
Estimation of the Genetic Diversity and Population Structure of Thailand’s Rice Landraces Using SNP Markers

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
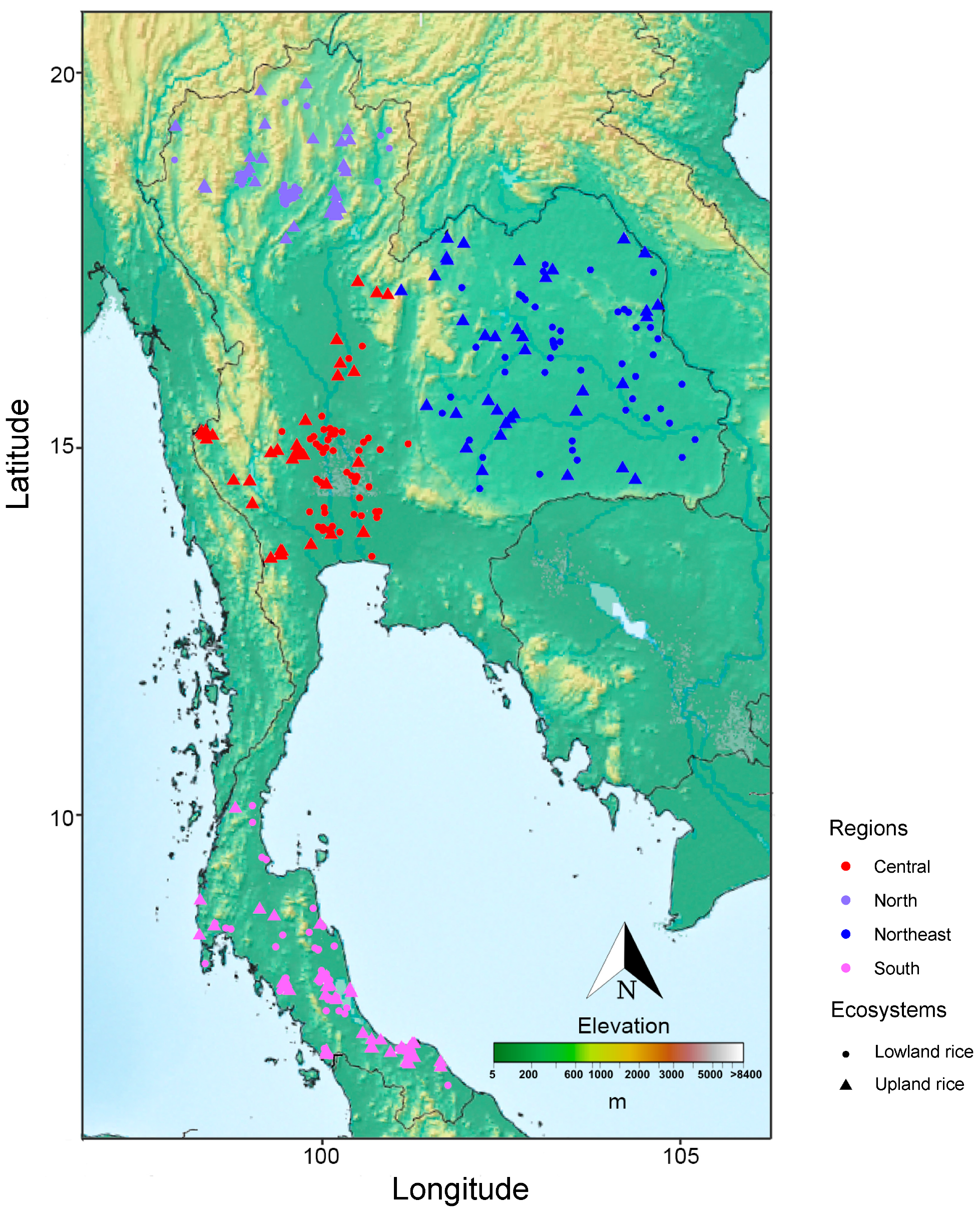
2.1. Plant Materials
2.2. DNA Extraction and SNP Genotyping
2.3. Data Management and Analysis
3. Results
3.1. Genetic Variability of 365 Rice Accessions Based on SNP Markers
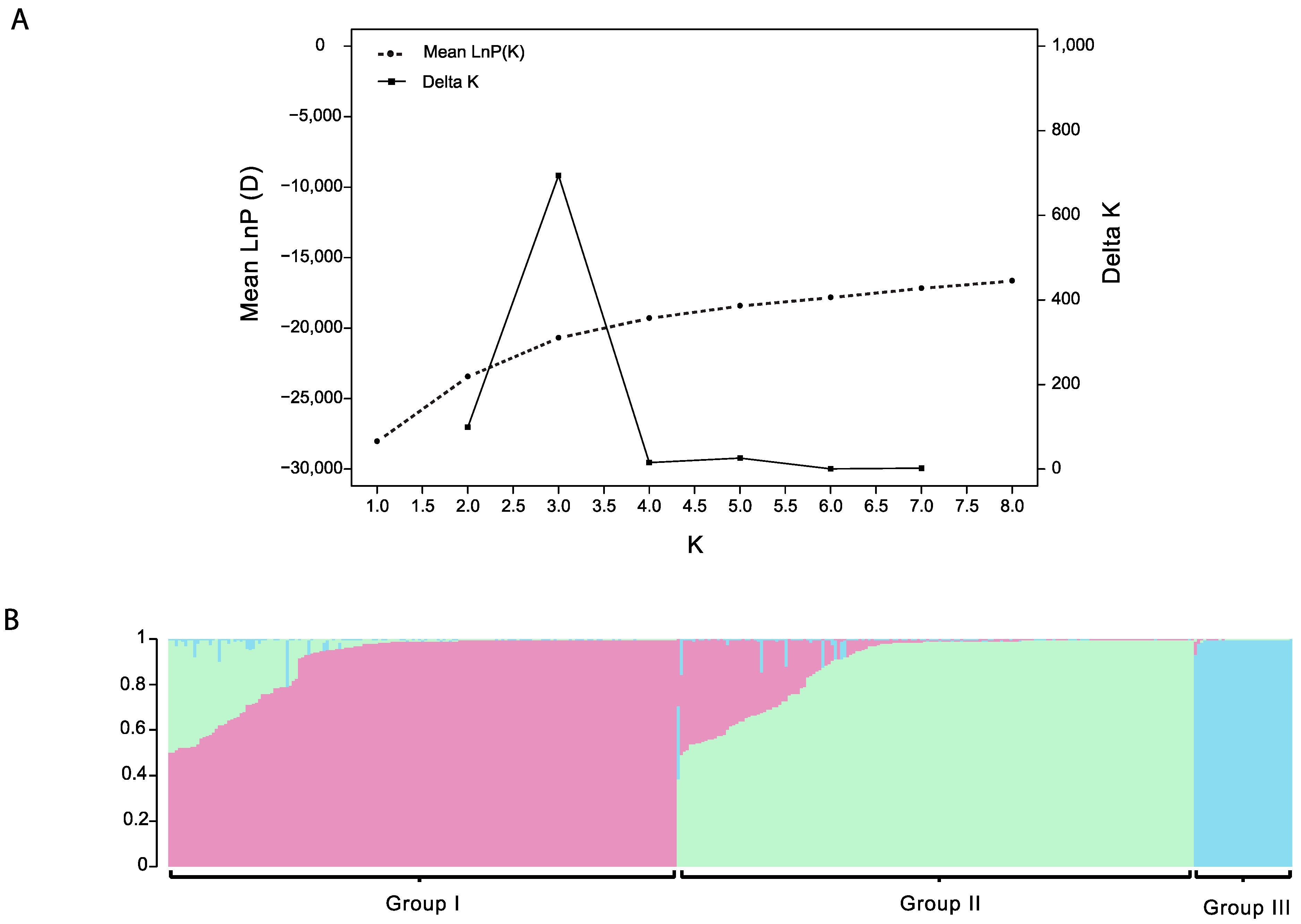
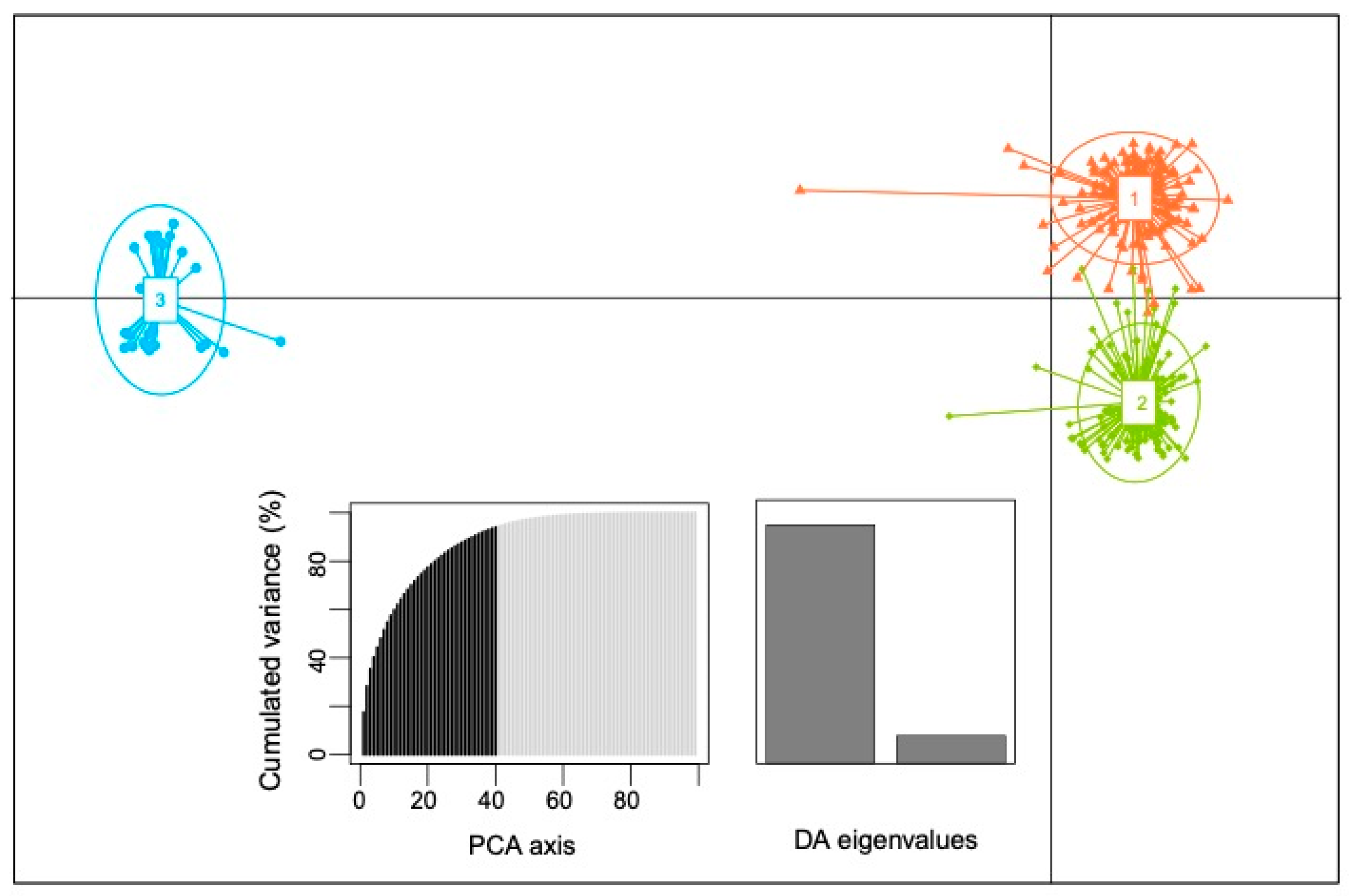
3.2. Population Structure of Thai Landraces
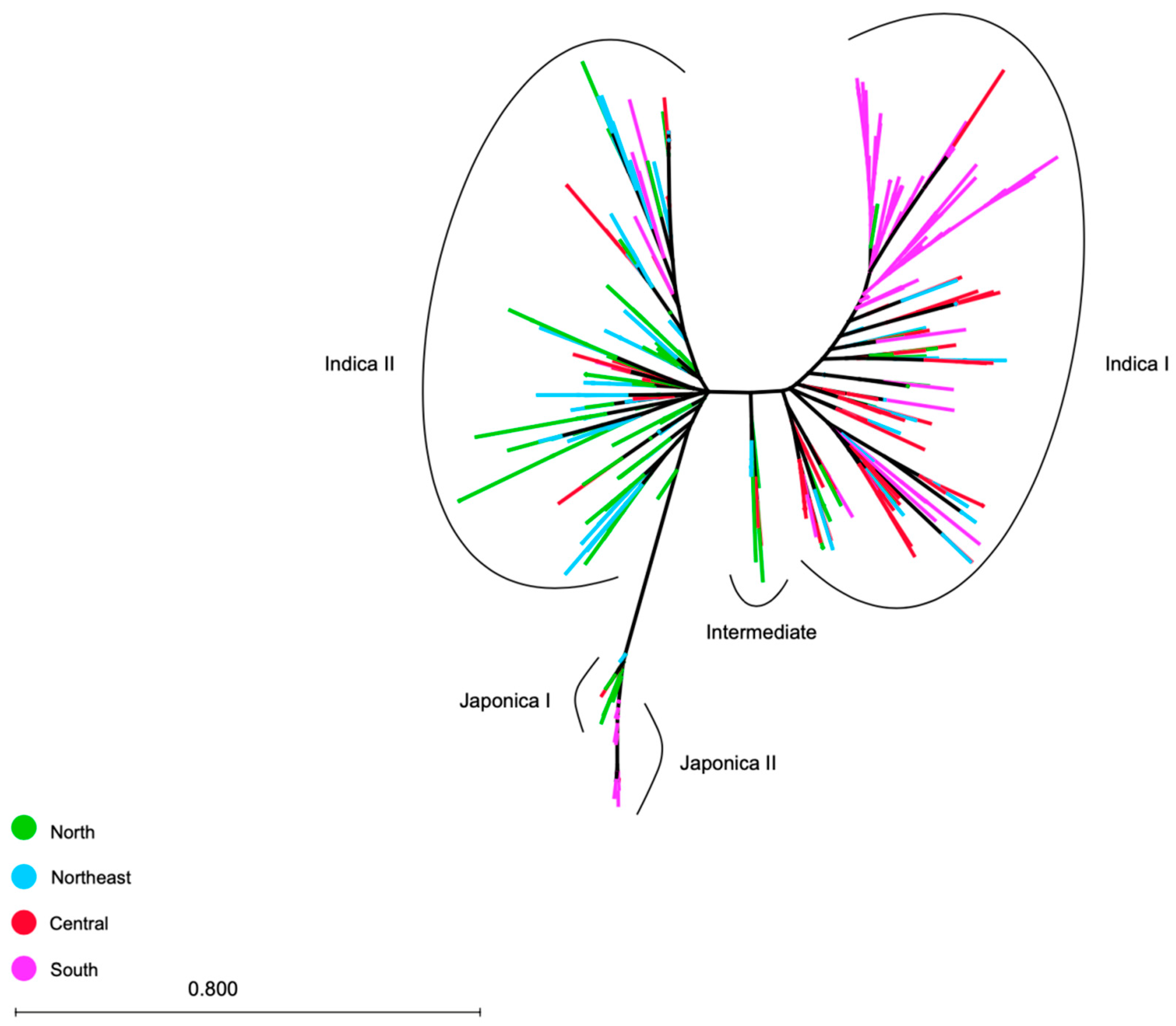
3.3. Genetic Distance and Phylogeny of the 365 Accessions
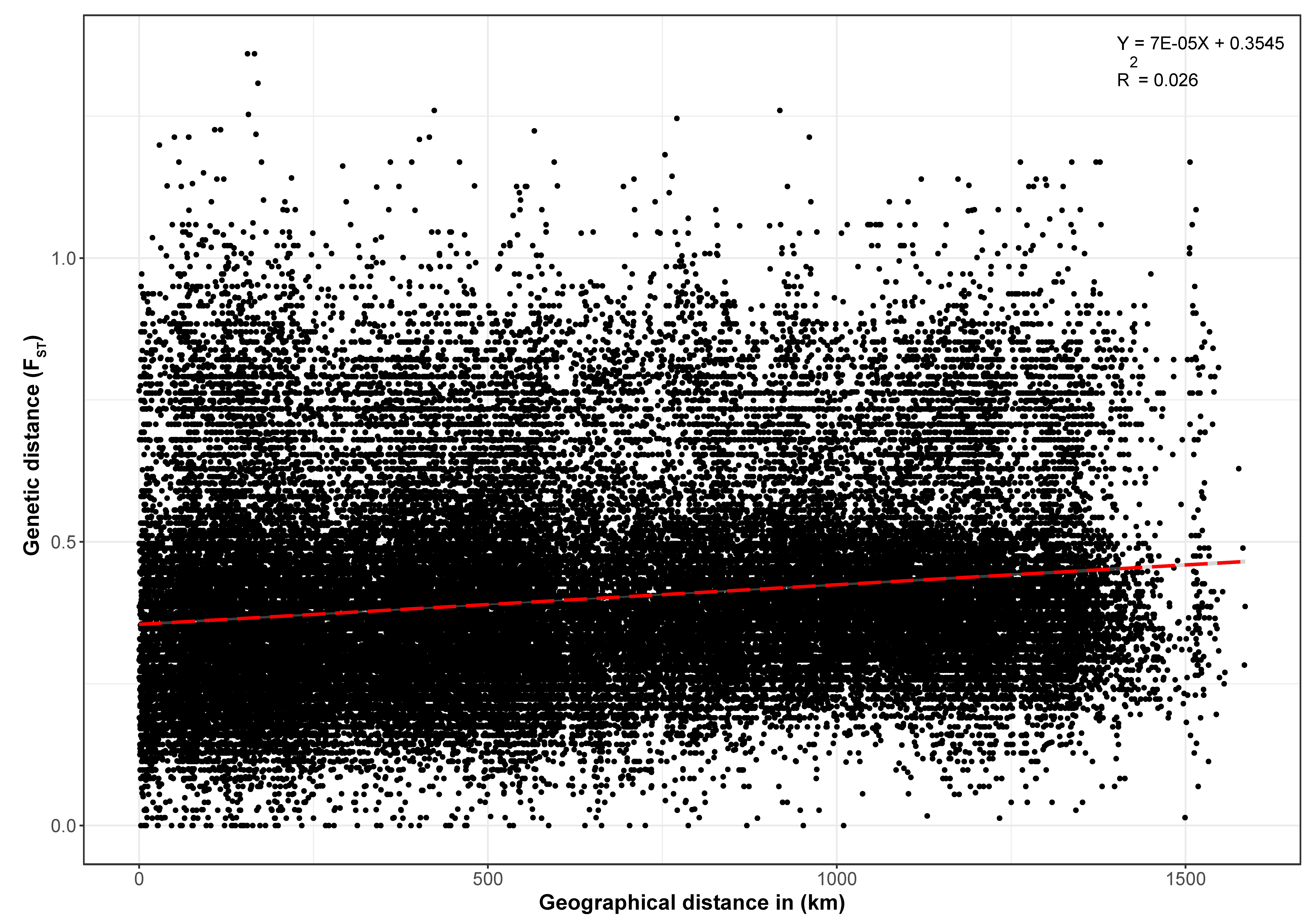
3.4. Genetic Differentiation, AMOVA, and Isolation-by-Distance Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khush, G.S. Origin, dispersal, cultivation and variation of rice. Plant Mol. Biol. 1997, 35, 25–34. [Google Scholar] [CrossRef]
- Bandumula, N. Rice production in asia: Key to global food security. Proc. Natl. Acad. Sci. USA 2018, 88, 1323–1328. [Google Scholar] [CrossRef]
- Silva, C.N. Urban Planning in Sub-Saharan Africa: Colonial and Post-Colonial Planning Cultures; Routledge: London, UK, 2015; ISBN 9781315797311. [Google Scholar]
