
Yield, Flower Quality, and Photo-Physiological Responses of Cut Rose Flowers Grafted onto Three Different Rootstocks in Summer Season


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Scanning Electron Microscopy (SEM)
2.3. Chlorophyll-a Fluorescence Fast Rise Curve (OJIP) and Photosynthesis
2.4. Root Activity
2.5. Experimental Design and Statistical Analysis
3. Results
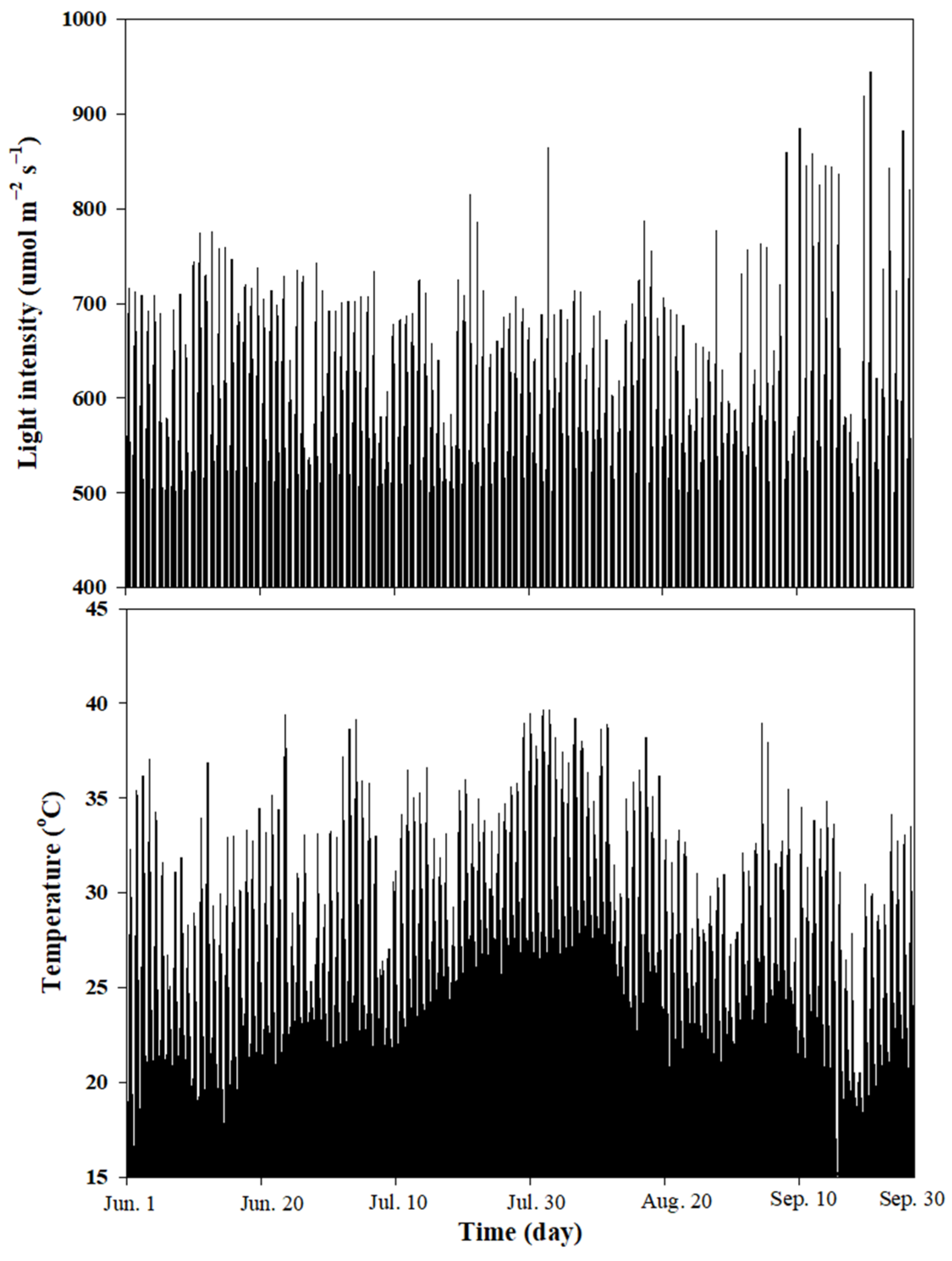
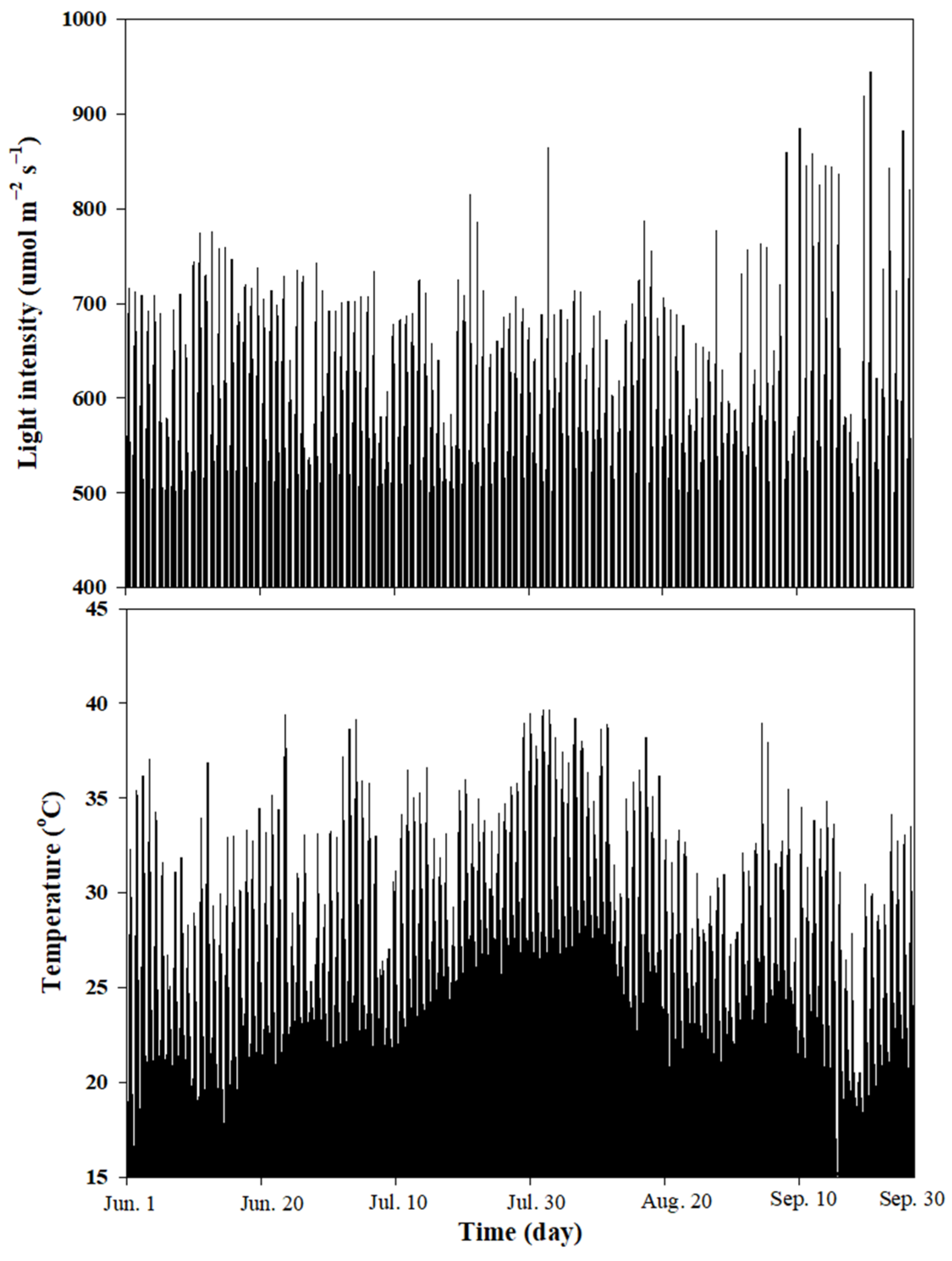
3.1. Ambient Light Intensity and Temperature in the Greenhouse
3.2. Stomata Images of Cut Rose Flowers
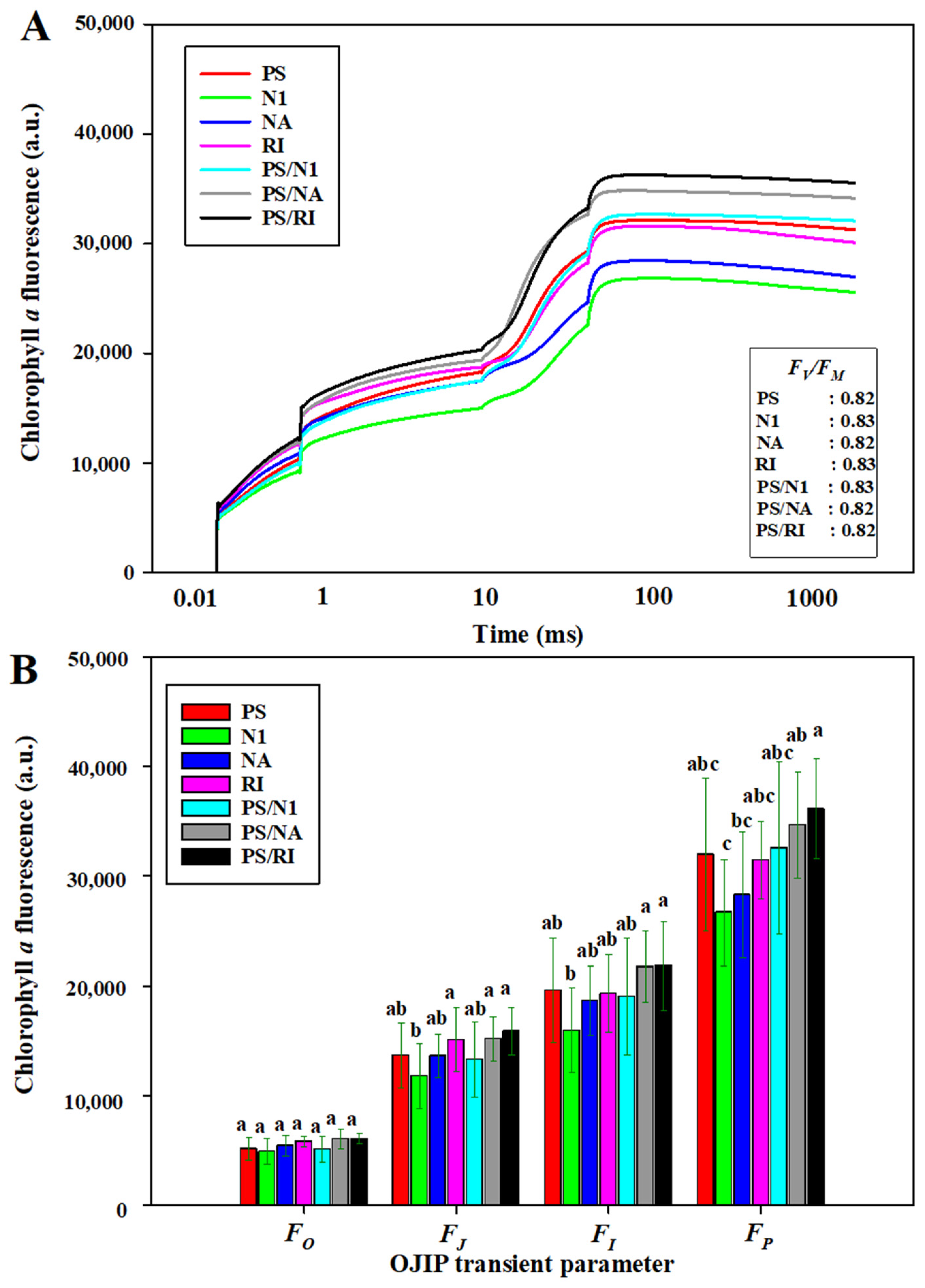
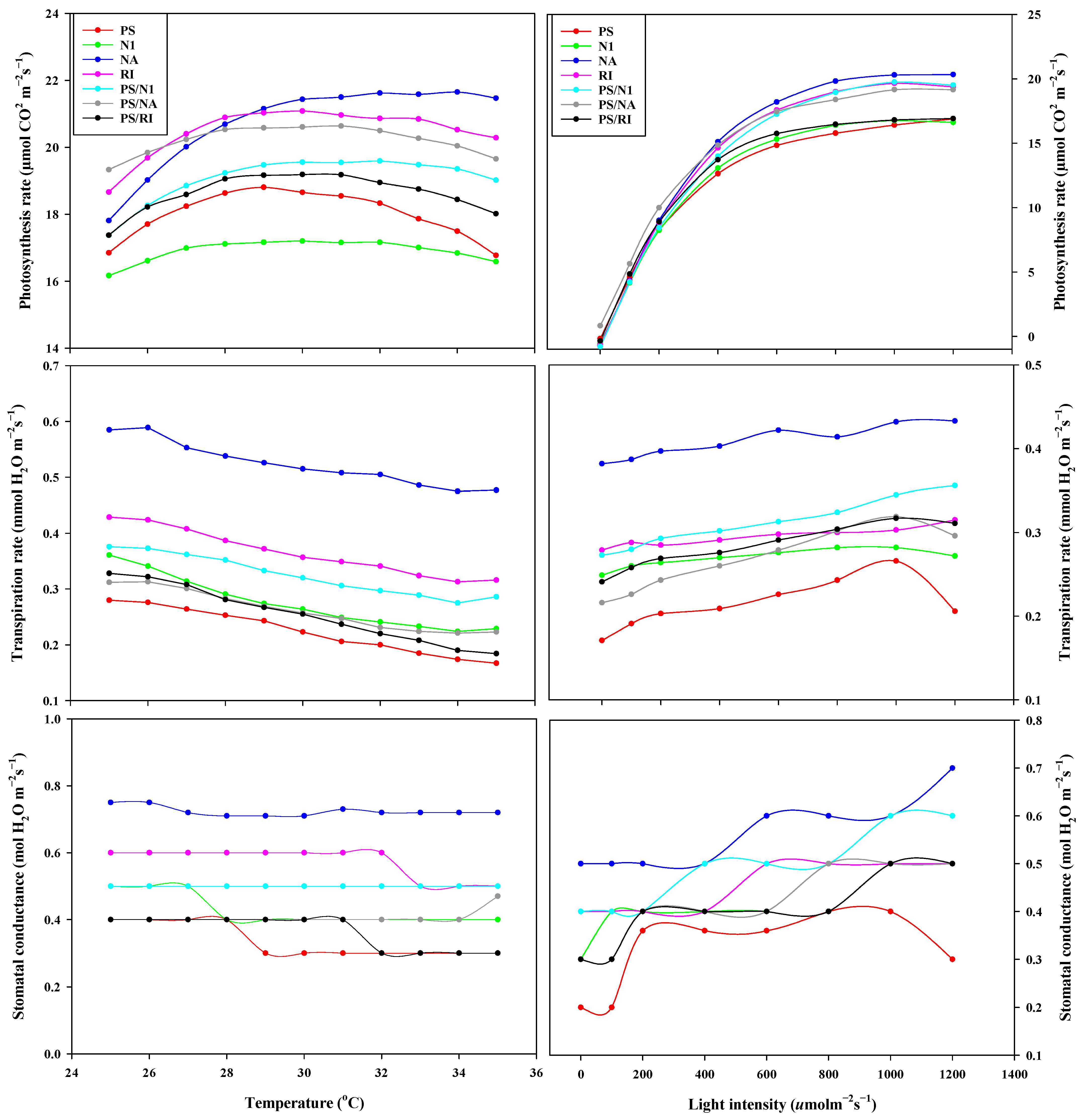
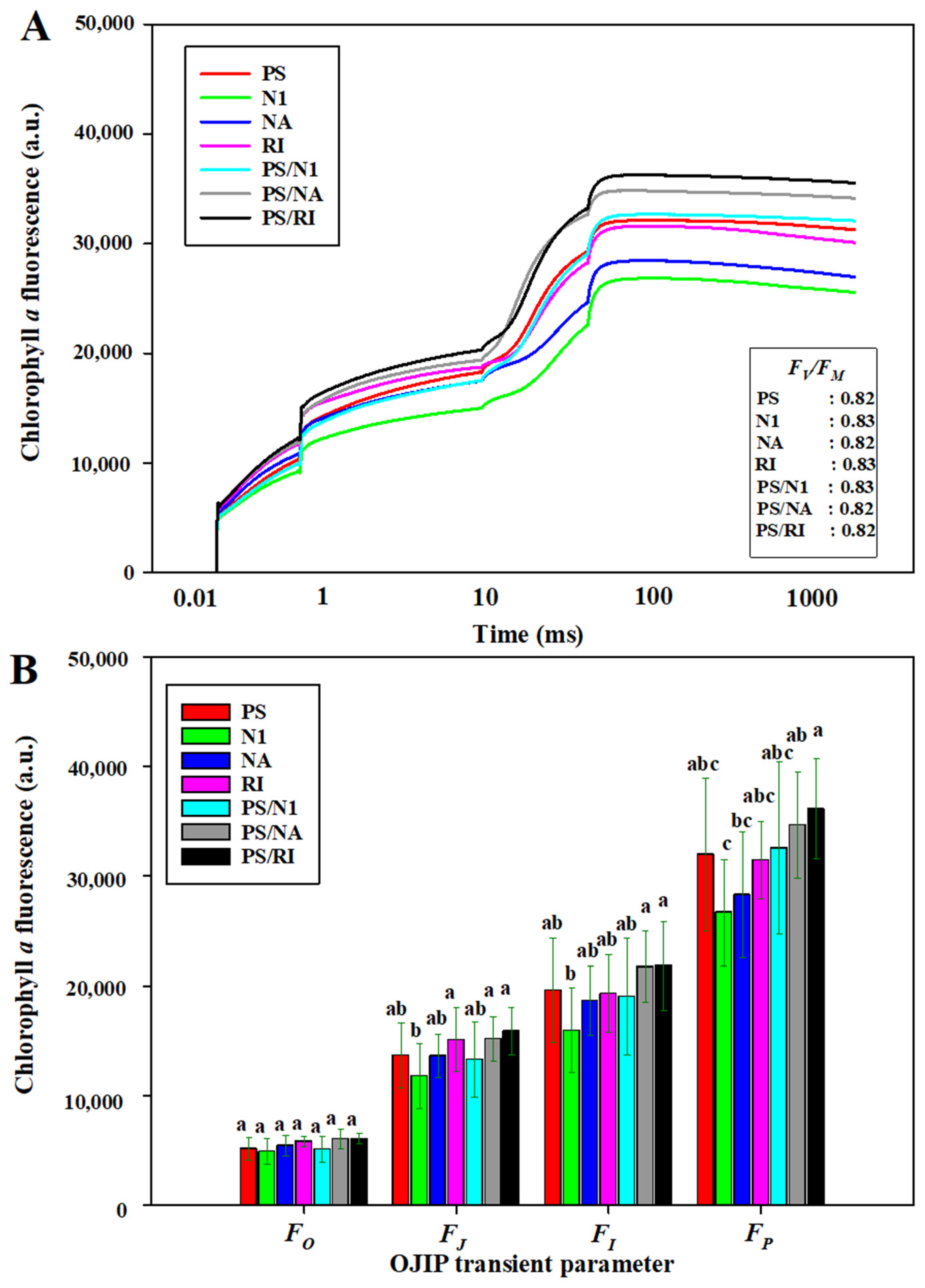
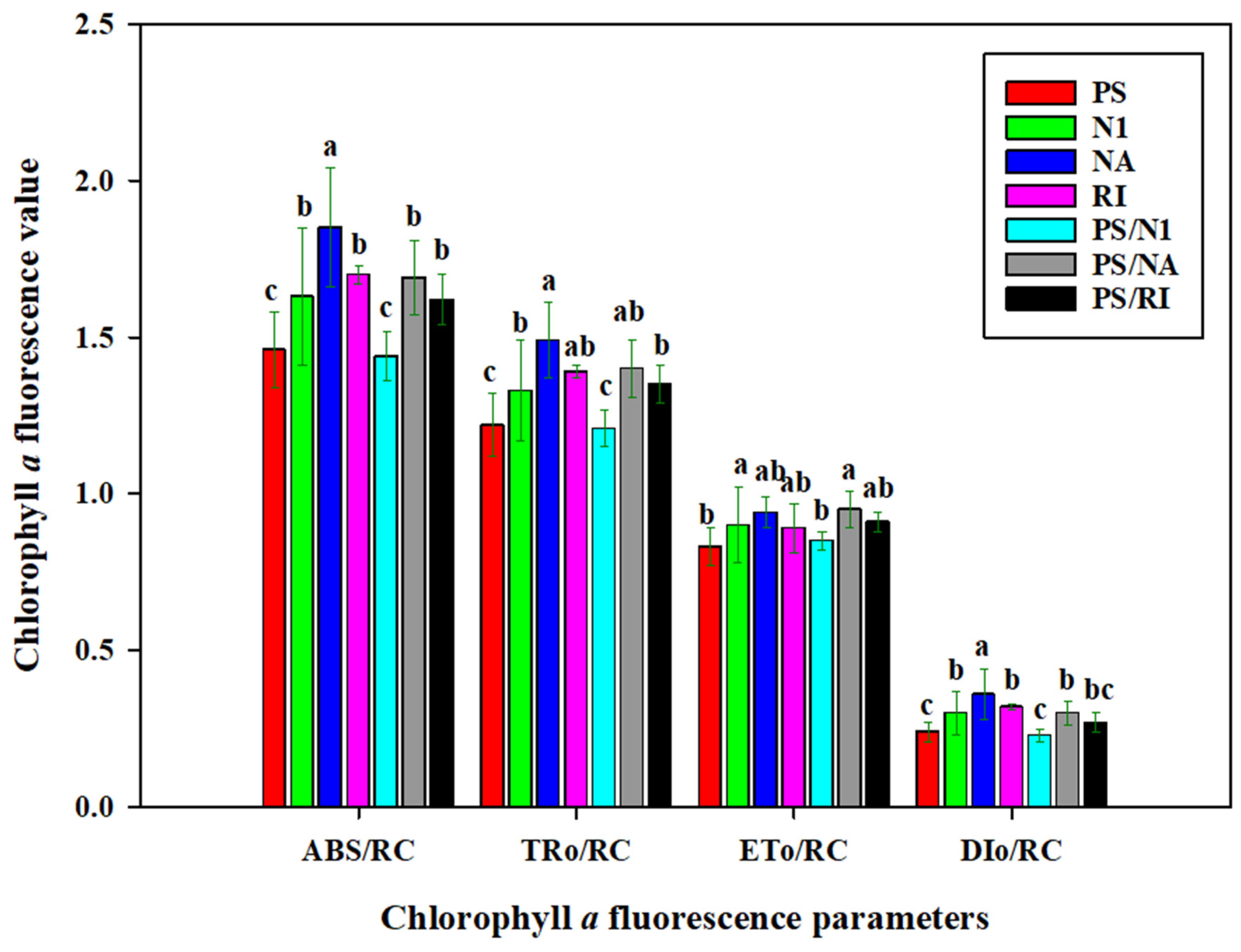
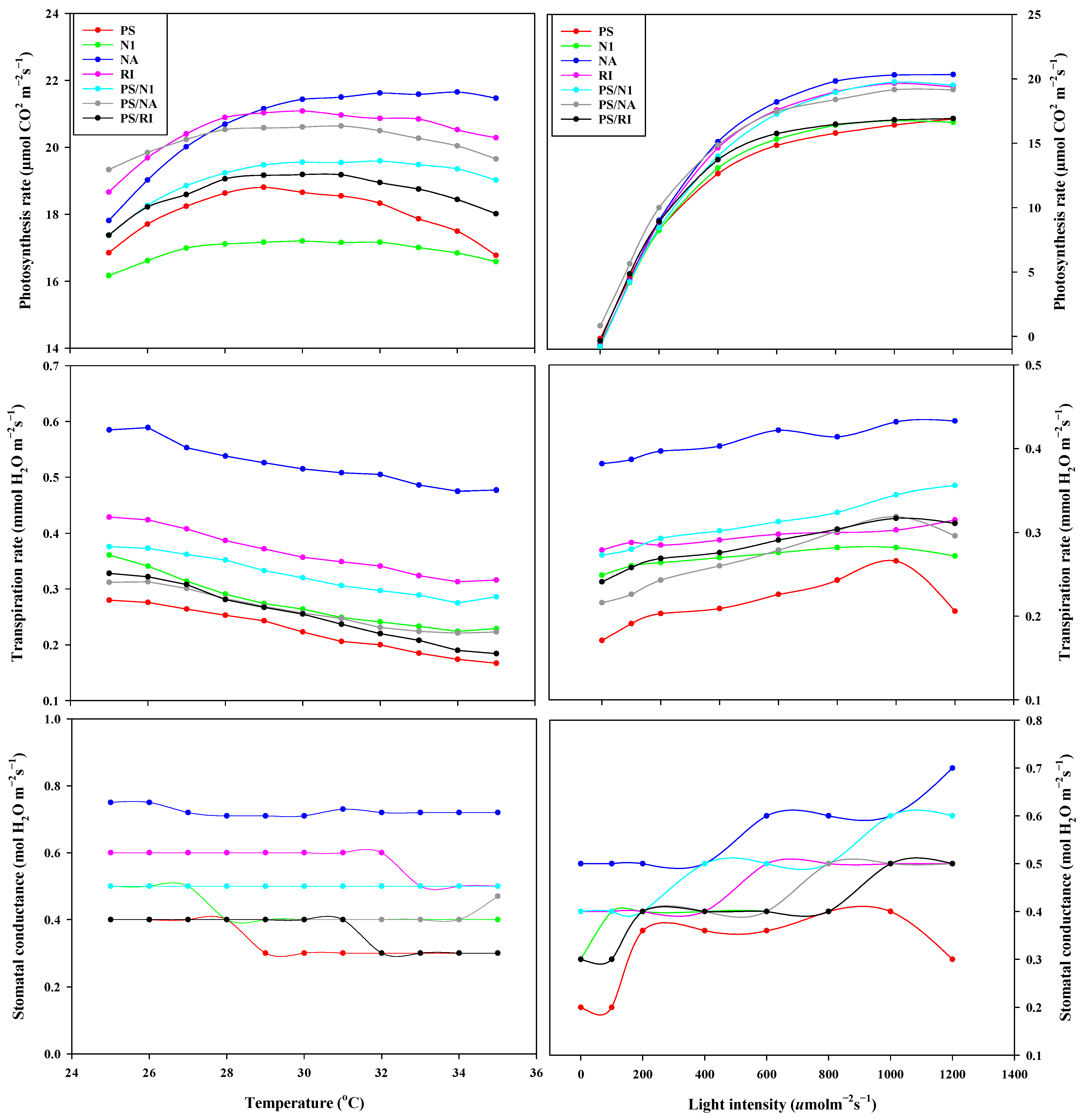
3.3. OJIP and Photosynthesis of Cut Rose Flowers
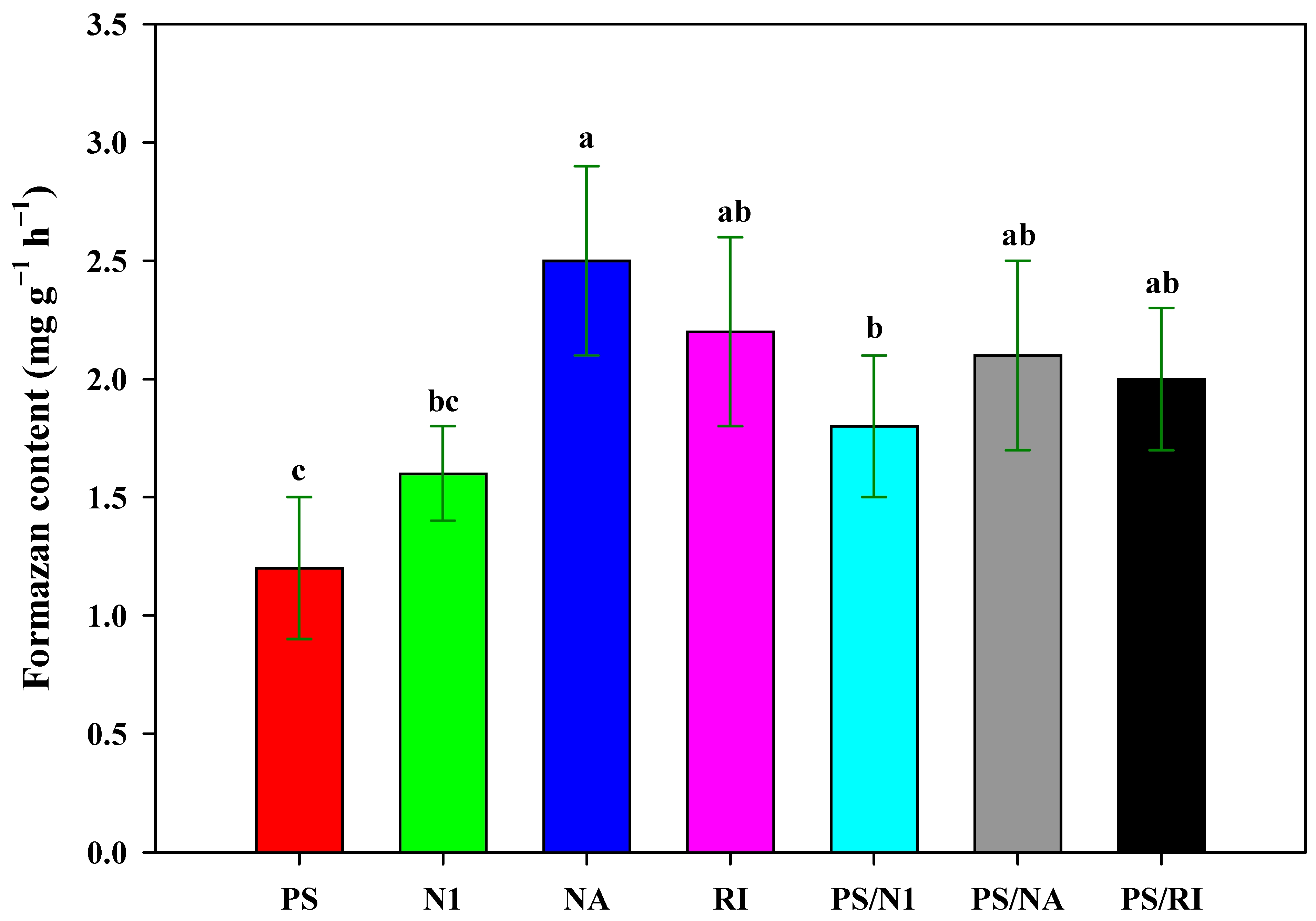
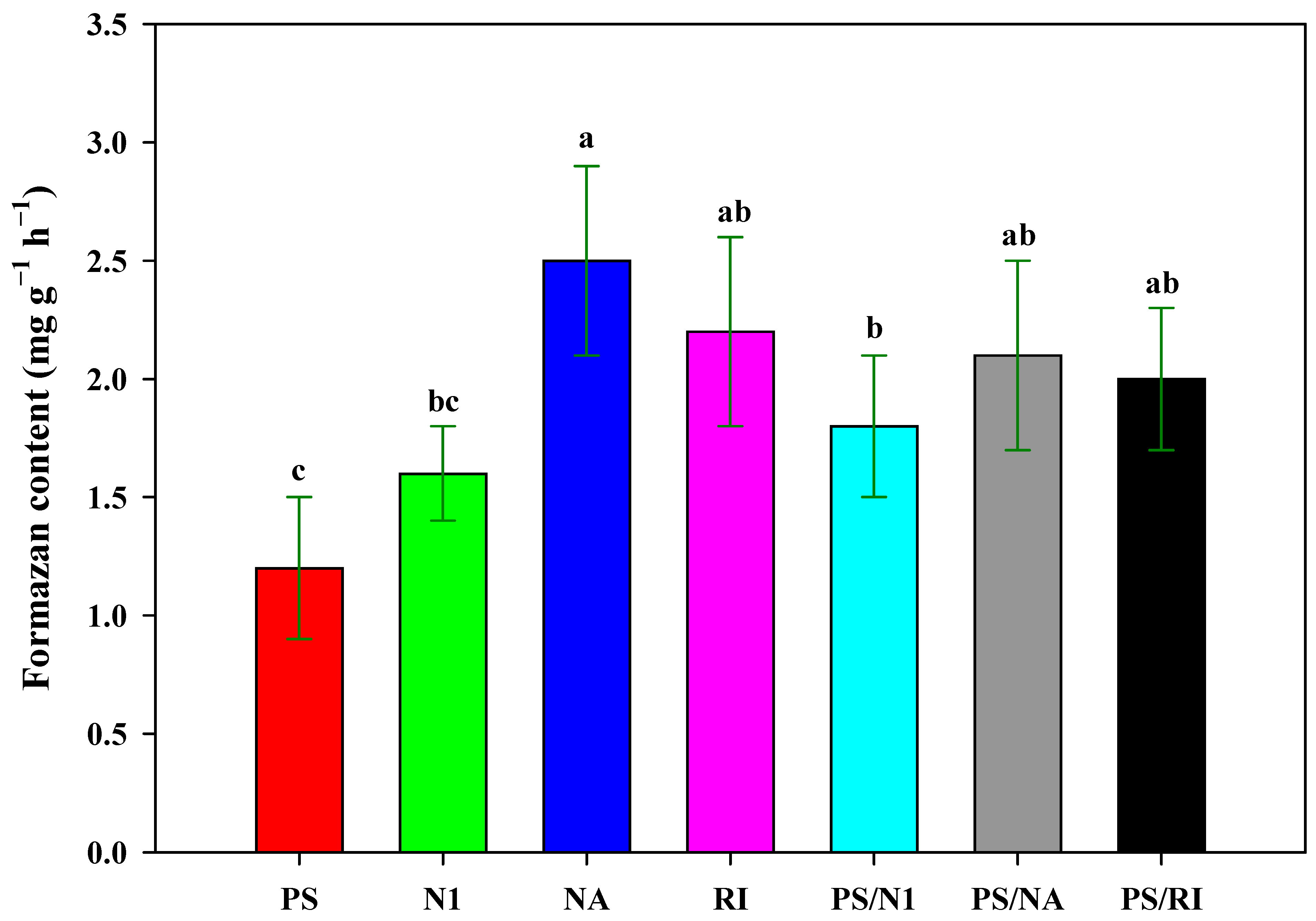
3.4. Root Activity of Cut Rose Flowers
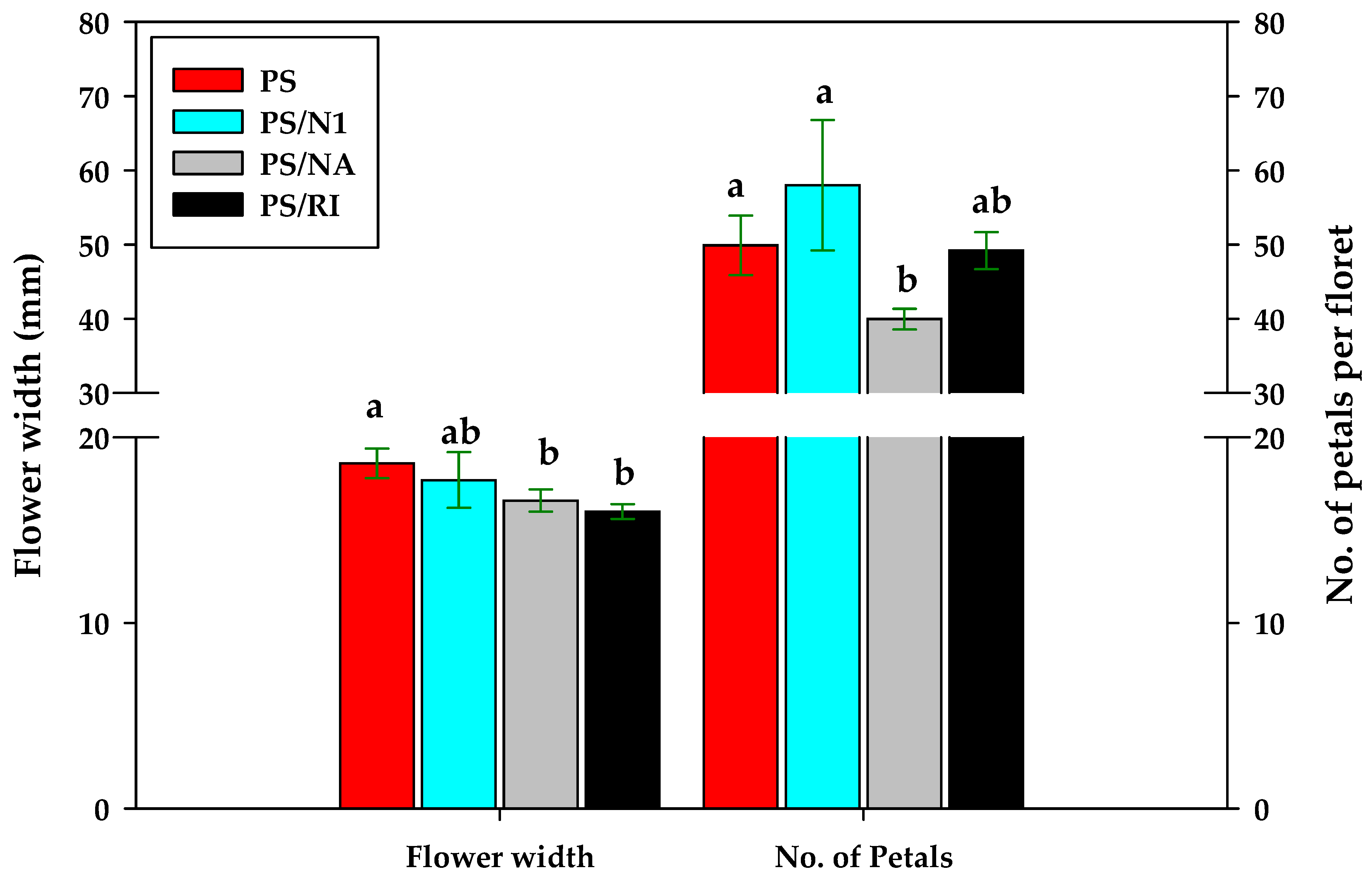
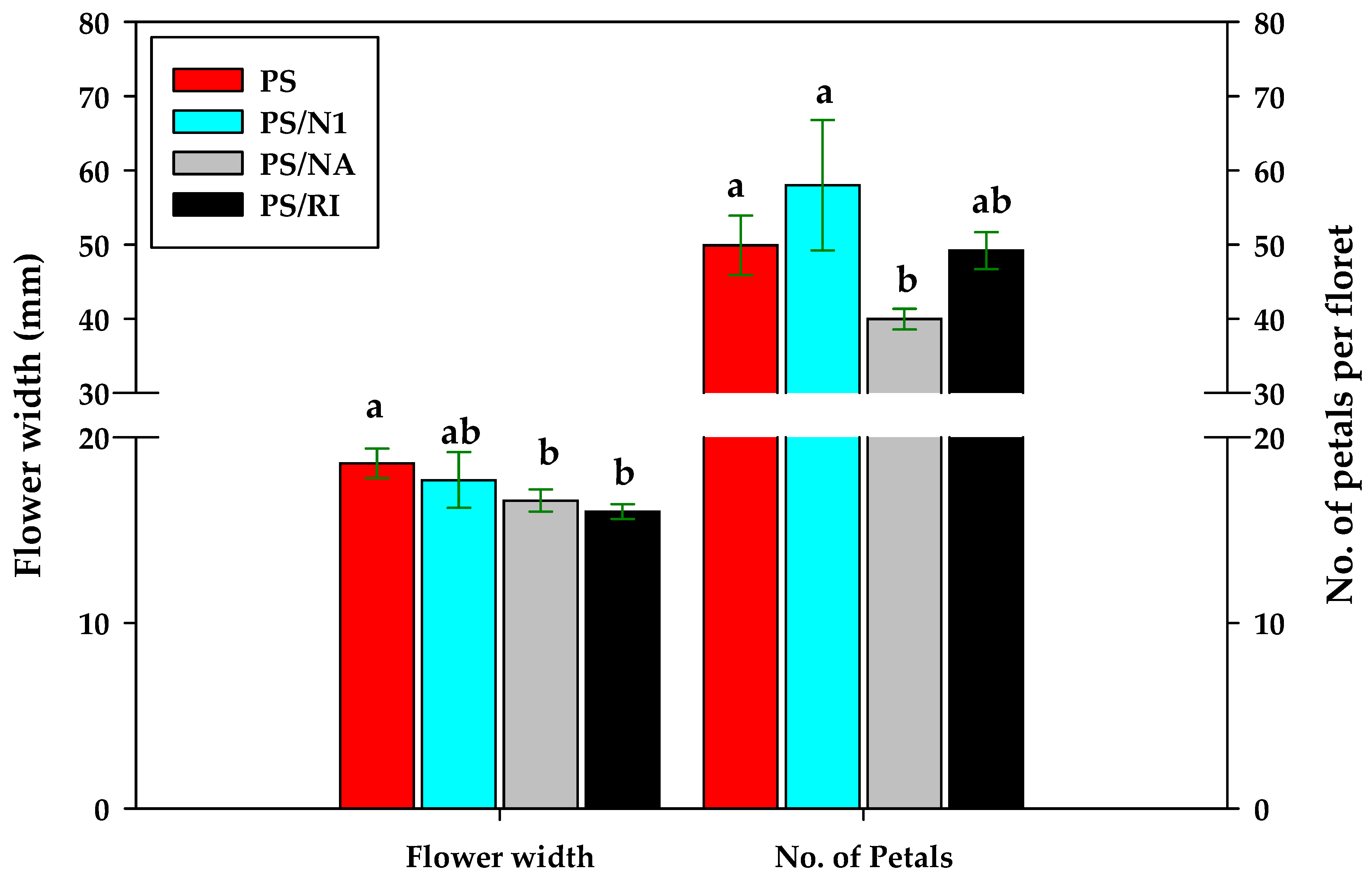
3.5. Yield and Flower Quality of Cut Rose Flowers
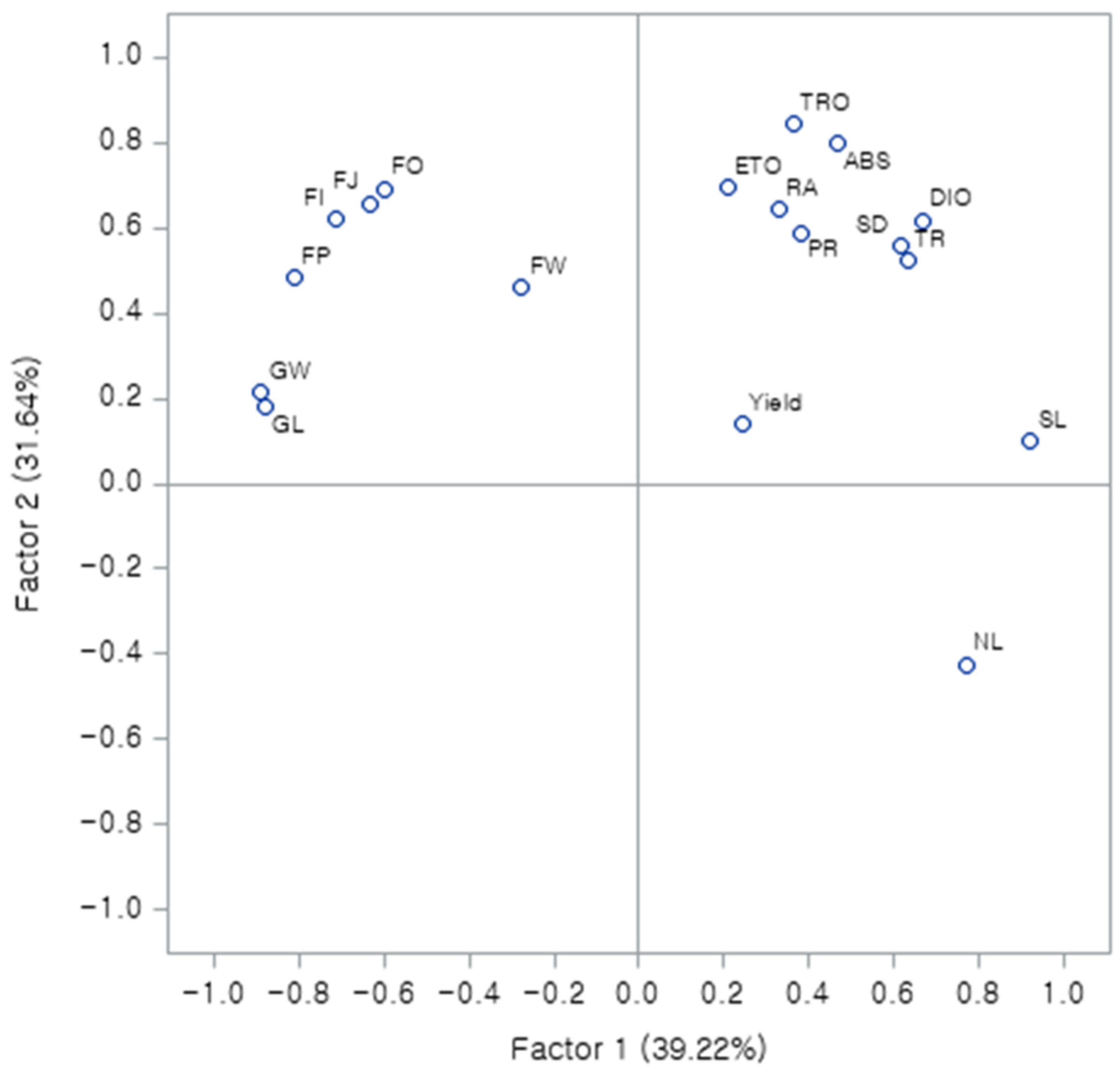
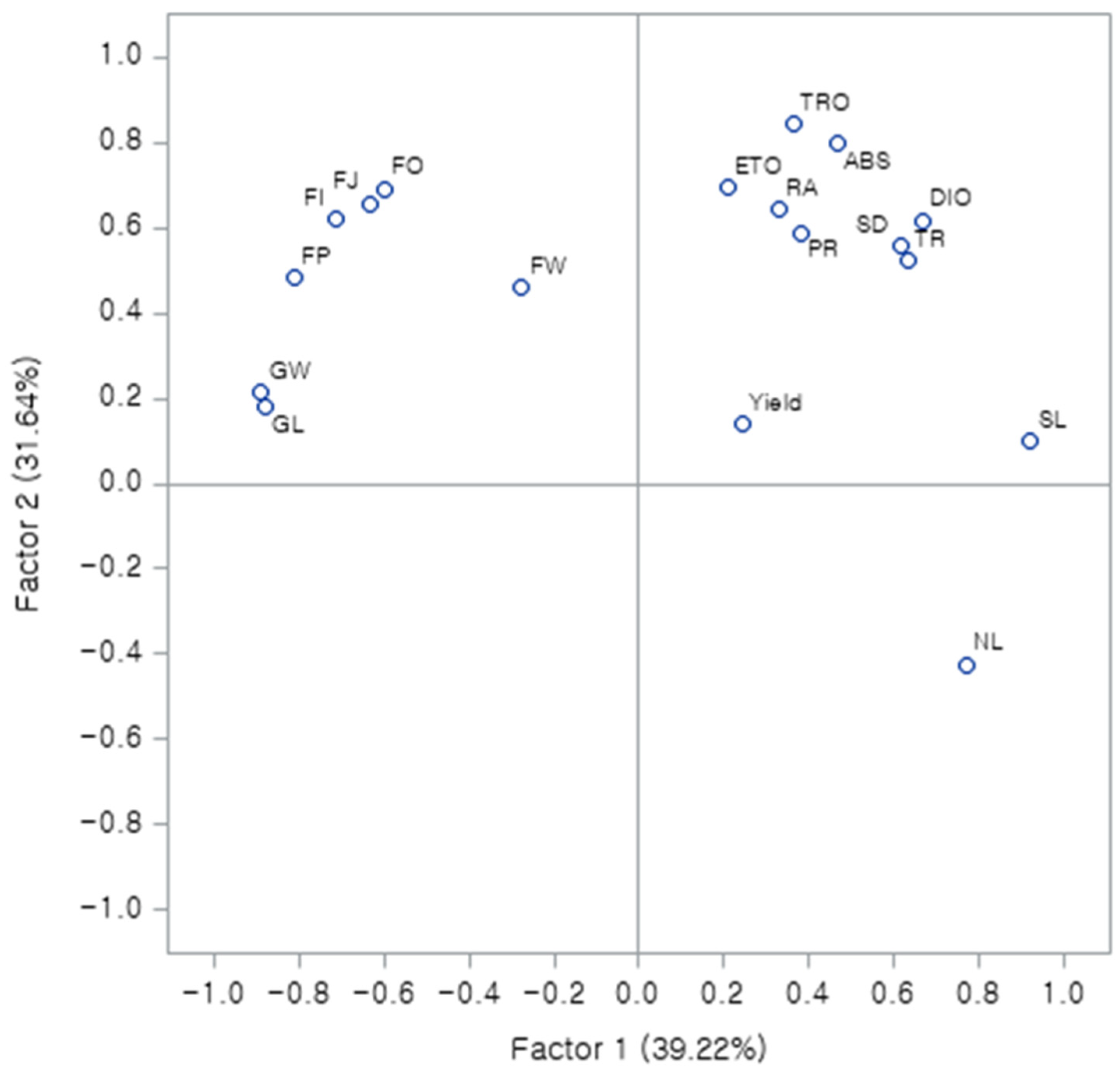
3.6. Principal Component Analysis (PCA)
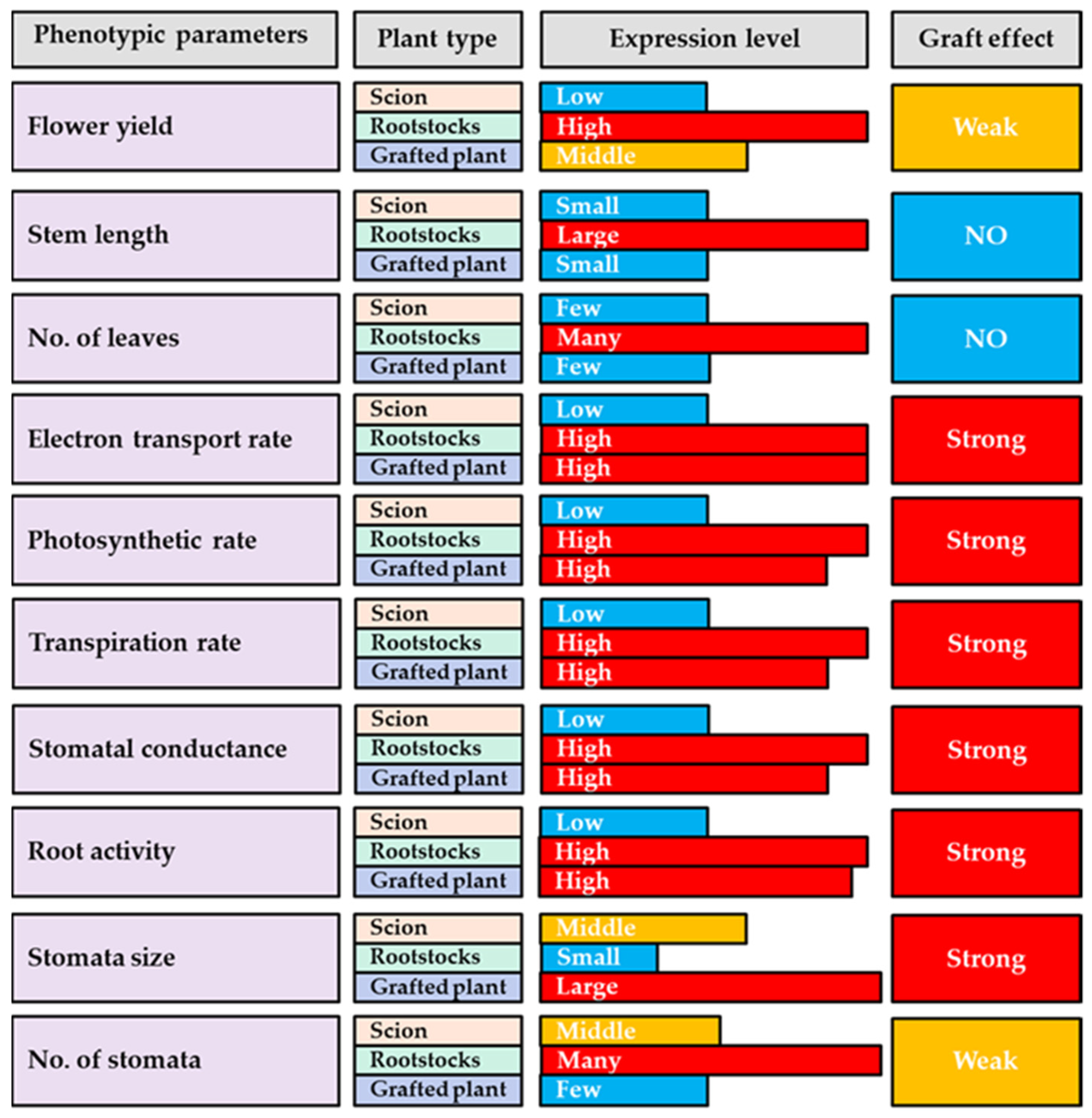
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferrante, A.; Mariani, L. Agronomic management for enhancing plant tolerance to abiotic stresses: High and low values of temperature, light intensity, and relative humditiy. Horticulturae 2018, 4, 21. [Google Scholar] [CrossRef] [Green Version]
- Toscano, S.; Trivellini, A.; Cocetta, G.; Bulgari, R.; Francini, A.; Romano, D.; Ferrante, A. Effect of preharvest abiotic stresses on the accumulation of bioactivie compounds in horticultural produce. Front. Plant Sci. 2019, 10, 1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, L.; Guo, L.; Li, R.; Cheng, Y.; Huang, L.; Zhou, H.; Xu, M.; Li, F.; Zhang, X.; Zheng, Y. Responses of photosynthesis to high temperature stress associated with changes in leaf structure and biochemistry of blueberry (Vaccinium corymbosum L.). Sci. Hortic. 2019, 246, 251–264. [Google Scholar] [CrossRef]
