
Variations in Plant Growth Characteristics Due to Oxygen Plasma Irradiation on Leaf and Seed


Abstract
1. Introduction
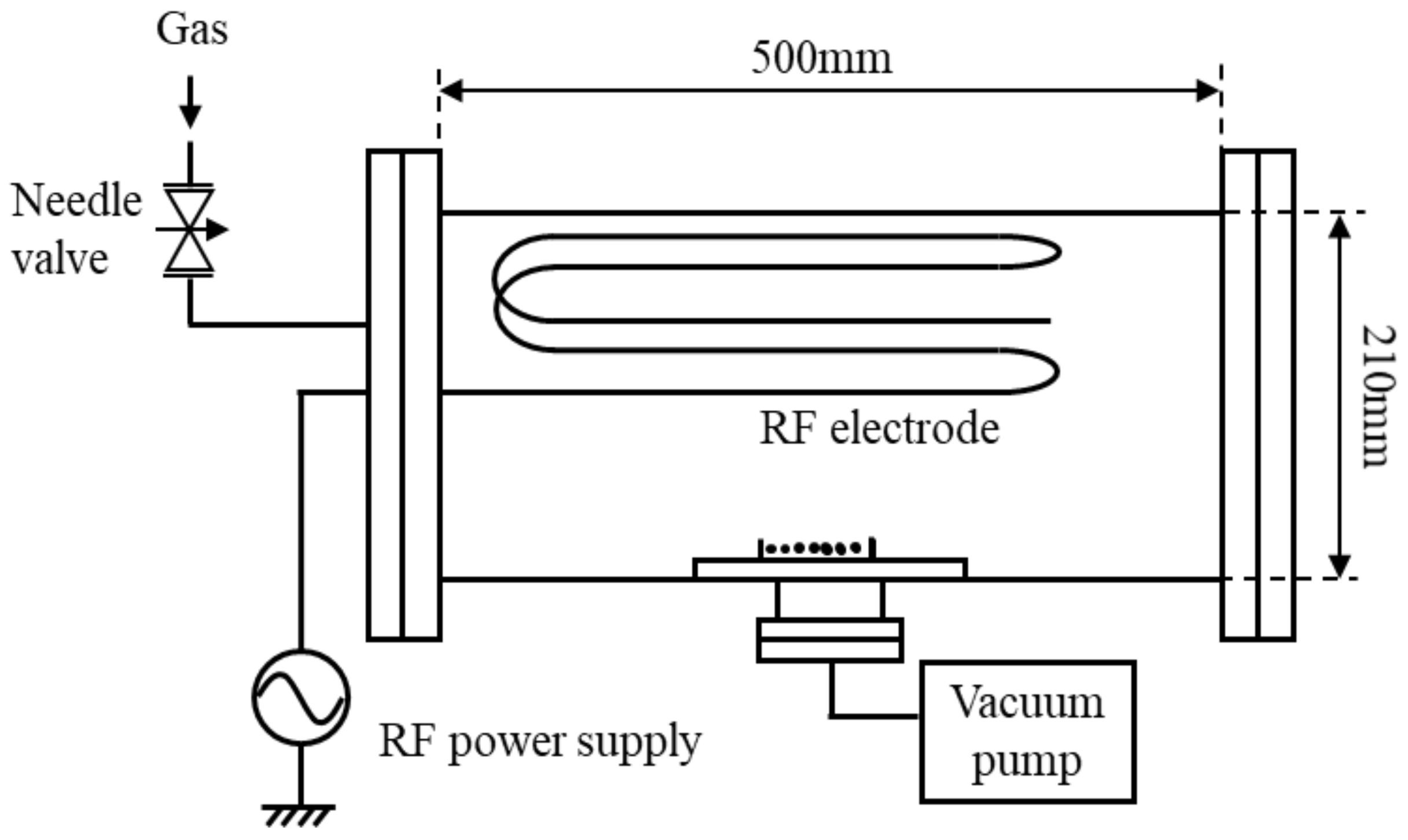
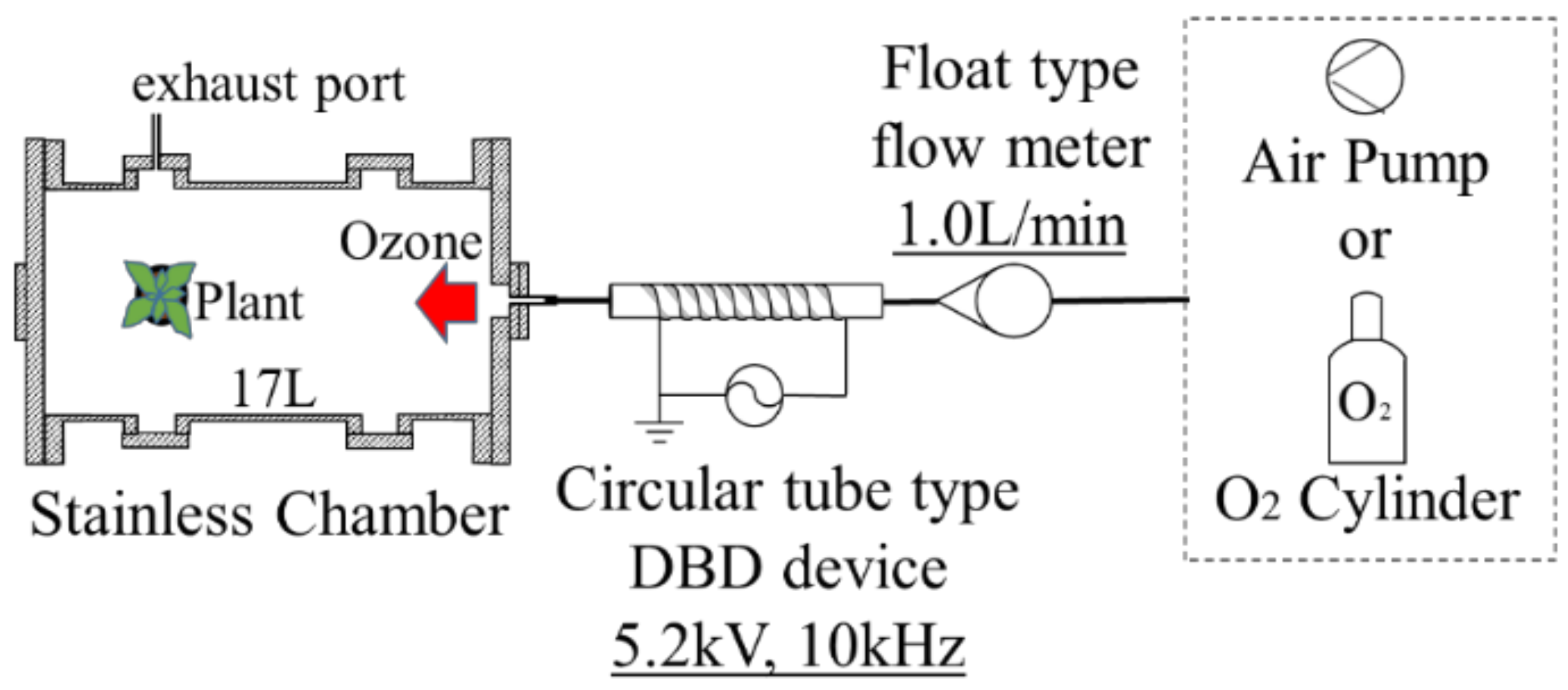
2. Experimental Apparatus and Methods
3. Experimental Results and Discussion
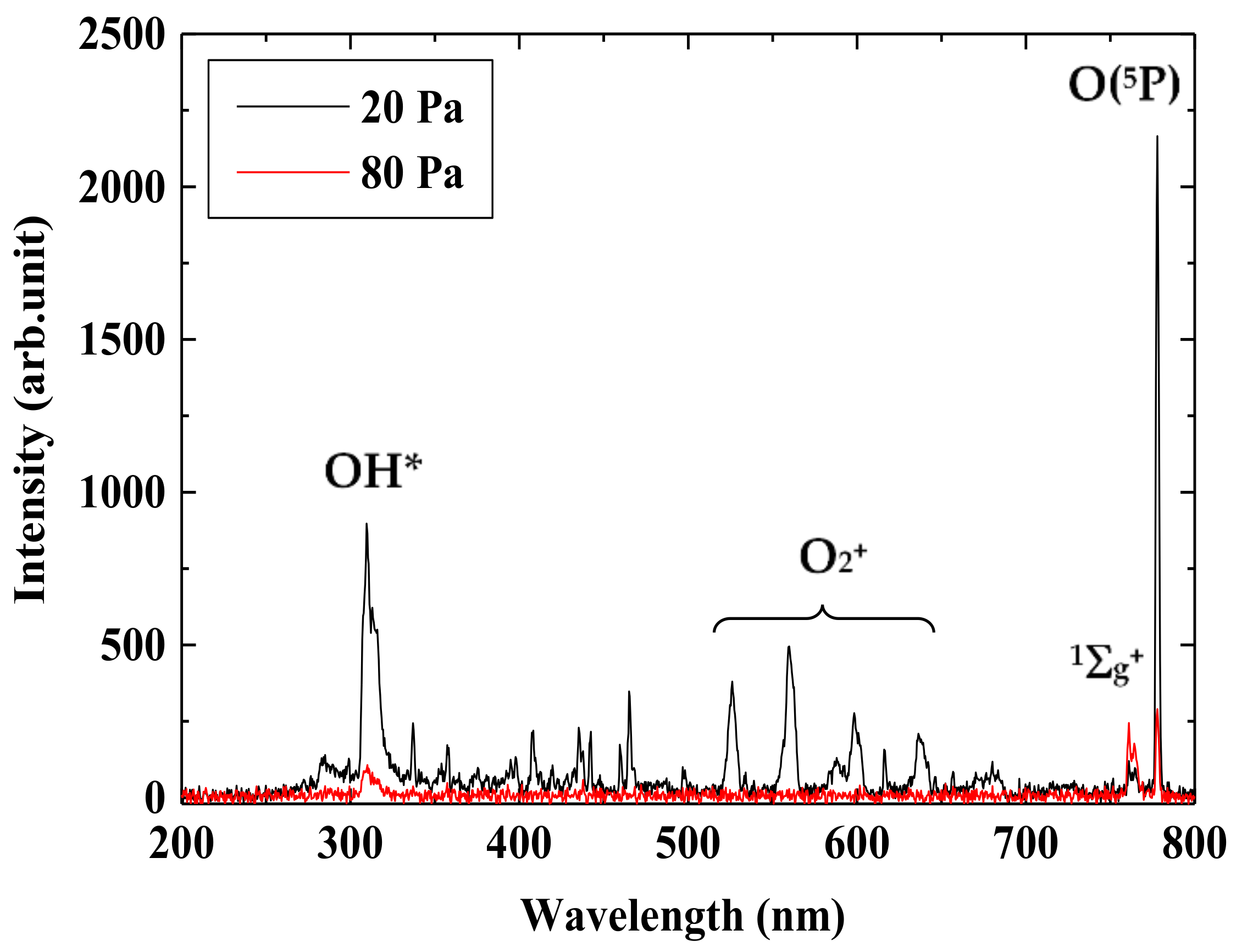
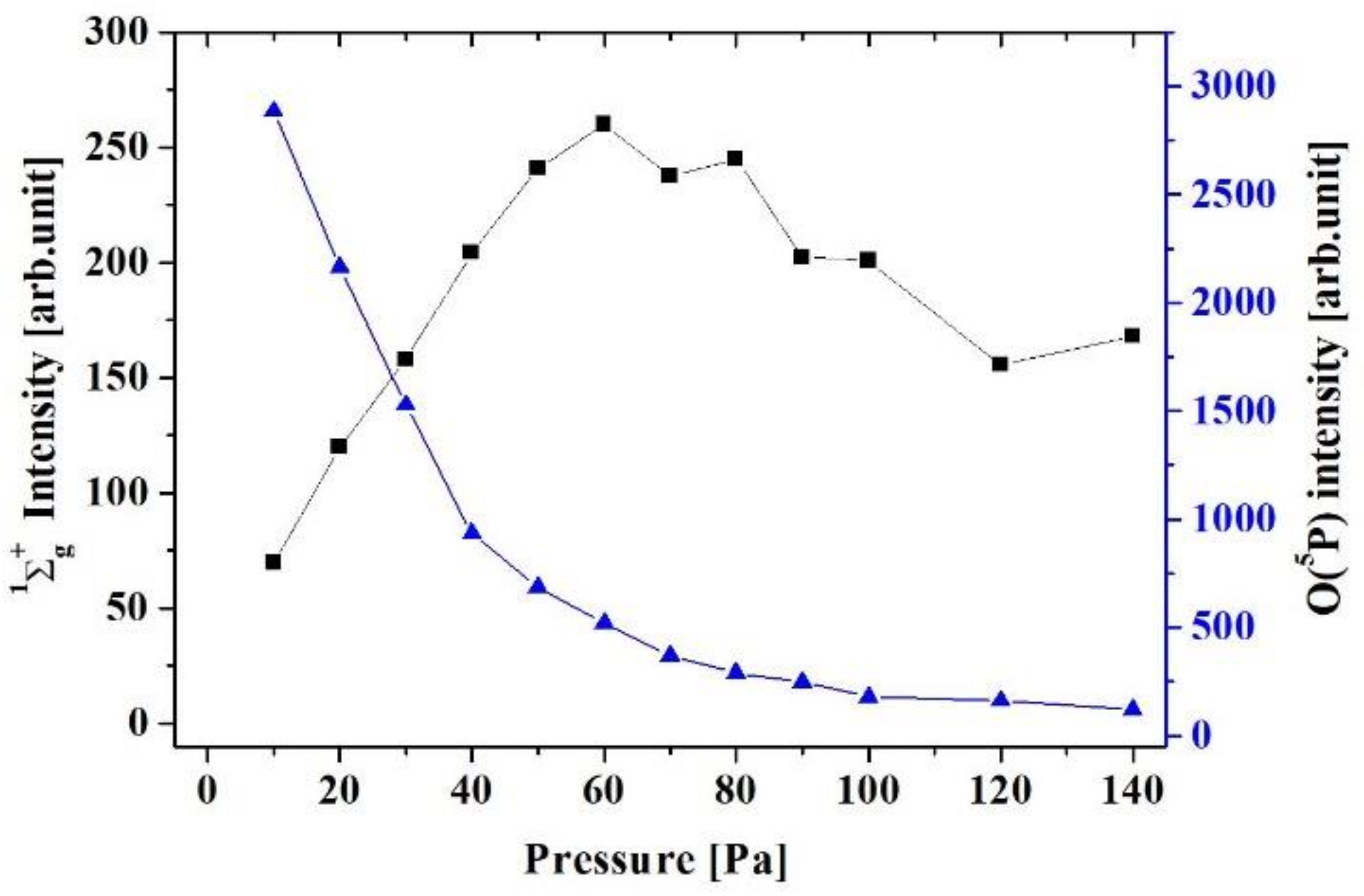
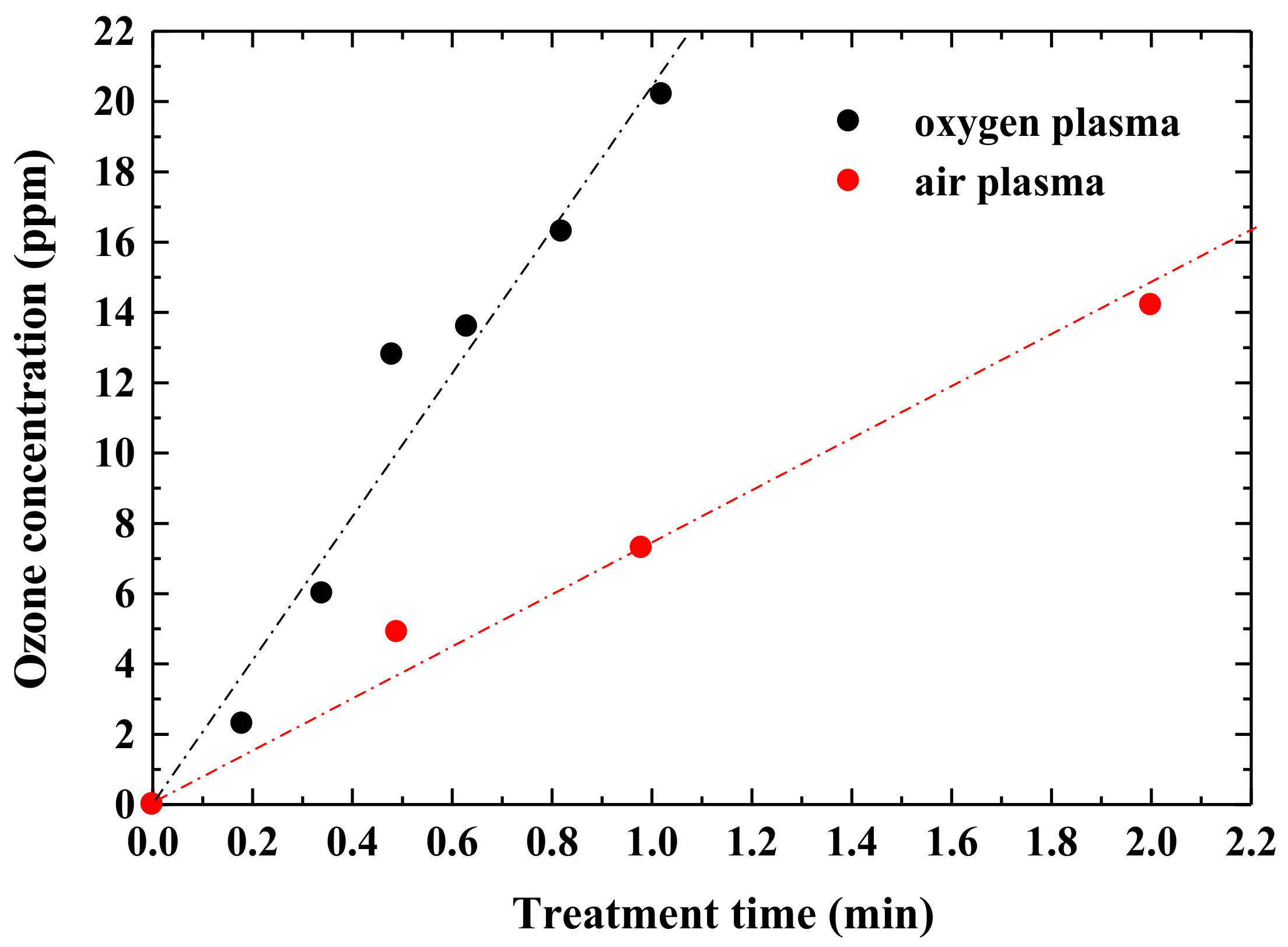
3.1. Production of Active Oxygen Species
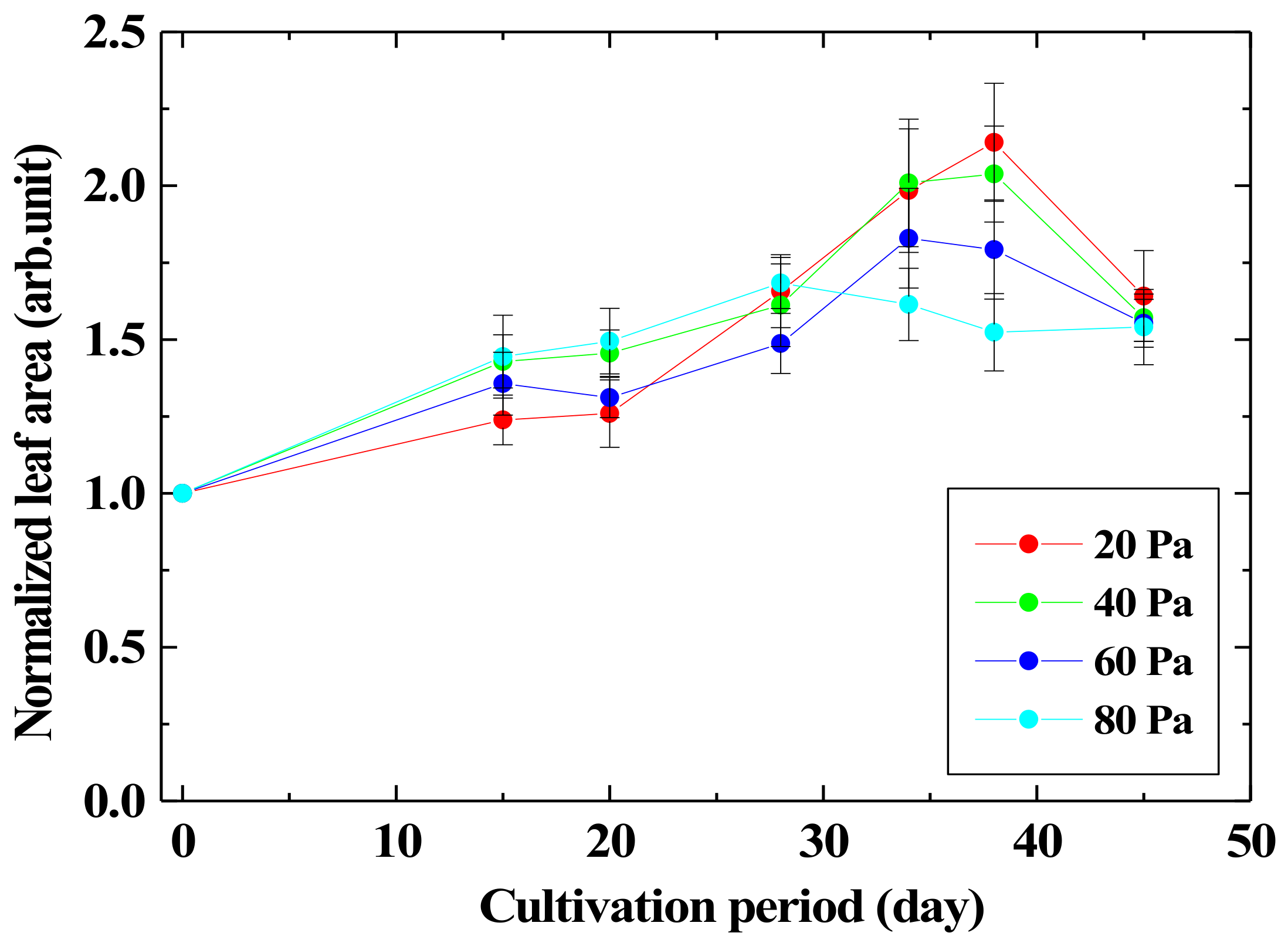
3.2. Growth Enhancement Characteristics of Leaf by Irradiating Seed with Oxygen Plasma
3.3. Growth Enhancement Characteristics of Leaf by Irradiating Seed with Ozone
3.4. Gene Expression Analysis of Leaf by Irradiating Seeds with Oxygen Plasma
- (1)
- Improvement of antioxidative activity: Increased expression of the gene group was observed. Table 2 shows the GO terms of the increased gene expression concerning antioxidative ability by irradiating seeds with oxygen plasma, where the p-value is the probability that the null hypothesis of a GO is adopted, and the Benjamini is the ratio of an acceptable correct null hypothesis, even if the null hypothesis is rejected. Moreover, the count is the number of genes corresponding to the GO. The GO in the table involved in enzymes with antioxidant activity such as oxidoreductase and superoxide dismutase (SOD) by irradiating oxygen plasma for 3 min. The redox enzymes, oxidoreductase, reduces the active oxygen in the plant cell. It is inferred that the increase of gene expression of the oxidoreductase enhances the sugar production by photosynthesis and the enhancement of growth by removing active oxygen in cells and blocking the inhibition of the dark reaction. Although the gene expression of the CuZn-SOD has been enhanced by the irradiation with ozone and