Radiation Use Efficiency and Agronomic Performance of Biomass Sorghum under Different Sowing Dates

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Experiments and Measured Data
2.2. Computation and Statistical Analysis of Field Data
2.3. Economic Analysis
3. Results
3.1. Phenological Development
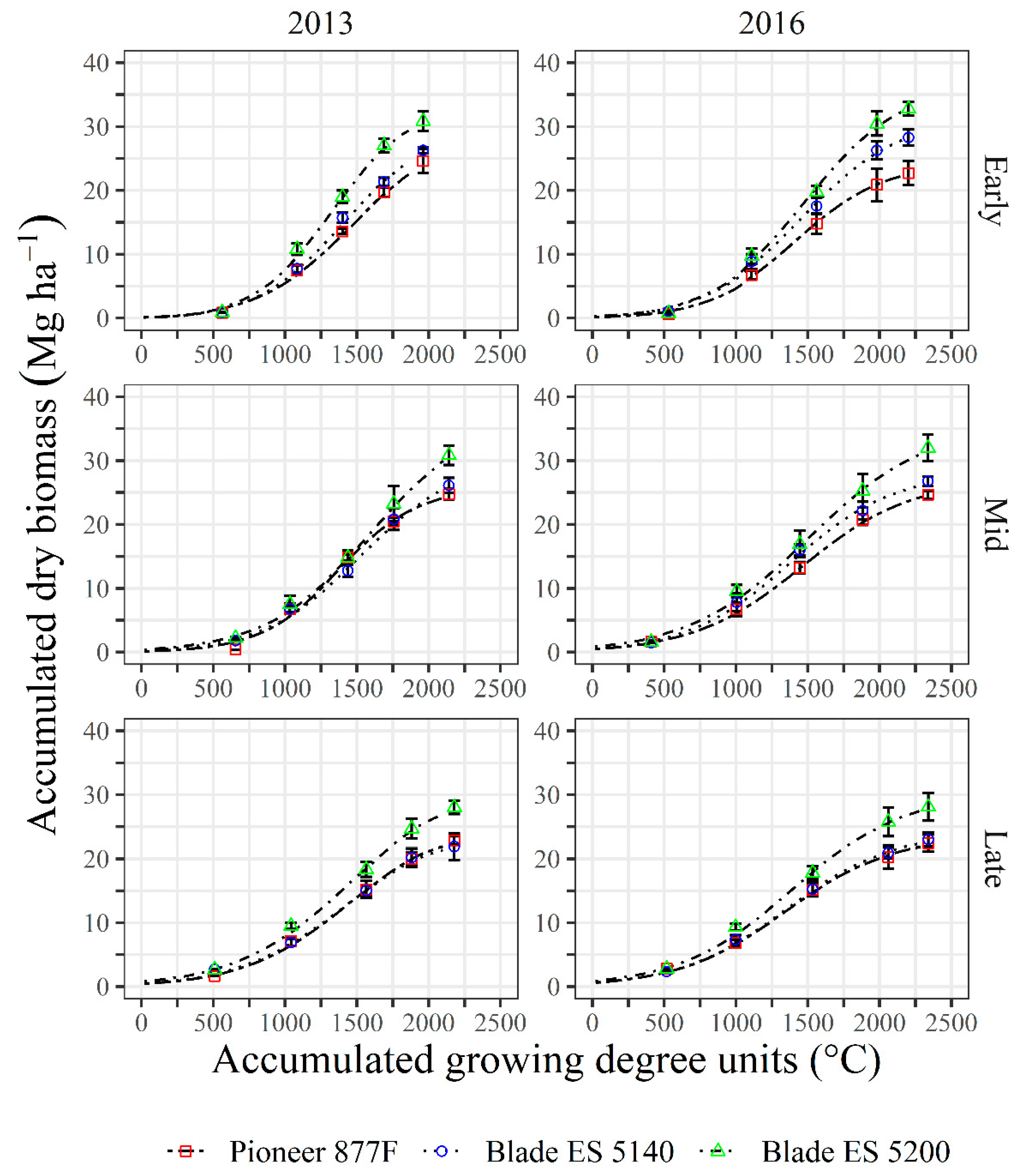
3.2. Dry Biomass Accumulation
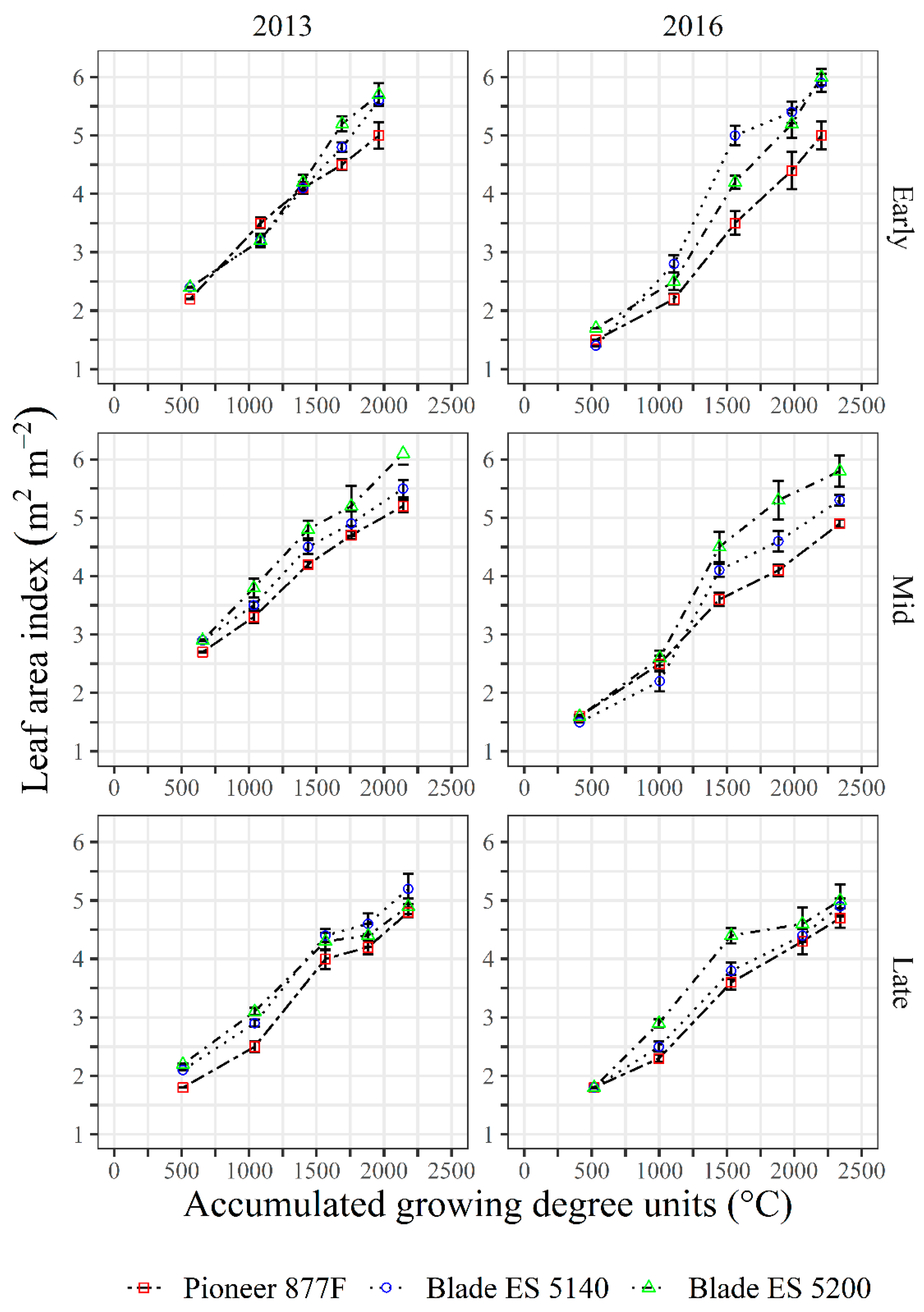
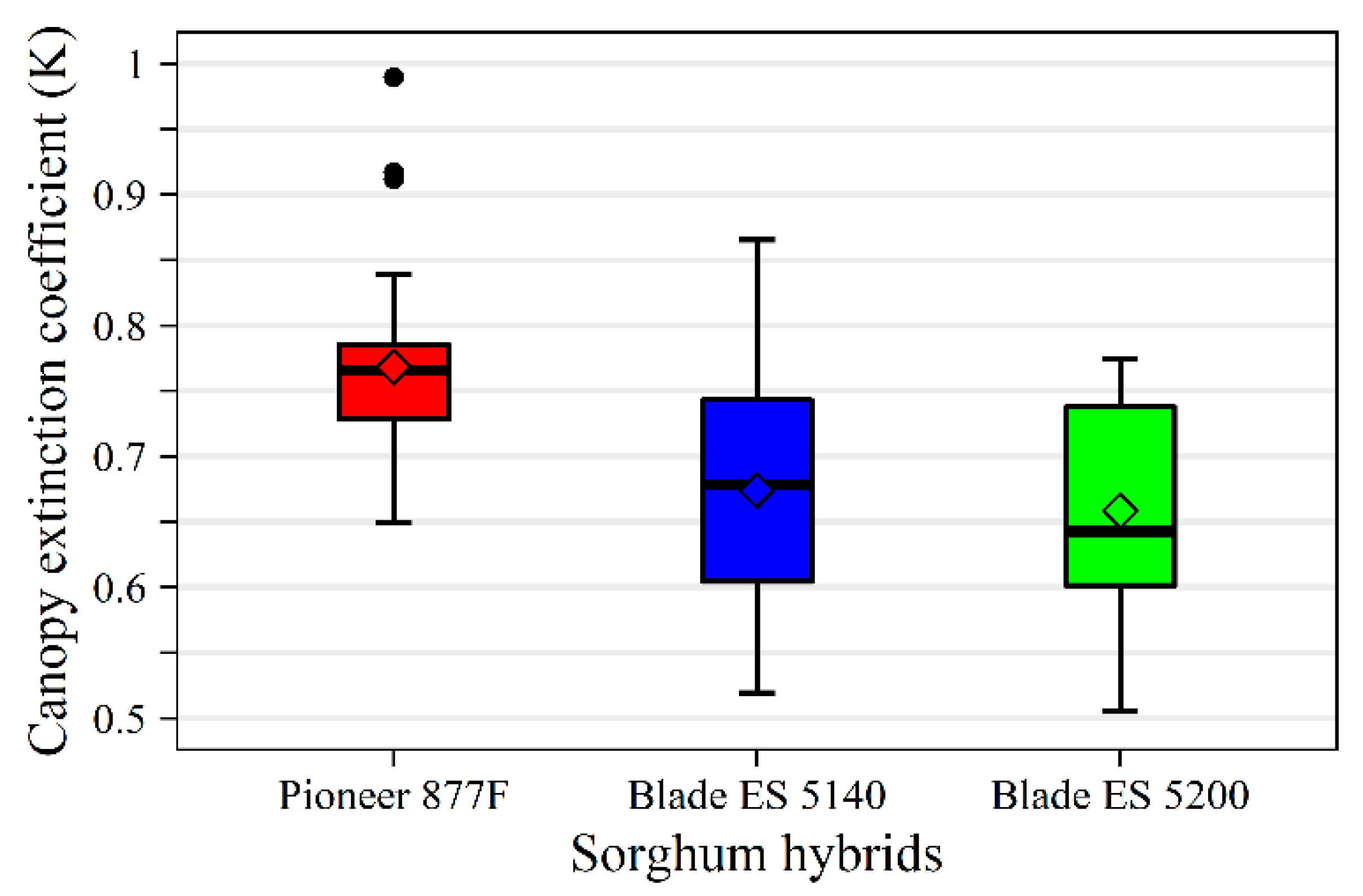
3.3. Leaf Area Index, Canopy Extinction Coefficients and Intercepted Photosynthetic Active Radiation
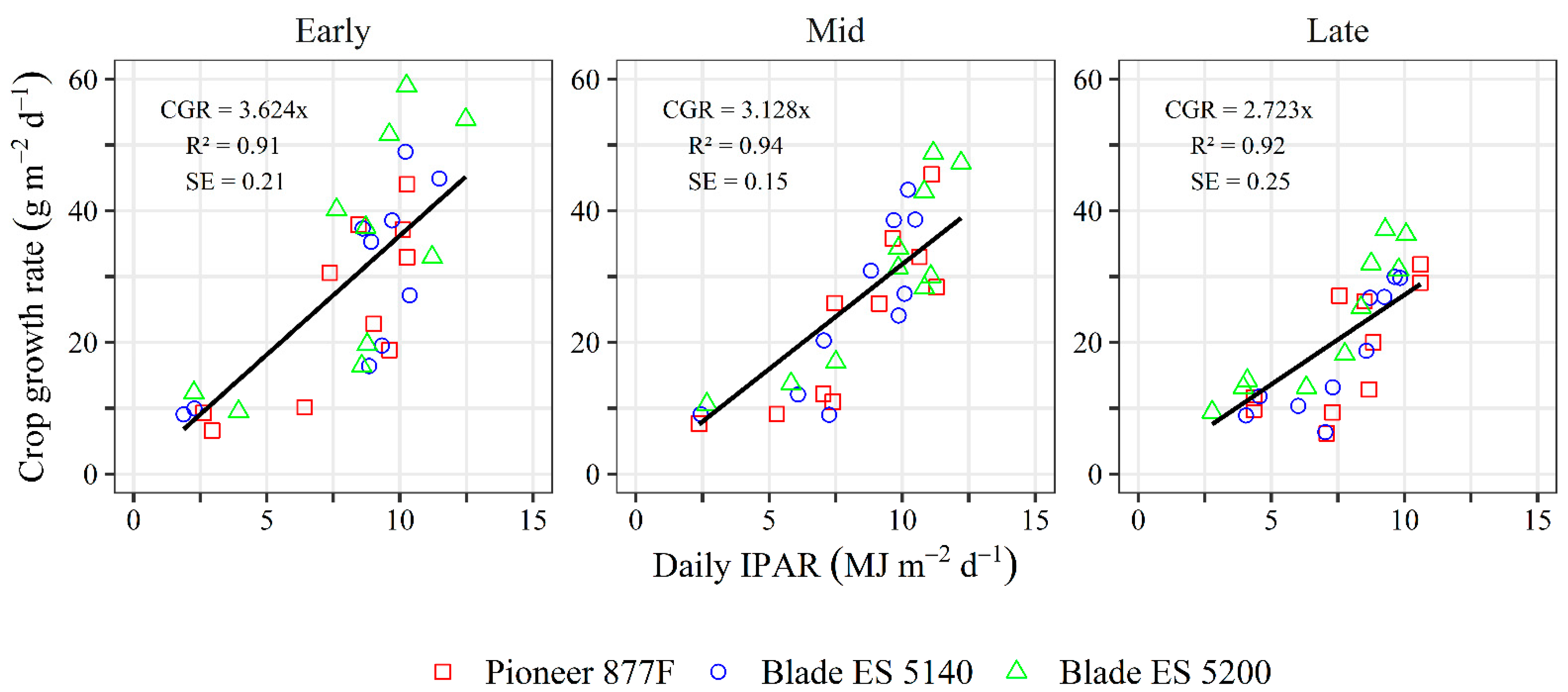
3.4. Plant Growth Analysis
3.5. Radiation Use Efficiency
3.6. Economic Feasibility Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Connor, D.J.; Loomis, R.S.; Cassman, K.G. Crop Ecology: Productivity and Management in Agricultural Systems; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Kiniry, J.; Jones, C.; O’Toole, J.; Blanchet, R.; Cabelguenne, M.; Spanel, D. Radiation-use efficiency in biomass accumulation prior to grain-filling for five grain-crop species. Field Crops Res. 1989, 20, 51–64. [Google Scholar] [CrossRef]
- Soltani, A.; Sinclair, T.R. Modeling Physiology of Crop Development, Growth and Yield; CABI: Wallingford, UK, 2012. [Google Scholar]
- Houx, J.H., III; Fritschi, F.B. Influence of late planting on light interception, radiation use efficiency and biomass production of four sweet sorghum cultivars. Ind. Crops Prod. 2015, 76, 62–68. [Google Scholar] [CrossRef]