- Posch, B.C.; Kariyawasam, B.C.; Bramely, H.; Coast, O.; Richards, R.A.; Reynolds, M.P.; Trethowan, R.; Atkin, O.K. Exploring high temperature responses of photosynthesis and respiration to improve heat tolerance in wheat. J. Exp. Bot. 2019, 70, 5051–5069. [Google Scholar] [CrossRef]
- Qi, W.; Zhang, C.; Wang, W.; Cao, Z.; Li, S.; Li, H.; Zhu, W.; Huang, Y.; Bao, M.; He, Y.; et al. Comparative transcriptome analysis of different heat stress responses between self-root grafting line and heterogeneous grafting line in rose. Hortic. Plant J. 2021, 7, 243–255. [Google Scholar] [CrossRef]
- Jiang, C.; Bi, Y.; Zhang, R.; Feng, S. Expression of RcHSP70, heat shock protein 70 gene from Chinese rose, enhances host resistance to abiotic stresses. Sci. Rep. 2020, 10, 2445. [Google Scholar] [CrossRef]
- Choi, H.G. Correlation among phenotypic parameters related to the growth and photosynthesis of the growth (Fragaria × ananassa Duch.) grown under various light intensity conditions. Front. Plant Sci. 2021, 12, 647585. [Google Scholar] [CrossRef]
- Chen, S.; Yang, J.; Zhang, M.; Strasser, R.J.; Qiang, S. Classification and characteristics of heat tolerance in Ageratina adenophora populations using fast chlorophyll a fluorescence rise O-J-I-P. Environ. Exp. Bot. 2016, 122, 126–140. [Google Scholar] [CrossRef]
- Na, Y.W.; Jeong, H.J.; Lee, S.Y.; Choi, H.G.; Kim, S.H.; Rho, I.R. Chlorophyll fluorescence as a diagnostic tool for abiotic stress tolerance in wild and cultivated strawberry species. Hort. Environ. Biotechnol. 2014, 55, 208–286. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Choi, H.G.; Moon, B.Y.; Kang, N.J. Correlation between strawberry (Fragaria ananassa Duch.) productivity and photosynthesis-related parameters under various growth conditions. Front. Plant Sci. 2016, 7, 1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolla, A.; Voyiatzis, D.; Koukourikou-Petridou, M.; Chimonidou, D. Photosynthetic parameters and cut-flower yield of rose ‘Eurored’ (H.T.) are adversely affected by mild water stress irrespective of substrate composition. Sci. Hortic. 2010, 126, 390–394. [Google Scholar] [CrossRef]
