
Genome Enhanced Marker Improvement for Potato Virus Y Disease Resistance in Potato

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Sampling
2.2. Phenotyping
2.3. DNA Extractions
2.4. Bioinformatic Data Analysis
2.5. KASP Assay
2.6. Protein Characterisation
3. Results
Protein Characterisation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Statistical Database Food and Agriculture Organization of the United Nations; FAO: Rome, Italy, 2019. [Google Scholar]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Kehoe, M.A.; Jones, R.A.C. Improving Potato virus Y strain nomenclature: Lessons from comparing isolates obtained over a 73-year period. Plant Pathol. 2016, 65, 322–333. [Google Scholar] [CrossRef]
- Jeffries, C. Technical Guidelines for the Safe Movement of Germplasm. No19 Potato; FAO: Rome, Italy, 1998; Volume 177. [Google Scholar]
- Nolte, P.; Whitworth, J.L.; Thornton, M.K.; McIntosh, C.S. Effect of seedborne Potato virus Y on performance of Russet Burbank, Russet Norkotah, and Shepody potato. Plant Dis. 2004, 88, 248–252. [Google Scholar] [CrossRef]
- Rodriguez-Rodriguez, M.; Chikh-Ali, M.; Johnson, S.B.; Gray, S.; Malseed, N.; Crump, N.S.; Karasev, A.V. The recombinant Potato Virus Y (PVY) strain, PVYNTN, identified in potato fields in Victoria, South Eastern Australia. Plant Dis. 2020, 104. [Google Scholar] [CrossRef] [PubMed]
- Swiezynski, K. Inheritance of resistance to viruses. Potato Genetics; Scottish Crop Research Institute: Dundee, UK, 1994. [Google Scholar]
- Bradshaw, J.; Mackay, G. Breeding Strategies for Clonally Propagated Potatoes; Scottish Crop Research Institute: Dundee, UK, 1994. [Google Scholar]
- Gebhardt, C. Bridging the gap between genome analysis and precision breeding in potato. Trends Genet. 2013, 29, 248–256. [Google Scholar] [CrossRef]
- Ortega, F.; Lopez-Vizcon, C. Application of molecular marker-assisted selection (MAS) for disease resistance in a practical potato breeding programme. Potato Res. 2012, 55, 1–13. [Google Scholar] [CrossRef]
- Lindhout, P.; Meijer, D.; Schotte, T.; Hutten, R.C.; Visser, R.G.; van Eck, H.J. Towards F1 hybrid seed potato breeding. Potato Res. 2011, 54, 301–312. [Google Scholar] [CrossRef]
- Slater, A.T.; Cogan, N.O.I.; Forster, J.W. Cost Analysis of the application of marker-assisted selection in potato breeding. Mol. Breed. 2013, 32, 299–310. [Google Scholar] [CrossRef]
- Hospital, F. Challenges for effective marker-assisted selection in plants. Genetica 2009, 136, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Barker, H. Extreme resistance to Potato virus V in clones of Solanum tuberosum that are also resistant to Potato viruses Y and A: Evidence for a Locus Conferring Broad-Spectrum Potyvirus Resistance. Theor. Appl. Genet. 1997, 95, 1258–1262. [Google Scholar] [CrossRef]
