Morphological, Biochemical, and Metabolomic Strategies of the Date Palm (Phoenix dactylifera L., cv. Deglet Nour) Roots Response to Salt Stress

 ,
,  ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Germination Assay
2.2.1. Germination Potential (GP)
2.2.2. Germination Rate (GR)
2.2.3. Germination Speed (GS)
2.2.4. Germination Mean Time (GMT)
2.2.5. Seeds Mortality Rate (SMR)
2.3. Evaluation of Biochemical Parameters
2.3.1. Total Soluble Proteins Extraction
2.3.2. Catalase (CAT) Activity Assay
2.3.3. Superoxide Dismutase (SOD) Activity Assay
2.4. Liquid Chromatography-Mass Spectrometry (LCMS) Analysis
Sample Extraction
2.5. XCMS Online for Untargeted Metabolomics
2.6. Targeted Metabolomics
2.6.1. Data Processing
2.6.2. Dereplication Process of the Extracts for the New Hit
2.7. Statistical Methods and Data Analysis
3. Results
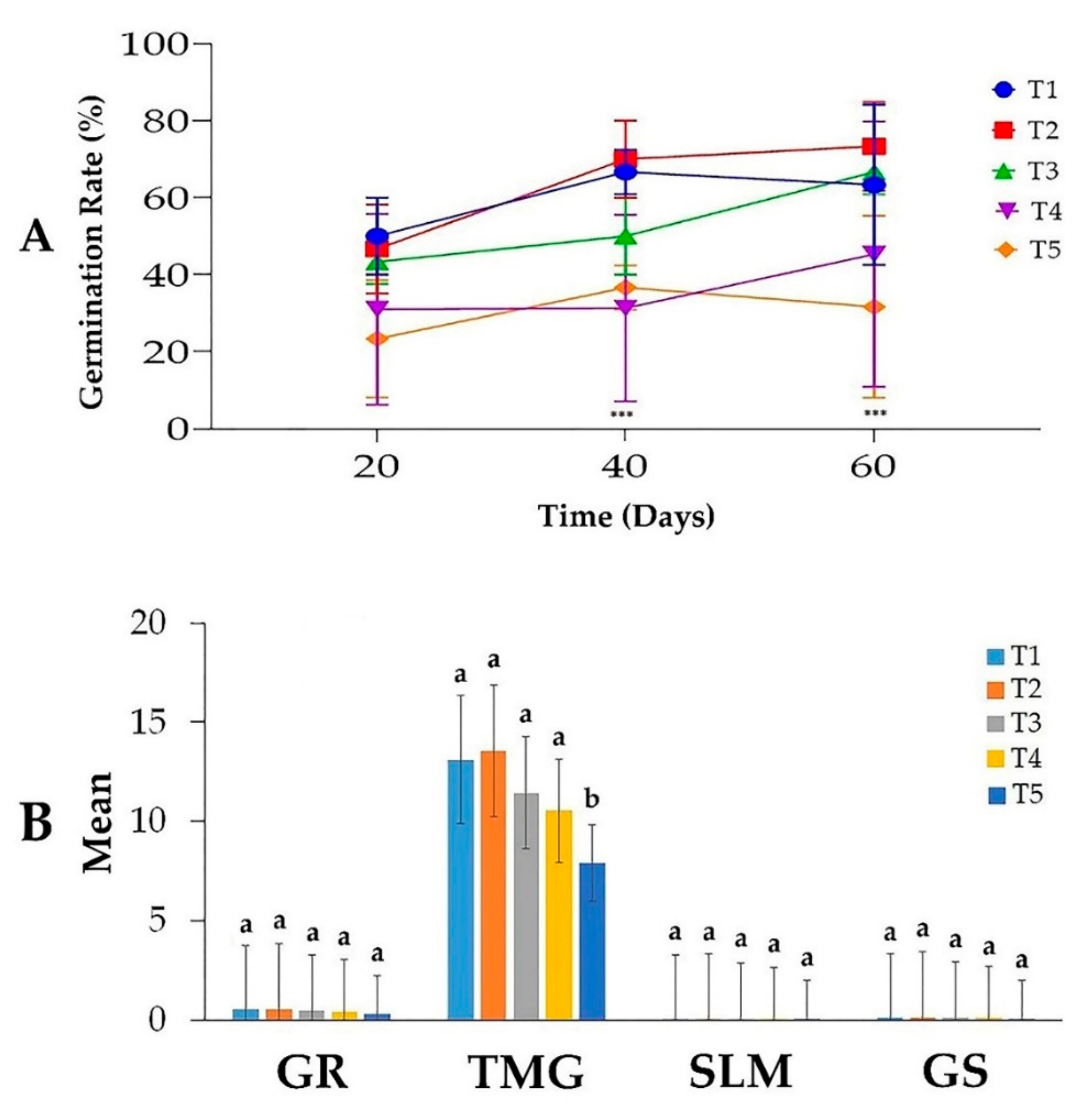
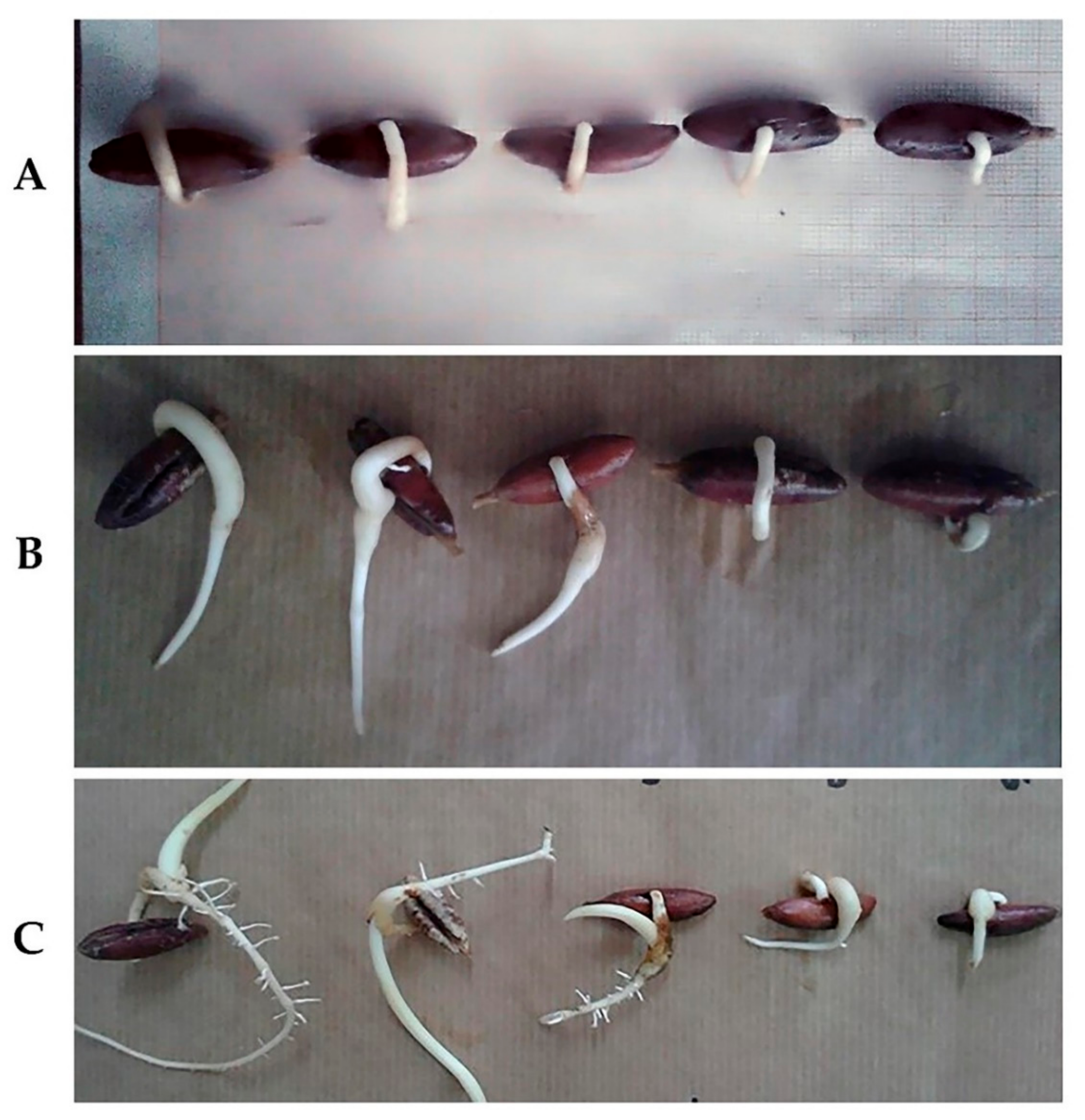
3.1. Effects of Salt Stress on Date Palm Seeds Germination
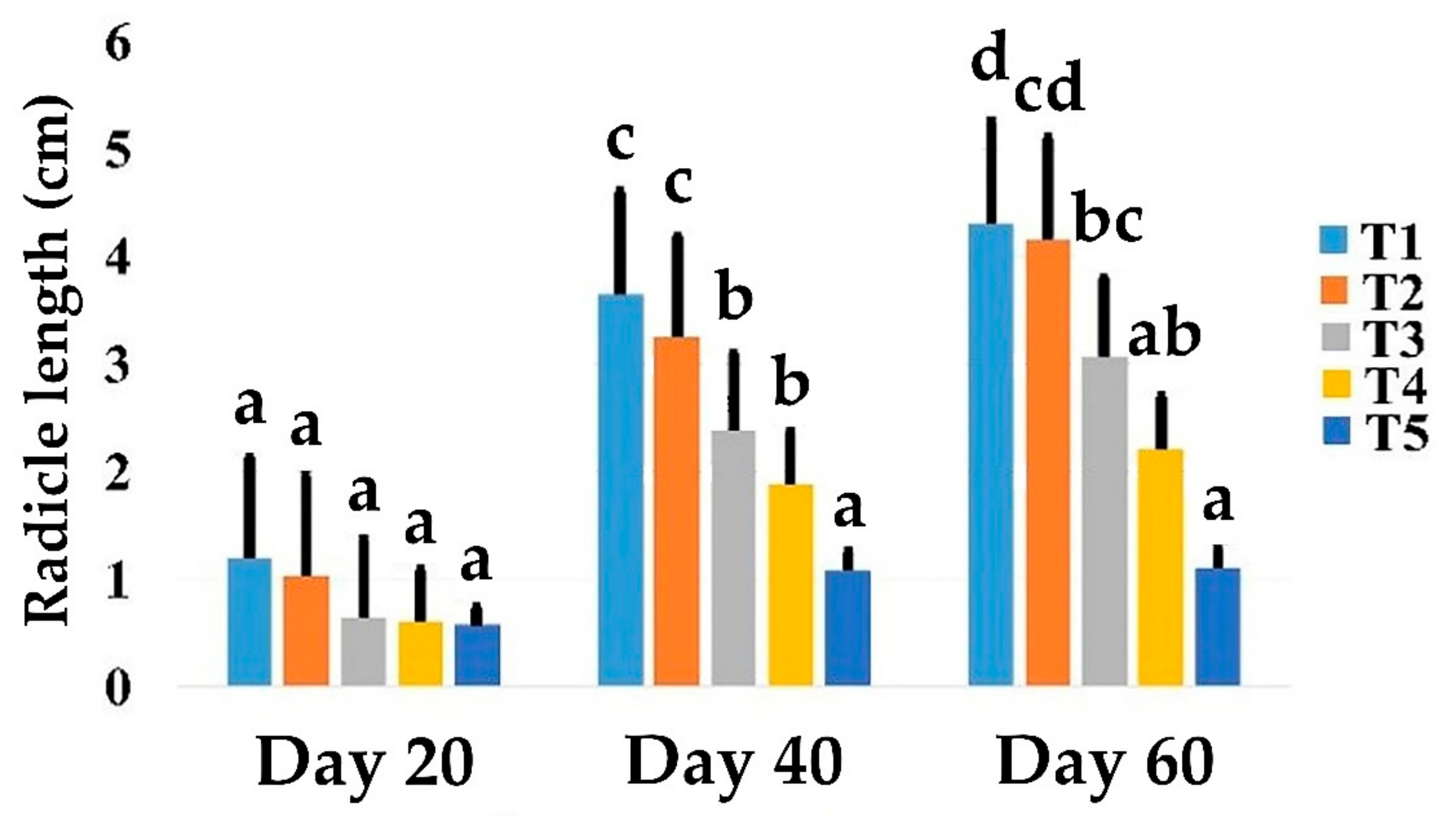
3.2. Effects of Salt Stress on Date Palm Radicle Length
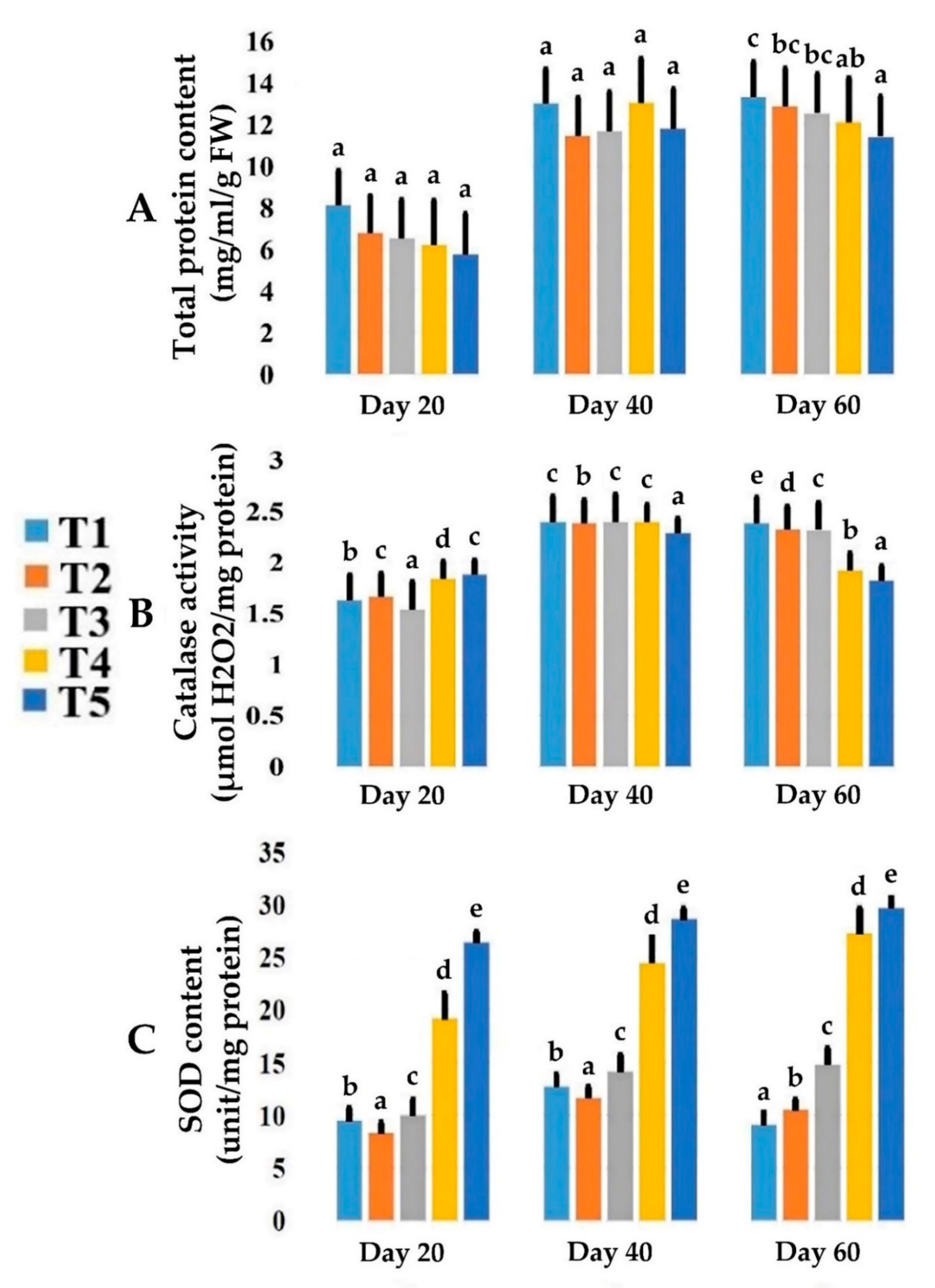
3.3. Effects of Date Palm Salt Stress on Total Proteins Content
3.4. Effects of Date Palm Salt Stress on CAT and SOD Enzymes
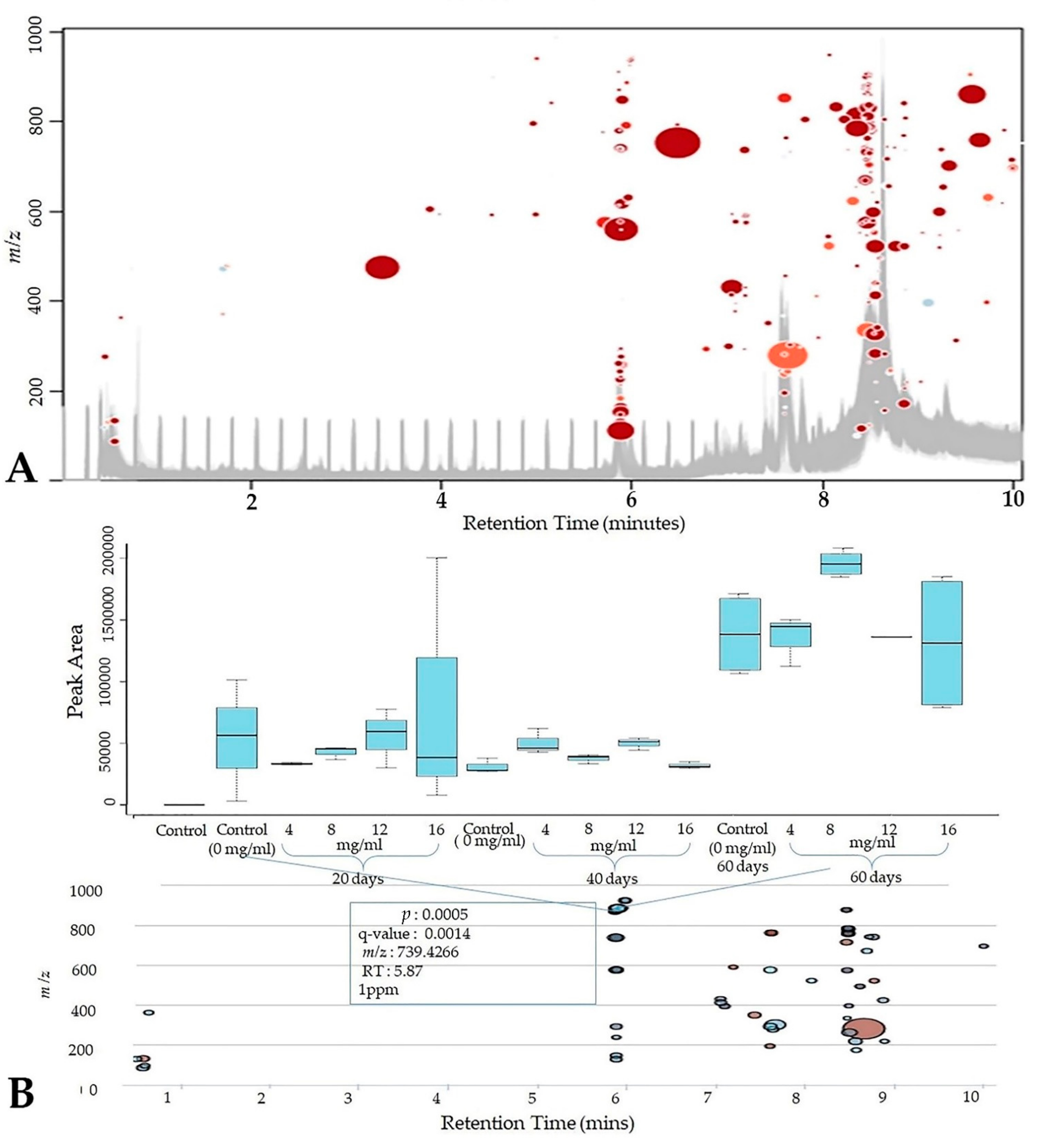
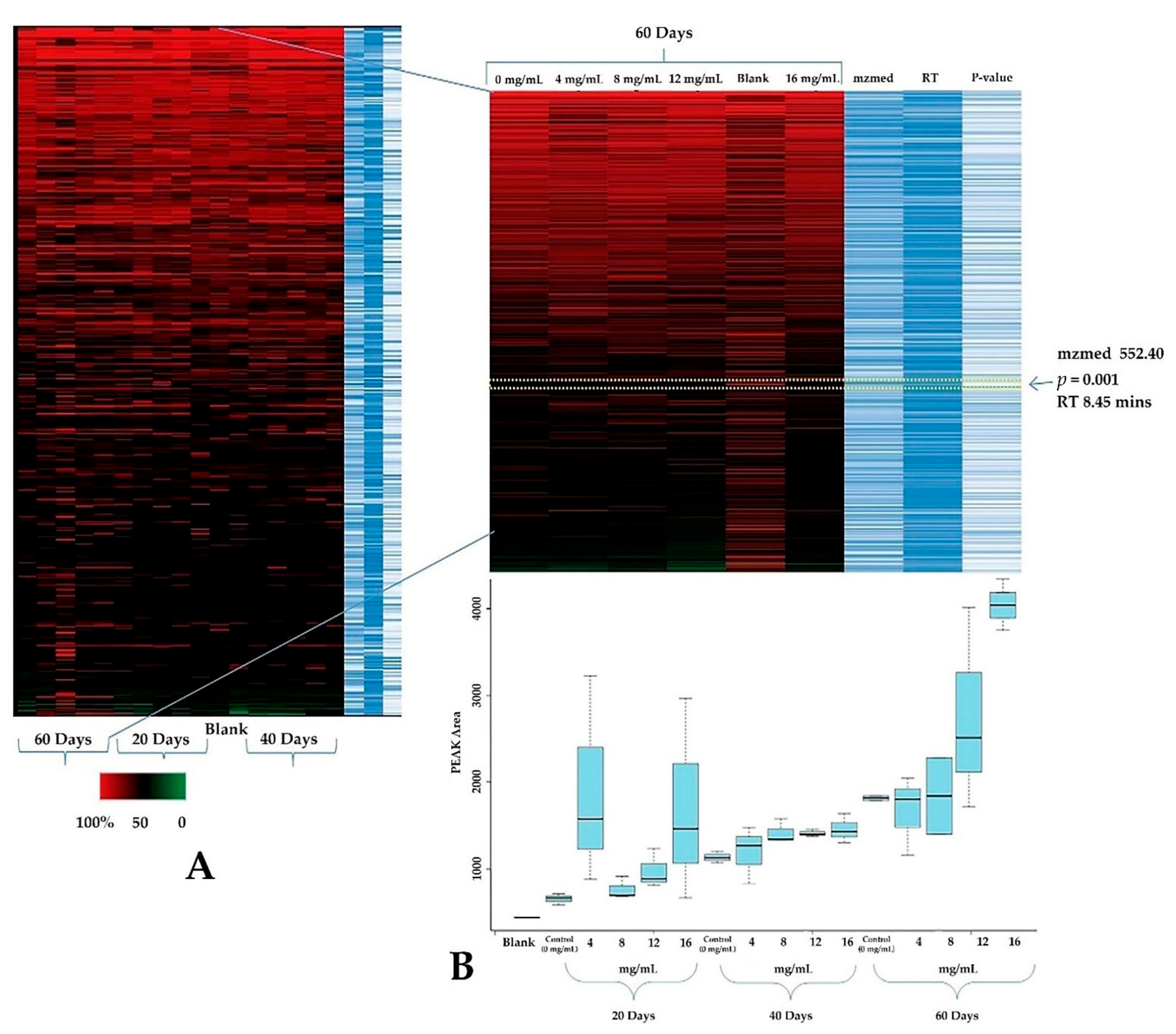
3.5. Metabolomics Profiling
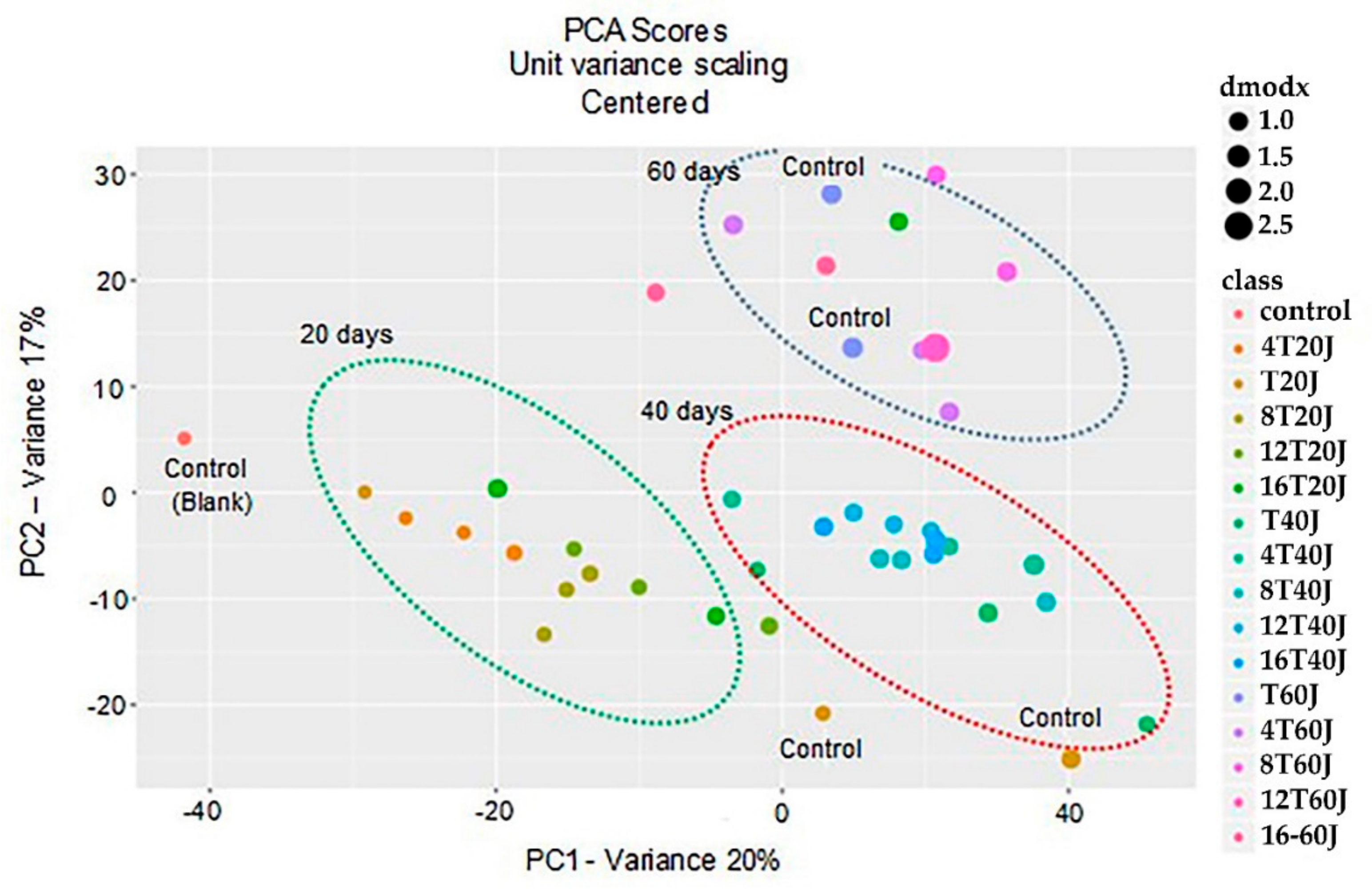
3.6. Principal Component Analysis (PCA)
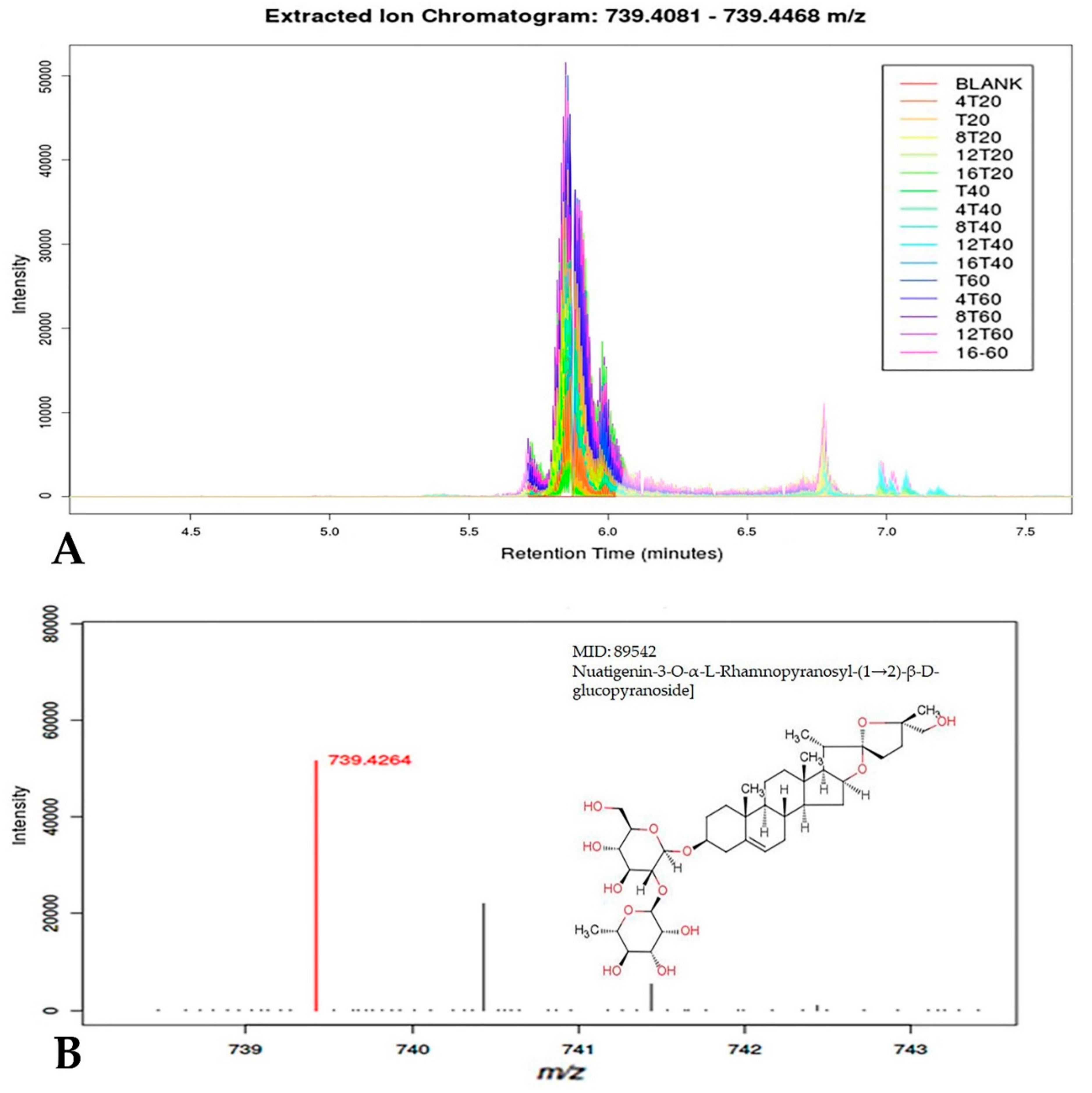
3.7. Customized Metabolomics Data
3.8. Secondary Metabolites Prediction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kerbab, S.; Silini, A.; Chenari Bouket, A.; Cherif-Silini, H.; Eshelli, M.; El Houda Rabhi, N.; Belbahri, L. Mitigation of NaCl stress in wheat by rhizosphere engineering using salt habitat adapted PGPR halotolerant bacteria. Appl. Sci. 2021, 11, 1034. [Google Scholar] [CrossRef]
- Magallon, K.J.; Dinneny, J.R. Environmental stress: Salinity ruins a plant’s day in the sun. Curr. Biol. 2019, 29, R360–R362. [Google Scholar]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar]
- Numan, M.; Bashir, S.; Khan, Y.; Mumtaz, R.; Shinwari, Z.K.; Khan, A.L.; Khan, A.; Ahmed, A.-H. Plant growth promoting bacteria as an alternative strategy for salt tolerance in plants: A review. Microbiol. Res. 2018, 209, 21–32. [Google Scholar]
- Kumar, M.; Kumar, R.; Jain, V.; Jain, S. Differential behavior of the antioxidant system in response to salinity induced oxidative stress in salt-tolerant and salt-sensitive cultivars of Brassica juncea L. Biocatal. Agric. Biotechnol. 2018, 13, 12–19. [Google Scholar]
- Gondek, M.; Weindorf, D.C.; Thiel, C.; Kleinheinz, G. Soluble salts in compost and their effects on soil and plants: A review. Compost Sci. Util. 2020, 28, 59–75. [Google Scholar]
- Alhammadi, M.S.; Kurup, S.S. Impact of salinity stress on date palm (Phoenix Dactylifera L.)—A review. In Crop Production Technologies; Sharma, P., Abrol, V., Eds.; Books on Demand: Norderstedt, Germany, 2012; pp. 169–173. [Google Scholar]
