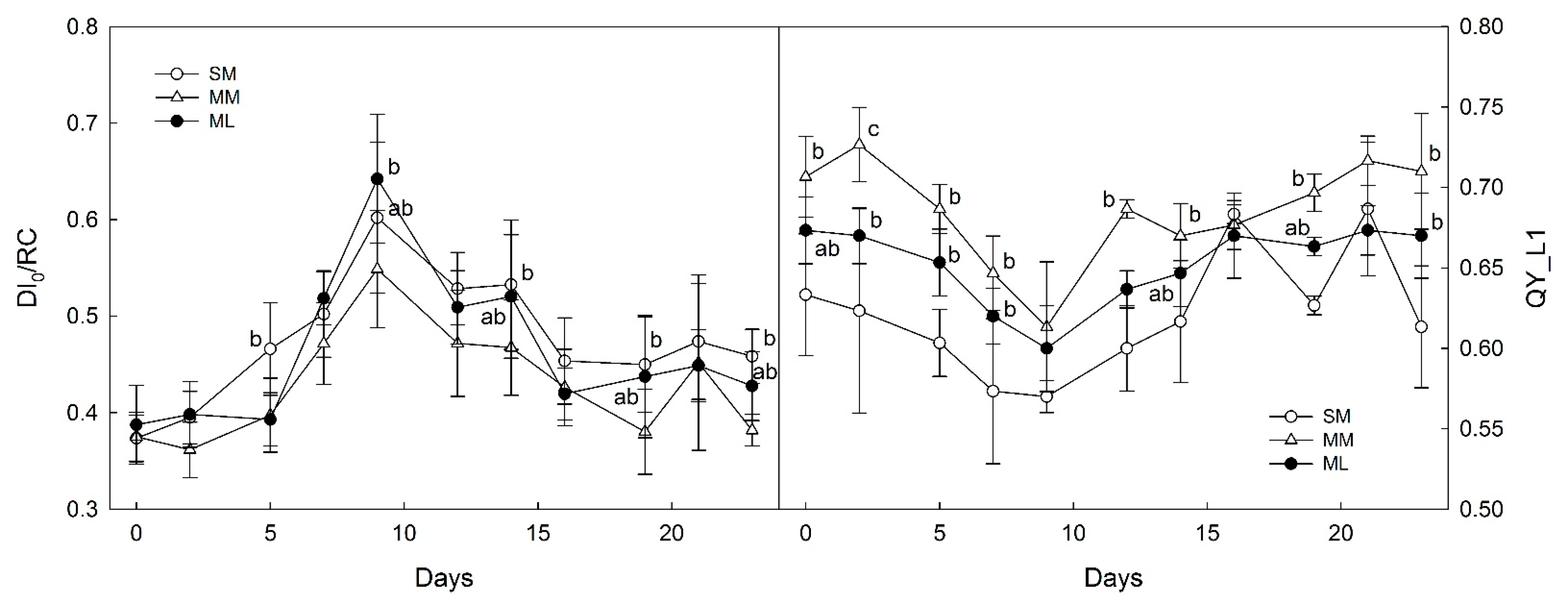
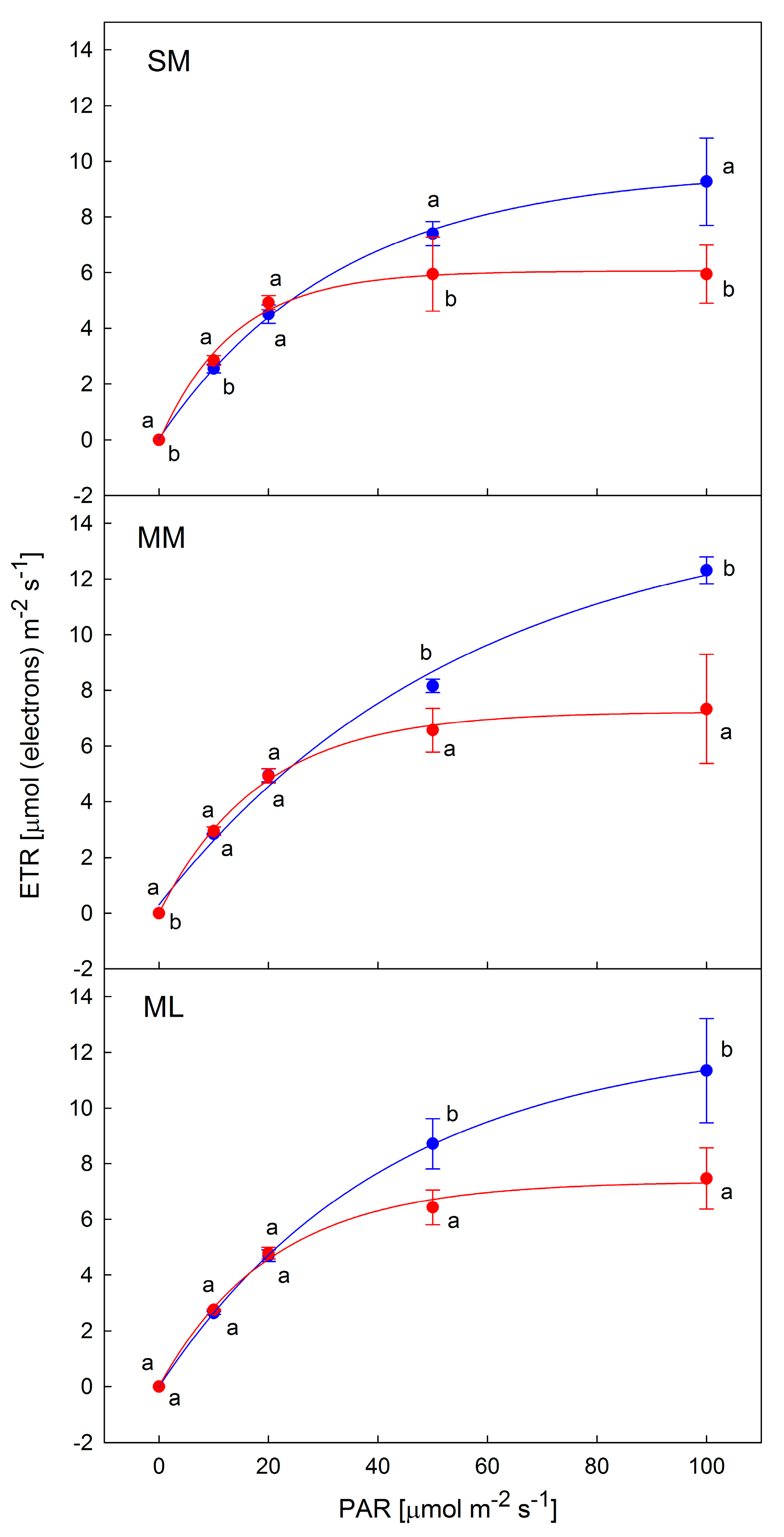
3.2. OJIP-Derived Parameters of Photosynthesis
4. Discussion
4.1. Microclimate
4.2. Photosynthetic Parameters Derived from OJIPs
4.3. Light Response Curves of ETR
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schulz, V.S.; Schumann, C.; Weisenburger, S.; Müller-Lindenlauf, M.; Stolzenburg, K.; Möller, K. Row-intercropping maize (Zea mays L.) with biodiversity-enhancing flowering-partners—Effect on plant growth, silage yield, and composition of harvest material. Agriculture 2020, 10, 524. [Google Scholar] [CrossRef]
- Kintl, A.; Vítěz, T.; Elbl, J.; Vítězová, M.; Dokulilová, T.; Nedělník, J.; Skladanka, J.; Brtnický, M. Mixed culture of corn and white lupine as an alternative to silage made from corn monoculture intended for biogas production. BioEnerg. Res. 2019, 12, 694–702. [Google Scholar] [CrossRef]
- Kintl, A.; Elbl, J.; Vítěz, T.; Brtnický, M.; Skládanka, J.; Hammerschmiedt, T.; Vítězová, M. Possibilities of using white sweetclover grown in mixture with maize for biomethane production. Agronomy 2020, 10, 1407. [Google Scholar] [CrossRef]
- Turkington, R.; Cavers, P.; Rempel, E. The biology of Canadian weeds. 29. Melilotus alba Desr. and M. officinalis (L.) Lam. Can. J. Plant Sci. 1978, 58, 523–537. [Google Scholar] [CrossRef]
- Kadaňková, P.; Kintl, A.; Koukalová, V.; Kučerová, J.; Brtnický, M. Coumarin content in silages made of mixed cropping biomass comprising maize and white sweet clover. In Proceedings of the 19th International Multidisciplinary Scientific GeoConference, Sofia: Surveying Geology & Mining Ecology Management (SGEM) 2019, Albena, Bulgaria, 28 June 2019; pp. 115–121. [Google Scholar] [CrossRef]
- Kintl, A.; Huňady, I.; Holátko, J.; Vítěz, T.; Hammerschmiedt, T.; Brtnický, M.; Ondrisková, V.; Elbl, J. Using the mixed culture of fodder mallow (Malva verticillate L.) and white sweet clover (Melilotus albus Medik.) for methane production. Fermentation 2022, 8, 94. [Google Scholar] [CrossRef]
- Suárez, J.C.; Anzola, J.A.; Contreras, A.T.; Salas, D.L.; Vanegas, J.I.; Urban, M.O.; Beebe, S.E.; Rao, I.M. Influence of simultaneous intercropping of maize-bean with input of inorganic or organic fertilizer on growth, development, and dry matter partitioning to yield components of two lines of common bean. Agronomy 2022, 12, 1216. [Google Scholar] [CrossRef]
