
Use of Dual-Purpose Winter-Grain Cover Crops as Emergency Forage and for Management of High Soil Phosphorous in Manured Fields


Abstract
1. Introduction
2. Materials and Methods
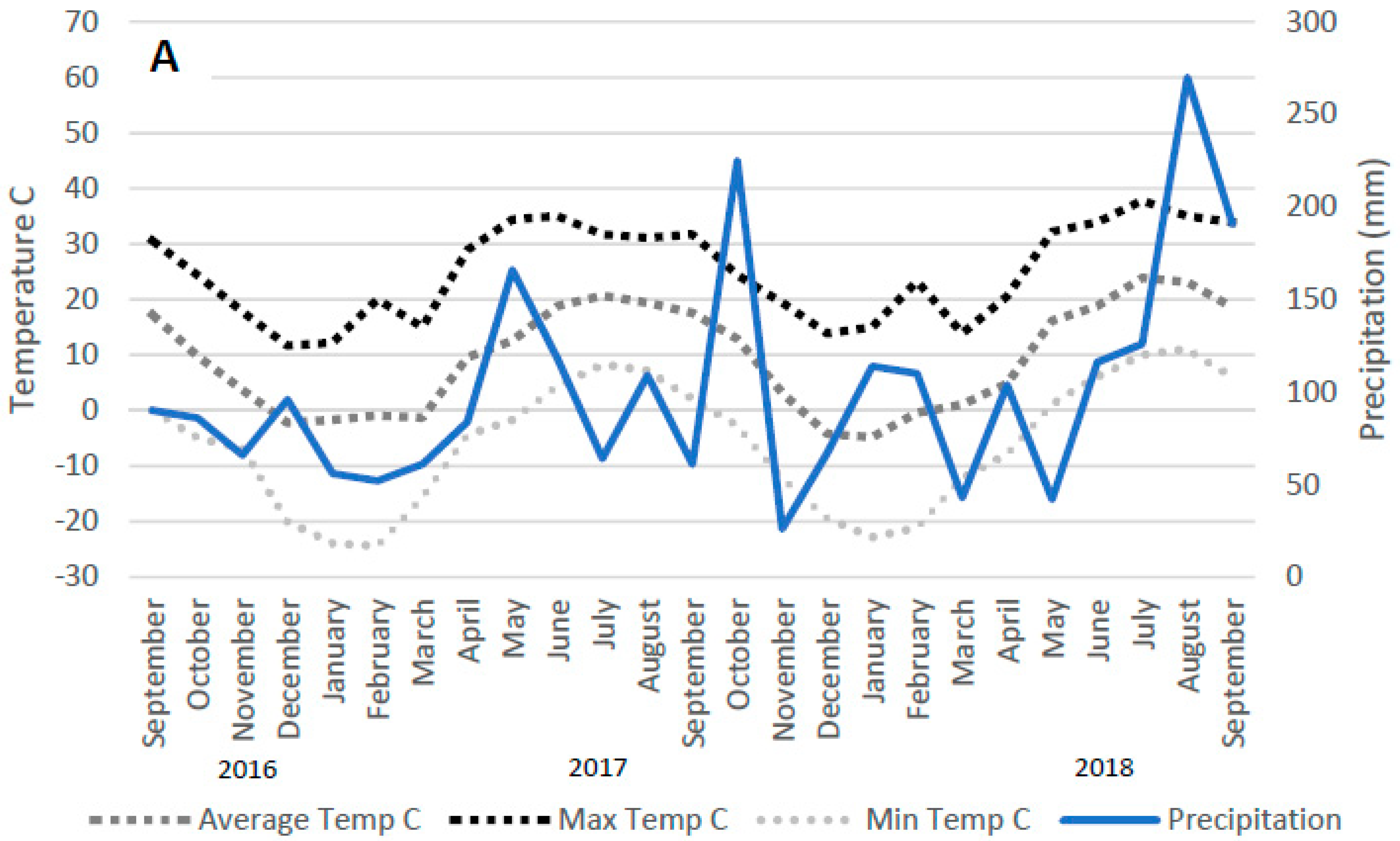
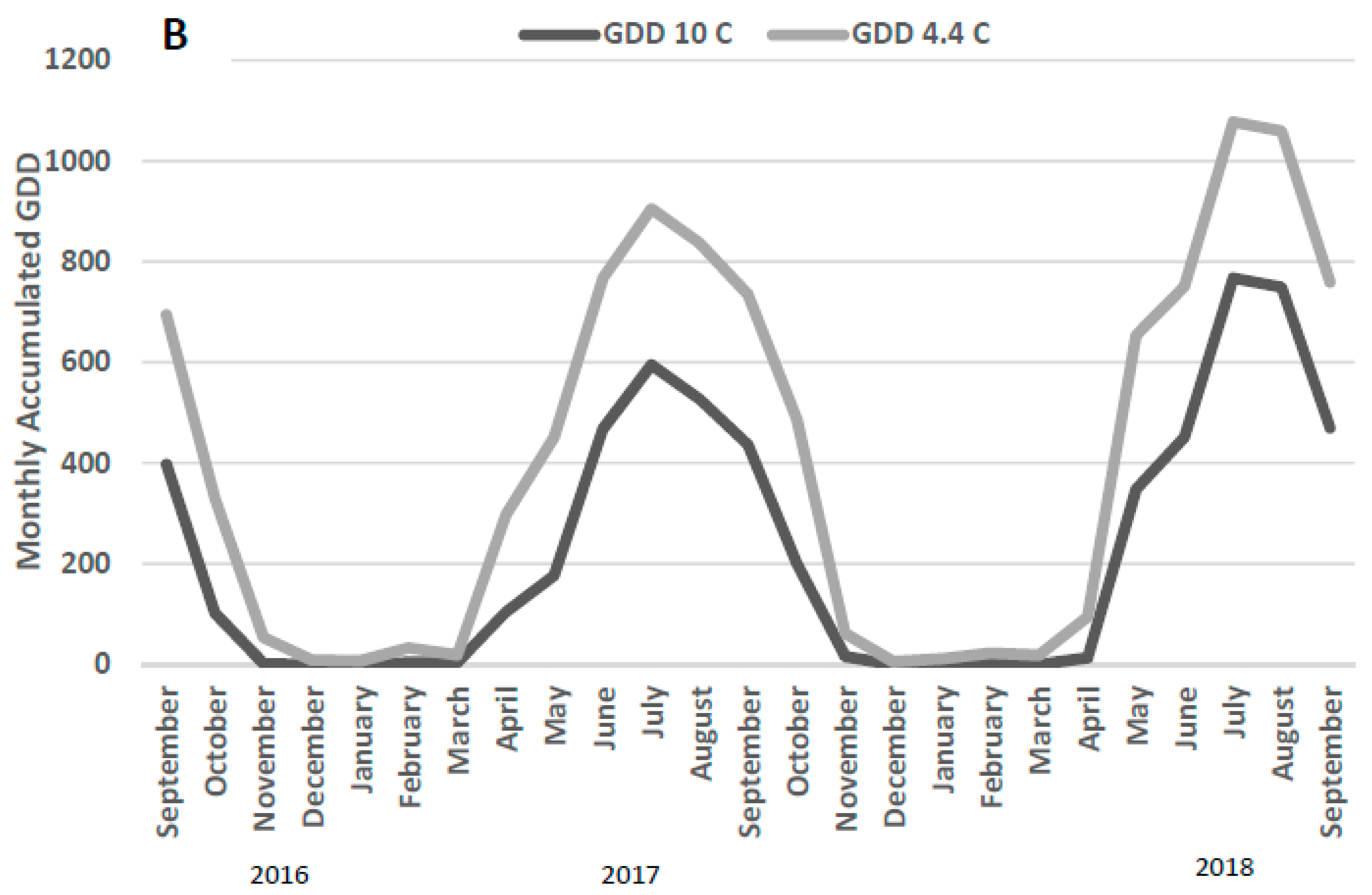
2.1. Experimental Site
2.2. Experimental Design and Management
2.3. Laboratory Analysis
2.4. Statistical Analysis
3. Results
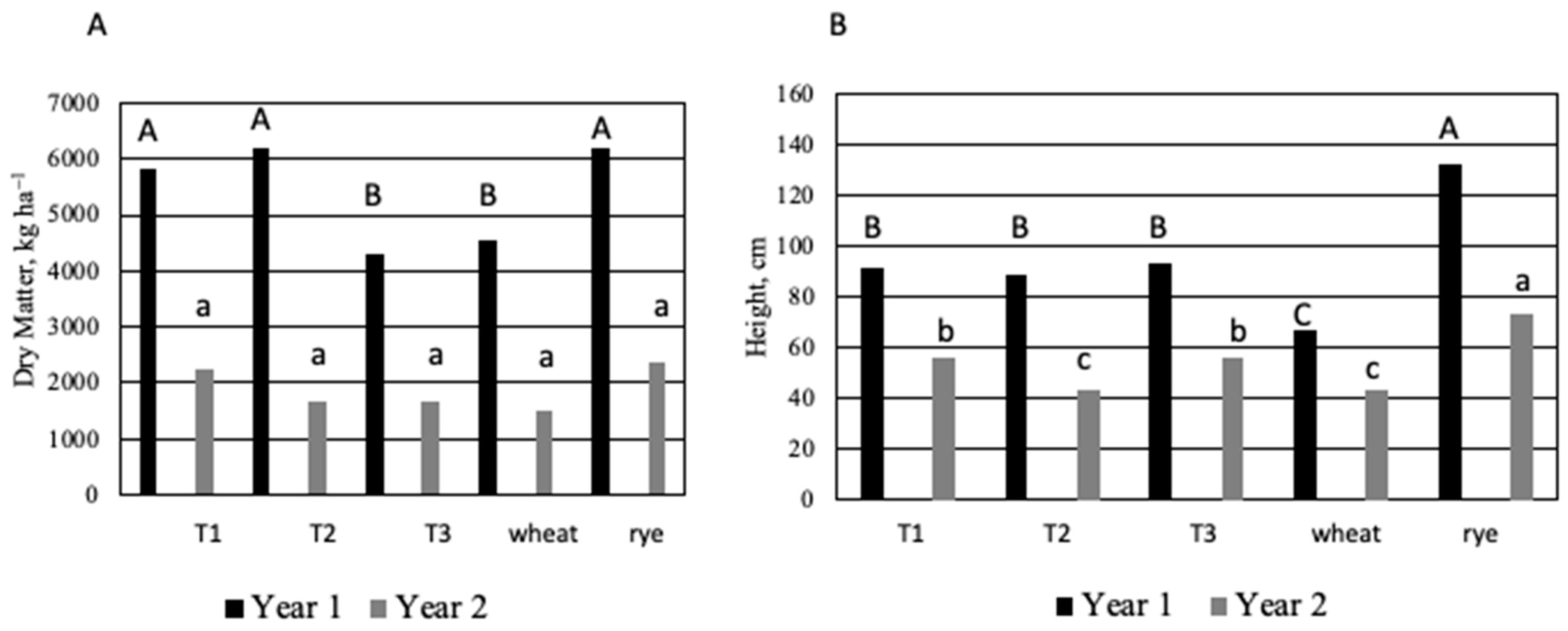
3.1. DPCC Yields, Heights, Moisture Contents, and Water Recoveries
3.2. N and P Recovery
3.3. Soil P and Organic Matter Changes
3.4. DPCC Forage-Quality Characteristics
4. Discussion
4.1. DPCC Yields, Heights, and Moisture Contents
4.2. N and P Recovery
4.3. DPCC Forage Quality
4.4. Final P and Soil-Organic-Matter Levels
5. Conclusions
- -
- DPCCs, in general, are an effective addition to the nutrient management strategies of manured systems. DPCCs should be considered as a tool for the recovery of the N residue in soil, thus preventing N loss and remediating or preventing P accumulation. DPCCs also contribute to on-farm nutrient cycling, and they keep the orthophosphate active in the biological cycle, rather than it becoming fixed by aluminum and iron and lost to the chemical cycle;
- -
- Wheat may not be as quick to mature in the northeast as rye and triticale. This may be desirable for wet fields that are entered later in the spring, or for fields in rotations with other summer crops, such as squash. Conversely, in fields that must be ready for harvest from early to mid-spring, the slow maturity of wheat is likely to result in low DPCC yields and a limited nutrient-scavenging capacity, and thus, both ecosystem services and the economic value of DPCCs are compromised;
- -
- Varieties bred for use as forage should be used. Many varieties have been bred for dual-purpose use as a grain crop, but these varieties appear to perform well as the sole forage crop in corn-silage rotations;