- Du, B.; Kreuzwieser, J.; Winkler, J.B.; Ghirardo, A.; Schnitzler, J.-P.; Ache, P.; Alfarraj, S.; Hedrich, R.; White, P.; Rennenberg, H. Physiological responses of date palm (Phoenix dactylifera) seedlings to acute ozone exposure at high temperature. Environ. Pollut. 2018, 242, 905–913. [Google Scholar]
- Safronov, O.; Kreuzwieser, J.; Haberer, G.; Alyousif, M.S.; Schulze, W.; Al-Harbi, N.; Arab, L.; Ache, P.; Stempfl, T.; Kruse, J. Detecting early signs of heat and drought stress in Phoenix dactylifera (date palm). PLoS ONE 2017, 12, e0177883. [Google Scholar]
- Saghafi, D.; Delangiz, N.; Lajayer, B.A.; Ghorbanpour, M. An overview on improvement of crop productivity in saline soils by halotolerant and halophilic PGPRs. 3 Biotech 2019, 9, 1–14. [Google Scholar]
- Hazzouri, K.M.; Flowers, J.M.; Nelson, D.; Lemansour, A.; Masmoudi, K.; Amiri, K.M.A. Prospects for the study and improvement of abiotic stress tolerance in date palms in the post-genomics era. Front. Plant Sci. 2020, 11, 293. [Google Scholar] [CrossRef]
- Slama, H.; Cherif-Silini, H.; Chenari Bouket, A.; Qader, M.; Silini, A.; Yahiaoui, B.; Alenezi, F.; Luptakova, L.; Triki, M.; Vallat, A.; et al. Screening for Fusarium antagonistic bacteria from contrasting niches designated the endophyte Bacillus halotolerans as plant warden against Fusarium. Front. Microbiol. 2019, 9, 3236. [Google Scholar] [CrossRef] [Green Version]
- Munier, P. Le Palmier-Dattier-Techniques Agricoles et Productions Tropicales; Maison Neuve et Larose: Paris, France, 1973; p. 217. [Google Scholar]
- El-Juhany, L.I. Degradation of date palm trees and date production in arab countries: Causes and potential rehabilitation. Australian J. Basic Appl. Sci. 2010, 4, 3998–4010. [Google Scholar]
- El-Far, A.H.; Oyinloye, B.E.; Sepehrimanesh, M.; Allah, M.A.G.; Abu-Reidah, I.; Shaheen, H.M.; Razeghian-Jahromi, I.; Noreldin, A.E.; Al Jaouni, S.K.; Mousa, S.A. Date palm (Phoenix Dactylifera): Novel findings and future directions for food and drug discovery. Curr. Drug Discov. Technol. 2019, 16, 2–10. [Google Scholar]
- Johnson, D.V.; Al-Khayri, J.M.; Jain, S.M. Introduction: Date production status and prospects in Africa and the Americas. In Date Palm Genetic Resources and Utilization; Springer: Berlin/Heidelberg, Germany, 2015; pp. 3–18. [Google Scholar]
- Ramoliya, P.J.; Pandey, A.N. Soil salinity and water status affect growth of Phoenix dactylifera seedlings. N. Z. J. Crop Hort. Sci. 2003, 31, 345–353. [Google Scholar]
- Al Kharusi, L.; Al Yahyai, R.; Yaish, M.W. Antioxidant response to salinity in salt-tolerant and salt-susceptible cultivars of date palm. Agriculture 2019, 9, 8. [Google Scholar]
- El Rabey, H.A.; Al-Malki, A.L.; Abulnaja, K.O. Proteome analysis of date palm (Phoenix dactylifera L.) under severe drought and salt stress. Int. J. Genom. 2016, 2016, 7840759. [Google Scholar]
- Al-Muaini, A.; Green, S.; Dakheel, A.; Abdullah, A.-H.; Abou Dahr, W.A.; Dixon, S.; Kemp, P.; Clothier, B. Irrigation management with saline groundwater of a date palm cultivar in the hyper-arid united Arab Emirates. Agric. Water Manag. 2019, 211, 123–131. [Google Scholar]