O(1D), the CuZn-SOD does not act as a catalyst for the removal of active oxygen. As ozone and O (1D) are the same oxidizing substances for DNA and related molecules as O2−, it is possible that the gene expression of the CuZn-SOD has been induced by the irradiation with ozone and O(1D) due to oxidation of transcription factor, even though the CuZn-SOD does not act as a catalyst for the removal of active oxygen. Moreover, GO terms of iron and copper appear in this table. The corresponding functions of these terms are the production of iron and copper compounds in the plant. Since iron and copper are contained in the active center of coenzymes for Oxidoreductase and SOD and are indispensable to activate the catalytic effect of these enzymes, their expression is considered to have changed with Oxidoreductase and SOD by irradiation with active oxygen. The GO term of Removal of superoxide radicals represents the functions of the superoxide removal using methods other than SOD, such as ascorbic acid, flavonoids, and polyphenols. From the above, it was found that the improvement of antioxidant activity occurs over a long period of time by changing the gene expression by short-time oxygen plasma irradiation.
- (2)
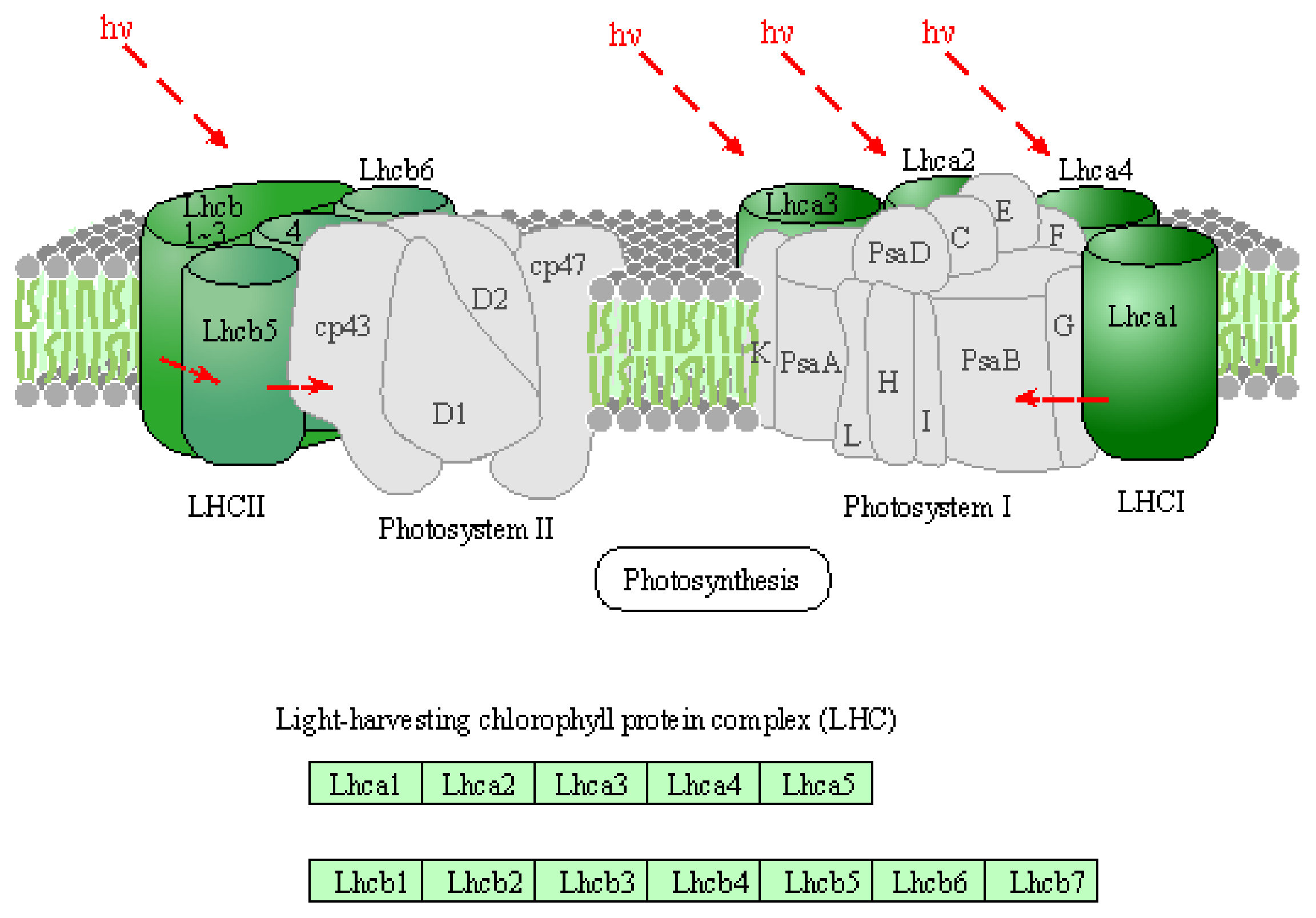
- Enhancement of photosynthesis: From the expression analysis results, it was found that the expression of genes related to the photosynthetic electron transport chain, including chlorophyll, was significantly increased by irradiation for 3 min, as shown in Table 3. The photosynthetic reaction obtains energy from light and produces organic molecules such as ATP from carbon dioxide and water in the atmosphere. Some enzymes involved in the Calvin cycle have one or more sets of -SH groups (thiol groups) that respond to redox, and when these