- Wang, Q.; Sun, D.; Hao, H.; Zhao, X.; Hao, W.; Liu, Q. Photosynthetically active radiation determining yields for an intercrop of maize with cabbage. Eur. J. Agron. 2015, 69, 32–40. [Google Scholar] [CrossRef]
- Li, Y.H.; Shi, D.Y.; Li, G.H.; Zhao, B.; Zhang, J.W.; Liu, P.; Ren, B.Z.; Dong, S.T. Maize/peanut intercropping increases photosynthetic characteristics, 13C-photosynthate distribution, and grain yield of summer maize. J. Integr. Agric. 2019, 18, 2219–2229. [Google Scholar] [CrossRef]
- Ofori, F.; Stern, W. Relative sowing time and density of component crops in a maize/cowpea intercrop system. Exp. Agric. 1987, 23, 41–52. [Google Scholar] [CrossRef]
- Hauggaard-Nielsen, H.; Andersen, M.K. Intercropping grain legumes and cereals in organic cropping systems. Grain Legumes 2000, 30, 18–19. [Google Scholar]
- Brooker, R.W.; Bennett, A.E.; Cong, W.-F.; Daniell, T.J.; George, T.S.; Hallett, P.D.; Hawes, C.; Iannetta, P.P.M.; Jones, H.G.; Karley, A.J.; et al. Improving intercropping: A synthesis of research in agronomy, plant physiology and ecology. New Phytol. 2015, 206, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Liu, Z.; Han, S.; Jiang, H.; Xu, S.; Zhao, H.; Ren, S. Using the diurnal variation characteristics of effective quantum yield of PSII photochemistry for drought stress detection in maize. Ecol. Indic. 2022, 138, 108842. [Google Scholar] [CrossRef]
- Chen, F.; Wang, H.; Zhao, F.; Wang, R.; Qi, Y.; Zhang, K.; Zhao, H.; Tang, G.; Yang, Y. The response mechanism and threshold of spring wheat to rapid drought. Atmosphere 2022, 13, 596. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fuorescence. Advances in Photosynthesis and Respiration; Govindjee, P.G.C., Ed.; Springer: Dordrecht, The Netherlands, 2004; Volume 19, pp. 321–362. [Google Scholar]
- Malan, C.; Berner, J.M. Comparative PSII photochemistry of quinoa and maize under mild to severe drought stress. Photosynthetica 2022, 60, 362–371. [Google Scholar] [CrossRef]
- Barboričová, M.; Filaček, A.; Mlynáriková Vysoká, D.; Gašparovič, K.; Živčák, M.; Brestič, M. Sensitivity of fast chlorophyll fluorescence parameters to combined heat and drought stress in wheat genotypes. Plant Soil Environ. 2022, 68, 309–316. [Google Scholar] [CrossRef]
- Yan, K.; Chen, P.; Shao, H.; Zhao, S.; Zhang, L.; Zhang, L.; Xu, G.; Sun, J. Responses of photosynthesis and photosystem ii to higher temperature and salt stress in Sorghum. J. Agron. Crop Sci. 2012, 198, 218–225. [Google Scholar] [CrossRef]