- Oka, H.-I.; Morishima, H. Phylogenetic differentiation of cultivated rice, XXIII. Potentiality of wild progenitors to evolve the Indica and Japonica types of rice cultivars. Euphytica 1982, 31, 41–50. [Google Scholar] [CrossRef]
- Mackill, D.J.; Lei, X. Genetic variation for traits related to temperate adaptation of rice cultivars. Crop Sci. 1997, 37, 1340–1346. [Google Scholar] [CrossRef]
- Mackill, D.J.; Khush, G.S. IR64: A high-quality and high-yielding mega variety. Rice 2018, 11, 18. [Google Scholar] [CrossRef]
- Singh, R.K. Genetic resource and the role of international collaboration in rice breeding. Genome 1999, 42, 635–641. [Google Scholar] [CrossRef]
- Harlan, J.R. Our vanishing genetic resources. Science 1975, 188, 617–621. [Google Scholar] [CrossRef]
- Kovach, M.J.; McCouch, S.R. Leveraging natural diversity: Back through the bottleneck. Curr. Opin. Plant Biol. 2008, 11, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Govindaraj, M.; Vetriventhan, M.; Srinivasan, M. Importance of genetic diversity assessment in crop plants and its recent advances: An overview of its analytical perspectives. Genet Res Int 2015, 2015, 431487. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, H.R.; Bhanu, A.N.; Srivastava, K.; Singh, M.N. Assessment of genetic diversity in crop plants-an overview. Adv. Plants Agric. 2017, 7, 255. [Google Scholar]