- Nomura, K.; Saito, M.; Tada, I.; Iwao, T.; Yamazaki, T.; Kira, N.; Nishimura, Y.; Mori, M.; Baeza, E.; Kitano, M. Estimation of photosynthesis loss due to greenhouse superstructures and shade nets: A case study with paprika and tomato canopies. HortScience 2022, 57, 464–471. [Google Scholar] [CrossRef]
- Franco, J.A.; Bañón, S.; Vicente, M.J.; Miralles, J.; Martínez-Sánchez, J.J. Root development in horticultural plants grown under abiotic stress conditions—A review. J. Hortic. Sci. Biotechnol. 2011, 86, 543–556. [Google Scholar] [CrossRef]
- Li, C.; Wang, P.; Wei, Z.; Liang, D.; Liu, C.; Yin, L.; Jia, D.; Fu, M.; Ma, F. The mitigation effects of exogenous melatonin on salinity-inducedstress in Malus hupehensis. J. Pineal Res. 2012, 53, 298–306. [Google Scholar] [CrossRef]
- Choi, H.G.; Jeong, H.J. Comparison of chlorophyll fluorescence and photosynthesis of two strawberry cultivars in response to relative humidity. Hortic. Sci. Technol. 2020, 38, 66–77. [Google Scholar]
- Sotelo-Cuitiva, Y.M.; Restrepo-Díaz, H.; García-Castro, A.; Ramírez-Godoy, A.; Flórez-Roncancio, V.J. Effect of kaolin film particle applications (Surround WP®) and water deficit on physiological characteristics in rose cut plants (Rose spp L.). Am. J. Plant Sci. 2011, 2, 354–358. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Yoon, J.W.; Oh, S.I.; Lee, A.K. Relationship between cultivation environment and postharvest quality of cut rose ‘Lovely lydia’. Hortic. Sci. Technol. 2020, 38, 263–270. [Google Scholar]
- Izadi, Z.; Zarei, H.; Alizadeh, M. Role of grafting technique on the success of stenting propagation of two rose (Rosa sp.) varieties. Am. J. Plant Sci. 2013, 4, 41–44. [Google Scholar] [CrossRef] [Green Version]
- Auler, P.A.; Freire, F.B.S.; Lima, V.F.; Daloso, D.M. On the role of guard cells in sensing environmental signals and memorizing stress periods. Theor. Exp. Plant Physiol. 2022, 34, 1–23. [Google Scholar] [CrossRef]
- Melis, A.; Zeiger, E. Chlorophyll a fluorescence transients in mesophyll and guard cells. Plant Physiol. 1982, 69, 642–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohkawa, K.; Suematsu, M. Arching cultivation techniques for growing cut-roses. Acta Hort. 1999, 482, 47–51. [Google Scholar] [CrossRef]