are oxidized, they become -S-S- bonds (disulfide bonds). Enzymes with disulfide bonds are known to have reduced activity. Antioxidants induced by short-lifetime active oxygen irradiation may reduce the enzymes involved in the Calvin cycle and enhance the carbon fixation reaction. Figure 9 shows the KEGG pathway for the light-collecting chlorophyll protein complex of green plants whose gene expression was significantly increased by irradiation seeds with active oxygen species in the oxygen plasma for 3 min. Acronyms Lhca1-a5 in LHCI and Lhcb1-b6 in LHCII stand for light-harvesting chlorophyll protein complexes [36]. Moreover, acronyms in Photosystem I and II refer to the reference pages [37,38]. It is considered that the plasma irradiation for a relatively short time, 3 min enhanced the function of photosynthesis by increasing the expression of some genes, and promoted the production of energy such as sugar and ATP essential for protein synthesis and then plant growth. On the other hand, when the seeds were irradiated with oxygen plasma for a long time, 30 min, there was no significant difference in the function of expressed genes, which was significantly different from that of the leaves irradiated for 3 min. There are two possibilities for the lack of growth-enhancement effect after 30-min irradiation: (i) direct non-gene-mediated damage by active oxygen species to the seeds or (ii) the Calvin cycle catalytic enzyme is inactivated by being oxidized by plasma irradiation, the Calvin cycle is suppressed and the growth of leaves is suppressed.
- (3)