- -
- Winter annual rye is already a popular cover crop, and it offers excellent yields, quality, and nitrogen capture. This study demonstrates that rye is a superior crop for dual-purpose use. With small changes to management practices, such as the planting date or seeding method, many growers can readily begin using rye cover crops for forage.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Harrigan, T.M.; Bickert, W.G.; Rotz, C.A. Simulation of Dairy Manure Management and Cropping Systems. Appl. Eng. Agric. 1996, 12, 563–574. [Google Scholar] [CrossRef]
- Eghball, B.; Power, J.F. Phosphorus- and Nitrogen-Based Manure and Compost Applications Corn Production and Soil Phosphorus. Soil Sci. Soc. Am. J. 1999, 63, 895–901. [Google Scholar] [CrossRef]
- Wironen, M.B.; Bennett, E.M.; Erickson, J.D. Phosphorus flows and legacy accumulation in an animal-dominated agricultural region from 1925 to 2012. Glob. Environ. Chang. 2018, 50, 88–99. [Google Scholar] [CrossRef]
- Bennett, E.M.; Carpenter, S.R.; Caraco, N.F. Human Impact on Erodable Phosphorus and Eutrophication: A Global Perspective: Increasing accumulation of phosphorus in soil threatens rivers, lakes, and coastal oceans with eutrophication. Bioscience 2001, 51, 227–234. [Google Scholar] [CrossRef]
- Daniel, T.C.; Sharpley, A.N.; Lemunyon, J.L. Agricultural Phosphorus and Eutrophication: A Symposium Overview. J. Environ. Qual. 1998, 27, 251–257. [Google Scholar]
- Hammond, L.L.; Chien, S.H.; Roy, A.H.; Mokwunye, A.U. Solubility and agronomic effectiveness of partially acidulated phosphate rocks as influenced by their iron and aluminium oxide content. Nutr. Cycl. Agroecosystems 1989, 19, 93–98. [Google Scholar] [CrossRef]
- Cordell, D. Global phosphorus scarcity: A food secure future? J. Nutr. Intermed. Metab. 2017, 8, 61–62. [Google Scholar] [CrossRef]
