
Efficiency of Methodologies Used in the Evaluation of the Weed Seed Bank under Mediterranean Conditions


Abstract
1. Introduction
2. Materials and Methods
2.1. Soil Seed Bank Sampling, Experimental Design and Methods of Quantification
2.2. Seed Germination Test (SG)
2.3. Seed Physical Extraction Test (SPE)
2.4. Statistical Analysis
3. Results
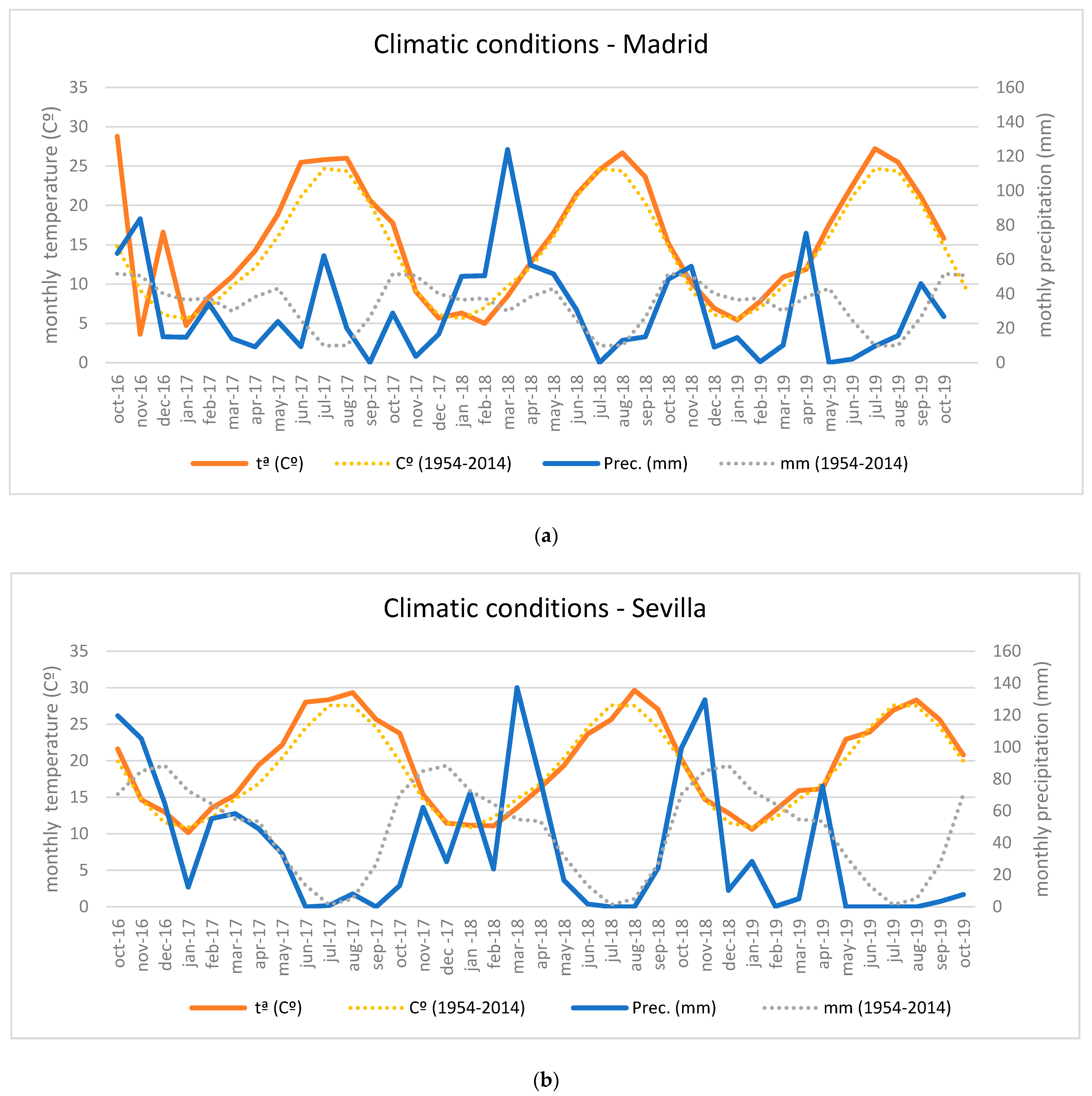
3.1. Climatic Conditions during the Study Period
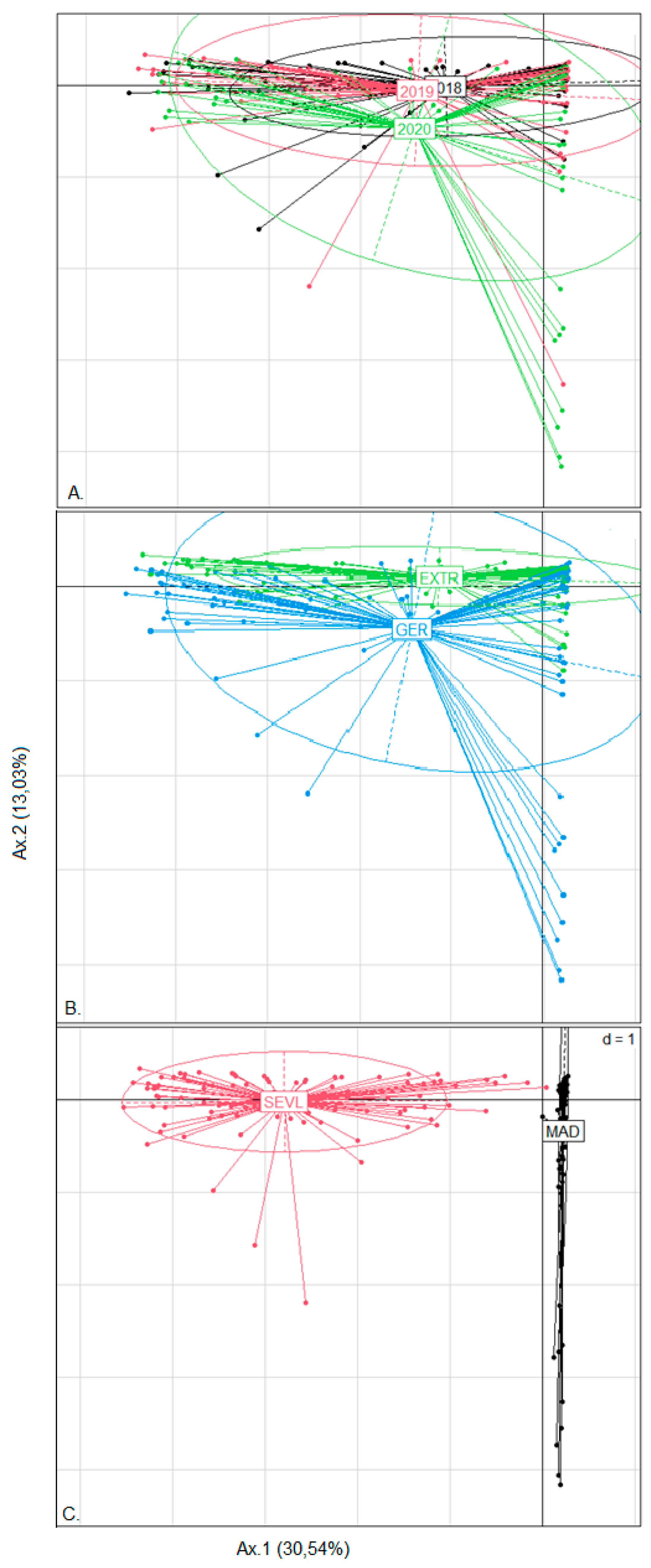
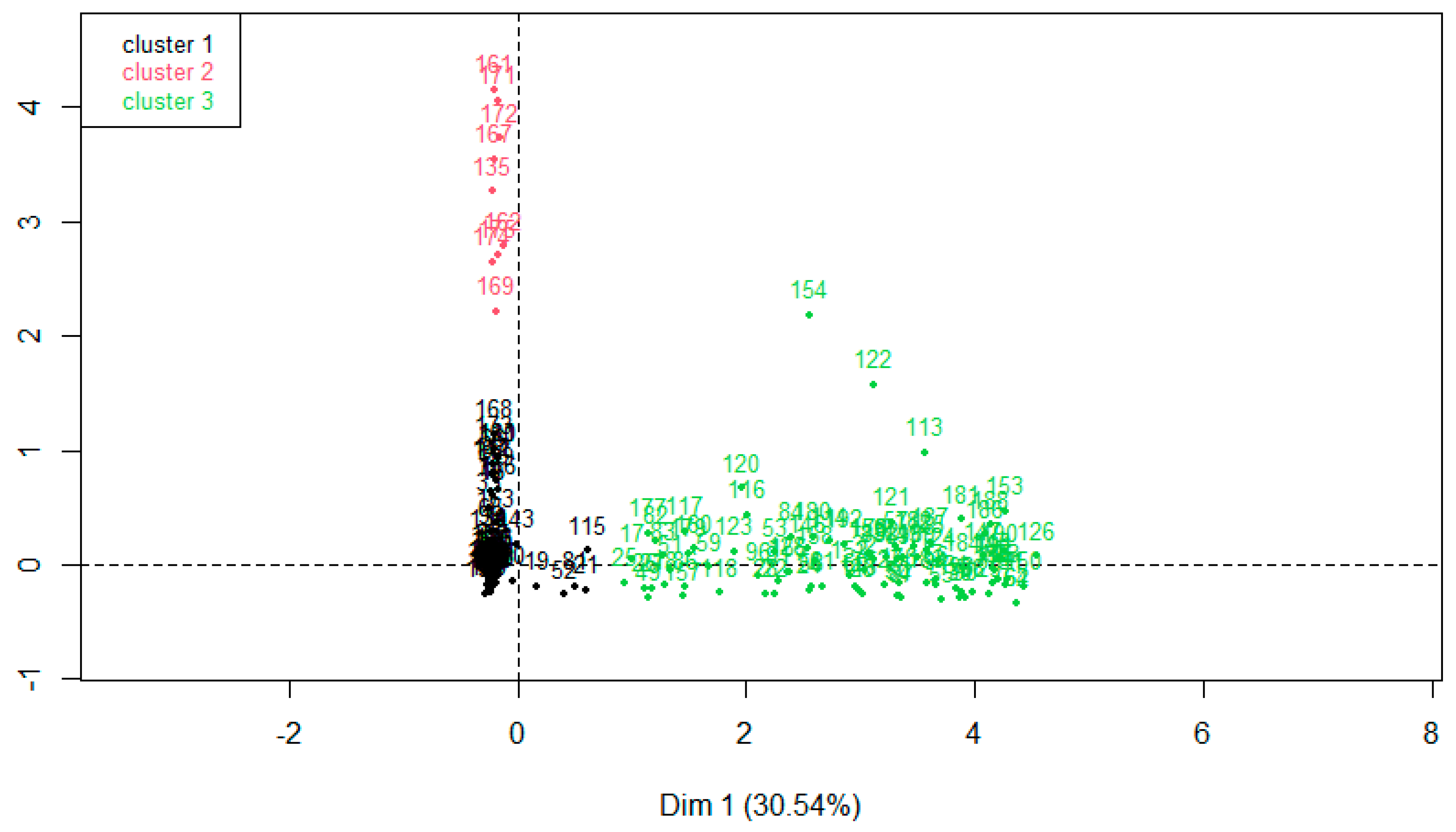
3.2. Correspondence Analysis (CA) Hierarchical Clustering on Principal Components (HCPC) of Weed Seed Banks
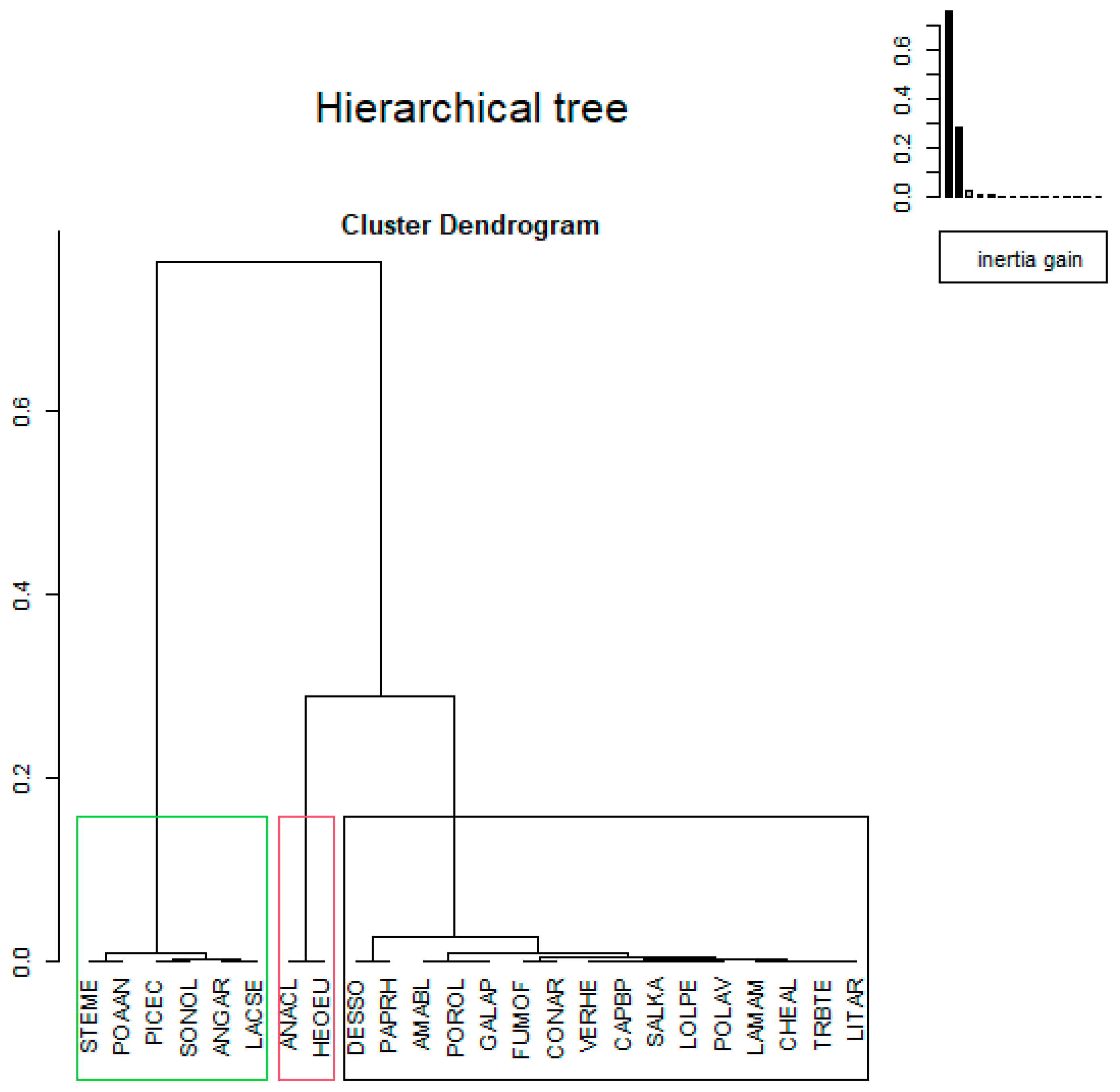
3.3. The HCPC from Weed Seed Bank in M Experiment
3.4. The HCPC from Weed Seed Bank in S Experiment
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kigel, J. Seed Development and Germination; CRC Press: New York, NY, USA, 1995; ISBN 978-0-8247-9229-9. [Google Scholar]
- Taiwo, D.M.; Oyelowo, O.J.; Ogedengbe, T.C.; Woghiren, A.I. The Role of Soil Seed Bank in Forest Regeneration. Asian J. Res. Agric. For. 2018, 1, 1–10. [Google Scholar] [CrossRef]