- Activation of molecular chaperone: Hsp90 had a high gene expression level after 30 min of irradiation. Hsp90 is a molecular chaperone that interacts with various proteins, has a function of preventing protein aggregation during stress, and at the same time plays an important role in signal transduction [39,40]. The increased expression of HSP suggests that plasma irradiation caused abnormalities in the protein biosynthesis system and that the growth-enhancement effect was suppressed compared to 3-min irradiation.
- (4)
- Suppression of DNA methylation: Gene concerning DNA methylation and epigenetics are found to be expressed by irradiation with the oxygen plasma [7,8,9]. The plasma irradiation for 3 min reduced the expression level of genes related to DNA methylation. Transcription and translation of the methylated portion of DNA are suppressed. Details of the observed epigenetic gene expression induced by the plasma irradiation are described in the previous papers [7,8,9].
- (5)
- Decrease of DNA replication: The rate of cell division in the leaves decreases. On the other hand, as the leaf area was expanded, it is considered that this area expansion is not due to the increase in the rate of cell division but to the cell elongation. However, no increase in gene expression related to cell elongation could be confirmed. From these facts, it is possible that the expansion of the leaf area is due to the increase in turgor pressure of the cells due to the storage of water. If the photosynthetic function is improved, it may require a large amount of H2O. In addition, the pores used for gas exchange with the atmosphere, which are essential for photosynthesis, close to suppress transpiration when the turgor pressure is low. Thus, storing water and keeping the turgor pressure high is necessary for efficient photosynthesis.
- Plasma irradiation of seeds
- Seeds are subjected to oxidative stress and improved antioxidant activity
- Obtained functions remains after germination (methylation: epigenetics)