- Rinaldi, M.; Garofalo, P. Radiation-use efficiency of irrigated biomass sorghum in a Mediterranean environment. Crop Pasture Sci. 2011, 62, 830–839. [Google Scholar] [CrossRef]
- Rooney, W.L.; Blumenthal, J.; Bean, B.; Mullet, J.E. Designing sorghum as a dedicated bioenergy feedstock. Biofuels Bioprod. Biorefin. 2007, 1, 147–157. [Google Scholar] [CrossRef]
- Enciso, J.; Chavez, J.C.; Ganjegunte, G.; Zapata, S.D. Energy Sorghum Production under Arid and Semi-Arid Environments of Texas. Water 2019, 11, 1344. [Google Scholar] [CrossRef] [Green Version]
- Enciso, J.; Jifon, J.; Ribera, L.; Zapata, S.; Ganjegunte, G. Yield, water use efficiency and economic analysis of energy sorghum in South Texas. Biomass Bioenergy 2015, 81, 339–344. [Google Scholar] [CrossRef]
- Garofalo, P.; Vonella, A.V.; Ruggieri, S.; Rinaldi, M. Water and radiation use efficiencies of irrigated biomass sorghum in a Mediterranean environment. Ital. J. Agron. 2011, 6, 21. [Google Scholar] [CrossRef]
- Rinaldi, M.; De Luca, D. Application of EPIC model to assess climate change impact on sorghum in southern Italy. Ital. J. Agron. 2012, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Teetor, V.H.; Duclos, D.V.; Wittenberg, E.T.; Young, K.M.; Chawhuaymak, J.; Riley, M.R.; Ray, D.T. Effects of planting date on sugar and ethanol yield of sweet sorghum grown in Arizona. Ind. Crops Prod. 2011, 34, 1293–1300. [Google Scholar] [CrossRef]
- Rao, S.S.; Patil, J.V.; Prasad, P.V.V.; Reddy, D.C.S.; Mishra, J.S.; Umakanth, A.V.; Reddy, B.V.S.; Kumar, A.A. Sweet sorghum planting effects on stalk yield and sugar quality in Semi-Arid tropical environment. Agron. J. 2013, 105, 1458–1465. [Google Scholar] [CrossRef] [Green Version]