- Feledyn-Szewczyk, B.; Smagacz, J.; Kwiatkowski, C.A.; Harasim, E.; Woźniak, A. Weed Flora and Soil Seed Bank Composition as Affected by Tillage System in Three-Year Crop Rotation. Agriculture 2020, 10, 186. [Google Scholar] [CrossRef]
- Hopfensperger, K.N. A Review of Similarity between Seed Bank and Standing Vegetation across Ecosystems. Oikos 2007, 116, 1438–1448. [Google Scholar] [CrossRef]
- Fisher, B.; Turner, R.K.; Morling, P. Defining and Classifying Ecosystem Services for Decision Making. Ecol. Econ. 2009, 68, 643–653. [Google Scholar] [CrossRef]
- Harper, J.L. Population Biology of Plants; Academic Press: London, UK, 1977; ISBN 0-12-325850-2. [Google Scholar]
- Laskurain, N.A.; Escudero, A.; Olano, J.M.; Loidi, J. Seedling Dynamics of Shrubs in a Fully Closed Temperate Forest: Greater than Expected. Ecography 2004, 27, 650–658. [Google Scholar] [CrossRef]
- Olano, J.M.; Caballero, I.; Laskurain, N.A.; Loidi, J.; Escudero, A. Seed Bank Spatial Pattern in a Temperate Secondary Forest. J. Veg. Sci. 2002, 13, 775–784. [Google Scholar] [CrossRef]
- Radosevich, S.R.; Holt, J.S.; Ghersa, C. Weed Ecology: Implications for Management; John Wiley & Sons: New York, NY, USA, 1997; ISBN 978-0-471-11606-6. [Google Scholar]