- Sarvajayakesavalu, S.; Lu, Y.; Withers, P.J.A.; Pavinato, P.S.; Pan, G.; Chareonsudjai, P. Phosphorus recovery: A need for an integrated approach. Ecosyst. Health Sustain. 2018, 4, 48–57. [Google Scholar] [CrossRef]
- Hashemi, M.; Farsad, A.; Sadeghpour, A.; Weis, S.A.; Herbert, S.J. Cover-crop seeding-date influence on fall nitrogen recovery. J. Plant Nutr. Soil Sci. 2013, 176, 69–75. [Google Scholar] [CrossRef]
- Martin, N.P.; Russelle, M.P.; Powell, J.M.; Sniffen, C.J.; Smith, S.I.; Tricarico, J.M.; Grant, R.J. Invited review: Sustainable forage and grain crop production for the US dairy industry. J. Dairy Sci. 2017, 100, 9479–9494. [Google Scholar] [CrossRef]
- Krueger, E.S.; Ochsner, T.E.; Baker, J.M.; Porter, P.M.; Reicosky, D.C. Rye–Corn Silage Double-Cropping Reduces Corn Yield but Improves Environmental Impacts. Agron. J. 2012, 104, 888–896. [Google Scholar] [CrossRef]
- Laughton, C. 2013 Northeast Dairy Farm Summary; Farm Credit East and Yankee Farm Credit: Enfield, CT, USA, 2014. [Google Scholar]
- Laughton, C. 2016 Northeast Dairy Farm Summary; Farm Credit East and Yankee Farm Credit: Enfield, CT, USA, 2017. [Google Scholar]
- Laughton, C. 2018 Northeast Dairy Farm Summary; Farm Credit East: Enfield, CT, USA, 2019. [Google Scholar]
- Laughton, C.; Zwiegibaum, B. 2014 Northeast Dairy Farm Summary; Farm Credit East and Yankee Farm Credit: Enfield, CT, USA, 2015. [Google Scholar]
- Lidback, J.; Laughton, C. 2011 Northeast Dairy Farm Summary; Farm Credit East and Yankee Farm Credit: Enfield, CT, USA, 2012. [Google Scholar]
- Vujić, S.; Krstić, D.; Mačkić, K.; Čabilovski, R.; Radanović, Z.; Zhan, A.; Ćupina, B. Effect of winter cover crops on water soil storage, total forage production, and quality of silage corn. Eur. J. Agron. 2021, 130, 126366. [Google Scholar] [CrossRef]
- Coale, F.J.; Costa, J.M.; Bollero, G.A.; Schlosnagle, S.P. Small grain winter cover crops for conservation of residual soil nitrogen in the mid-Atlantic Coastal Plain. Am. J. Altern. Agric. 2001, 16, 66. [Google Scholar] [CrossRef]