- Hipp, B.W.; Cowley, W.R.; Gerard, C.J.; Smith, B.A. Influence of Solar Radiation and Date of Planting on Yield of Sweet Sorghum. Crop Sci. 1970, 10, 91–92. [Google Scholar] [CrossRef]
- Zapata, S.D.; Ribera, L.A.; Palma, M.A. Effect of Production Parameters on the Economic Feasibility of a Biofuel Enterprise. J. Agric. Appl. Econ. 2017, 49, 347–362. [Google Scholar] [CrossRef] [Green Version]
- Coyle, W.T. Next-Generation Biofuels: Near-Term Challenges and Implications for Agriculture; DIANE Publishing: Collingdale, PA, USA, 2010. [Google Scholar]
- Henggeler, J.C.; Enciso, J.M.; Multer, W.L.; Unruh, B.L. Deficit Subsurface Drip Irrigation of Cotton. In Deficit Irrigation Practices; FAO: Rome, Italy, 2002. [Google Scholar]
- Gerik, T.; Bean, B.W.; Vanderlip, R. Sorghum Growth and Development. In Texas FARMER Collection; Texas Cooperative Extension: College Station, TX, USA, 2003. [Google Scholar]
- Richards, F. A flexible growth function for empirical use. J. Exp. Bot. 1959, 10, 290–301. [Google Scholar] [CrossRef]
- Gregorczyk, A. The logistic function-its application to the description and prognosis of plant growth. Acta Soc. Bot. Pol. 1991, 60, 67–76. [Google Scholar] [CrossRef]
- Monteith, J.; Unsworth, M. Principles of Environmental Physics; Academic Press: Cambridge, MA, USA, 2007. [Google Scholar]
- Zhang, L.; Hu, Z.; Fan, J.; Zhou, D.; Tang, F. A meta-analysis of the canopy light extinction coefficient in terrestrial ecosystems. Front. Earth Sci. 2014, 8, 599–609. [Google Scholar] [CrossRef]