- Khatri, K.; Rathore, M.S. Salt and osmotic stress-induced changes in physio-chemical responses, PSII photochemistry and chlorophyll a fluorescence in peanut. Plant Stress 2022, 3, 100063. [Google Scholar] [CrossRef]
- Zhu, L.; Wen, W.; Thorpe, M.R.; Hocart, C.H.; Song, X. Combining heat stress with pre-existing drought exacerbated the effects on chlorophyll fluorescence rise kinetics in four contrasting plant species. Int. J. Mol. Sci. 2021, 22, 10682. [Google Scholar] [CrossRef]
- Ayyaz, A.; Farooq, M.A.; Dawood, M.; Majid, A.; Javed, M.; Athar, H.U.; Bano, H.; Zafar, Z.U. Exogenous melatonin regulates chromium stress-induced feedback inhibition of photosynthesis and antioxidative protection in Brassica napus cultivars. Plant Cell Rep. 2021, 40, 2063–2080. [Google Scholar] [CrossRef]
- Kožnarová, V.; Klabzuba, J. Recommendation of World Meteorological Organization to describing meteorological or climatological conditions. Plant Prod./Rostl. Výroba 2002, 48, 190–192. [Google Scholar] [CrossRef]
- Jedmowski, C.; Brüggemann, W. Imaging of fast chlorophyll fluorescence induction curve (OJIP) parameters, applied in a screening study with wild barley (Hordeum spontaneum) genotypes under heat stress. J. Photoch. Photobiol. B 2015, 151, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yang, J.; Zhang, M.; Strasser, R.J.; Qiang, S. Classification and characteristics of heat tolerance in Ageratina adenophora populations using fast chlorophyll a fluorescence rise O-J-I-P. Environ. Exp. Bot. 2016, 122, 126–140. [Google Scholar] [CrossRef]
- Choi, H.G.; Jeong, H.J. Comparison of chlorophyll fluorescence and photosynthesis of two strawberry cultivars in response to relative humidity. Hortic. Sci. Technol. 2020, 38, 66–77. [Google Scholar] [CrossRef]
- Liu, T.D.; Song, F.B. Maize photosynthesis and microclimate within the canopies at grain-filling stage in response to narrow-wide row planting patterns. Photosynthetica 2012, 50, 215–222. [Google Scholar] [CrossRef]
- Gao, Y.; Duan, A.; Qiu, X.; Sun, J.; Zhang, J.; Liu, H.; Wang, H. Distribution and use efficiency of photosynthetically active radiation in strip intercropping of maize and soybean. Agron. J. 2010, 102, 1149–1157. [Google Scholar] [CrossRef]
- Keshavamurthy; Yadav, J.S. Effect of row direction and row spacing on micro-climate in castor based intercropping system. J. Pharmacogn. Phytochem. 2019, 8, 2167–2170. [Google Scholar]