- Improvement of leaf antioxidant activity
- Increase chlorophyll and activate photosynthetic electron transfer reaction
- Increase water storage to improve photosynthetic efficiency, adjust circadian rhythm
- Increase ATP synthesis
- Promote growth (increase in leaf area).
3.5. Gene Expression Analysis of Leaf Irradiated with Ozone
- Ozone exposure to leaves
- Damage to the photosystem antenna complex due to oxidative stress
- Decrease in the reaction rate of photosynthetic electron transfer reaction
- Increased production of chaperone for protein repair
- Decrease of ATP and sugar synthesis amount due to carbon fixation reaction
- Protein decrease in biosynthesis
- Promote growth (increase in leaf area)
3.6. Comparison of Leaf Gene Expression Variation Analysis before and after Germination
4. Conclusions
- The leaf area of Arabidopsis thaliana promotes growth up to 2.1 times by irradiation of seeds with low-pressure oxygen plasma. On the other hand, plasma irradiation of the leaves suppressed the growth about 0.8 times.
- Plasma irradiation of seeds significantly increased gene expression related to leaf photosynthesis, antioxidant activity, and water storage. From this, it is presumed that the production of NADPH was increased by photosynthesis as the oxidation resistance to plasma irradiation and the water storage was improved accordingly, leading to the promotion of growth.
- Comparing the results of gene expression analysis of seeds and leaves after the plasma irradiation of seeds, it was confirmed that both seeds and leaves had increased gene expression related to the same function, such as photosynthesis. This suggests that plasma irradiation of seeds also affects leaf gene expression, and it is considered that this phenomenon is related to epigenetic changes such as methylation.