- Monsi, M.; Saeki, T. The light factor in plant communities and its significance for dry matter production. Jpn. J. Bot. 1953, 14, 22–52. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Mendiburu, F. agricolae: Statistical Procedures for Agricultural Research. R Package Version 1.3-5. 2021. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 17 April 2022).
- AMSU. National Biomass Energy Report. 2022. Available online: https://mymarketnews.ams.usda.gov/viewReport/2804 (accessed on 2 February 2022).
- Meki, M.N.; Ogoshi, R.M.; Kiniry, J.R.; Crow, S.E.; Youkhana, A.H.; Nakahata, M.H.; Littlejohn, K. Performance evaluation of biomass sorghum in Hawaii and Texas. Ind. Crops Prod. 2017, 103, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Dercas, N.; Liakatas, A. Water and radiation effect on sweet sorghum productivity. Water Resour. Manag. 2007, 21, 1585–1600. [Google Scholar] [CrossRef]
- Sinclair, T. Leaf Area Development in Field-Grown Soybeans 1. Agron. J. 1984, 76, 141–146. [Google Scholar] [CrossRef]
- Ceotto, E.; Di Candilo, M.; Castelli, F.; Badeck, F.-W.; Rizza, F.; Soave, C.; Volta, A.; Villani, G.; Marletto, V. Comparing solar radiation interception and use efficiency for the energy crops giant reed (Arundo donax L.) and sweet sorghum (Sorghum bicolor L. Moench). Field Crops Res. 2013, 149, 159–166. [Google Scholar] [CrossRef]
- Olson, S.N.; Ritter, K.; Rooney, W.; Kemanian, A.; McCarl, B.A.; Zhang, Y.; Hall, S.; Packer, D.; Mullet, J. High biomass yield energy sorghum: Developing a genetic model for C4 grass bioenergy crops. Biofuels Bioprod. Biorefin. 2012, 6, 640–655. [Google Scholar] [CrossRef]