- Anli, M.; Baslam, M.; Tahiri, A.; Raklami, A.; Symanczik, S.; Boutasknit, A.; Ait-El-Mokhtar, M.; Ben-Laouane, R.; Toubali, S.; Rahou, Y.A. Biofertilizers as strategies to improve photosynthetic apparatus, growth, and drought stress tolerance in the date palm. Front. Plant Sci. 2020, 11, 516818. [Google Scholar] [CrossRef]
- Ben Mefteh, F.; Daoud, A.; Chenari Bouket, A.; Thissera, B.; Kadri, Y.; Cherif-Silini, H.; Eshelli, M.; Alenezi, F.N.; Vallat, A.; Oszako, T. Date palm trees root-derived endophytes as fungal cell factories for diverse bioactive metabolites. Int. J. Mol. Sci. 2018, 19, 1986. [Google Scholar]
- Daoud, A.; Malika, D.; Bakari, S.; Hfaiedh, N.; Mnafgui, K.; Kadri, A.; Gharsallah, N. Assessment of polyphenol composition, antioxidant and antimicrobial properties of various extracts of date palm pollen (DPP) from two Tunisian cultivars. Arabian J. Chem. 2019, 12, 3075–3086. [Google Scholar]
- Darwesh, R.S. Improving growth of date palm plantlets grown under salt stress with yeast and amino acids applications. Ann. Agric. Sci. 2013, 58, 247–256. [Google Scholar]
- Sperling, O.; Lazarovitch, N.; Schwartz, A.; Shapira, O. Effects of high salinity irrigation on growth, gas-exchange, and photoprotection in date palms (Phoenix dactylifera L., Cv. Medjool). Environ. Exp. Bot. 2014, 99, 100–109. [Google Scholar]
- Alfarrah, N.; Walraevens, K. Groundwater overexploitation and seawater intrusion in coastal areas of arid and semi-arid regions. Water 2018, 10, 143. [Google Scholar]
- Ait-El-Mokhtar, M.; Baslam, M.; Ben-Laouane, R.; Anli, M.; Boutasknit, A.; Mitsui, T.; Wahbi, S.; Meddich, A. Alleviation of detrimental effects of salt stress on date palm (Phoenix dactylifera L.) by the application of arbuscular mycorrhizal fungi and/or compost. Front. Sustain. Food Syst. 2020, 4, 131. [Google Scholar] [CrossRef]
- Rodziewicz, P.; Swarcewicz, B.; Chmielewska, K.; Wojakowska, A.; Stobiecki, M. Influence of abiotic stresses on plant proteome and metabolome changes. Acta Physiol. Plant 2014, 36, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Al Kharusi, L.; Jana, G.A.; Patankar, H.V.; Yaish, M.W. Comparative metabolic profiling of two contrasting date palm genotypes under salinity. Plant Mol. Biol. Rep. 2020, 39, 351–363. [Google Scholar]
- Rekik, I.; Chaabane, Z.; Missaoui, A.; Bouket, A.C.; Luptakova, L.; Elleuch, A.; Belbahri, L. Effects of untreated and treated wastewater at the morphological, physiological and biochemical levels on seed germination and development of sorghum (Sorghum bicolor (L.) Moench), alfalfa (Medicago sativa L.) and fescue (Festuca arundinacea Schreb.). J. Hazard Mater. 2017, 326, 165–176. [Google Scholar]
- Bradford, M.M. A Rapid and Sensitive Method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar]
- Aebi, H. [13] Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar]