- A Hierarchical Consideration of Causes and Mechanisms of Succession, SpringerLink. Available online: https://link.springer.com/chapter/10.1007/978-94-009-4061-1_10 (accessed on 19 November 2021).
- Baraibar, B.; Canadell, C.; Torra, J.; Royo-Esnal, A.; Recasens, J. Weed Seed Fate during Summer Fallow: The Importance of Seed Predation and Seed Burial. Weed Sci. 2017, 65, 515–524. [Google Scholar] [CrossRef]
- Hyatt, L.A.; Casper, B.B. Seed Bank Formation during Early Secondary Succession in a Temperate Deciduous Forest. J. Ecol. 2000, 88, 516–527. [Google Scholar] [CrossRef]
- Menalled, F.D.; Gross, K.L.; Hammond, M. Weed Aboveground and Seedbank Community Responses to Agricultural Management Systems. Ecol. Appl. 2001, 11, 1586–1601. [Google Scholar] [CrossRef]
- Murphy, S.D.; Clements, D.R.; Belaoussoff, S.; Kevan, P.G.; Swanton, C.J. Promotion of Weed Species Diversity and Reduction of Weed Seedbanks with Conservation Tillage and Crop Rotation. Weed Sci. 2006, 54, 69–77. [Google Scholar] [CrossRef]
- Albrecht, H.; Auerswald, K. Seed Traits in Arable Weed Seed Banks and Their Relationship to Land-Use Changes. Basic Appl. Ecol. 2009, 10, 516–524. [Google Scholar] [CrossRef]