- Ozone exposure to leaves reduced gene expression related to leaf photosynthesis. On the other hand, the antioxidant activity of the leaves increased.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubinov, A.E.; Lazarenko, E.R.; Selemir, V.D. Effect of glow discharge air plasma on grain crops seed. IEEE Trans. Plasma Sci. 2000, 28, 180–183. [Google Scholar] [CrossRef]
- Hayashi, N.; Ono, R.; Uchida, S. Growth Enhancement of Plant by Plasma and UV Light Irradiation to Seeds. J. Photopolym. Sci. Technol. 2015, 28, 445–448. [Google Scholar] [CrossRef][Green Version]
- Ono, R.; Hayashi, N. Variation of antioxidative activity and growth enhancement of Brassicaceae induced by low-pressure oxygen plasma. Jpn. J. Appl. Phys. 2015, 54, 06GD03-1–06GD03-4. [Google Scholar] [CrossRef]
- Jiafeng, J.; Xin, H.; Ling, L.; Jiangang, L.; Hanliang, S.; Qilai, X.; Renhong, Y.; Yuanhua, D. Effect of Cold Plasma Treatment on Seed Germination and Growth of Wheat. Plasma Sci. Technol. 2014, 16, 54–58. [Google Scholar]
- Ling, L.; Jiafeng, J.; Jiangang, L.; Minchong, S.; Xin, H.; Hanliang, S.; Yuanhua, D. Effects of cold plasma treatment on seed germination and seedling growth of soybean. Sci. Rep. 2014, 4, 5859. [Google Scholar] [CrossRef]
- Watanabe, S.; Ono, R.; Hayashi, N.; Tashiro, K.; Kuhara, S.; Inoue, A.; Yasuda, K.; Hagiwara, H. Growth Enhancement and Gene Expression of Arabidopsis thaliana irradiated by active oxygen species. Jpn. J. Appl. Phys. 2016, 55, 07LG10-1–07LG10-6. [Google Scholar] [CrossRef]
- Nakano, R.; Tashiro, K.; Aijima, R.; Hayashi, N. Effect of oxygen plasma irradiation on gene expression in plant seeds induced by active oxygen species. Plasma Med. 2016, 6, 303–313. [Google Scholar] [CrossRef]
- Hayashi, N.; Ono, R.; Nakano, R.; Shiratani, M.; Tashiro, K.; Kuhara, S.; Yasuda, K.; Hagiwara, H. DNA microarray analysis of plant seeds irradiated by active oxygen species in oxygen plasma. Plasma Med. 2016, 6, 459–471. [Google Scholar] [CrossRef]
- Nakano, R.; Yamashita, Y.; Kobayashi, A.; Hayashi, N. Gene expression effect of plant seeds by irradiation with low-pressure oxygen plasma. J. IAPS 2018, 26, 91–95. [Google Scholar]
- Koichi, T.; Nobuya, H.; Douyan, W.; Takayuki, O. High-voltage technologies for agriculture and food processing. J. Phys. D Appl. Phys. 2019, 52, 473001. [Google Scholar]
- Maruyama-Nakashita, A.; Ishibashi, Y.; Yamamoto, K.; Zhang, L.; Morikawa-Ichinose, T.; Kim, S.; Hayashi, N. Oxygen plasma modulates glucosinolate levels without affecting lipid contents and composition in Brassica napus seeds. Biosci. Biotechnol. Biochem. 2021, 85, 2434–2441. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, N.; Yagyu, Y. Treatment of protein using oxygen plasma produced by RF discharge. Trans. Mater. Res. Soc. Jpn. 2008, 33, 791–794. [Google Scholar] [CrossRef]
- Yagyu, Y.; Hayashi, N.; Guan, W.; Kawasaki, H. Influence of Atomic and Singlet Molecular Oxygen Generated by RF Plasma on Reduction of Protein. J. Plasma Fusion Res. 2009, 8, 578–581. [Google Scholar]
- Hayashi, N.; Nakahigashi, A.; Liu, H.; Goto, M. Treatment of second order structures of protein using oxygen RF plasma. Jpn. J. Appl. Phys. 2010, 49, 08JH02-1–08JH02-4. [Google Scholar] [CrossRef]
- Hayashi, N.; Kometani, R.; Yoshida, Y. Treatment of dipicolinic acid and inactivation mechanism of thermophile spore using active oxygen. Jpn. J. Appl. Phys. 2013, 52, 11NF03-1–11NF03-4. [Google Scholar] [CrossRef]
- Ono, R.; Tanaka, A.; Uchida, S.; Kitazaki, S.; Itarashiki, T.; Hayashi, N. Effect of Active Oxygen Species in Low Pressure Oxygen Plasma on Antioxidative Substances. Front. Appl. Plasma Technol. 2014, 7, 45–46. [Google Scholar]
- Hayashi, N.; Ono, R.; Yagyu, Y.; Yonesu, A. Application of atmospheric discharge plasma to agricultural and marine products. J. Jpn. Soc. Appl. Electromagn. Mech. 2014, 22, 447–452. [Google Scholar]
- Hayashi, N.; Miyamaru, Y.; Aijima, R.; Yamashita, Y. Activation of p53-mediated apoptosis pathway in HSC3 cancer cell irradiated by atmospheric DBD oxygen plasma. IEEE Trans. Plasma Sci. 2018, 47, 1093–1099. [Google Scholar] [CrossRef]
- Hayashi, N.; Inoue, Y.; Kyumoto, Y.; Kukita, T. Characteristics of differentiation of osteoclast cells irradiated with active species in atmospheric oxygen plasma. Jpn. J. Appl. Phys. 2020, 59, SJJF02. [Google Scholar] [CrossRef]
- Subaedah, S.; Uematsu, H.; Hayashi, N. Activation of EL-4 T-cells by irradiation with atmospheric oxygen plasma. Jpn. J. Appl. Phys. 2020, 59, SJJF03. [Google Scholar] [CrossRef]
- Gordon, G. The chemistry and reactions of ozone in our environment. Prog. Nucl. Energy 1995, 29, 89–96. [Google Scholar] [CrossRef]
- Mitsugi, F.; Abiru, T.; Ikegami, T.; Ebihara, K.; Aoqui, S.; Nagahama, K. Influence of Ozone Generated by Surface Barrier Discharge on Nematode and Plant Growth. IEEE Trans. Plasma Sci. 2016, 44, 3071–3076. [Google Scholar] [CrossRef]
- Ohshiro, S.; Katsuto, M.; Satahira, K.; Iriyama, Y.; Nakamura, K.; Ito, S.; Ihara, T. Fabrication of the Plasma-Chemical Indicator and It’s Application. J. Photopolym. Sci. 2013, 26, 533–538. [Google Scholar] [CrossRef]
- Satahira, K.; Ohshiro, S.; Nakamura, K.; Ito, S.; Ihara, T. Visualization Plasma Diagnosis Indicator Using Discoloration Reaction of Au and Pt Ions. J. Photopolym. Sci. 2015, 28, 435–438. [Google Scholar] [CrossRef][Green Version]
- Bolstad, B.M.; Irizarry, R.A.; Åstrand, M.; Speed, T.P. A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinformatics 2003, 19, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Gentleman, R.C.; Carey, V.J.; Bates, D.M.; Bolstad, B.; Dettling, M.; Dudoit, S.; Ellis, B.; Gautier, L.; Ge, Y.; Gentry, J.; et al. Bioconductor: Open software development for computational biology and bioinformatics. Genome Biol. 2004, 5, R80. [Google Scholar] [CrossRef] [PubMed]
- Quackenbush, J. Microarray data normalization and transformation. Nat. Genet. 2002, 32, 496–501. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef]
- Germany, G.A.; Anderson, R.J.; Salamo, G.J. Electron impact excitation of the 3p(5P) state of atomic oxygen. J. Chem. Phys. 1988, 89, 1999–2002. [Google Scholar] [CrossRef]
- Katsch, H.M.; Tewes, A.; Quandt, E.; Goehlich, A.; Kawetzki, T.; Döbele, H.F. Detection of atomic oxygen: Improvement of actinometry and comparison with laser spectroscopy. J. Appl. Phys. 2000, 88, 6232–6238. [Google Scholar] [CrossRef]
- Kaufman, F. The air afterglow and its use in the study of some reactions of atomic oxygen. Proc. R. Soc. Lond. Ser. A 1958, 247, 123–139. [Google Scholar]
- Waskoenig, J.; Niemi, K.; Knake, N.; Graham, L.M.; Reuter, S.; der Gathen, V.S.; Gans, T. Atomic oxygen formation in a radio-frequency driven micro-atmospheric pressure plasma jet. Plasma Sources Sci. Technol. 2010, 19, 045018. [Google Scholar] [CrossRef]
- Rich, S. Ozone damage to plants. Annu. Rev. Phytopathol. 1964, 2, 253–266. [Google Scholar] [CrossRef]
- Miller, P.R.; Parmeter, J.R., Jr.; Taylor, O.C.; Cardiff, E.A. Ozone injury to the foliage of Pinus ponderosa. Phytopathology 1963, 53, 1072–1076. [Google Scholar]
- Kyoto Encyclopedia of Genes and Genomes, LHC. Available online: https://www.genome.jp/dbget-bin/get_linkdb?-t+genes+path:ath00196 (accessed on 1 December 2021).