- Bégué, A. Leaf area index, intercepted photosynthetically active radiation, and spectral vegetation indices: A sensitivity analysis for regular-clumped canopies. Remote Sens. Environ. 1993, 46, 45–59. [Google Scholar] [CrossRef]
- Narayanan, S.; Aiken, R.M.; Prasad, P.V.V.; Xin, Z.; Yu, J. Water and Radiation Use Efficiencies in Sorghum. Agron. J. 2013, 105, 649–656. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, T.R. A reminder of the limitations in using Beer’s Law to estimate daily radiation interception by vegetation. Crop Sci. 2006, 46, 2343–2347. [Google Scholar] [CrossRef]
- Hammer, G.L.; Vanderlip, R.L.; Gibson, G.; Wade, L.J.; Henzell, R.G.; Younger, D.R.; Warren, J.; Dale, A.B. Genotype-by-Environment Interaction in Grain Sorghum. II. Effects of Temperature and Photoperiod on Ontogeny. Crop Sci. 1989, 29, 376–384. [Google Scholar] [CrossRef]
- Muchow, R. Comparative productivity of maize, sorghum and pearl millet in a semi-arid tropical environment II. Effect of water deficits. Field Crops Res. 1989, 20, 207–219. [Google Scholar] [CrossRef]
- Mastrorilli, M.; Katerji, N.; Rana, G.; Steduto, P. Sweet sorghum in Mediterranean climate: Radiation use and biomass water use efficiencies. Ind. Crops Prod. 1995, 3, 253–260. [Google Scholar] [CrossRef]
- Demetriades-Shah, T.H.; Fuchs, M.; Kanemasu, E.T.; Flitcroft, I. A note of caution concerning the relationship between cumulated intercepted solar radiation and crop growth. Agric. For. Meteorol. 1992, 58, 193–207. [Google Scholar] [CrossRef]
