


Improving Broad Spectrum Blast Resistance by Introduction of the Pita2 Gene: Encoding the NB-ARC Domain of Blast-Resistant Proteins into Upland Rice Breeding Programs


Abstract
1. Introduction
2. Materials and Methods
2.1. Screening for Blast Resistance
2.2. DNA Extraction, PCR Analysis and DNA Amplification
2.3. Sequencing and Analysis Data
2.4. Field Evaluation of Blast Resistance
3. Results
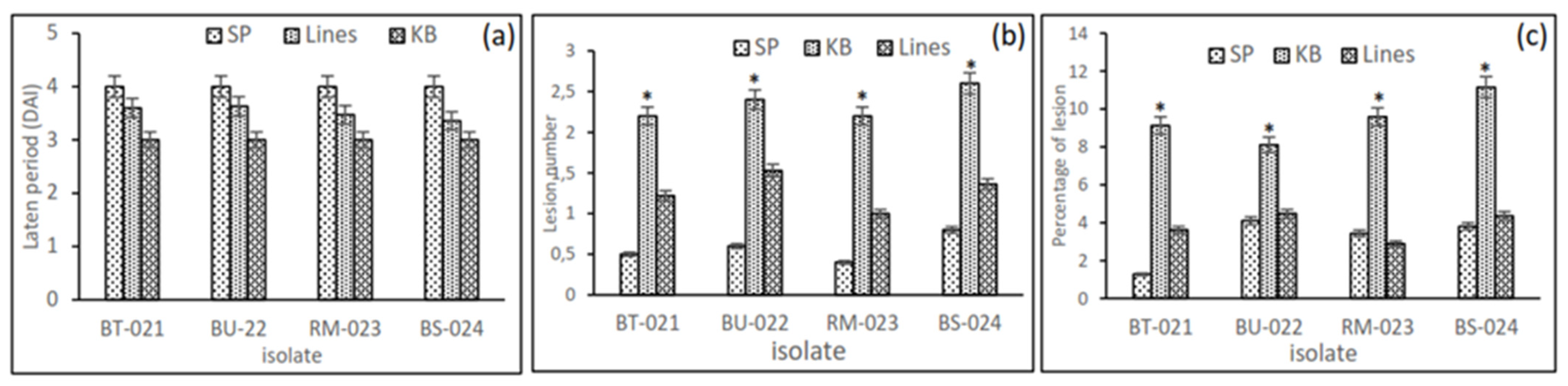
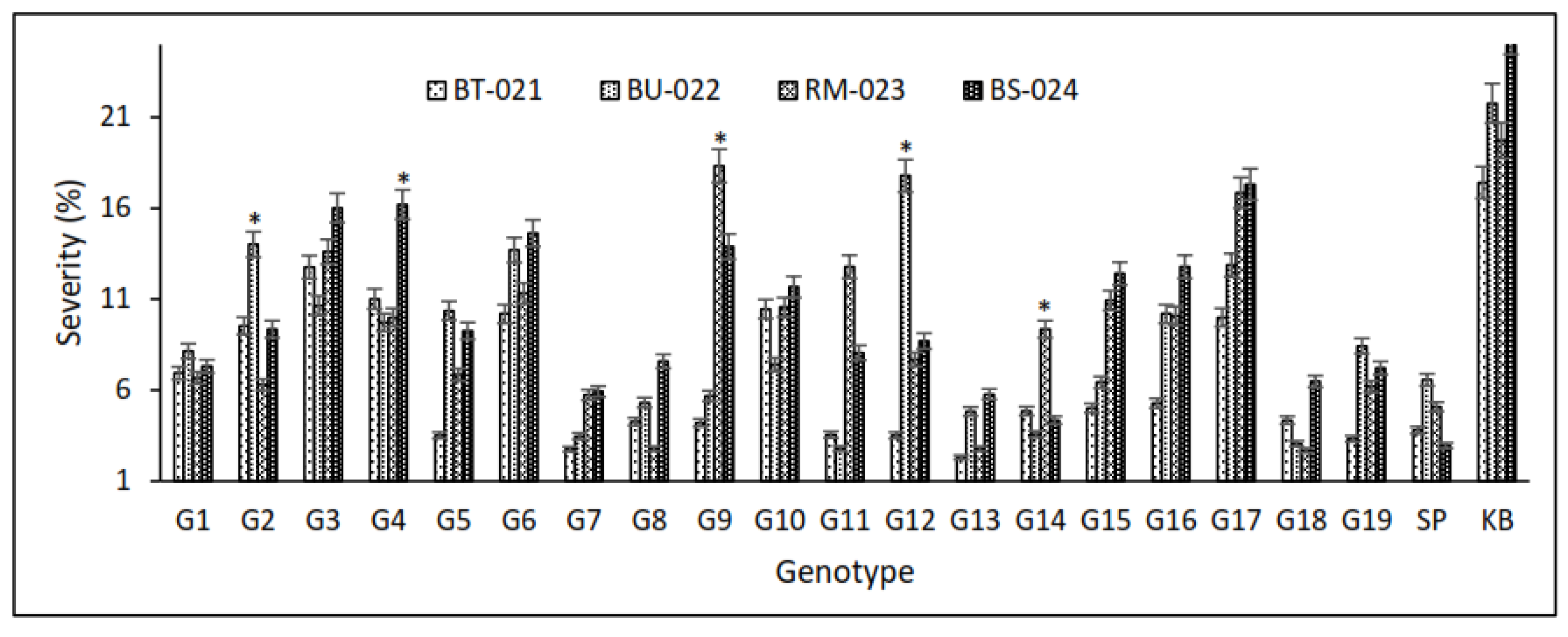
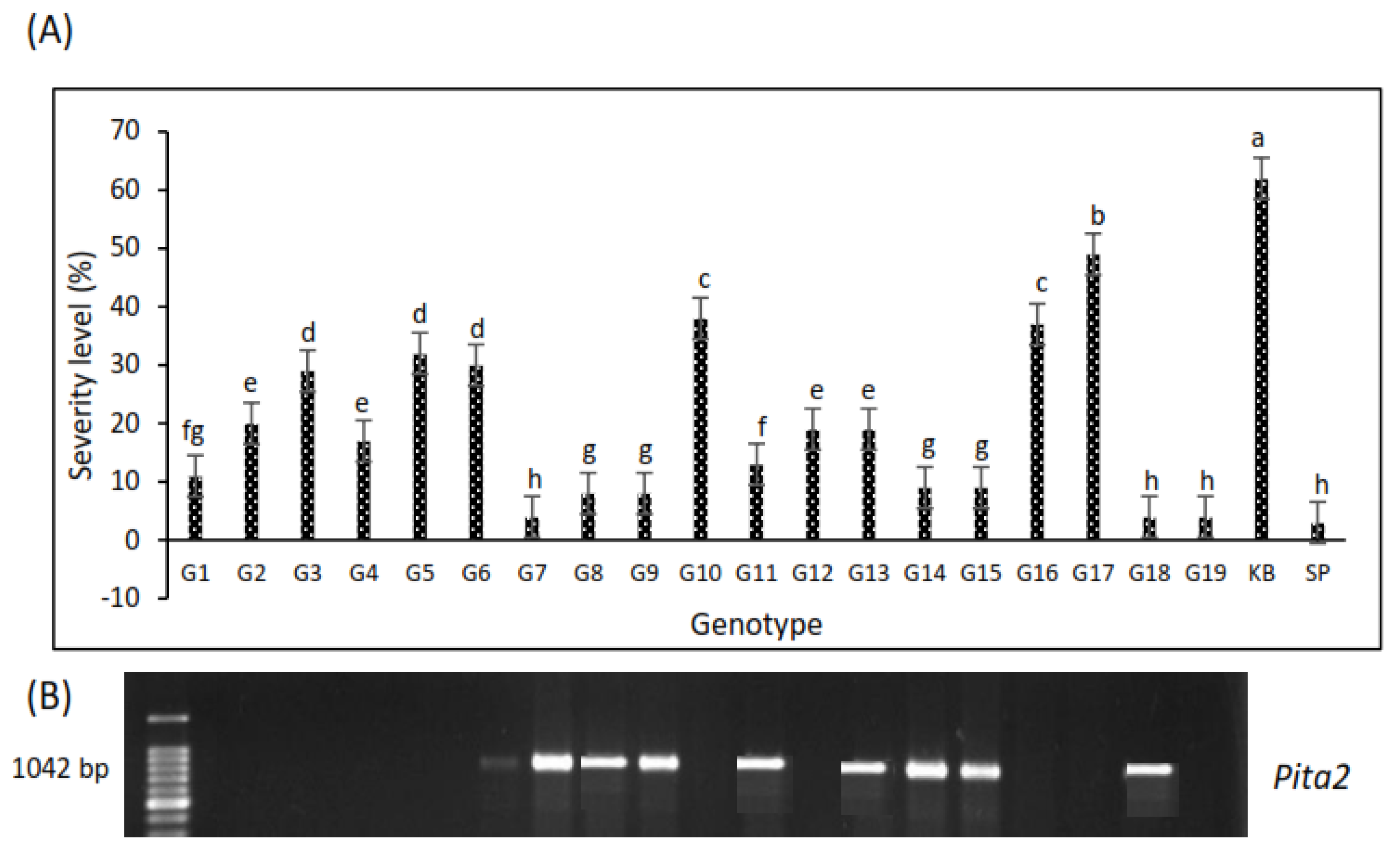
3.1. Screening for Blast Resistance
3.2. Detection of Blast-Resistant Genes
3.3. Sequence Analysis to Determine Genes Encoding the NB-ARC Domain Proteins for Blast Resistance
3.4. Field Evaluation of Blast Resistance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, X.; Lee, S.; Wang, J.; Ma, J.; Bianco, T.; Ji, Y. Current Advances on Genetic Resistance to Rice Blast Disease. In Rice—Germplasm, Genetics and Improvement; InTech: London, UK, 2014. [Google Scholar] [CrossRef]
- Suganda, T.; Yulia, E.; Widiantini, F.; Hersanti, H. Intensitas penyakit blas (Pyricularia Oryzae Cav.) pada padi varietas ciherang di lokasi endemik dan pengaruhnya terhadap kehilangan hasil. Agrikultura 2016, 27, 154–159. [Google Scholar] [CrossRef]
- Nickolas, H.; Jayalekshmy, V.G.; Yamini Varma, C.K.; Vighneswaran, V. Molecular and field level screening for blast resistance gene donors among traditional rice varieties of Kerala. J. Trop. Agric. 2018, 56, 93–98. [Google Scholar]
- Yadav, M.K.; Aravindan, S.; Ngangkham, U.; Raghu, S.; Prabhukarthikeyan, S.R.; Keerthana, U.; Marndi, B.C.; Adak, T.; Munda, S.; Deshmukh, R.; et al. Blast resistance in Indian rice landraces: Genetic dissection by gene specific markers. PLoS ONE 2019, 14, e0211061. [Google Scholar] [CrossRef]