- Kyoto Encyclopedia of Genes and Genomes, Photosystem I. Available online: https://www.kegg.jp/kegg-bin/view_ortholog_table?md=M00163 (accessed on 1 December 2021).
- Kyoto Encyclopedia of Genes and Genomes, Photosystem II. Available online: https://www.kegg.jp/kegg-bin/view_ortholog_table?md=M00161 (accessed on 1 December 2021).
- Richter, K.; Buchne, J. Hsp90: Chaperoning signal transduction. J. Cell. Physiol. 2001, 188, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Prodromou, C. Mechanisms of Hsp90 regulation. Biochem. J. 2016, 473, 2439–2452. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Gao, J.; Kim, J.I.; Chen, K.; Bressan, R.A.; Zhu, J.-K. Control of Plant Water Use by ABA Induction of Senescence and Dormancy: An Overlooked Lesson from Evolution. Plant Cell Physiol. 2017, 58, 1319–1327. [Google Scholar] [CrossRef] [PubMed]
- Ron, D. Translational control in the endoplasmic reticulum stress response. J. Clin. Investig. 2002, 110, 1383–1388. [Google Scholar] [CrossRef]
- Gelhaye, E.; Rouhier, N.; Navrot, N.; Jacquot, J.P. The plant thioredoxin system. Cell. Mol. Life Sci. 2005, 62, 24–35. [Google Scholar] [CrossRef]
- Lu, J.; Holmgren, A. The thioredoxin antioxidant system. Free Radic. Biol. Med. 2014, 66, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Edman, J.C.; Ellis, L.; Blacher, R.W.; Roth, R.A.; Rutter, W.J. Sequence of protein disulphide isomerase and implications of its relationship to thioredoxin. Nature 1985, 317, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Freedman, R.B.; Hirst, T.R.; Tuite, M.F. Protein disulphide isomerase: Building bridges in protein folding. Trends Biochem. Sci. 1994, 19, 331–336. [Google Scholar] [CrossRef]
- Jansson, S. The light-harvesting chlorophyll ab-binding proteins. Biochim. Biophys. Acta 1994, 1184, 1–19. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functions and Enzymes with Increased Gene Expression | Functions and Enzymes with Reduced Gene Expression |
|---|---|
| Antioxidative activity | DNA Methylation |
| Photosynthetic electron transport chain | DNA replication |
| Molecular Chaperone | Histone |
| Response to Abscisic acid |
| GO Terms | Count | p-Value | Benjamini |
|---|---|---|---|
| Oxidoreductase | 37 | 3.30 × 10−5 | 6.90 × 10−4 |
| Oxidation–reduction process | 39 | 6.20 × 10−4 | 1.80 × 10−2 |
| Iron | 19 | 3.00 × 10−2 | 2.10 × 10−1 |
| Superoxide dismutase, copper/zinc-binding domain | 3 | 1.40 × 10−3 | 5.70 × 10−2 |
| Superoxide dismutase, (Cu/Zn)/chaperones | 3 | 1.40 × 10−3 | 5.70 × 10−2 |
| Removal of superoxide radicals | 3 | 1.60 × 10−2 | 2.50 × 10−1 |
| Copper | 3 | 3.00 × 10−1 | 1.00 × 100 |
| GO Terms | Count | p-Value | Benjamini |
|---|---|---|---|
| Pigment binding | 13 | 3.80 × 10−17 | 1.10 × 10−14 |
| Photosynthesis, light harvesting in photosystem I | 13 | 1.70 × 10−16 | 7.40 × 10−14 |
| Photosynthesis–antenna proteins | 13 | 3.80 × 10−16 | 2.40 × 10−14 |
| Chlorophyll a/b binding_protein domain | 13 | 5.60 × 10−16 | 2.50 × 10−13 |
| Chlorophyll A-B binding_protein, plant | 12 | 1.30 × 10−15 | 2.70 × 10−13 |
| Chlorophyll a/b binding_protein | 13 | 1.80 × 10−15 | 2.70 × 10−13 |
| Chlorophyll | 13 | 7.50 × 10−15 | 1.50 × 10−12 |
| Photosystem I | 13 | 2.80 × 10−14 | 2.30 × 10−12 |
| Light-harvesting complex | 11 | 5.60 × 10−14 | 2.30 × 10−12 |
| Chromophore | 14 | 8.70 × 10−14 | 8.90 × 10−12 |
| Chlorophyll-binding | 13 | 1.90 × 10−13 | 2.80 × 10−11 |
| Protein–chrophore linkage | 14 | 2.20 × 10−13 | 4.80 × 10−11 |
| Photosystem I | 13 | 3.70 × 10−13 | 2.60 × 10−11 |
| photosynthesis | 18 | 6.40 × 10−13 | 3.30 × 10−11 |
| Plastoglobule | 15 | 3.40 × 10−12 | 9.00 × 10−11 |
| Photosystem II | 12 | 7.80 × 10−13 | 1.60 × 10−10 |
| Photosystem II | 13 | 1.40 × 10−11 | 5.60 × 10−10 |
| Photosynthesis | 17 | 1.10 × 10−9 | 1.50 × 10−7 |
| Thylakoid | 20 | 2.60 × 10−8 | 8.80 × 10−7 |
| Thylakoid | 17 | 5.90 × 10−8 | 9.40 × 10−7 |
| Chloroplast thylakoid | 16 | 3.50 × 10−7 | 4.70 × 10−6 |
| Chloroplast thylakoid membrane | 21 | 2.30 × 10−6 | 2.60 × 10−5 |
| Functions and Enzymes with Increased Gene Expression | Functions and Enzymes with Reduced Gene Expression |
|---|---|
| Heat shock protein | Photosynthesis |
| Redox reaction | Glycoprotein |
| Methylation |
| GO Terms | Count | p-Value | Benjamini |
|---|---|---|---|