- Pallavicini, Y.; Hernandez Plaza, E.; Bastida, F.; Izquierdo, J.; Gallart, M.; Gonzalez-Andujar, J.L. Weed Seed Bank Diversity in Dryland Cereal Fields: Does It Differ Along the Field and Between Fields with Different Landscape Structure? Agronomy 2020, 10, 575. [Google Scholar] [CrossRef]
- Pritchard, H.W. Seeds. Ecology, biogeography, and evolution of dormancy and germination. Ann. Bot. 2000, 86, 705–707. [Google Scholar] [CrossRef]
- Murdoch, A.J. Longevity, Viability and Dormancy. Seeds Ecol. Regen. Plant Communities 1992, 2, 193–229. [Google Scholar]
- De Souza Maia, M.; Maia, F.C.; Pérez, M.A. Bancos de semillas en el suelo. Agriscientia 2006, 23, 33–44. [Google Scholar]
- McCloskey, M.; Firbank, L.G.; Watkinson, A.R.; Webb, D.J. The Dynamics of Experimental Arable Weed Communities under Different Management Practices. J. Veg. Sci. 1996, 7, 799–808. [Google Scholar] [CrossRef]
- Effects of Crop and Weed Management on Density and Vertical Distribution of Weed Seeds in Soil Agronomy Journal. Available online: https://acsess.onlinelibrary.wiley.com/doi/abs/10.2134/agronj1998.00021962009000060013xa (accessed on 19 November 2021).