- Tautenhahn, R.; Patti, G.J.; Rinehart, D.; Siuzdak, G. XCMS Online: A web-based platform to process untargeted metabolomic data. Anal. Chem. 2012, 84, 5035–5039. [Google Scholar] [CrossRef] [Green Version]
- Rekik, I.; Chaabane, Z.; Grubb, C.D.; Drira, N.; Cheour, F.; Elleuch, A. In silico characterization and Molecular modeling of double-strand break repair protein MRE11 from Phoenix dactyliferav deglet nour. Theor. Biol. Med. Model. 2015, 12, 23. [Google Scholar] [CrossRef] [Green Version]
- Haj-Amor, Z.; Ibrahimi, M.-K.; Feki, N.; Lhomme, J.-P.; Bouri, S. Soil salinisation and irrigation management of date palms in a saharan environment. Environ. Monit. Assess. 2016, 188, 1–17. [Google Scholar]
- Youssef, T.; Awad, M.A. Mechanisms of enhancing photosynthetic gas exchange in date palm seedlings (Phoenix dactylifera L.) under salinity stress by a 5-aminolevulinic acid-based fertilizer. J. Plant Growth Regul. 2008, 27, 1–9. [Google Scholar]
- Yaish, M.W.; Kumar, P.P. Salt tolerance research in date palm tree (Phoenix dactylifera L.), past, present, and future perspectives. Front. Plant Sci. 2015, 6, 348. [Google Scholar] [CrossRef] [Green Version]
- Elshibli, S.; Elshibli, E.M.; Korpelainen, H. Growth and photosynthetic CO2 responses of date palm plants to water availability. Emir. J. Food Agric. 2016, 28, 58–65. [Google Scholar] [CrossRef]
- Müller, H.M.; Schäfer, N.; Bauer, H.; Geiger, D.; Lautner, S.; Fromm, J.; Riederer, M.; Bueno, A.; Nussbaumer, T.; Mayer, K. The desert plant Phoenix dactylifera closes stomata via nitrate-regulated SLAC 1 anion channel. New Phytol. 2017, 216, 150–162. [Google Scholar]
- Zai, X.-M.; Fan, J.-J.; Hao, Z.-P.; Liu, X.-M.; Zhang, W.-X. Effect of co-inoculation with arbuscular mycorrhizal fungi and phosphate solubilizing fungi on nutrient uptake and photosynthesis of beach palm under salt stress environment. Sci. Rep. 2021, 11, 5761. [Google Scholar] [CrossRef]
- Hussain, N.; Al-Rasbi, S.; Al-Wahaibi, N.S.; Al-Ghanum, G.; El-Sharief Abdalla, O.A. Salinity problems and their management in date palm production. In Dates: Production, Processing, Food, and Medicinal Value; Manickavasagan, A., Mohamed Essa, M., Sukumar, E., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 88–108. [Google Scholar]
- Alrasbi, S.A.R.; Hussain, N.; Schmeisky, H. Evaluation of the growth of date palm seedlings irrigated with saline water in the Sultanate of Oman. In Proceedings of the IV International Date Palm Conference, Abu Dhabi, UAE, 15–17 March 2010; pp. 233–246. [Google Scholar]
- Yaish, M.W. Proline accumulation is a general response to abiotic stress in the date palm tree (Phoenix dactylifera L.). Genet. Mol. Res. 2015, 14, 9943–9950. [Google Scholar]
- Khudairi, A.K. Studies on the germination of date-palm seeds. The effect of sodium chloride. Physiol. Plant. 1958, 11, 16–22. [Google Scholar]
- Aljuburi, H.V. Effect of Sodium Chloride on Seedling Growth of Four Date Palm Varieties. Ann. Arid Zone 1992, 31, 259–262. [Google Scholar]