- Wang, G.L.; Valent, B. Durable resistance to rice blast. Science 2017, 355, 906–907. [Google Scholar] [CrossRef]
- Sheoran, N.; Ganesan, P.; Mughal, N.M.; Yadav, I.S.; Kumar, A. Genome assisted molecular typing and pathotyping of rice blast pathogen, Magnaporthe oryzae, reveals a genetically homogenous population with high virulence diversity. Fungal Biol. 2021, 125, 733–747. [Google Scholar] [CrossRef]
- Herawati, R.; Entang Inoriah, S. Genetics Diversity and Agronomic Characters of F3 Lines Selected by Recurrent Selection for Drought Tolerance and Blast Resistance of Bengkulu Local Rice Varieties. Int. J. Adv. Sci. Eng. Inf. Technol. 2017, 7, 922–927. [Google Scholar] [CrossRef][Green Version]
- Herawati, R.; Alnopri, A.; Masdar, M.; Simarmata, M.; Sipriyadi, S.; Sutrawati, M. Identification of drought tolerant markers, DREB2A and BADH2 genes, and yield potential from single-crossing varieties of rice in Bengkulu, Indonesia. Biodiversitas 2021, 22, 785–793. [Google Scholar] [CrossRef]
- Herawati, R.; Lestari, A.P.; Nurmegawati; Ganefianti, D.W.; Romeida, A. Comparative study on the stability and adaptability of different models to develop a high-yield inbred line from landrace rice varieties. Ann. Agric. Sci. 2021, 66, 184–192. [Google Scholar] [CrossRef]
- Khan, M.A.I.; Sen, P.P.; Bhuiyan, R.; Ali, E.K.A.; Chowdhury, A.K.; Latif, M.A. Phenotypic screening and molecular analysis of blast resistance in fragrant rice for marker-assisted selection. Comptes Rendus Biol. 2014, 337, 318–324. [Google Scholar] [CrossRef]
- Deng, Y.; Zhai, K.; Xie, Z.; Yang, D.; Zhu, X.; Liu, J.; Wang, X.; Qin, P.; Yang, Y.; Zhang, G.; et al. Epigenetic regulation of antagonistic receptors confers rice blast resistance with yield balance. Science 2017, 355, 962–965. [Google Scholar] [CrossRef]