- Dhakal, D.; Erwin, Z.L.; Nelson, K.A. Grazing cover crops in a no-till corn and soybean rotation. Agron. J. 2022, 114, 1255–1268. [Google Scholar] [CrossRef]
- Komatsuzaki, M.; Wagger, M.G. Nitrogen recovery by cover crops in relation to time of planting and growth termination. J. Soil Water Conserv. 2015, 70, 385–398. [Google Scholar] [CrossRef]
- Schomberg, H.H.; Fisher, D.S.; Reeves, D.W. Grazing winter rye cover crop in a cotton no-till system: Yield and economics. Agron. J. 2014, 106, 1041–1050. [Google Scholar] [CrossRef]
- Tumbalam, P.; Hard, K.; Thelen, K.D. Integrating Winter Annual Cereal Rye or Triticale into a Corn Forage Biofuel Production System. J. Crop Improv. 2016, 30, 526–540. [Google Scholar] [CrossRef]
- Kumssa, T.T.; Anderson, J.D.; Butler, T.J.; Ma, X. Small Grains as Winter Pasture in the Southern Great Plains of the United States. In Grasses and Grassland Aspects; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Melillo, J.M.; Richmond, T.C.; Yohe, G.W. (Eds.) Climate Change Impacts in the United States: The Third National Climate Assessment. U.S.; Global Change Research Program: Washington, DC, USA, 2014; pp. 371–395. [CrossRef]
- Schlegel, R.H. Rye: Genetics, Breeding, and Cultivation; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
- Phillips, H.N.; Heins, B.J.; Delate, K.; Turnbull, R. Biomass Yield and Nutritive Value of Rye (Secale cereale L.) and Wheat (Triticum aestivum L.) Forages While Grazed by Cattle. Crops 2021, 1, 42–53. [Google Scholar] [CrossRef]
- Obour, A.K.; Holman, J.D.; Assefa, Y. Single and multispecies dual-purpose cover crop productivity, nutritive value, and profitability. Agrosystems, Geosci. Environ. 2022, 5, e20275. [Google Scholar] [CrossRef]