- Pelech, E.A.; Alexander, B.C.S.; Bernacchi, C.J. Photosynthesis, yield, energy balance, and water-use of intercropped maize and soybean. Plant Direct 2021, 5, e365. [Google Scholar] [CrossRef]
- Mao, L.; Zhang, L.; Li, W.; van der Werf, W.; Sun, J.; Spiertz, H.L.; Li, L. Yield advantage and water saving in maize/pea intercrop. Field Crops Res. 2012, 138, 11–20. [Google Scholar] [CrossRef]
- Zhang, Y.; Duan, Y.; Nie, J.; Yang, J.; Ren, J.; van der Werf, W.; Evers, J.B.; Zhang, J.; Su, Z.; Zhang, L. A lack of complementarity for water acquisition limits yield advantage of oats/vetch intercropping in a semi-arid condition. Agric. Water Manag. 2019, 225, 105778. [Google Scholar] [CrossRef]
- Wise, R.R.; Olson, A.J.; Schrader, S.M.; Sharkey, T.D. Electron transport is the functional limitation of photosynthesis in field-grown Pima cotton plants at high temperature. Plant Cell Environ. 2004, 27, 717–724. [Google Scholar] [CrossRef]
- Yamori, W.; Noguchi, K.; Kashino, Y.; Terashima, I. The role of electron transport in determining the temperature dependence of the photosynthetic rate in spinach leaves grown at contrasting temperatures. Plant Cell Physiol. 2008, 49, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Percival, G.C. The use of chlorophyll fluorescence to identify chemical and environmental stress in leaf tissue of three oak (Quercus) species. J. Arbor. 2005, 31, 215–227. [Google Scholar] [CrossRef]
- Guo, Y.P.; Zhou, H.F.; Zhang, L.C. Photosynthetic characteristics and protective mechanisms against photooxidation during high temperature stress in two citrus species. Sci. Hort. 2006, 108, 260–267. [Google Scholar] [CrossRef]
- Costa, E.S.; Bressan-Smith, R.; Oliveira, J.G.; Campostrini, E. Chlorophyll a fluorescence analysis in response to excitation irradiance in bean plants (Phaseolus vulgaris L. and Vigna unguiculata L. Walp) submitted to high temperature stress. Photosynthetica 2003, 41, 77–82. [Google Scholar] [CrossRef]
- Kitao, M.; Yasuda, Y.; Kodani, E.; Harayama, H.; Awaya, Y.; Komatsu, M.; Yazaki, K.; Tobita, H.; Agathokleous, E. Integration of electron flow partitioning improves estimation of photosynthetic rate under various environmental conditions based on chlorophyll fluorescence. Remote Sens. Environ. 2021, 254, 112273. [Google Scholar] [CrossRef]
- Song, X.; Zhou, G.; He, Q.; Zhou, H. Stomatal limitations to photosynthesis and their critical Water conditions in different growth stages of maize under water stress. Agric. Water Manag. 2020, 241, 106330. [Google Scholar] [CrossRef]