- Furr, J.R.; Ream, C.L. Salinity effects on growth and salt uptake of seedlings of the date, Phoenix dactylifera L. In Procceedings of American Society of Horticultural Sciences; American Society of Horticultural Sciences: Alexandria, VA, USA, 1968; Volume 92, pp. 268–273. [Google Scholar]
- Hewitt, A.A. Effect of different salts and salt concentration on the germination and subsequent growth of deglet Noor date seeds. Date Growers’ Inst. Rept. 1963, 40, 4–6. [Google Scholar]
- Xiao, T.T.; Raygoza, A.A.; Pérez, J.C.; Kirschner, G.; Deng, Y.; Atkinson, B.; Sturrock, C.; Lube, V.; Wang, J.Y.; Lubineau, G. Emergent protective organogenesis in date palms: A morpho-devo-dynamic adaptive strategy during early development. Plant Cell. 2019, 31, 1751–1766. [Google Scholar]
- Verslues, P.E.; Agarwal, M.; Katiyar-Agarwal, S.; Zhu, J.; Zhu, J.-K. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J. 2006, 45, 523–539. [Google Scholar]
- Flowers, T.J.; Colmer, T.D. Plant Salt Tolerance: Adaptations in halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar]
- Hanin, M.; Ebel, C.; Ngom, M.; Laplaze, L.; Masmoudi, K. New insights on plant salt tolerance mechanisms and their potential use for breeding. Front. Plant Sci. 2016, 7, 1787. [Google Scholar]
- You, J.; Chan, Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar]
- Dreyer, A.; Dietz, K.-J. Reactive oxygen species and the redox-regulatory network in cold stress acclimation. Antioxidants 2018, 7, 169. [Google Scholar]
- Foyer, C.H. Reactive oxygen species, oxidative signaling and the regulation of photosynthesis. Environ. Exp. Bot. 2018, 154, 134–142. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Oxidant and antioxidant signalling in plants: A re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ. 2005, 28, 1056–1071. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar]
- Huang, H.; Ullah, F.; Zhou, D.-X.; Yi, M.; Zhao, Y. Mechanisms of ROS regulation of plant development and stress responses. Front. Plant Sci. 2019, 10, 800. [Google Scholar]
- Mohammadi, Z.; Rastegar, S.; Abdollahi, F.; Hosseini, Y. Morphological and antioxidant enzymatic activity responses of sapodilla rootstock to salinity stress. J. Plant Process Funct. 2018, 6, 24. [Google Scholar]
- Qiu-Fang, Z.; Yuan-Yuan, L.; Cai-Hong, P.; Cong-Ming, L.; Bao-Shan, W. NaCl enhances thylakoid-bound SOD activity in the leaves of C3 halophyte Suaeda salsa L. Plant Sci. 2005, 168, 423–430. [Google Scholar]
- Ye, Y.; Gu, Y.T.; Gao, H.Y.; Lu, C.Y. Combined effects of simulated tidal sea-level rise and salinity on seedlings of a mangrove species, Kandelia candel (L.) Druce. Hydrobiologia 2010, 641, 287–300. [Google Scholar]
- Wang, N.; Qiao, W.; Liu, X.; Shi, J.; Xu, Q.; Zhou, H.; Yan, G.; Huang, Q. Relative contribution of Na+/K+ homeostasis, photochemical efficiency and antioxidant defense system to differential salt tolerance in cotton (Gossypium hirsutum L.) cultivars. Plant Physiol. Biochem. 2017, 119, 121–131. [Google Scholar]