- Clément, C.; Burrus, M.; Audran, J.C. Floral organ growth and carbohydrate content during pollen development in Lilium. Am. J. Bot. 1996, 83, 459–469. [Google Scholar] [CrossRef]
- Stirbet, A.; Govindjee. On the relation between the kautsky effect (chlorophyll a fluorescence induction) and Photosystem II: Basics and applications of the OJIP fluorescence transient. J. Photochem. Photobiol. B Biol. 2011, 104, 236–257. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.B.; Lee, I.B.; Park, J.M.; Lim, T.J. Effect of waterlogging conditions on the growth, root activities and nutrient content of ‘Campbell Early’ grapevine. Korean J. Hort. Sci. Technol. 2010, 28, 172–179. [Google Scholar]
- Zhang, X.; Feng, Y.; Khan, A.; Ullah, N.; Li, Z.; Zaheer, S.; Zhou, R.; Zhang, Z. Quantitative proteomics-based analysis reveals molecular mechanisms of chilling tolerance in grafted cotton seedlings. Agronomy 2022, 12, 1152. [Google Scholar] [CrossRef]
- Reig, G.; Iglesias, I.; Zazurca, L.; Torguet, L.; Martinez, G.; Miarnau, X. Physiological and agronomical responses of ‘Vairo’ almond and ‘Big Top’ nectarine cultivars grafted onto different prunus rootstocks and grown under semiarid Mediterranean conditions. Agronomy 2022, 12, 821. [Google Scholar] [CrossRef]
- Lin, S.; Chen, P.A.; Zhuang, B.W. The stomatal conductance and Fv/Fm as the indicators of stress tolerance of avocado seedling under short-term waterlogging. Agronomy 2022, 12, 1084. [Google Scholar] [CrossRef]
- Biswal, A.K.; Kohli, A. Cereal flag leaf adaptations for grain yield under drought: Knowledge status and gaps. Mol. Breed. 2013, 31, 749–766. [Google Scholar] [CrossRef]
- Hetherington, A.M.; Woodward, F.I. The role of stomata in sensing and driving environmental change. Nature 2003, 424, 901–908. [Google Scholar] [CrossRef]
- Shahinnia, F.; Roy, J.L.; Laborde, B.; Sznajder, B.; Kalambettu, P.; Mahjourimaid, S.; Tilbrook, J.; Fleury, D. Genetic association of stomatal traits and yield in wheat grown in low rainfall environments. BMC Plant Biol. 2016, 16, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, D.C.; Sack, F.D. Stomatal development. Annu. Rev. Plant Biol. 2007, 58, 163–181. [Google Scholar] [CrossRef] [PubMed]