- Hämäläinen, J.H.; Sorri, V.A.; Watanabe, K.N.; Gebhardt, C.; Valkonen, J.P.T. Molecular examination of a chromosome region that controls resistance to Potato Y and a Potyviruses in potato. Theor. Appl. Genet. 1998, 96, 1036–1043. [Google Scholar] [CrossRef]
- Cockerham, G. Genetical studies on resistance to potato viruses X and Y. Heredity 1970, 25, 309–348. [Google Scholar] [CrossRef]
- Gebhardt, C.; Bellin, D.; Henselewski, H.; Lehmann, W.; Schwarzfischer, J.; Valkonen, J.P.T. Marker-assisted combination of major genes for pathogen resistance in potato. Theor. Appl. Genet. 2006, 112, 1458–1464. [Google Scholar] [CrossRef]
- Valkonen, J.P.T.; Wiegmann, K.; Hämäläinen, J.H.; Marczewski, W.; Watanabe, K.N. Evidence for utility of the same PCR-based markers for selection of extreme resistance to Potato virus Y controlled by Rysto of Solanum stoloniferum derived from different sources. Ann. Appl. Biol. 2008, 152, 121–130. [Google Scholar] [CrossRef]
- Ottoman, R.J.; Hane, D.C.; Brown, C.R.; Yilma, S.; James, S.R.; Mosley, A.R.; Crosslin, J.M.; Vales, M.I. Validation and implementation of marker-assisted selection (MAS) for PVY resistance (Ryadg gene) in a tetraploid potato breeding program. Am. J. Potato Res. 2009, 86, 304–314. [Google Scholar] [CrossRef]
- Ross, H.; Hunnius, W. Potato Breeding: Problems and Perspectives; V.P. Parey: Berlin, Germany, 1986. [Google Scholar]
- Mendoza, H.A.; Mihovilovich, E.J.; Saguma, E. Identification of triplex (YYYy) potato virus y (PVY) immune progenitors derived from Solanum tuberosum ssp.andigena. Am. Potato J. 1996, 73, 7. [Google Scholar] [CrossRef]
- Kasai, K.; Morikawa, Y.; Sorri, V.A.; Valkonen, J.; Gebhardt, C.; Watanabe, K.N. Development of SCAR markers to the PVY resistance gene Ryadg based on a common feature of plant disease resistance genes. Genome 2000, 43, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Brigneti, G.; Garcia-Mas, J.; Baulcombe, D.C. Molecular mapping of the Potato virus Y resistance gene Rysto in potato. Theor. Appl. Genet. 1997, 94, 198–203. [Google Scholar] [CrossRef]
- Slater, A.T. Molecular and Quantitative Genetic Studies for Potato Germplasm Enhancement. Ph.D. Thesis, La Trobe University, Bundoora, Australia, 2013. [Google Scholar]
- Slater, A.T.; Schultz, L.; Lombardi, M.; Rodoni, B.C.; Bottcher, C.; Cogan, N.O.I.; Forster, J.W. Screening for resistance to PVY in Australian potato germplasm. Genes 2020, 11, 429. [Google Scholar] [CrossRef]
- Dalla Rizza, M.; Vilaró, F.L.; Torres, D.G.; Maeso, D. Detection of PVY extreme resistance genes in potato germplasm from the Uruguayan breeding program. Am. J. Potato Res. 2006, 83, 297–304. [Google Scholar] [CrossRef]