- Bheemanahalli, R.; Vennam, R.R.; Ramamoorthy, P.; Reddy, K.R. Effects of post-flowering heat and drought stresses on physiology, yield, and quality in maize (Zea mays L.). Plant Stress 2022, 6, 100106. [Google Scholar] [CrossRef]
- Müllers, Y.; Postma, J.A.; Poorter, H.; van Dusschoten, D. Stomatal conductance tracks soil-to-leaf hydraulic conductance in faba bean and maize during soil drying. Plant Physiol. 2022, 190, 2279–2294. [Google Scholar] [CrossRef]
- Pierre, J.F.; Latournerie-Moreno, L.; Garruña, R.; Jacobsen, K.L.; Laboski, C.A.M.; Us-Santamaría, R.; Ruiz-Sánchez, E. Effect of Maize–Legume Intercropping on Maize Physio-Agronomic Parameters and Beneficial Insect Abundance. Sustainability 2022, 14, 12385. [Google Scholar] [CrossRef]
- Pierre, J.F.; Latournerie-Moreno, L.; Garruña-Hernández, R.; Jacobsen, K.L.; Guevara-Hernández, F.; Laboski, C.A.M.; RuizSánchez, E. Maize legume intercropping systems in southern Mexico: A review of benefits and challenges. Ciênc. Rural 2022, 52, 11. [Google Scholar] [CrossRef]
- Cárceles Rodríguez, B.; Durán Zuazo, V.H.; Soriano Rodríguez, M.; García-Tejero, I.F.; Gálvez Ruiz, B.; Repullo-Ruibérriz de Torres, M.A.; Ordóñez-Fernández, R.; Carbonell-Bojollo, R.M.; Cuadros Tavira, S. Chapter 13—Legumes protect the soil erosion and ecosystem services. In Advances in Legumes for Sustainable Intensification; Meena, R.S., Kumar, S., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 247–266. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Soil Reaction (pH) | Plant Available Nutrient Content (mg/kg) | ||||
|---|---|---|---|---|---|---|
| Nan-100% | K | Mg | P | Ca | ||
| Arable Soil | 7.1 ± 0.2 | 23.9 ± 5.3 | 191 ± 34.2 | 153 ± 5.1 | 58.3 ± 5.1 | 5968 ± 556 |
| Abbrev. | Formula/Equations | Explanation |
|---|---|---|
| FV/FM | Maximal quantum yield of PSII fluorescence | |
| VJ | VJ = (FJ − F0)/(FM − F0) | Relative variable fluorescence at the J-step |
| PIABS | PIABS = (RC/ABS)·[φP0/(1 − φP0)]·[ψ0/(1 − ψ0)] | Performance index (potential) for energy conservation from exciton to the reduction of intersystem electron acceptors |
| ET0/RC | ET0/RC = M0·(1/VJ)·ψ0 | Electron transport flux (further than QA) per RC |
| DI0/RC | DI0/RC = (ABS/RC)–(TR0/RC) | The flux of dissipated excitation energy at time 0 |
| QY_L1 | Equivalent to effective quantum yield of PSII (ΦPSII) |