- Drake, P.L.; Froend, R.H.; Franks, P.J. Smaller, faster stomata: Scaling of stomatal size, rate of response, and stomatal conductance. J. Exp. Bot. 2013, 64, 495–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadras, V.O.; Montoro, A.; Moran, M.A.; Aphalo, P.J. Elevated temperature altered the reaction norms of stomatal conductance in field-grown grapevine. Agric. For. Meteorol. 2012, 165, 35–42. [Google Scholar] [CrossRef]
- Yarsi, G.; Sivaci, A.; Dasgan, H.Y.; Altuntas, O.; Binzet, R.; Akhoundnejad, Y. Effects of salinity stress on chlorophyll and carotenoid contents and stomata size of grafted and ungrafted Galia C8 melon cultivar. Pak. J. Bot. 2017, 49, 421–426. [Google Scholar]
- Oustric, J.; Herbette, S.; Quilichini, Y.; Morillon, R.; Giannettini, J.; Berti, L.; Santini, J. Tetraploid Citrumelo 4475 rootstocks improve diploid common clementine tolerance to long-term nutrient deficiency. Sci. Rep. 2021, 11, 8902. [Google Scholar] [CrossRef]
- Liu, S.; Li, H.; Lv, X.; Ahammed, G.J.; Xia, X.; Zhou, J.; Shi, K.; Asami, T.; Yu, J.; Zhou, Y. Grafting cucumber onto luffa improves drought tolerance by increasing ABA biosynthesis and sensitivity. Sci. Rep. 2016, 6, 20212. [Google Scholar] [CrossRef] [Green Version]
- Yan, K.; Chen, P.; Shao, H.; Shao, C.; Zhao, S.; Brestic, M. Dissection of photosynthetic electron transport process in sweet sorghum under heat stress. PLoS ONE 2013, 8, e62100. [Google Scholar] [CrossRef] [Green Version]
- Zushi, K.; Kajiwara, S.; Matsuzoe, N. Chlorophyll a fluorescence OJIP transient as a tool to characterize and evaluate response to heat and chilling stress in tomato leaf and fruit. Sci. Hortic. 2012, 148, 39–46. [Google Scholar] [CrossRef]
- Poudyal, D.; Rosenqvist, E.; Ottosen, C.O. Phenotyping from lab to field—Tomato lines screened for heat stress using Fv/Fm maintain high fruit yield during thermal stress in the field. Funct. Plant Biol. 2018, 46, 44–55. [Google Scholar] [CrossRef]