- Du, B.; Ma, Y.; Yáñez-Serrano, A.M.; Arab, L.; Fasbender, L.; Alfarraj, S.; Albasher, G.; Hedrich, R.; White, P.J.; Werner, C.; et al. Physiological responses of date palm (Phoenix dactylifera) seedlings to seawater and flooding. New Phytol. 2021, 229, 3318–3329. [Google Scholar] [CrossRef]
- Patankar, H.V.; Al-Harrasi, I.; Al Kharusi, L.; Jana, G.A.; Al-Yahyai, R.; Sunkar, R.; Yaish, M.W. Overexpression of a metallothionein 2A gene from date palm confers abiotic stress tolerance to yeast and Arabidopsis thaliana. Int. J. Mol. Sci. 2019, 20, 2871. [Google Scholar]
- Diboun, I.; Mathew, S.; Al-Rayyashi, M.; Elrayess, M.; Torres, M.; Halama, A.; Méret, M.; Mohney, R.P.; Karoly, E.D.; Malek, J.; et al. Metabolomics of dates (Phoenix dactylifera) reveals a highly dynamic ripening process accounting for major variation in fruit composition. BMC Plant Biol. 2015, 15, 291. [Google Scholar] [CrossRef] [Green Version]
- Jana, G.A.; Kharusi, L.A.; Sunkar, R.; Al-Yahyai, R.; Yaish, M.W. Metabolomic analysis of date palm seedlings exposed to salinity and silicon treatments. Plant Signal. Behav. 2019, 14, 1663112. [Google Scholar] [CrossRef]
- Abdul-Hamid, N.A.; Abas, F.; Ismail, I.S.; Tham, C.L.; Maulidiani, M.; Mediani, A.; Swarup, S.; Umashankar, S.; Zolkeflee, N.K.Z. Metabolites and biological activities of Phoenix dactylifera L. pulp and seeds: A comparative MS and NMR based metabolomics approach. Phytochem. Lett. 2019, 31, 20–32. [Google Scholar]
- Hamad, I.; AbdElgawad, H.; Al Jaouni, S.; Zinta, G.; Asard, H.; Hassan, S.; Hegab, M.; Hagagy, N.; Selim, S. Metabolic analysis of various date palm fruit (Phoenix dactylifera L.) cultivars from Saudi Arabia to assess their nutritional quality. Molecules 2015, 20, 13620–13641. [Google Scholar]
- Al-Alawi, R.A.; Al-Mashiqri, J.H.; Al-Nadabi, J.S.M.; Al-Shihi, B.I.; Baqi, Y. Date palm tree (Phoenix dactylifera L.): Natural products and therapeutic options. Front. Plant Sci. 2017, 8, 845. [Google Scholar] [CrossRef] [Green Version]
- Abdul-Hamid, N.A.; Mustaffer, N.H.; Maulidiani, M.; Mediani, A.; Ismail, I.S.; Tham, C.L.; Shadid, K.; Abas, F. Quality evaluation of the physical properties, phytochemicals, biological activities and proximate analysis of nine Saudi date palm fruit varieties. J. Saudi Soc. Agric. Sci. 2020, 19, 151–160. [Google Scholar] [CrossRef]
- Wu, S.-B.; Meyer, R.S.; Whitaker, B.D.; Litt, A.; Kennelly, E.J. A new liquid chromatography–mass spectrometry-based strategy to integrate chemistry, morphology, and evolution of eggplant (Solanum) species. J. Chromatogr. A. 2013, 1314, 154–172. [Google Scholar]
- Akula, R.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar]
- Bistgani, Z.E.; Hashemi, M.; DaCosta, M.; Craker, L.; Maggi, F.; Morshedloo, M.R. Effect of salinity stress on the physiological characteristics, phenolic compounds and antioxidant activity of Thymus vulgaris L. and Thymus daenensis Celak. Indust. Crops Prod. 2019, 135, 311–320. [Google Scholar]
- Hassini, I.; Rios, J.J.; Garcia-Ibañez, P.; Baenas, N.; Carvajal, M.; Moreno, D.A. Comparative effect of elicitors on the physiology and secondary metabolites in broccoli plants. J. Plant Physiol. 2019, 239, 1–9. [Google Scholar]