- Lindquist, J.L.; Arkebauer, T.J.; Walters, D.T.; Cassman, K.G.; Dobermann, A. Maize radiation use efficiency under optimal growth conditions. Agron. J. 2005, 97, 72–78. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, J.T.; Singh, U.; Godwin, D.C.; Bowen, W.T. Cereal Growth, Development and Yield. In Understanding Options for Agricultural Production; Tsuji, G.Y., Hoogenboom, G., Thornton, P.K., Eds.; Springer: Dordrecht, The Netherlands, 1998; pp. 79–98. [Google Scholar]
- Childs, K.; Miller, F.R.; Cordonnier-Pratt, M.M.; Pratt, L.H.; Morgan, P.W.; Mullet, J.E. The Sorghum Photoperiod Sensitivity Gene, Ma3, Encodes a Phytochrome B. Plant Physiol. 1997, 113, 611–619. [Google Scholar] [CrossRef] [Green Version]
- Epplin, F.M.; Clark, C.D.; Roberts, R.K.; Hwang, S. Challenges to the Development of a Dedicated Energy Crop. Am. J. Agric. Econ. 2007, 89, 1296–1302. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agronomic Data | 2013 | 2016 | ||||
|---|---|---|---|---|---|---|
| Sowing date | 1 April | 9 May | 14 June | 4 April | 11 May | 15 June |
| Harvest date | 26 July | 29 August | 8 October | 2 August | 8 September | 13 October |
| Length of season | 116 days | 112 days | 116 days | 120 days | 120 days | 120 days |
| Sampling (DAS) | 42, 70, 86, 101, 116 | 36, 55, 76, 92, 112 | 26, 53, 80, 98, 116 | 32, 64, 87, 108, 120 | 22, 53, 75, 97, 120 | 26, 50, 77, 104, 120 |
| Min. Temp. | 21.9 °C | 24.4 °C | 24.3 °C | 23.7 °C | 25.1 °C | 24.9 °C |
| Max. Temp. | 32.6 °C | 34.8 °C | 34.5 °C | 33.9 °C | 35.4 °C | 35.8 °C |
| Water irrigation | 356 mm | 457 mm | 279 mm | 381 mm | 406 mm | 432 mm |