- Gisbert-Mullor, R.; Padilla, Y.G.; Martínez-Cuenca, M.R.; López-Galarza, S.; Calatayud, Á. Suitable rootstocks can alleviate the effects of heat stress on pepper plants. Sci. Hortic. 2021, 290, 110529. [Google Scholar] [CrossRef]
- Fan, Y.; Li, Z.; Xie, B.; Liang, X.; Huang, X. A study of shoot growth, leaf photosynthesis, and nutrients in ‘Lingfengjing’ litchi grafted onto seedlings of different cultivars. Horticulturae 2022, 8, 282. [Google Scholar] [CrossRef]
- Bai, Y.; Zhang, J.; Wu, Y.; Huang, R.; Chang, Y.; Lei, X.; Song, X.; Pei, D. Possibility of increasing the growth and photosynthetic properties of precocious walnut by grafting. Sustainability 2020, 12, 5178. [Google Scholar] [CrossRef]
- Geboloğlu, N.; Yilmaz, E.; Çakmak, P.; Aydin, M.; Kasap, Y. Determining of the yield, quality and nutrient content of tomatoes grafted on different rootstocks in soilless culture. Sci. Res. Essays 2011, 6, 2147–2153. [Google Scholar]
- Zhang, X.; Khan, A.; Zhou, R.; Liu, Y.; Zhang, B.; Wang, Q.; Zhang, Z. Grafting in cotton: A mechanistic approach for stress tolerance and sustainable development. Ind. Crop. Prod. 2022, 175, 114227. [Google Scholar] [CrossRef]
- Rouphael, Y.; Cardarelli, M.; Colla, G.; Rea, E. Yield, mineral composition, water relations, and water use efficiency of grafted mini-watermelon plants under deficit irrigation. HortScience 2008, 43, 730–736. [Google Scholar] [CrossRef] [Green Version]
- Fang, S.; Tao, Y.; Zhang, Y.; Kong, F.; Wang, Y. Effects of metalaxyl enantiomers stress on root activity and leaf antioxidant enzyme activities in tobacco seedlings. Chirality 2018, 30, 469–474. [Google Scholar] [CrossRef]
- Salehi-Mohammadi, R.; Kashi, A.; Lee, S.G.; Hun, Y.C.; Lee, J.M.; Babalar, M.; Delshad, M. Assessing the survival and growth performance of Iranian melon to grafting onto Cucurbita rootstocks. Korean J. Hort. Sci. Technol. 2009, 27, 1–6. [Google Scholar]
- Canales, F.J.; Rispail, N.; García-Tejera, O.; Arbona, V.; Pérez-de-Luque, A.; Prats, E. Drought resistance in oat involves ABA-mediated modulation of transpiration and root hydraulic conductivity. Environ. Exp. Bot. 2021, 182, 104333. [Google Scholar] [CrossRef]