| Heat Shock Protein 70, conserved site | 7 | 5.60 × 10−8 | 2.80 × 10−5 |
| Heat Shock Protein 70 family | 7 | 2.90 × 10−7 | 4.80 × 10−5 |
| Response to virus | 4 | 2.30 × 10−2 | 3.40 × 10−1 |
| Heat shock protein Hsp90, N-terminal | 4 | 1.20 × 10−4 | 1.20 × 10−2 |
| Heat shock protein Hsp90, conserved site | 4 | 1.20 × 10−4 | 1.20 × 10−2 |
| Heat shock protein Hsp90 | 4 | 1.20 × 10−4 | 1.20 × 10−2 |
| Heat shock protein, HS90/HTPG types | 4 | 1.40 × 10−4 | 2.10 × 10−3 |
| Ribosomal protein S5 domain 2-type fold | 5 | 2.30 × 10−2 | 4.90 × 10−1 |
| Histidine kinase-like ATPase, ATP-binding domain | 4 | 2.50 × 10−2 | 5.00 × 10−1 |
| HATPase c | 3 | 3.40 × 10−2 | 6.90 × 10−1 |
| Plant–pathogen interaction | 8 | 3.60 × 10−2 | 2.50 × 10−1 |
| GO Terms | Count | p-Value | Benjamini |
|---|---|---|---|
| Response to endoplasmic reticulum stress | 7 | 3.9 × 10−7 | 8.7 × 10−5 |
| Redox-active center | 12 | 3.1 × 10−6 | 1.6 × 10−4 |
| Disulfide isomerase | 4 | 7.0 × 10−5 | 8.5 × 10−3 |
| Protein disulfide isomerase activity | 5 | 8.4 × 10−4 | 1.2 × 10−1 |
| Domain: Thioredoxin 1 | 3 | 8.5 × 10−4 | 4.3 × 10−2 |
| Site: Lowers pKa of C-terminal Cys of the second active site | 3 | 8.5 × 10−4 | 4.3 × 10−2 |
| Site: Lowers pKa of C-terminal Cys of the first active site | 3 | 8.5 × 10−4 | 4.3 × 10−2 |
| Domain: Thioredoxin 2 | 3 | 8.5 × 10−4 | 4.3 × 10−2 |
| Protein disulfide isomerase | 3 | 1.4 × 10−3 | 9.3 × 10−2 |
| Thioredoxin, conserved site | 5 | 1.6 × 10−3 | 9.3 × 10−2 |
| Thioredoxin domain | 7 | 2.1 × 10−3 | 1.1 × 10−1 |
| Cell redox homeostasis | 9 | 2.7 × 10−3 | 1.0 × 10−1 |
| Thioredoxin-like fold | 11 | 5.4 × 10−3 | 2.2 × 10−1 |
| GO Terms | Count | p-Value | Benjamini |
|---|---|---|---|
| Light-harvesting complex | 7 | 4.40 × 10−6 | 1.80 × 10−4 |
| Pigment binding | 7 | 5.50 × 10−6 | 2.00 × 10−3 |
| Photosynthesis–antenna proteins | 7 | 6.50 × 10−6 | 4.40 × 10−4 |
| Photosynthesis light-harvesting in photosystem I | 7 | 1.10 × 10−5 | 6.30 × 10−3 |
| Chlorophyll a/b binding_protein domain | 7 | 1.60 × 10−5 | 1.00 × 10−2 |
| Chlorophyll A/B binding_protein | 7 | 2.60 × 10−5 | 8.10 × 10−3 |
| Chlorophyll A/B binding_protein, plant | 6 | 6.80 × 10−5 | 1.40 × 10−2 |
| Chlorophyll | 7 | 7.40 × 10−5 | 4.30 × 10−3 |
| Photosystem I | 7 | 1.10 × 10−4 | 2.90 × 10−3 |
| Chlorophyll-binding | 7 | 1.70 × 10−4 | 3.10 × 10−2 |
| Photosystem II | 7 | 2.90 × 10−4 | 6.10 × 10−3 |
| Plastoglobule | 9 | 3.40 × 10−4 | 6.00 × 10−3 |
| Photosystem I | 7 | 3.70 × 10−4 | 1.40 × 10−2 |
| Protein–chromophore linkage | 7 | 7.60 × 10−4 | 1.00 × 10−1 |
| Chromophore | 7 | 7.90 × 10−4 | 2.60 × 10−2 |
| Photosystem II | 7 | 1.60 × 10−3 | 4.20 × 10−2 |
| Response to red light | 7 | 2.90 × 10−3 | 2.40 × 10−1 |
| Photosynthesis | 9 | 5.30 × 10−3 | 8.50 × 10−2 |
| Photosynthesis | 9 | 2.00 × 10−2 | 5.20 × 10−1 |
| Response to far-red light | 5 | 3.20 × 10−2 | 5.70 × 10−1 |
| GO Terms | Count | p-Value | Benjamini |
|---|---|---|---|
| Signal | 150 | 1.40 × 10−11 | 3.20 × 10−9 |
| Extracellular region | 99 | 3.20 × 10−6 | 2.00 × 10−4 |
| Glycoprotein | 71 | 7.70 × 10−6 | 9.00 × 10−4 |
| Signal peptide | 67 | 1.00 × 10−5 | 5.40 × 10−3 |
| Secreted | 54 | 4.20 × 10−5 | 3.30 × 10−3 |
| Glycosylation site: N-linked(GlcNAc…) | 46 | 2.10 × 10−4 | 5.40 × 10−2 |
| Functions and Enzymes with Increased Gene Expression | Functions and Enzymes with Reduced Gene Expression |
|---|---|
| Biodefense reaction | Glycoprotein |
| Redox reaction | Growth factor |
| Protein phosphorylation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayashi, N.; Yamamoto, K. Variations in Plant Growth Characteristics Due to Oxygen Plasma Irradiation on Leaf and Seed. Agronomy 2022, 12, 259. https://doi.org/10.3390/agronomy12020259
Hayashi N, Yamamoto K. Variations in Plant Growth Characteristics Due to Oxygen Plasma Irradiation on Leaf and Seed. Agronomy. 2022; 12(2):259. https://doi.org/10.3390/agronomy12020259
Chicago/Turabian StyleHayashi, Nobuya, and Kyotaro Yamamoto. 2022. "Variations in Plant Growth Characteristics Due to Oxygen Plasma Irradiation on Leaf and Seed" Agronomy 12, no. 2: 259. https://doi.org/10.3390/agronomy12020259
APA StyleHayashi, N., & Yamamoto, K. (2022). Variations in Plant Growth Characteristics Due to Oxygen Plasma Irradiation on Leaf and Seed. Agronomy, 12(2), 259. https://doi.org/10.3390/agronomy12020259