| Precipitation | 152 mm | 141 mm | 273 mm | 131 mm | 201 mm | 118 mm |
| Solar radiation | 2331 MJ m−2 | 2350 MJ m−2 | 2255 MJ m−2 | 2642 MJ m−2 | 2724 MJ m−2 | 2482 MJ m−2 |
| Total GDU | 1961 °C | 2141 °C | 2179 °C | 2177 °C | 2336 °C | 2342 °C |
| Daylight > 12:20 h | 116 days | 112 days | 91 days | 120 days | 120 days | 120 days |
| Year | Effect | DB | LAI | RUE |
|---|---|---|---|---|
| 2013 | Season (S) | 0.006 | 0.025 | 0.029 |
| Hybrid (H) | <0.001 | <0.001 | <0.001 | |
| 0.481 | 0.354 | 0.177 | ||
| 2016 | Season (S) | 0.009 | 0.032 | 0.002 |
| Hybrid (H) | <0.001 | <0.001 | <0.001 | |
| 0.061 | 0.159 | 0.014 |
| Sorghum Hybrid | Sowing Date | Average CGR | Maximum CGR |
|---|---|---|---|
| 2013 | |||
| Pioneer 877F | Early | 20.8 | 44.2 |
| Mid | 21.6 | 46.6 | |
| Late | 19.1 | 34.1 | |
| Blade ES 5140 | Early | 22.3 | 49.1 |
| Mid | 23.0 | 43.7 | |
| Late | 18.5 | 32.3 | |
| Blade ES 5200 | Early | 26.3 | 60.8 |
| Mid | 27.1 | 50.4 | |
| Late | 23.3 | 39.1 | |
| 2016 | |||
| Pioneer 877F | Early | 18.6 | 39.2 |
| Mid | 20.1 | 35.8 | |
| Late | 17.8 | 31.6 | |
| Blade ES 5140 | Early | 23.4 | 45.9 |
| Mid | 21.5 | 39.2 | |
| Late | 18.3 | 32.5 | |
| Blade ES 5200 | Early | 27.2 | 54.6 |
| Mid | 25.6 | 43.0 | |
| Late | 22.4 | 38.3 | |
| Sowing Date | Biomass Sorghum Hybrid | Revenue (USD ha−1) | Production Cost (USD ha−1) | Net Return (USD ha−1) | Average Cost per Unit of Mass (USD Mg−1) |
|---|---|---|---|---|---|
| Early | Pioneer 877F | 2082.67 | 1522.12 | 560.55 | 64.43 |
| Blade ES 5140 | 2392.86 | 1560.74 | 832.12 | 57.30 | |