- Yassin, H.; Hussen, S. Reivew on role of grafting on yield and quality of selected fruit vegetables. Glob. J. Sci. Front. Res. 2015, 15, 1–12. [Google Scholar]
- Kyriacou, M.C.; Rouphael, Y.; Colla, G.; Zrenner, R.; Schwarz, D. Vegetable grafting: The implications of a growing agronomic imperative for vegetable fruit quality and nutritive value. Front. Plant Sci. 2017, 8, 741. [Google Scholar] [CrossRef] [PubMed]
- Soteriou, G.A.; Kyriacou, M.C.; Siomos, A.S.; Gerasopoulos, D. Evolution of watermelon fruit physicochemical and phytochemical composition during ripening as affected by grafting. Food Chem. 2014, 165, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Świerczyński, S. The effect of rootstock activity for growth and root system soaking in trichoderma atroviride on the graft success and continued growth of beech (Fagus sylvatica L.) plants. Agronomy 2022, 12, 1259. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Definition |
|---|---|
| F0 | Minimal fluorescence when all photosystem II reaction centers are open (at 20 μs) |
| FJ | Fluorescence intensity at J step (at 3 ms) |
| FI | Fluorescence intensity at I step (at 30 ms) |
| FM (FP) | Maximal fluorescence intensity when all photosystem II reaction centers are closed |
| FV | Maximal variable fluorescence: FV = FM − F0 |
| FV/FM (ϕP0) | Maximal quantum yield of primary photochemistry. Expresses the probability that an absorbed photon leads to a reduction in QA: ϕP0 = TR0/ABS = FV/FM |
| MO | Initial slope of induction curve: MO = 4 (F300μs − F0)/(FM − F0) = TR0/RC − ET0/RC |
| ABS/RC | Average absorbed photon flux per photosystem II reaction center:ABS/RC = MO (1/VJ) (1/ϕP0) |
| TR0/RC | Trapped energy flux leading to a reduction in QA: TR0/RC = MO (1/VJ) |
| ET0/RC | Rate of electron transport flux from QA to QB per photosystem II reaction center:ET0/RC = MO (1/VJ) (1 − VJ) |
| DI0/RC | Dissipated energy flux per reaction center: DI0/RC = ABS/RC − TR0/RC |
| Treatment | Size Characteristics (µm) | |||
|---|---|---|---|---|
| Guard Cell Length | Guard Cell Width | Stoma (Pore) Length | Stoma (Pore) Width | |
| PS | 28.18 ± 2.26 b z | 11.02 ± 1.12 b | 17.95 ± 1.35bc | 2.49 ± 0.17 b |