- Undersander, D.; Combs, D.; Shaver, R.; Hoffman, P. Milk 2006, University of Wisconsin Alfalfa/Grass Evaluation System. 2006. Available online: https://fyi.extension.wisc.edu/forage/files/2016/09/milk2006-alfalfa-grass-final.xls (accessed on 29 August 2022).
- Glaze-Corcoran, S. Developing Alternative Forage Production Strategies for Environmental and Economic Resiliency on New England Dairy Farms. Ph.D. Thesis, University of Massachusetts, Amherst, MA, USA, 2019. [Google Scholar]
- Baenziger, P.S.; Janninks, J.-L.; Gibson, L.R. Registration of ‘NE426GT’ Winter Triticale. Crop Sci. 2005, 45, 796. [Google Scholar] [CrossRef]
- Ayalew, H.; Kumssa, T.T.; Butler, T.; Ma, X.-F. Triticale Improvement for Forage and Cover Crop Uses in the Southern Great Plains of the United States. Front. Plant Sci. 2018, 9, 1130. [Google Scholar] [CrossRef]
- Coblentz, W.K.; Akins, M.S.; Cavadini, J.S. Fall dry matter yield and nutritive value of winter rye, wheat, and triticale cultivars in Wisconsin. Crop. Forage Turfgrass Manag. 2020, 6, e20075. [Google Scholar] [CrossRef]
- Kim, K.S.; Anderson, J.D.; Webb, S.L.; Newell, M.A.; Butler, T.J. Variation in winter forage production of four small grain species-oat, rye, triticale, and wheat. Pak. J. Bot. 2017, 49, 553–559. [Google Scholar]
- Bruckner, P.L.; Raymer, P.L. Factors Influencing Species and Cultivar Choice of Small Grains for Winter Forage. J. Prod. Agric. 1990, 3, 349–355. [Google Scholar] [CrossRef]
- Sheng, Q.; Hunt, L.A. Shoot and root dry weight and soil water in wheat, triticale and rye. Can. J. Plant Sci. 1991, 71, 41–49. [Google Scholar] [CrossRef]
- Martinez-Feria, R.A.; Dietzel, R.; Liebman, M.; Helmers, M.J.; Archontoulis, S.V. Rye cover crop effects on maize: A system-level analysis. Field Crop. Res. 2016, 196, 145–159. [Google Scholar] [CrossRef]
- Schroeder, J.W. Haylage and Other Fermented Forages. Quality Forage Series, AS1252. 2013. Available online: http://library.nd.gov/statedocs/NDSUExtensionService/as125220131223.pdf (accessed on 29 August 2022).