- Vasudevan, K.; Gruissem, W.; Bhullar, N.K. Identification of novel alleles of the rice blast resistance gene Pi54. Sci. Rep. 2015, 5, 17920. [Google Scholar] [CrossRef] [PubMed]
- Ning, X.; Yunyu, W.; Aihong, L. Strategy for Use of Rice Blast Resistance Genes in Rice Molecular Breeding. Rice Sci. 2020, 27, 263–277. [Google Scholar] [CrossRef]
- Lin, F.; Chen, S.; Que, Z.; Wang, L.; Liu, X.P.Q. The Blas Resistance Gene Pi37 Encodes a Nucleotide Binding Site Leucine-Rich Repeat Protein and is a Member of a Resistance Gene Cluster on Rice Chromosome 1. Genetics 2007, 177, 1871–1880. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.; Liu, G.; Zhou, B.; Bellizzi, M.; Zeng, L.; Dai, L.; Han, B.; Wang, G.-L. The broad-spectrum blast resistance gene Pi9 encodes a nucleotide-binding site-leucine-rich repeat protein and is a member of a multigene family in rice. Genetics 2006, 172, 1901–1914. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Shang, J.; Chen, D.; Lei, C.; Zhou, Y.; Zhai, W.; Liu, G.; Xu, J.; Ling, Z.; Cao, G.; et al. A B-Lectin Receptor Kinase Gene Conferring Rice Blas Resistance. Plant J. 2006, 46, 794–804. [Google Scholar] [CrossRef] [PubMed]
- Bryan, G.T.; Wu, K.S.; Farrall, L.; Jia, Y.; Hershey, H.P.; McAdams, S.A.; Faulk, K.N.; Donaldson, G.K.; Tarchini, R.V.B. A Single Amino Acid Difference Distinguishes Resistant and Susceptible Alleles of the Rice Blas Resistance Gene Pi-ta. Plant Cell 2000, 12, 2033–2045. [Google Scholar] [CrossRef]
- Heuer, S.; Lu, X.; Chin, J.H.; Tanaka, J.P.; Kanamon, H.; Matsumoto, T.; Leon, T.D.; Ulat, V.J.; Ismail, A.M.; Yano, M.; et al. Comparative sequence analyses of the major quantitative trait locus phosphorus uptake 1 (Pup1) reveal a complex genetic structure. Plant Biotech. J. 2009, 7, 456–471. [Google Scholar] [CrossRef]