- Pappalardo, H.D.; Toscano, V.; Puglia, G.D.; Genovese, C.; Raccuia, S.A. Cynara cardunculus L. as a multipurpose crop for plant secondary metabolites production in marginal stressed lands. Front. Plant Sci. 2020, 11, 240. [Google Scholar] [CrossRef]
- Hadrami, A.E.; Daayf, F.; Hadrami, I.E. Secondary metabolites of date palm. In Date Palm Biotechnology; Jain, S.M., Al-Khayri, J.M., Johnson, D.V., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 653–674. [Google Scholar]
- Ashraf, M.A.; Iqbal, M.; Rasheed, R.; Hussain, I.; Riaz, M.; Arif, M.S. Environmental stress and secondary metabolites in plants: An overview. In Plant Metabolites and Regulation under Environmental Stress; Ahmad, P., Ahanger, M.A., Singh, V.P., Tripathi, D.K., Alam, P., Alyemeni, M.N., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 153–167. [Google Scholar]
- Bartwal, A.; Mall, R.; Lohani, P.; Guru, S.K.; Arora, S. Role of secondary metabolites and brassinosteroids in plant defense against environmental stresses. J. Plant Growth Regul. 2013, 32, 216–232. [Google Scholar] [CrossRef]
- Rienth, M.; Vigneron, N.; Darriet, P.; Sweetman, C.; Burbidge, C.; Bonghi, C.; Walker, R.P.; Famiani, F.; Castellarin, S.D. Grape berry secondary metabolites and their modulation by abiotic factors in a climate change scenario—A review. Front. Plant Sci. 2021, 12, 262. [Google Scholar] [CrossRef]
- Watts, E.J.; Shen, Y.; Lansky, E.P.; Nevo, E.; Bobe, G.; Traber, M.G. High environmental stress yields greater tocotrienol content while changing vitamin E profiles of wild emmer wheat seeds. J. Med. Food 2015, 18, 216–223. [Google Scholar] [CrossRef]
- Sattler, S.E.; Mène-Saffrané, L.; Farmer, E.E.; Krischke, M.; Mueller, M.J.; DellaPenna, D. Nonenzymatic lipid peroxidation reprograms gene expression and activates defense markers in Arabidopsis tocopherol-deficient mutants. Plant Cell. 2006, 18, 3706–3720. [Google Scholar]
- Cela, J.; Chang, C.; Munné-Bosch, S. Accumulation of γ- rather than α-tocopherol alters ethylene signaling gene expression in the Vte4 mutant of Arabidopsis thaliana. Plant Cell Physiol. 2011, 52, 1389–1400. [Google Scholar] [CrossRef] [Green Version]
- Yusuf, M.A.; Kumar, D.; Rajwanshi, R.; Strasser, R.J.; Tsimilli-Michael, M.; Sarin, N.B. Overexpression of γ-tocopherol methyl transferase gene in transgenic Brassica juncea plants alleviates abiotic stress: Physiological and chlorophyll a fluorescence measurements. Biochim. Biophys. Acta Bioenerg. BBA-Bioenerg. 2010, 1797, 1428–1438. [Google Scholar]
- Awad, A.M.; Soaud, A.A.; El-Konaissi, M.S.; Zaid, A.; Eshkandi, O.H.; Badawi, M.A. Effect of elemental sulfur, some antioxidants and growth regulators on tolerance ability of in-vitro produced plantlets, and nutrient uptake, yield and fruit quality of mature date palm trees–Part I. Tolerance ability of in-vitro produced plantlets. In Proceedings of the 6th Annual Research Conference at UAE University, Al Ain, UAE, 24–26 April 2005; pp. 24–26. [Google Scholar]