| N1 | 18.85 ± 1.37cd | 7.42 ± 1.03cd | 14.31 ± 1.16 d | 2.38 ± 0.11 b |
| NA | 17.53 ± 1.56 d | 6.69 ± 1.23 d | 14.16 ± 1.07 d | 2.15 ± 0.23 b |
| RI | 20.98 ± 1.48 c | 8.75 ± 1.08 c | 16.51 ± 1.26 c | 2.46 ± 0.13 b |
| PS/N1 | 33.46 ± 1.96ab | 12.24 ± 1.38 b | 20.29 ± 1.42ab | 3.12 ± 0.33 a |
| PS/NA | 30.15 ± 1.56 b | 12.01 ± 1.06 b | 19.18 ± 1.21 b | 2.97 ± 0.25 a |
| PS/RI | 36.52 ± 1.74 a | 14.37 ± 1.36 a | 21.18 ± 1.35 a | 3.62 ± 0.15 a |
| Treatment | Stem Length (cm) | No. of Leaves (/Stem) | Fresh Weight (g/Stem) | Yield (No. of Flowers/Plant) |
|---|---|---|---|---|
| PS | 52.0 ± 0.6cd z | 8.4 ± 0.2 b | 39.0 ± 2.6 a | 4.4 ± 0.5 c |
| N1 | 99.5 ± 13.3 b | 32.5 ± 4.9 a | 20.0 ± 12.1 a | 5.3 ± 0.6 b |
| NA | 125.6 ± 15.8 a | 34.7 ± 1.9 a | 40.6 ± 12.0 a | 4.8 ± 0.6 bc |
| RI | 122.7 ± 14.4 a | 33.6 ± 1.2 a | 33.8 ± 4.9 a | 7.0 ± 1.5 a |
| PS/N1 | 50.3 ± 0.8 d | 8.2 ± 0.3 b | 41.5 ± 4.1 a | 5.3 ± 0.3 b |
| PS/NA | 54.0 ± 1.9 c | 8.3 ± 0.3 b | 40.8 ± 3.6 a | 4.9 ± 0.1 bc |
| PS/RI | 50.2 ± 0.2 d | 8.5 ± 0.2 b | 41.4 ± 1.8 a | 4.9 ± 0.1 bc |
| LSD (0.05) | 14.063 | 3.601 | 11.127 | 1.384 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, O.-H.; Choi, H.-G. Yield, Flower Quality, and Photo-Physiological Responses of Cut Rose Flowers Grafted onto Three Different Rootstocks in Summer Season. Agronomy 2022, 12, 1468. https://doi.org/10.3390/agronomy12061468
Kwon O-H, Choi H-G. Yield, Flower Quality, and Photo-Physiological Responses of Cut Rose Flowers Grafted onto Three Different Rootstocks in Summer Season. Agronomy. 2022; 12(6):1468. https://doi.org/10.3390/agronomy12061468
Chicago/Turabian StyleKwon, O-Hyeon, and Hyo-Gil Choi. 2022. "Yield, Flower Quality, and Photo-Physiological Responses of Cut Rose Flowers Grafted onto Three Different Rootstocks in Summer Season" Agronomy 12, no. 6: 1468. https://doi.org/10.3390/agronomy12061468
APA StyleKwon, O.-H., & Choi, H.-G. (2022). Yield, Flower Quality, and Photo-Physiological Responses of Cut Rose Flowers Grafted onto Three Different Rootstocks in Summer Season. Agronomy, 12(6), 1468. https://doi.org/10.3390/agronomy12061468