- Yan, L.; Bai-Yuan, Y.; Yun-Liang, P.; Zhi-Juan, J.; Yu-Xiang, Z.; Han-Lin, W.; Chang-Deng, Y. Molecular Screening of Blast Resistance Genes in Rice Germplasms Resistant to Magnaporthe oryzae. Rice Sci. 2017, 24, 41–47. [Google Scholar] [CrossRef]
- Kiyosawa, S. Inheritance of resistance of the rice variety PI no. 4 to blast. Jpn. J. Breed. 1967, 17, 165–172. [Google Scholar] [CrossRef]
- Rybka, K.; Miyamoto, M.; Ando, I.; Saito, A.; Kawasaki, S. High resolution mapping of the Indica -derived rice blast resistance genes II. Pi-ta2 and Pi-ta and a consideration of their origin. Mol. Plant-Microbe Interact. 1997, 10, 517–524. [Google Scholar] [CrossRef]
- Liu, J.; Wang, X.; Mitchell, T.; Hu, Y.; Liu, X.; Dai, L.; Wang, G.L. Recent progress and understanding of the molecular mechanisms of the rice- Magnaporthe oryzae interaction. Mol. Plant Pathol. 2020, 11, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Xiao, G.; Telebanco-Yanoria, M.J.; Siazon, P.M.; Padilla, J.; Opulencia, R.; Bigirimana, J.; Habarugira, G.; Wu, J.; Li, M.; et al. The broad-spectrum rice blast resistance (R) gene Pita2 encodes a novel R protein unique from Pita. Rice 2020, 13, 19. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Kazmi, A.Z.; Ahmed, Z.; Roychowdhury, G.; Kumari, V.; Kumar, M.; Mukhopadhyay, K. Genome-wide identification and characterization of NB-ARC resistant genes in wheat (Triticum aestivum L.) and their expression during leaf rust infection. Plant Cell Rep. 2017, 36, 1097–1112. [Google Scholar] [CrossRef] [PubMed]
- Urbach, J.M.; Ausubel, F.M. The NBS-LRR architectures of plant R-proteins and metazoan NLRs evolved in independent events. Proc. Natl. Acad. Sci. USA 2017, 114, 1063–1068. [Google Scholar] [CrossRef]