- del Rosario Herrera, M.; Vidalon, L.J.; Montenegro, J.D.; Riccio, C.; Guzman, F.; Bartolini, I.; Ghislain, M. Molecular and genetic characterization of the Ryadg locus on chromosome XI from Andigena potatoes conferring extreme resistance to Potato virus Y. Theor. Appl. Genet. 2018, 131, 1925–1938. [Google Scholar] [CrossRef] [PubMed]
- The Potato Genome Sequencing Consortium. Genome sequence and analysis of the tuber crop potato. Nature 2011, 475, 189–195. [Google Scholar] [CrossRef]
- Uitdewilligen, J.G.A.M.L.; Wolters, A.-M.A.; D’hoop, B.B.; Borm, T.J.A.; Visser, R.G.F.; van Eck, H.J. A next-generation sequencing method for genotyping-by-sequencing of highly heterozygous autotetraploid potato. PLoS ONE 2013, 8, e62355. [Google Scholar] [CrossRef]
- Vos, P.G.; Uitdewilligen, J.G.A.M.L.; Voorrips, R.E.; Visser, R.G.F.; van Eck, H.J. Development and analysis of a 20K SNP array for potato (Solanum tuberosum): An insight into the breeding history. Theor. Appl. Genet. 2015, 128, 2387–2401. [Google Scholar] [CrossRef] [PubMed]
- Caruana, B.M.; Pembleton, L.W.; Constable, F.; Rodoni, B.; Slater, A.T.; Cogan, N.O.I. Validation of genotyping by sequencing using transcriptomics for diversity and application of genomic selection in tetraploid potato. Front. Plant Sci. 2019, 10, 670. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Sharma, S.K.; Bolser, D.; de Boer, J.; Sønderkær, M.; Amoros, W.; Carboni, M.F.; D’Ambrosio, J.M.; de la Cruz, G.; Di Genova, A.; Douches, D.S.; et al. Construction of reference chromosome-scale pseudomolecules for potato: Integrating the potato genome with genetic and physical maps. G3 GenesGenomesGenetics 2013, 3, 2031–2047. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The genome analysis toolkit: A mapreduce framework for analyzing next-generation dna sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.L.; Attwood, T.K.; Babbitt, P.C.; Blum, M.; Bork, P.; Bridge, A.; Brown, S.D.; Chang, H.-Y.; El-Gebali, S.; Fraser, M.I.; et al. InterPro in 2019: Improving coverage, classification and access to protein sequence annotations. Nucleic Acids Res. 2019, 47, D351–D360. [Google Scholar] [CrossRef]
- Zhang, C.; Freddolino, P.L.; Zhang, Y. COFACTOR: Improved protein function prediction by combining structure, sequence and protein–protein interaction information. Nucleic Acids Res. 2017, 45, W291–W299. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, Y. I-TASSER Server: New development for protein structure and function predictions. Nucleic Acids Res. 2015, 43, W174–W181. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, Scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Munoz, F.J.; Plaisted, R.L.; Thurston, H.D. Resistance to Potato virus Y in Solanum tuberosum spp. Andigena. Am. Potato J. 1975, 52, 107–115. [Google Scholar] [CrossRef]
- Galvez, R.; Brown, C.R. Inheritance of Extreme resistance to PVY derived from Solanum tuberosum ssp. Andigena. Am. Potato J. 1980, 57, 476–477. [Google Scholar]