- Glaszmann, J.C. Isozymes and classification of Asian rice varieties. Theor. Appl. Genet. 1987, 74, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Garris, A.J.; Tai, T.H.; Coburn, J.; Kresovich, S.; McCouch, S. Genetic structure and diversity in Oryza sativa L. Genetics 2005, 169, 1631–1638. [Google Scholar] [CrossRef]
- Sow, M.; Ndjiondjop, M.-N.; Sido, A.; Mariac, C.; Laing, M.; Bezançon, G. Genetic diversity, population structure and differentiation of rice species from Niger and their potential for rice genetic resources conservation and enhancement. Genet. Resour. Crop Evol. 2014, 61, 199–213. [Google Scholar] [CrossRef]
- Caicedo, A.L.; Williamson, S.H.; Hernandez, R.D.; Boyko, A.; Fledel-Alon, A.; York, T.L.; Polato, N.R.; Olsen, K.M.; Nielsen, R.; McCouch, S.R.; et al. Genome-wide patterns of nucleotide polymorphism in domesticated rice. PLoS Genet. 2007, 3, 1745–1756. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Kurata, N.; Wei, X.; Wang, Z.-X.; Wang, A.; Zhao, Q.; Zhao, Y.; Liu, K.; Lu, H.; Li, W.; et al. A map of rice genome variation reveals the origin of cultivated rice. Nature 2012, 490, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Tam, N.T.; Dwiyanti, M.S.; Koide, Y.; Nagano, A.J.; Ky, H.; Tin, H.Q.; Hien, N.L.; Dung, L.V.; Kishima, Y. Profiling SNP and nucleotide diversity to characterize mekong delta rice landraces in southeast asian populations. Plant Genome 2019, 12, 190042. [Google Scholar] [CrossRef]
- Zhao, K.; Wright, M.; Kimball, J.; Eizenga, G.; McClung, A.; Kovach, M.; Tyagi, W.; Ali, M.L.; Tung, C.-W.; Reynolds, A.; et al. Genomic diversity and introgression in O. sativa reveal the impact of domestication and breeding on the rice genome. PLoS ONE 2010, 5, e10780. [Google Scholar] [CrossRef] [PubMed]
- Sang, T.; Ge, S. Genetics and phylogenetics of rice domestication. Curr. Opin. Genet. Dev. 2007, 17, 533–538. [Google Scholar] [CrossRef]
- Sweeney, M.; McCouch, S. The complex history of the domestication of rice. Ann. Bot. 2007, 100, 951–957. [Google Scholar] [CrossRef]
- Kladmook, M.; Kumchoo, T.; Hongtrakul, V. Genetic diversity analysis and subspecies classification of Thailand rice landraces using DNA markers. Afr. J. Biotechnol. 2012, 11, 14044–14053. [Google Scholar] [CrossRef]
- Nilthong, S.; Chukeatirote, E. Assessment of genetic diversity in Thai upland rice varieties using SSR markers. Aust. J. Crop. Sci. 2020. [Google Scholar] [CrossRef]