- Meyers, B.C.; Dickerman, A.W.; Michelmore, R.W.; Sivaramakrishnan, S.; Sobral, B.W.; Young, N.D. Plant disease resistance genes encode members of an ancient and diverse protein family within the nucleotide-binding superfamily. Plant J. 1999, 20, 317–332. [Google Scholar] [CrossRef]
- Nimchuk, Z.; Eulgem, T.; Holt, B.F.; Dangl, J.L. Recognition and Response in the Plant Immune System. Annu. Rev. Genet. 2003, 37, 579–609. [Google Scholar] [CrossRef]
- van der Biezen, E.A.; Jones, J.D. The NB-ARC domain: A novel signalling motif shared by plant resistance gene products and regulators of cell death in animals. Curr. Biol. 1998, 8, R225–R227. [Google Scholar] [CrossRef]
- Danot, O.; Marquenet, E.; Vidal-Ingigliardi, D.; Richet, E. Wheel of Life, Wheel of Death: A Mechanistic Insight into Signaling by STAND Proteins. Structure 2009, 17, 172–182. [Google Scholar] [CrossRef]
- Bentham, A.; Burdett, H.; Anderson, P.A.; Williams, S.J.; Kobe, B. Animal NLRs provide structural insights into plant NLR function. Ann. Bot. 2017, 119, 689–702. [Google Scholar] [CrossRef]
- Hu, Z.; Zhou, Q.; Zhang, C.; Fan, S.; Cheng, W.; Zhao, Y.; Shao, F.; Wang, H.-W.; Sui, S.-F.; Chai, J. Structural and biochemical basis for induced self-propagation of NLRC4. Science 2015, 350, 399–404. [Google Scholar] [CrossRef]
- Devanna, N.B.; Vijayan, J.; Sharma, T.R. The blast resistance gene Pi54of cloned from Oryza officinalis interacts with Avr-Pi54 through its novel non-LRR domains. PLoS ONE 2014, 9, e104840. [Google Scholar] [CrossRef] [PubMed]
- International Rice Research Institut. Standard Evaluation System for Rice. Available online: http://www.knowledgebank.irri.org/images/docs/rice-standard-evaluation-system.pdf (accessed on 25 January 2022).