- Bent, A.F.; Kunkel, B.N.; Dahlbeck, D.; Brown, K.L.; Schmidt, R.; Giraudat, J.; Leung, J.; Staskawicz, B.J. RPS2 of Arabidopsis thaliana: A leucine-rich repeat class of plant disease resistance genes. Science 1994, 265, 1856–1860. [Google Scholar] [CrossRef]
- Whitham, S.; Dinesh-Kumar, S.P.; Choi, D.; Hehl, R.; Corr, C.; Baker, B. The product of the Tobacco mosaic virus resistance gene N: Similarity to toll and the interleukin-1 receptor. Cell 1994, 78, 1101–1115. [Google Scholar] [CrossRef]
- Mindrinos, M.; Katagiri, F.; Yu, G.-L.; Ausubel, F.M. The A. Thaliana disease resistance gene RPS2 encodes a protein containing a nucleotide-binding site and leucine-rich repeats. Cell 1994, 78, 1089–1099. [Google Scholar] [CrossRef]
- Barragan, A.C.; Weigel, D. Plant NLR Diversity: The known unknowns of pan-NLRomes. Plant Cell 2020, 1–18. [Google Scholar] [CrossRef]
- Ganal, M.W.; Tanksley, S.D. Recombination around the Tm2a and Mi resistance genes in different crosses of Lycopersicon peruvianum. Theor. Appl. Genet. 1996, 92, 101–108. [Google Scholar] [CrossRef]
- Wei, F.; Wing, R.A.; Wise, R.P. Genome dynamics and evolution of the Mla (powdery mildew) resistance locus in barley. Plant Cell 2002, 14, 1903–1917. [Google Scholar] [CrossRef]
- Finkers-Tomczak, A.; Bakker, E.; de Boer, J.; van der Vossen, E.; Achenbach, U.; Golas, T.; Suryaningrat, S.; Smant, G.; Bakker, J.; Goverse, A. Comparative sequence analysis of the potato cyst nematode resistance locus H1 reveals a major lack of co-linearity between three haplotypes in potato (Solanum tuberosum ssp.). Theor. Appl. Genet. 2011, 122, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Schultz, L.; Cogan, N.O.I.; McLean, K.; Dale, M.F.B.; Bryan, G.J.; Forster, J.W.; Slater, A.T. Evaluation and implementation of a potential diagnostic molecular marker for H1-Conferred potato cyst nematode resistance in potato (Solanum tuberosum L.). Plant Breed. 2012, 131, 315–321. [Google Scholar] [CrossRef]
- Bayless, A.M.; Nishimura, M.T. Enzymatic functions for toll/interleukin-1 receptor domain proteins in the plant immune system. Front. Genet. 2020, 11, 539. [Google Scholar] [CrossRef]
- Leipe, D.D.; Koonin, E.V.; Aravind, L. STAND, a class of p-loop NTPases including animal and plant regulators of programmed cell death: Multiple, complex domain architectures, unusual phyletic patterns, and evolution by horizontal gene transfer. J. Mol. Biol. 2004, 343, 1–28. [Google Scholar] [CrossRef]
- Gajiwala, K.S.; Burley, S.K. Winged helix proteins. Curr. Opin. Struct. Biol. 2000, 10, 110–116. [Google Scholar] [CrossRef]
- Tameling, W.I.L.; Takken, F.L.W. Resistance proteins: Scouts of the plant innate immune system. Eur. J. Plant Pathol. 2008, 121, 243–255. [Google Scholar] [CrossRef]


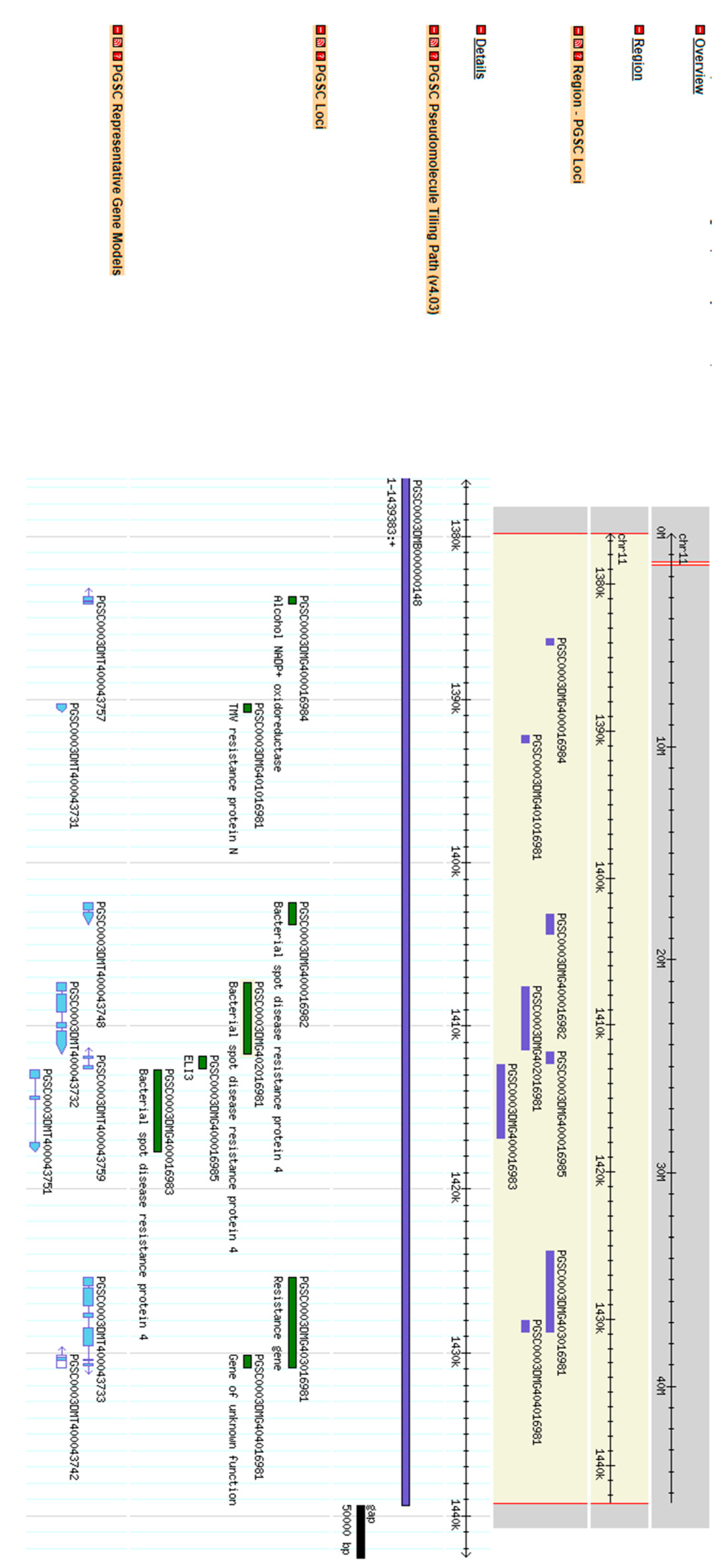{kind=link}