- Pathaichindachote, W.; Panyawut, N.; Sikaewtung, K.; Patarapuwadol, S.; Muangprom, A. Genetic diversity and allelic frequency of selected thai and exotic rice germplasm using SSR markers. Rice Sci. 2019, 26, 393–403. [Google Scholar] [CrossRef]
- Chakhonkaen, S.; Pitnjam, K.; Saisuk, W.; Ukoskit, K.; Muangprom, A. Genetic structure of Thai rice and rice accessions obtained from the International Rice Research Institute. Rice 2012, 5, 19. [Google Scholar] [CrossRef] [PubMed]
- Pusadee, T.; Wongtamee, A.; Rerkasem, B.; Olsen, K.M.; Jamjod, S. Farmers Drive Genetic Diversity of Thai Purple Rice (Oryza sativa L.) Landraces. Econ Bot 2019, 73, 76–85. [Google Scholar] [CrossRef]
- Nei, M. Genetic Distance between Populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Liu, K.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef]
- Alexandrov, N.; Tai, S.; Wang, W.; Mansueto, L.; Palis, K.; Fuentes, R.R.; Ulat, V.J.; Chebotarov, D.; Zhang, G.; Li, Z.; et al. SNP-Seek database of SNPs derived from 3000 rice genomes. Nucleic Acids Res. 2015, 43, D1023–D1027. [Google Scholar] [CrossRef]
- Perrier, X.; Jacquemoud-Collet, J.P. DARwin software. 2006. Available online: https://darwin.cirad.fr (accessed on 4 May 2021).