- Dessaint, F.; Barralis, G.; Beuret, E.; Caixinhas, M.L.; Post, B.J.; Zanin, G. Etude coopérative EWRS: La détermination du potentiel semencier. Weed Res. 1990, 30, 421–429. [Google Scholar] [CrossRef]
- Bàrberi, P.; Lo Cascio, B. Long-Term Tillage and Crop Rotation Effects on Weed Seedbank Size and Composition. Weed Res. 2001, 41, 325–340. [Google Scholar] [CrossRef]
- Rao, N.; Dong, L.; Li, J.; Zhang, H. Influence of Environmental Factors on Seed Germination and Seedling Emergence of American Sloughgrass (Beckmannia Syzigachne). Weed Sci. 2008, 56, 529–533. [Google Scholar] [CrossRef]
- Lal, B.; Gautam, P.; Raja, R.; Tripathi, R.; Shahid, M.; Mohanty, S.; Panda, B.B.; Bhattacharyya, P.; Nayak, A.K. Weed Seed Bank Diversity and Community Shift in a Four-Decade-Old Fertilization Experiment in Rice–Rice System. Ecol. Eng. 2016, 86, 135–145. [Google Scholar] [CrossRef]
- Buhler, D.D.; Stoltenberg, D.E.; Becker, R.L.; Gunsolus, J.L. Perennial Weed Populations After 14 Years of Variable Tillage and Cropping Practices. Weed Sci. 1994, 42, 205–209. [Google Scholar] [CrossRef]
- Cardina, J.; Regnier, E.; Harrison, K. Long-Term Tillage Effects on Seed Banks in Three Ohio Soils. Weed Sci. 1991, 39, 186–194. [Google Scholar] [CrossRef]
- Ball, D.A. Weed Seedbank Response to Tillage, Herbicides, and Crop Rotation Sequence. Weed Sci. 1992, 40, 654–659. [Google Scholar] [CrossRef]
- Auškalnienė, O.; Auškalnis, A. The Influence of Tillage System on Diversities of Soil Weed Seed Bank. Agron. Res. 2009, 7, 156–161. [Google Scholar]
- Skuodienė, R. Assessment of the Weed Incidence and Weed Seed Bank of Crops under Different Pedological Traits. Appl. Ecol. Environ. Res. 2018, 16, 1131–1142. [Google Scholar] [CrossRef]
- Santín-Montanyá, M.I.; Martín-Lammerding, D.; Zambrana, E.; Tenorio, J.L. Management of Weed Emergence and Weed Seed Bank in Response to Different Tillage, Cropping Systems and Selected Soil Properties. Soil Tillage Res. 2016, 161, 38–46. [Google Scholar] [CrossRef]
- Auffret, A.G.; Cousins, S.A.O. Past and Present Management Influences the Seed Bank and Seed Rain in a Rural Landscape Mosaic: Seed Bank and Seed Rain in the Rural Landscape. J. Appl. Ecol. 2011, 48, 1278–1285. [Google Scholar] [CrossRef]
- Forcella, F.; Fao, R.; Webster, T.; Cardina, J.; Labrada, R. Protocolos Para La Determinacion de Bancos de Semillas de Malezas En Los Agrosistemas; FAO: Rome, Italy, 2004. [Google Scholar]
- Recasens, J.; Riba, F.; Izquierdo, J.; Taberner, A.L. analisis del banc de llavors de males herbes de sòls agrícoles. Aspectes metodològics. Ilerda. Ciències 1991, 49, 83–102. [Google Scholar]
- Jastrzebska, M.; Jastrzebski, W.P.; Holdynski, C.; Kostrzewska, M.K. Weed Species Diversity in Organic and Integrated Farming Systems. Acta Agrobot. 2013, 66, 113–124. [Google Scholar] [CrossRef][Green Version]