- Sudir, S.; Nasution, A.; Santoso, S.; Nuryanto, B. Penyakit Blas Pyricularia grisea pada Tanaman Padi dan Strategi Pengendaliannya. Iptek Tanam. Pangan. 2014, 9, 85–96. [Google Scholar]
- Xiao, W.M.; Yang, Q.Y.; Sun, D.Y.; Wang, H.; Guo, T.; Liu, Y.Z.; Zhu, X.Y. CZQ. Identification of three major R genes responsible for broad spectrum blast resistance in an indica rice accession. Mol. Breed. 2015, 35, 49. [Google Scholar] [CrossRef]
- Xiao, W.M.; Luo, L.X.; Hui, W.A.N.G.; Tao, G.U.O.; Liu, Y.Z.; Zhou, J.Y.; Zhu, X.Y.; Yang, Q.Y.; Chen, Z.Q. Pyramiding of Pi46 and Pita to improve blast resistance and to evaluate the resistance effect of the two R genes. J. Integr. Agric. 2016, 15, 2290–2298. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, X.; Jia, Y.; Minkenberg, B.; Wheatley, M.; Fan, J.; Jia, M.H.; Famoso, A.; Edwards, J.; Wamishe, Y.; et al. The rice blast resistance gene Ptr encodes an atypical protein required for broad-spectrum disease resistance. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Orasen, G.; Greco, R.; Puja, E.; Pozzi, C.; Stile, M.R. Blast resistance R genes pyramiding in temperate japonica rice. Euphytica 2020, 216, 40. [Google Scholar] [CrossRef]
- Bonman, J.; Khush, G.N.R. Breeding rice for resistance to pests. Annu. Rev. Phytopathol. 1992, 30, 507–528. [Google Scholar] [CrossRef]
- Koide, Y.; Telebanco-Yanoria, M.; Fukuta, Y.; Kobayashi, N. Detection of novel blast resistance genes, Pi58(t) and Pi59(t), in a Myanmar rice landrace based on a standard differential system. Mol. Breed. 2013, 32, 241–252. [Google Scholar] [CrossRef]
- Sharma, T.R.; Rai, A.K.; Gupta, S.K.; Vijayan, J.; Devanna, B.N.; Ray, S. Rice Blast Management through Host-Plant Resistance: Retrospect and Prospects. Agric. Res. 2012, 1, 37–52. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Gururani, M.A.; Venkatesh, J.; Upadhyaya, C.P.; Nookaraju, A.; Pandey, S.K.; Park, S.W. Plant disease resistance genes: Current status and future directions. Physiol. Mol. Plant Pathol. 2012, 78, 51–65. [Google Scholar] [CrossRef]
- Leipe, D.D.; Koonin, E.V.; Aravind, L. STAND, a class of P-loop NTPases including animal and plant regulators of programmed cell death: Multiple, complex domain architectures, unusual phyletic patterns, and evolution by horizontal gene transfer. J. Mol. Biol. 2004, 343, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Takken, F.L.; Albrecht, M.; Tameling, W.I.L. Resistance proteins: Molecular switches of plant defence. Curr. Opin. Plant Biol. 2006, 9, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Rairdan, G.J.; Moffett, P. Distinct domains in the ARC region of the potato resistance protein Rx mediate LRR binding and inhibition of activation. Plant Cell 2006, 18, 2082–2093. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Accession | The Lowest Score | The Highest Score | Mean Score | Reaction |