- Villanueva, R.A.M.; Chen, Z.J. ggplot2: Elegant Graphics for Data Analysis (2nd ed.). Meas. Interdiscip. Res. Perspect. 2019, 17, 160–167. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Genalex, S. 6: Genetic analysis in Excel. Population genetic software for teaching and research. Bioinformatics 2012, 28, 19. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P. Vegan: Community Ecology Package. R-Package Version 2.0-10. 2013. Available online: http://CRAN.R (accessed on 4 May 2021).
- Wright, S. Evolution and the genetics of populations, volume 4: Variability within and among natural populations. Biometrics 1979, 35, 359. [Google Scholar] [CrossRef]
- Govindaraju, D.R. Variation in gene flow levels among predominantly self-pollinated plants. J. Evol. Biol. 1989, 2, 173–181. [Google Scholar] [CrossRef]
- Eltaher, S.; Sallam, A.; Belamkar, V.; Emara, H.A.; Nower, A.A.; Salem, K.F.M.; Poland, J.; Baenziger, P.S. Genetic Diversity and Population Structure of F3:6 Nebraska Winter Wheat Genotypes Using Genotyping-By-Sequencing. Front. Genet. 2018, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Oka, H.I.; Chang, W.T. A note on rice varieties of Japonica type found in northern Thailand. Bot. Bull. Acad. Sinica 1963, 4, 163–168. [Google Scholar]
- Frankham, R.; Ballou, J.D.; Briscoe, D.A.; McInnes, K.H. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2002; ISBN 9780511808999. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Chr. | Position | REF | ALT | MAF | Na | Ho | He | PIC |
|---|---|---|---|---|---|---|---|---|---|
| R01002684973 | 1 | 2684973 | C | A | 0.65 | 2 | 0 | 0.46 | 0.35 |
| R01008961781 | 1 | 8961781 | C | A | 0.89 | 2 | 0.01 | 0.19 | 0.17 |
| R01023762056 | 1 | 23762056 | G | A | 0.7 | 2 | 0 | 0.42 | 0.33 |
| R01033064986 | 1 | 33064986 | T | C | 0.91 | 2 | 0 | 0.16 | 0.15 |
| R01033454937 | 1 | 33454937 | A | T | 0.64 | 2 | 0 | 0.46 | 0.36 |
| R01034609753 | 1 | 34609753 | C | G | 0.93 | 2 | 0 | 0.13 | 0.12 |
| R01036673058 | 1 | 36673058 | C | T | 0.91 | 2 | 0 | 0.17 | 0.15 |
| R02008303326 | 2 | 8303326 | G | A | 0.89 | 2 | 0.01 | 0.20 | 0.18 |
| R02008313120 | 2 | 8313120 | G | T | 0.69 | 2 | 0.01 | 0.43 | 0.34 |
| R02008315835 | 2 | 8315835 | T | A | 0.94 | 2 | 0 | 0.12 | 0.11 |
| R02010864977 | 2 | 10864977 | G | T | 0.82 | 2 | 0 | 0.30 | 0.25 |
| R02019361361 | 2 | 19361361 | A | C | 0.9 | 2 | 0 | 0.18 | 0.17 |
| R02019361451 | 2 | 19361451 | G | T | 0.9 | 2 | 0 | 0.18 | 0.17 |
| R03016733359 | 3 | 16733359 | A | C | 0.72 | 2 | 0.02 | 0.40 | 0.32 |
| R03017286744 | 3 | 17286744 | G | T | 0.81 | 2 | 0 | 0.31 | 0.26 |
| R03017571575 | 3 | 17571575 | G | A | 0.84 | 2 | 0.01 | 0.27 | 0.23 |
| R03031335170 | 3 | 31335170 | C | T | 0.84 | 2 | 0.01 | 0.27 | 0.23 |
| R03031437173 | 3 | 31437173 | C | A | 0.91 | 2 | 0 | 0.16 | 0.15 |
| R04001019335 | 4 | 1019335 | A | G | 0.79 | 2 | 0 | 0.33 | 0.28 |
| R04005867320 | 4 | 5867320 | T | G | 0.66 | 2 | 0 | 0.45 | 0.35 |
| R04006451939 | 4 | 6451939 | C | T | 0.86 | 2 | 0 | 0.24 | 0.21 |
| R04006968050 | 4 | 6968050 | A | G | 0.93 | 2 | 0 | 0.13 | 0.12 |
| R04011087541 | 4 | 11087541 | G | T | 0.69 | 2 | 0.02 | 0.43 | 0.34 |