- Gross, K.L. A Comparison of Methods for Estimating Seed Numbers in the Soil. J. Ecol. 1990, 78, 1079–1093. [Google Scholar] [CrossRef]
- CardiNa, J.; Sparrow, D.H.; McCoy, E.L. Spatial Relationships between Seedbank and Seedling Populations of Common Lambsquarters (Chenopodium album) and Annual Grasses. Weed Sci. 1996, 44, 298–308. [Google Scholar] [CrossRef]
- Malone, C.R. A Rapid Method for Enumeration of Viable Seeds in Soil. Weeds 1967, 15, 381–382. [Google Scholar] [CrossRef]
- Tsuyuzaki, S. Rapid Seed Extraction from Soils by a Flotation Method. Weed Res. 1994, 34, 433–436. [Google Scholar] [CrossRef]
- Gonzalez, S.; Ghermandi, L. Comparison of Methods to Estimate Soil Seed Banks: The Role of Seed Size and Mass. Community Ecol. 2012, 13, 238–242. [Google Scholar] [CrossRef]
- Mesgaran, M.B.; Mashhadi, H.R.; Zand, E.; Alizadeh, H.M. Comparison of Three Methodologies for Efficient Seed Extraction in Studies of Soil Weed Seedbanks. Weed Res. 2007, 47, 472–478. [Google Scholar] [CrossRef]
- Auškalnienė, O.; Kadžienė, G.; Janušauskaitė, D.; Supronienė, S. Changes in Weed Seed Bank and Flora as Affected by Soil Tillage Systems. Zemdirb. Agric. 2018, 105, 221–226. [Google Scholar] [CrossRef]
- Ball, D.A.; Miller, S.D. A Comparison of Techniques for Estimation of Arable Soil Seedbanks and Their Relationship to Weed Flora. Weed Res. 1989, 29, 365–373. [Google Scholar] [CrossRef]
- Espeland, E.K.; Perkins, L.B.; Leger, E.A. Comparison of Seed Bank Estimation Techniques Using Six Weed Species in Two Soil Types. Rangel. Ecol. Manag. 2010, 63, 243–247. [Google Scholar] [CrossRef]
- Brown, D. Estimating the Composition of a Forest Seed Bank: A Comparison of the Seed Extraction and Seedling Emergence Methods. Can. J. Bot. 1992, 70, 1603–1612. [Google Scholar] [CrossRef]
- Reinhardt, T.; Leon, R.G. Extractable and Germinable Seedbank Methods Provide Different Quantifications of Weed Communities. Weed Sci. 2018, 66, 715–720. [Google Scholar] [CrossRef]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA, 2014.
- Flora Iberica. Plantas Vasculares de la Península Ibérica e Islas Baleares. Real Jardín Botánico CSIC. 2020. [Date Accessed: 2018–2020]. Available online: http://www.floraiberica.org/ (accessed on 28 November 2021).
- Thompson, K.; Bakker, J.P.; Bekker, R.M. The Soil Seed Banks of North West Europe: Methodology, Density and Longevity; University Press: Cambridge, UK, 1997; ISBN 978-0-521-49519-6. [Google Scholar]
- Zanin, G.; Otto, S.; Riello, L.; Borin, M. Ecological Interpretation of Weed Flora Dynamics under Different Tillage Systems. Agric. Ecosyst. Environ. 1997, 66, 177–188. [Google Scholar] [CrossRef]
- Seed Identification Guide. 2018. [Date Accessed: 2018–2020]. Available online: https://www.idseed.org (accessed on 19 November 2021).
- Arable Seed Identification System. The James Hutton Institute. 2020. [Date Accessed: 2018–2020]. Available online: https://Asis.Hutton.Ac.Uk (accessed on 19 November 2021).