|---|---|---|---|---|---|
| G1 | BKL1-RS1-1-247-13 | 0 | 2 | 1.4 | R |
| G2 | BKL1-RS1-1-248-14 | 0 | 3 | 1.8 | R |
| G3 | BKL1-RS1-2-249-15 | 0 | 3 | 3.4 | MR |
| G4 | BKL2-RS1-1-251-17 | 0 | 2 | 1.8 | R |
| G5 | BKL3-RS1-1-253-18 | 2 | 3 | 3.4 | MR |
| G6 | BKL4-RS1-1-256-21 | 0 | 3 | 3.2 | MR |
| G7 | BKL4-RS1-2-257-22 | 0 | 1 | 0.4 | R |
| G8 | BKL4-RS1-3-258-23 | 0 | 2 | 1 | R |
| G9 | BKL1 B-1-259-1 | 0 | 1 | 0.8 | R |
| G10 | BKL1 B-2-260-2 | 3 | 5 | 4.6 | MS |
| G11 | BKL1 B-3-261-3 | 0 | 2 | 2.4 | R |
| G12 | BKL2 B-1-262-4 | 0 | 2 | 2.6 | R |
| G13 | BKL2 B-2-263-5 | 0 | 2 | 2.8 | R |
| G14 | BKL2 B-2-264-6 | 0 | 2 | 1.6 | R |
| G15 | BKL3 B-2-266-8 | 0 | 2 | 2.2 | R |
| G16 | BKL3 B-3-267-9 | 2 | 4 | 4.2 | MS |
| G17 | BKL4 B-1-268-10 | 4 | 4 | 5 | MS |
| G18 | BKL4-B2-269-11 | 0 | 1 | 0.6 | R |
| G19 | BKL4 B-3-270-12 | 0 | 1 | 0.6 | R |
| G20 | Kencana Bali | 5 | 6 | 5.4 | MS |
| G21 | Situ Patenggang | 0 | 1 | 0.6 | R |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herawati, R.; Herlinda, S.; Ganefianti, D.W.; Bustamam, H.; Sipriyadi. Improving Broad Spectrum Blast Resistance by Introduction of the Pita2 Gene: Encoding the NB-ARC Domain of Blast-Resistant Proteins into Upland Rice Breeding Programs. Agronomy 2022, 12, 2373. https://doi.org/10.3390/agronomy12102373
Herawati R, Herlinda S, Ganefianti DW, Bustamam H, Sipriyadi. Improving Broad Spectrum Blast Resistance by Introduction of the Pita2 Gene: Encoding the NB-ARC Domain of Blast-Resistant Proteins into Upland Rice Breeding Programs. Agronomy. 2022; 12(10):2373. https://doi.org/10.3390/agronomy12102373
Chicago/Turabian StyleHerawati, Reny, Siti Herlinda, Dwi Wahyuni Ganefianti, Hendri Bustamam, and Sipriyadi. 2022. "Improving Broad Spectrum Blast Resistance by Introduction of the Pita2 Gene: Encoding the NB-ARC Domain of Blast-Resistant Proteins into Upland Rice Breeding Programs" Agronomy 12, no. 10: 2373. https://doi.org/10.3390/agronomy12102373
APA StyleHerawati, R., Herlinda, S., Ganefianti, D. W., Bustamam, H., & Sipriyadi. (2022). Improving Broad Spectrum Blast Resistance by Introduction of the Pita2 Gene: Encoding the NB-ARC Domain of Blast-Resistant Proteins into Upland Rice Breeding Programs. Agronomy, 12(10), 2373. https://doi.org/10.3390/agronomy12102373