| R04011803874 | 4 | 11803874 | T | A | 0.58 | 2 | 0 | 0.49 | 0.37 |
| R04016214692 | 4 | 16214692 | CA | TG | 0.57 | 2 | 0 | 0.49 | 0.37 |
| R04016218749 | 4 | 16218749 | T | G | 0.57 | 2 | 0.01 | 0.49 | 0.37 |
| R04022184296 | 4 | 22184296 | A | G | 0.64 | 2 | 0.02 | 0.46 | 0.35 |
| R04023172729 | 4 | 23172729 | T | G | 0.74 | 2 | 0 | 0.38 | 0.31 |
| R04023175725 | 4 | 23175725 | T | C | 0.75 | 2 | 0 | 0.37 | 0.3 |
| R05003673333 | 5 | 3673333 | T | C | 0.68 | 2 | 0.01 | 0.44 | 0.34 |
| R05019155333 | 5 | 19155333 | A | G | 0.59 | 2 | 0.01 | 0.48 | 0.37 |
| R05019155705 | 5 | 19155705 | G | T | 0.5 | 2 | 0.01 | 0.50 | 0.37 |
| R05023218617 | 5 | 23218617 | A | C | 0.7 | 2 | 0 | 0.42 | 0.33 |
| R05026487913 | 5 | 26487913 | A | G | 0.91 | 2 | 0 | 0.17 | 0.15 |
| R05028876504 | 5 | 28876504 | A | T | 0.91 | 2 | 0.01 | 0.16 | 0.15 |
| R05028876779 | 5 | 28876779 | A | C | 0.91 | 2 | 0.01 | 0.16 | 0.15 |
| R06001693194 | 6 | 1693194 | G | A | 0.54 | 2 | 0.01 | 0.50 | 0.37 |
| R06001693411 | 6 | 1693411 | A | C | 0.56 | 2 | 0.01 | 0.49 | 0.37 |
| R06001765760 | 6 | 1765760 | G | T | 0.54 | 2 | 0 | 0.50 | 0.37 |
| R06001768006 | 6 | 1768006 | A | C | 0.79 | 2 | 0 | 0.33 | 0.27 |
| R06001768724 | 6 | 1768724 | T | C | 0.82 | 2 | 0 | 0.29 | 0.25 |
| R06001768997 | 6 | 1768997 | C | T | 0.69 | 2 | 0.01 | 0.43 | 0.34 |
| R06006752886 | 6 | 6752886 | G | T | 0.55 | 2 | 0.01 | 0.49 | 0.37 |
| R07005873563 | 7 | 5873563 | G | A | 0.79 | 2 | 0 | 0.33 | 0.28 |
| R07020013105 | 7 | 20013105 | G | T | 0.55 | 2 | 0 | 0.49 | 0.37 |
| R07020826100 | 7 | 20826100 | G | T | 0.82 | 2 | 0.01 | 0.30 | 0.26 |
| R07024350575 | 7 | 24350575 | C | T | 0.91 | 2 | 0 | 0.17 | 0.15 |
| R07025982551 | 7 | 25982551 | A | G | 0.91 | 2 | 0 | 0.16 | 0.15 |
| R07027746661 | 7 | 27746661 | C | G | 0.53 | 2 | 0.01 | 0.50 | 0.37 |
| R08002269285 | 8 | 2269285 | G | C | 0.56 | 2 | 0 | 0.49 | 0.37 |
| R08002890407 | 8 | 2890407 | G | A | 0.63 | 2 | 0 | 0.46 | 0.36 |
| R08003007900 | 8 | 3007900 | C | G | 0.54 | 2 | 0 | 0.50 | 0.37 |
| R08020382861 | 8 | 20382861 | ATTATGGC | -:- | 0.87 | 2 | 0 | 0.23 | 0.2 |
| R08027057202 | 8 | 27057202 | G | A | 0.83 | 2 | 0 | 0.28 | 0.24 |
| R08027176617 | 8 | 27176617 | C | A | 0.72 | 2 | 0 | 0.40 | 0.32 |
| R08027943348 | 8 | 27943348 | G | A | 0.86 | 2 | 0 | 0.24 | 0.21 |
| R09004198183 | 9 | 4198183 | C | A | 0.77 | 2 | 0.01 | 0.35 | 0.29 |
| R09007245205 | 9 | 7245205 | A | T | 0.9 | 2 | 0.01 | 0.18 | 0.16 |
| R09007245448 | 9 | 7245448 | C | T | 0.9 | 2 | 0.01 | 0.18 | 0.16 |
| R09007245650 | 9 | 7245650 | A | G | 0.9 | 2 | 0 | 0.17 | 0.16 |
| R09007245739 | 9 | 7245739 | C | G | 0.9 | 2 | 0.01 | 0.17 | 0.16 |
| R09007246222 | 9 | 7246223 | T | G | 0.9 | 2 | 0.01 | 0.18 | 0.16 |
| R09007246804 | 9 | 7246804 | T | G | 0.85 | 2 | 0.01 | 0.26 | 0.23 |
| R10015680316 | 10 | 15680316 | C | A | 0.92 | 2 | 0 | 0.15 | 0.14 |
| R10021955049 | 10 | 21955049 | A | G | 0.9 | 2 | 0 | 0.18 | 0.16 |
| R11013840467 | 11 | 13840467 | A | G | 0.81 | 2 | 0 | 0.30 | 0.26 |
| R11014118135 | 11 | 14118135 | A | G | 0.6 | 2 | 0.01 | 0.48 | 0.36 |