- Smutný, V.; Křen, J. Improvement of an Elutriation Method for Estimation of Weed Seedbank in the Soil. Plant Soil Environ. 2011, 48, 271–278. [Google Scholar] [CrossRef]
- Porčová, L.; Smutný, V. The Impact of Under-Vine Management on the Weed Seedbank in the Soil. Acta Univ. Agric. Silvic. Mendel. Brun. 2018, 66, 1165–1170. [Google Scholar] [CrossRef]
- Dray, S.; Dufour, A.-B. The Ade4 Package: Implementing the Duality Diagram for Ecologists. J. Stat. Soft. 2007, 22. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘Vegan’. Community Ecol. Package Version 2013, 2, 1–295. [Google Scholar]
- Husson, F.; Josse, J.; Le, S.; Mazet, J.; Husson, M.F. Package ‘FactoMineR’. R Package 2016, 96, 698. [Google Scholar]
- Dorado, J.; López-Fando, C. The Effect of Tillage System and Use of a Paraplow on Weed Flora in a Semiarid Soil from Central Spain. Weed Res. 2006, 46, 424–431. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Singh, R.G.; Mahajan, G. Ecology and Management of Weeds under Conservation Agriculture: A Review. Crop Prot. 2012, 38, 57–65. [Google Scholar] [CrossRef]
- Trichard, A.; Alignier, A.; Chauvel, B.; Petit, S. Identification of Weed Community Traits Response to Conservation Agriculture. Agric. Ecosyst. Environ. 2013, 179, 179–186. [Google Scholar] [CrossRef]
- Marshall, E.J.P.; Brown, V.K.; Boatman, N.D.; Lutman, P.J.W.; Squire, G.R.; Ward, L.K. The Role of Weeds in Supporting Biological Diversity within Crop Fields*: Weeds and Biodiversity. Weed Res. 2003, 43, 77–89. [Google Scholar] [CrossRef]
- Smith, R.G. Timing of Tillage Is an Important Filter on the Assembly of Weed Communities. Weed Sci. 2006, 54, 705–712. [Google Scholar] [CrossRef]
- Ryan, M.R.; Smith, R.G.; Mirsky, S.B.; Mortensen, D.A.; Seidel, R. Management Filters and Species Traits: Weed Community Assembly in Long-Term Organic and Conventional Systems. Weed Sci. 2010, 58, 265–277. [Google Scholar] [CrossRef]
- Smith, R.G.; Atwood, L.W.; Morris, M.B.; Mortensen, D.A.; Koide, R.T. Evidence for Indirect Effects of Pesticide Seed Treatments on Weed Seed Banks in Maize and Soybean. Agric. Ecosyst. Environ. 2016, 216, 269–273. [Google Scholar] [CrossRef]
- PUL, L.V. Principies of Dispersal Inhigherplants, 3rd ed.; Springer: Berlin, Germany, 1982. [Google Scholar]
- Zohary, M. Plant Life of Palestine: Israel and Jordan, 6th ed.; The Ronald Press: New York, NY, USA, 1962. [Google Scholar]
- Hughes, P.W. Minimal-Risk Seed Heteromorphism: Proportions of Seed Morphs for Optimal Risk-Averse Heteromorphic Strategies. Front. Plant Sci. 2018, 9, 1412. [Google Scholar] [CrossRef]
- Hopkins, D.R.; Parker, V.T. A study of the seed bank of a salt marsh in northern San Francisco Bay. Am. J. Bot. 1984, 71, 348–355. [Google Scholar] [CrossRef]
- Poiani, K.A.; Johnson, W.C. Evaluation of the Emergence Method in Estimating Seed Bank Composition of Prairie Wetlands. Aquat. Bot. 1988, 32, 91–97. [Google Scholar] [CrossRef]
- Ter Heerdt, G.N.J.; Verweij, G.L.; Bekker, R.M.; Bakker, J.P. An Improved Method for Seed-Bank Analysis: Seedling Emergence after Removing the Soil by Sieving. Funct. Ecol. 1996, 144–151. [Google Scholar] [CrossRef]
- Moles, A.T.; Westoby, M. Seed Size and Plant Strategy across the Whole Life Cycle. Oikos 2006, 113, 91–105. [Google Scholar] [CrossRef]
- Saatkamp, A.; Affre, L.; Dutoit, T.; Poschlod, P. The Seed Bank Longevity Index Revisited: Limited Reliability Evident from a Burial Experiment and Database Analyses. Ann. Bot. 2009, 104, 715–724. [Google Scholar] [CrossRef]
- Gunton, R.M.; Petit, S.; Gaba, S. Functional Traits Relating Arable Weed Communities to Crop Characteristics: Traits Relating Weed Communities to Crops. J. Veg. Sci. 2011, 22, 541–550. [Google Scholar] [CrossRef]
- Saatkamp, A.; Affre, L.; Dutoit, T.; Poschlod, P. Germination Traits Explain Soil Seed Persistence across Species: The Case of Mediterranean Annual Plants in Cereal Fields. Ann. Bot. 2011, 107, 415–426. [Google Scholar] [CrossRef]
- Grime, J.P. Plant Strategies and Vegetation Processes, 2nd ed.; John Wiley & Sons: Chichester, UK, 2001. [Google Scholar]