- Bennett, J.; Mutti, L.; Rao, P.; Jones, J. Interactive effects of nitrogen and water stresses on biomass accumulation, nitrogen uptake, and seed yield of maize. Field Crop. Res. 1989, 19, 297–311. [Google Scholar] [CrossRef]
- Hawkesford, M.J. An overview of nutrient use efficiency and strategies for crop improvement. In The Molecular and Physiological Basis of Nutrient Use Efficiency in Crops; Hawkesford, M.J., Barraclough, P., Eds.; Wiley-Blackwell: Chichester, UK, 2011; pp. 3–19. [Google Scholar]
- Karpinets, T.V.; Greenwood, D.J.; Sams, C.E.; Ammons, J.T. RNA: Protein ratio of the unicellular organism as a characteristic of phosphorus and nitrogen stoichiometry and of the cellular requirement of ribosomes for protein synthesis. BMC Biol. 2006, 4, 30. [Google Scholar] [CrossRef]
- Mugwira, L.M.; Elgawhary, S.M.; Allen, A.E. Nitrate Uptake Effectiveness of Different Cultivars of Triticale, Wheat, and Rye 1. Agron. J. 1980, 72, 585–588. [Google Scholar]
- Pandey, R.; Singh, B.; Nair, T.V.R. Impact of Arbuscular-Mycorrhizal Fungi on Phosphorus Efficiency of Wheat, Rye, and Triticale. J. Plant Nutr. 2005, 28, 1867–1876. [Google Scholar] [CrossRef]
- Lynch, J.P. Steep, cheap and deep: An ideotype to optimize water and N acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef] [PubMed]
- NOAA National Centers for Environmental Information. Climate at a Glance: Statewide Time Series. 2019. Available online: https://www.ncdc.noaa.gov/cag/ (accessed on 22 August 2019).
- Fick, G.W.; Wilkens, P.W.; Cherney, J.H. Modeling Forage Quality Changes in The Growing Crop; Wiley: New York, NY, USA, 1994; Volume 4, pp. 757–795. [Google Scholar]
- Sadeghpour, A.; Adeyemi, O.; Reed, B.; Fry, J.; Afshar, R.K. Profitability of dual-purpose rye cover crop as influenced by harvesting date. Agron. J. 2021, 114, 627–640. [Google Scholar] [CrossRef]
- USDA Economics, Statistics and Market Information System. Announcement of Advanced Prices and Pricing Factors. 2019. Available online: https://usda.library.cornell.edu/concern/publications/ms35t8620?locale=en (accessed on 11 September 2019).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2016–2017 | 2017–2018 | Pr > F | |
|---|---|---|---|
| Dry matter (kg ha−1) | 5442 a | 1866 b | <0.0001 |
| Height (cm) | 95 a | 54 b | <0.0001 |
| Percent moisture at harvest Water removed (m3 ha−1) | 74 b 16.7 a | 82 a 8.7 b | <0.0001 <0.0001 |
| N concentration (%) | 1.55 b | 1.90 a | <0.0001 |
| P concentration (%) | 0.39 b | 0.47 a | <0.0001 |
| N:P ratio | 4.06 | 4.07 | 0.7649 |
| N removed (kg ha−1) | 83.2 a | 35.3 b | <0.0001 |
| P removed (kg ha−1) | 21.9 a | 8.8 b | <0.0001 |
| Relative feed value | 111 b | 126 a | <0.0001 |