| Primer Name | Primer Details | Primer Sequence (5′-3′) |
|---|---|---|
| 3279_F1 | Allele specific primer 1 | (FAM-GAAGGTGACCAAGTTCATGCT)-AGCAGATATGAAGAAGAGGCG |
| 3279_F2 | Allele specific primer 2 | (VIC-GAAGGTCGGAGTCAACGGATT)-AGCAGATATGAAGAAGAGGCC |
| 3279_R | Common primer | CTATAATCCCGGGCTTGTG |
| Marker | Chromosome XI Position | Reference |
|---|---|---|
| RYSC3 (forward) | 1,826,029–1,826,051 | Kasai et al. 2001 |
| RYSC3 (reverse) | 1,826,323–1,826,346 | |
| M45 (forward) | 1,494,355–1,494,377 | Brigneti et al. 1997 |
| M45 (reverse) | 1,494,446–1,494,463 | |
| M6 (forward) | 1,708,102–1,708,121 | Herrera et al. 2018 |
| M6 (reverse) | 1,709,108–1,709,083 |
| Transcript | Position | Reference | Alternative | |
|---|---|---|---|---|
| PGSC0003DMT400043732 | 3047 | A | T 1, C | Triallelic |
| PGSC0003DMT400043732 | 3177 | T | A 1 | |
| PGSC0003DMT400043732 | 3179 | A | T, G | Triallelic |
| PGSC0003DMT400043732 | 3198 | G | C, T | Triallelic |
| PGSC0003DMT400043732 | 3279 | C | A, G 1 | Triallelic |
| Cultivar | RySC3 | M45 | SNP 3279 | PVYNTN Phenotype | PVY European Cultivated Potato Database Rating |
|---|---|---|---|---|---|
| Carlingford | - | + | + | Resistant | Low to medium (SASA 1) High (IPK 2) |
| Emma | + | + | - | Susceptible | NA |
| Friar | - | + | + | Resistant | Very high (SASA) |
| Galil | - | + | + | Resistant | NA |
| Lady Christl | - | + | + | Resistant | Very high (SASA) Very high (NIVAP 3) Very high (YN: Neiker 4) |
| PO3 | + | + | + | Resistant | NA |
| Royal Blue | - | + | + | Resistant | NA |
| Granola | - | - | + | Resistant | Low (SASA 1) High (IPK) High (YN: Neiker) |
| Suvi | - | - | + | Resistant | N/A |
| Foxton | - | - | + | Resistant | High (SASA 1) |
| Maranca | - | - | + | Resistant | N/A (Parent Impala screened as medium-high or high-very high resistance) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caruana, B.M.; Rodoni, B.C.; Constable, F.; Slater, A.T.; Cogan, N.O.I. Genome Enhanced Marker Improvement for Potato Virus Y Disease Resistance in Potato. Agronomy 2021, 11, 832. https://doi.org/10.3390/agronomy11050832
Caruana BM, Rodoni BC, Constable F, Slater AT, Cogan NOI. Genome Enhanced Marker Improvement for Potato Virus Y Disease Resistance in Potato. Agronomy. 2021; 11(5):832. https://doi.org/10.3390/agronomy11050832
Chicago/Turabian StyleCaruana, Brittney M., Brendan C. Rodoni, Fiona Constable, Anthony T. Slater, and Noel O. I. Cogan. 2021. "Genome Enhanced Marker Improvement for Potato Virus Y Disease Resistance in Potato" Agronomy 11, no. 5: 832. https://doi.org/10.3390/agronomy11050832
APA StyleCaruana, B. M., Rodoni, B. C., Constable, F., Slater, A. T., & Cogan, N. O. I. (2021). Genome Enhanced Marker Improvement for Potato Virus Y Disease Resistance in Potato. Agronomy, 11(5), 832. https://doi.org/10.3390/agronomy11050832