| Dependent Parameter | Climatic Parameter (Spearman Correlation, p < 0.05) | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| FV/FM | −0.341 | −0.050 | −0.111 | −0.457 | −0.078 | −0.332 | −0.025 | −0.553 |
| PIABS | −0.351 | −0.101 | −0.071 | −0.537 | −0.005 | −0.285 | 0.031 | −0.599 |
| ET0/RC | −0.152 | −0.276 | 0.166 | −0.299 | 0.268 | 0.083 | 0.413 | −0.110 |
| DI0/RC | 0.379 | 0.057 | 0.099 | 0.513 | 0.088 | 0.333 | 0.066 | 0.643 |
| 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | |
| FV/FM | −0.169 | 0.119 | −0.187 | −0.215 | 0.250 | −0.544 | −0.162 | 0.049 |
| PIABS | −0.253 | 0.115 | −0.296 | −0.271 | 0.149 | −0.493 | −0.247 | 0.103 |
| ET0/RC | −0.325 | 0.263 | −0.201 | −0.341 | 0.051 | 0.088 | −0.227 | 0.375 |
| DI0/RC | 0.180 | −0.086 | 0.268 | 0.211 | −0.206 | 0.592 | 0.215 | −0.019 |
| System | Z. mays [kg (DM) ha−1] | M. albus (MM) Lupinus sp. (ML) [kg (DM) ha−1] | Overall Productivity [kg (DM) ha−1] |
|---|---|---|---|
| Sole maize (SM) | 15 676 ± 508 | 15 676 ± 508 | |
| Intercropping (MM) | 14 514 ± 874 | 1 381 ± 102 | 15 895 ± 521 |
| Intercropping (ML) | 14 806 ± 1356 | 1 045 ± 221 | 15 851 ± 699 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lang, J.; Váczi, P.; Barták, M.; Hájek, J.; Kintl, A.; Zikmundová, B.; Elbl, J. Stimulative Effects of Lupinus sp. and Melilotus albus Underseed on the Photosynthetic Performance of Maize (Zea mays) in Two Intercropping Systems. Agronomy 2023, 13, 163. https://doi.org/10.3390/agronomy13010163
Lang J, Váczi P, Barták M, Hájek J, Kintl A, Zikmundová B, Elbl J. Stimulative Effects of Lupinus sp. and Melilotus albus Underseed on the Photosynthetic Performance of Maize (Zea mays) in Two Intercropping Systems. Agronomy. 2023; 13(1):163. https://doi.org/10.3390/agronomy13010163
Chicago/Turabian StyleLang, Jaroslav, Peter Váczi, Miloš Barták, Josef Hájek, Antonín Kintl, Barbora Zikmundová, and Jakub Elbl. 2023. "Stimulative Effects of Lupinus sp. and Melilotus albus Underseed on the Photosynthetic Performance of Maize (Zea mays) in Two Intercropping Systems" Agronomy 13, no. 1: 163. https://doi.org/10.3390/agronomy13010163
APA StyleLang, J., Váczi, P., Barták, M., Hájek, J., Kintl, A., Zikmundová, B., & Elbl, J. (2023). Stimulative Effects of Lupinus sp. and Melilotus albus Underseed on the Photosynthetic Performance of Maize (Zea mays) in Two Intercropping Systems. Agronomy, 13(1), 163. https://doi.org/10.3390/agronomy13010163