| Crude protein (%) | 11.9 b | 9.7 a | <0.0001 |
| Milk (kg ha−1) | 4402 a | 1563 b | <0.0001 |
| Milk (kg Mg−1) | 1621 b | 1715 a | 0.0044 |
| Dollar value of milk ha−1 of DPCC (USD) | 845 a | 300 b | <0.0001 |
| T1 | T2 | T3 | Wheat | Rye | Pr > F | |
|---|---|---|---|---|---|---|
| Dry matter (kg ha−1) | 4056 a | 3953 a | 2987 b | 2950 b | 4612 a | <0.0001 |
| Height (cm) | 74 b | 66 c | 74 b | 56 d | 106 a | <0.0001 |
| Percent moisture at harvest | 78 ab | 80 a | 77 b | 73 c | 81 a | <0.0001 |
| Water removed (L ha−1) | 14,000 bc | 15,250 b | 9520 c | 7745 c | 18,455 a | <0.0001 |
| Nitrogen concentration (%) | 1.66 b | 1.65 b | 1.71 b | 1.90 a | 1.69 b | 0.0004 |
| Phosphorous concentration (%) | 0.45 a | 0.44 a | 0.44 a | 0.38 b | 0.45 a | <0.0001 |
| Ratio of N:P | 3.68 b | 3.71 b | 3.95 b | 5.2 a | 3.79 b | <0.0001 |
| Nitrogen removed (kg ha−1) | 62.4 b | 57.9 b | 48.4 c | 53.7 b | 77.3 a | 0.0002 |
| Phosphorous removed (kg ha−1) | 17.3 ab | 16.6 b | 12.4 c | 10.2 c | 20.8 a | <0.0001 |
| Relative feed value | 112 b | 116 b | 120 b | 132 a | 112 b | <0.0001 |
| Crude protein (%) | 8.6 b | 8.8 b | 9.5 b | 11.0 a | 10.4 b | 0.0004 |
| Milk (kg ha−1) | 3080 a | 2992 a | 2470 b | 2718 b | 3713 a | 0.0216 |
| Milk (kg Mg−1) | 1543 b | 1636 ab | 1729 a | 1759 a | 1677 ab | 0.0005 |
| Dollar value of milk ha−1 of DPCC (USD) | 590 ab | 576 ab | 326 b | 474 b | 714 a | 0.0216 |
| Phosphorus (mg/g) | Soil Organic Matter (%) | |
|---|---|---|
| Baseline | 0.031 | 3.9 |
| T1 | 0.029 | 3.1 |
| T2 | 0.032 | 4.2 |
| T3 | 0.031 | 3.9 |
| Wheat | 0.030 | 3.9 |
| Rye | 0.029 | 4.7 |
| Average of all DPCCs | 0.030 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glaze-Corcoran, S.; Smychkovich, A.; Hashemi, M. Use of Dual-Purpose Winter-Grain Cover Crops as Emergency Forage and for Management of High Soil Phosphorous in Manured Fields. Agronomy 2022, 12, 2334. https://doi.org/10.3390/agronomy12102334
Glaze-Corcoran S, Smychkovich A, Hashemi M. Use of Dual-Purpose Winter-Grain Cover Crops as Emergency Forage and for Management of High Soil Phosphorous in Manured Fields. Agronomy. 2022; 12(10):2334. https://doi.org/10.3390/agronomy12102334
Chicago/Turabian StyleGlaze-Corcoran, Samantha, Alexandra Smychkovich, and Masoud Hashemi. 2022. "Use of Dual-Purpose Winter-Grain Cover Crops as Emergency Forage and for Management of High Soil Phosphorous in Manured Fields" Agronomy 12, no. 10: 2334. https://doi.org/10.3390/agronomy12102334
APA StyleGlaze-Corcoran, S., Smychkovich, A., & Hashemi, M. (2022). Use of Dual-Purpose Winter-Grain Cover Crops as Emergency Forage and for Management of High Soil Phosphorous in Manured Fields. Agronomy, 12(10), 2334. https://doi.org/10.3390/agronomy12102334