| Blade ES 5200 | 2774.12 | 1592.10 | 1182.01 | 50.41 | |
| Mid | Pioneer 877F | 2102.25 | 1540.54 | 561.71 | 62.12 |
| Blade ES 5140 | 2254.70 | 1515.89 | 738.82 | 56.99 | |
| Blade ES 5200 | 2669.90 | 1551.21 | 1118.68 | 49.26 | |
| Late | Pioneer 877F | 2263.04 | 1512.72 | 750.32 | 66.79 |
| Blade ES 5140 | 2243.72 | 1509.18 | 734.54 | 67.22 | |
| Blade ES 5200 | 2804.78 | 1549.72 | 1255.06 | 55.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chavez, J.C.; Ganjegunte, G.K.; Jeong, J.; Rajan, N.; Zapata, S.D.; Ruiz-Alvarez, O.; Enciso, J. Radiation Use Efficiency and Agronomic Performance of Biomass Sorghum under Different Sowing Dates. Agronomy 2022, 12, 1252. https://doi.org/10.3390/agronomy12061252
Chavez JC, Ganjegunte GK, Jeong J, Rajan N, Zapata SD, Ruiz-Alvarez O, Enciso J. Radiation Use Efficiency and Agronomic Performance of Biomass Sorghum under Different Sowing Dates. Agronomy. 2022; 12(6):1252. https://doi.org/10.3390/agronomy12061252
Chicago/Turabian StyleChavez, Jose C., Girisha K. Ganjegunte, Jaehak Jeong, Nithya Rajan, Samuel D. Zapata, Osias Ruiz-Alvarez, and Juan Enciso. 2022. "Radiation Use Efficiency and Agronomic Performance of Biomass Sorghum under Different Sowing Dates" Agronomy 12, no. 6: 1252. https://doi.org/10.3390/agronomy12061252
APA StyleChavez, J. C., Ganjegunte, G. K., Jeong, J., Rajan, N., Zapata, S. D., Ruiz-Alvarez, O., & Enciso, J. (2022). Radiation Use Efficiency and Agronomic Performance of Biomass Sorghum under Different Sowing Dates. Agronomy, 12(6), 1252. https://doi.org/10.3390/agronomy12061252