| R11014169508 | 11 | 14169508 | C | A | 0.75 | 2 | 0.01 | 0.37 | 0.3 |
| R11021481048 | 11 | 21481048 | ATT | -:- | 0.52 | 2 | 0 | 0.50 | 0.37 |
| R11028209211 | 11 | 28209211 | A | T | 0.7 | 2 | 0 | 0.42 | 0.33 |
| R12009613648 | 12 | 9613648 | A | C | 0.9 | 2 | 0.01 | 0.18 | 0.17 |
| R12018759236 | 12 | 18759236 | T | G | 0.85 | 2 | 0 | 0.25 | 0.22 |
| R12018856177 | 12 | 18856177 | G | A | 0.83 | 2 | 0 | 0.29 | 0.24 |
| R12022373643 | 12 | 22373643 | A | G | 0.69 | 2 | 0.01 | 0.43 | 0.34 |
| R12023173265 | 12 | 23173265 | T | A | 0.77 | 2 | 0 | 0.36 | 0.29 |
| Population | Sample Size | Major Allele Frequency/Locus | Mean Gene Diversity | Mean PIC Value |
|---|---|---|---|---|
| All subpopulations | 365 | 0.76 | 0.33 | 0.26 |
| Central (C) | 89 | 0.77 | 0.32 | 0.25 |
| Northern (N) | 96 | 0.80 | 0.29 | 0.24 |
| Northeastern (NE) | 86 | 0.81 | 0.28 | 0.23 |
| Southern (S) | 94 | 0.77 | 0.31 | 0.25 |
| Population | Sample Size | Major Allele Frequency /Locus | Mean Gene Diversity | Mean PIC Value |
|---|---|---|---|---|
| All subpopulations | 365 | 0.76 | 0.32 | 0.26 |
| Group I (indica) | 167 | 0.80 | 0.27 | 0.21 |
| Group II (indica) | 166 | 0.82 | 0.24 | 0.19 |
| Group III (japonica) | 32 | 0.93 | 0.09 | 0.07 |
| Subpopulations | Central | North | Northeast | South |
|---|---|---|---|---|
| Central (C) | 0.000 | |||
| Northern (N) | 0.039 | 0.000 | ||
| Northeast (NE) | 0.023 | 0.016 | 0.000 | |
| South (S) | 0.052 | 0.069 | 0.078 | 0.000 |
| Source | df | SS | MS | Estimated Variance | Percentage of Total Variance | Probability (p) |
|---|---|---|---|---|---|---|
| Among Pops | 3 | 1463.724 | 487.908 | 4.835 | 9% | <0.001 |
| Within Pops | 361 | 16970.874 | 47.011 | 47.011 | 91% | |
| Total | 364 | 18434.597 | 51.846 | 100% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aesomnuk, W.; Ruengphayak, S.; Ruanjaichon, V.; Sreewongchai, T.; Malumpong, C.; Vanavichit, A.; Toojinda, T.; Wanchana, S.; Arikit, S. Estimation of the Genetic Diversity and Population Structure of Thailand’s Rice Landraces Using SNP Markers. Agronomy 2021, 11, 995. https://doi.org/10.3390/agronomy11050995
Aesomnuk W, Ruengphayak S, Ruanjaichon V, Sreewongchai T, Malumpong C, Vanavichit A, Toojinda T, Wanchana S, Arikit S. Estimation of the Genetic Diversity and Population Structure of Thailand’s Rice Landraces Using SNP Markers. Agronomy. 2021; 11(5):995. https://doi.org/10.3390/agronomy11050995
Chicago/Turabian StyleAesomnuk, Wanchana, Siriphat Ruengphayak, Vinitchan Ruanjaichon, Tanee Sreewongchai, Chanate Malumpong, Apichart Vanavichit, Theerayut Toojinda, Samart Wanchana, and Siwaret Arikit. 2021. "Estimation of the Genetic Diversity and Population Structure of Thailand’s Rice Landraces Using SNP Markers" Agronomy 11, no. 5: 995. https://doi.org/10.3390/agronomy11050995
APA StyleAesomnuk, W., Ruengphayak, S., Ruanjaichon, V., Sreewongchai, T., Malumpong, C., Vanavichit, A., Toojinda, T., Wanchana, S., & Arikit, S. (2021). Estimation of the Genetic Diversity and Population Structure of Thailand’s Rice Landraces Using SNP Markers. Agronomy, 11(5), 995. https://doi.org/10.3390/agronomy11050995