- Gardarin, A.; Dürr, C.; Mannino, M.R.; Busset, H.; Colbach, N. Seed Mortality in the Soil Is Related to Seed Coat Thickness. Seed Sci. Res. 2010, 20, 243–256. [Google Scholar] [CrossRef]
- Dorado, J.; Monte, J.P.D.; López-Fando, C. Weed Seedbank Response to Crop Rotation and Tillage in Semiarid Agroecosystems. Weed Sci. 1999, 47, 67–73. [Google Scholar] [CrossRef]
- Dick, W.A.; Daniel, T.C. Soil Chemical and Biological Properties as Affected by Conservation Tillage: Environmental Implications. In Effects Conservation Tillage on Ground Water Quality: Nitrates and Pesticides; Logan, T.J., Ed.; CRC Press: Chelsea, MI, USA, 1987; pp. 125–147. [Google Scholar]
- Pareja, M.R.; Staniforth, D.W.; Pareja, G.P. Distribution of Weed Seed among Soil Structural Units. Weed Sci. 1985, 33, 182–189. [Google Scholar] [CrossRef]
- Andreasen, C.; Streibig, J.C.; Haas, H. Soil Properties Affecting the Distribution of 37 Weed Species in Danish Fields. Weed Res. 1991, 31, 181–187. [Google Scholar] [CrossRef]
- Shiferaw, W.; Lemenih, M.; Gole, T.W.M. Analysis of Plant Species Diversity and Forest Structure in Arero Dry Afromontane Forest of Borena Zone, South Ethiopia. Trop. Plant Res. 2018, 5, 129–140. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Latin Name | EPPO Code | Life History * | Relative Abundance MAD | Relative Abundance SEV |
|---|---|---|---|---|
| Amaranthus blitoides S. Watson | AMABL | SA | 0.300 | 0.024 |
| Anacyclus clavatus (Desf.) Pers. | ANACL | WA | 0.021 | 0.005 |
| Anagallis arvensis L. | ANGAR | SA | 0.000 | 0.008 |
| Capsella bursa-pastoris (L.) Medicus | CAPBP | WA | 0.001 | 0.000 |
| Chenopodium album L. | CHEAL | SA | 0.045 | 0.012 |
| Convolvulus arvensis L. | CONAR | P | 0.005 | 0.019 |
| Descurainia sophia (L.) Webb ex Prantl in Engler & Prantl | DESSO | WA | 0.008 | 0.000 |
| Fumaria officinalis L. | FUMOF | WA | 0.008 | 0.031 |
| Galium tricornutum L. | GALAP | WA | 0.001 | 0.001 |
| Heliotropium europaeum L. | HEOEU | WA | 0.001 | 0.000 |
| Lactuca serriola L. | LACSE | A | 0.000 | 0.018 |
| Lamium amplexicaule L. | LAMAM | WA | 0.039 | 0.001 |
| Buglossoides arvensis (L.) I.M. Johnst. | LITAR | WA | 0.021 | 0.001 |
| Lolium perenne L. | LOLPE | P | 0.015 | 0.001 |
| Papaver rhoeas L. | PAPRH | WA | 0.022 | 0.002 |
| Picris echioides L. | PICEC | A/B | 0.000 | 0.041 |
| Poa annua L. | POAAN | WA | 0.000 | 0.131 |
| Polygonum aviculare L. | POLAV | P | 0.003 | 0.000 |
| Portulaca oleracea L. | POROL | SA | 0.502 | 0.212 |
| Salsola kali L. | SALKA | SA | 0.002 | 0.000 |
| Sonchus oleraceus L. | SONOL | A | 0.000 | 0.119 |
| Stellaria media (L.) Vill. | STEME | SA | 0.000 | 0.375 |
| Tribulus terrestris L. | TRBTE | WA | 0.002 | 0.000 |
| Veronica hederifolia L. | VERHE | WA | 0.005 | 0.000 |
| M Experiment | ||
|---|---|---|
| Cluster 1 | Cluster 2 | |
| METHOD = EXTR | −5.51 | 5.51 |
| METHOD = GER | 5.51 | −5.51 |
| AMABL | −4.78 | 4.78 |
| ANACL | 1.96 | −1.96 |
| CHEAL | −4.16 | 4.16 |
| POROL | −8.37 | 8.37 |
| SALKA | −2.75 | 2.75 |
| TRBTE | −2.19 | 2.19 |
| S Experiment | ||
|---|---|---|
| Cluster 1 | Cluster 2 | |
| METHOD = EXTR | −1.9 | 1.9 |
| METHOD = GER | 1.9 | −1.9 |
| AMABL | 1.64 | −1.64 |
| CHEAL | −2.42 | 2.42 |
| POAAN | −2.21 | 2.21 |
| STEME | −8.22 | 8.22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gandía, M.L.; Del Monte, J.P.; Santín-Montanyá, M.I. Efficiency of Methodologies Used in the Evaluation of the Weed Seed Bank under Mediterranean Conditions. Agronomy 2022, 12, 138. https://doi.org/10.3390/agronomy12010138
Gandía ML, Del Monte JP, Santín-Montanyá MI. Efficiency of Methodologies Used in the Evaluation of the Weed Seed Bank under Mediterranean Conditions. Agronomy. 2022; 12(1):138. https://doi.org/10.3390/agronomy12010138
Chicago/Turabian StyleGandía, María Luisa, Juan Pablo Del Monte, and María Inés Santín-Montanyá. 2022. "Efficiency of Methodologies Used in the Evaluation of the Weed Seed Bank under Mediterranean Conditions" Agronomy 12, no. 1: 138. https://doi.org/10.3390/agronomy12010138
APA StyleGandía, M. L., Del Monte, J. P., & Santín-Montanyá, M. I. (2022). Efficiency of Methodologies Used in the Evaluation of the Weed Seed Bank under Mediterranean Conditions. Agronomy, 12(1), 138. https://doi.org/10.3390/agronomy12010138