- Hou, Q.; Ufer, G.; Bartels, D. Lipid signalling in plant responses to abiotic stress. Plant Cell Environ. 2016, 39, 1029–1048. [Google Scholar] [CrossRef]
- Sui, N.; Han, G. Salt-induced photoinhibition of PSII is alleviated in halophyte Thellungiella halophila by increases of unsaturated fatty acids in membrane lipids. Acta Physiol. Plant. 2014, 36, 983–992. [Google Scholar]
- Chaudhary, S.; Chaudhary, P.S.; Chikara, S.K.; Sharma, M.C.; Iriti, M. Review on fenugreek (Trigonella foenum-graecum L.) and its important secondary metabolite diosgenin. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 46, 22–31. [Google Scholar]
- Wu, B.; Xiao, X.; Li, S.; Zuo, G. Transcriptomics and metabonomics of the anti-aging properties of total flavones of epimedium in relation to lipid metabolism. J. Ethnopharmacol. 2019, 229, 73–80. [Google Scholar]
- Riaz, A.; Rasul, A.; Kanwal, N.; Hussain, G.; Shah, M.A.; Sarfraz, I.; Ishfaq, R.; Batool, R.; Rukhsar, F.; Adem, Ş. Germacrone: A potent secondary metabolite with therapeutic potential in metabolic diseases, cancer and viral infections. Curr. Drug Metab. 2020, 21, 1079–1090. [Google Scholar]
- Hanen, F.; Ksouri, R.; Megdiche, W.; Trabelsi, N.; Boulaaba, M.; Abdelly, C. Effect of salinity on growth, leaf-phenolic content and antioxidant scavenging activity in Cynara cardunculus L. In Biosaline Agriculture and High Salinity Tolerance; Springer: Berlin/Heidelberg, Germany, 2008; pp. 335–343. [Google Scholar]
- Jing-Yan, W.; Zhao-Pu, L.I.U. Alkaloid accumulation in Catharanthus roseus increases with addition of seawater salts to the nutrient solution. Pedosphere 2010, 20, 718–724. [Google Scholar]
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q. Response of plant secondary metabolites to environmental factors. Molecules 2018, 23, 762. [Google Scholar]
- Yasmin, H.; Naeem, S.; Bakhtawar, M.; Jabeen, Z.; Nosheen, A.; Naz, R.; Keyani, R.; Mumtaz, S.; Hassan, M.N. Halotolerant rhizobacteria Pseudomonas pseudoalcaligenes and Bacillus subtilis mediate systemic tolerance in hydroponically grown soybean (Glycine max L.) against salinity stress. PLoS ONE 2020, 15, e0231348. [Google Scholar] [CrossRef]
- Hassan, A.; Amjad, S.F.; Saleem, M.H.; Yasmin, H.; Imran, M.; Riaz, M.; Joyia, F.A.; Mobeen; Ahmed, S.; Ali, S.; et al. Foliar application of ascorbic acid enhances salinity stress tolerance in barley (Hordeum vulgare L.) through modulation of morpho-physio-biochemical attributes, ions uptake, osmo-protectants and stress response genes expression. Saudi J. Biol. Sci. 2021, 28, 4276–4290. [Google Scholar]
- Ilyas, N.; Mazhar, R.; Yasmin, H.; Khan, W.; Iqbal, S.; Enshasy, H.E.; Dailin, D.J. Rhizobacteria Isolated from Saline Soil Induce Systemic Tolerance in Wheat (Triticum aestivum L.) against Salinity Stress. Agronomy 2020, 10, 989. [Google Scholar]
- Jabeen, Z.; Fayyaz, H.A.; Irshad, F.; Hussain, N.; Hassan, M.N.; Li, J.; Rehman, S.; Haider, W.; Yasmin, H.; Mumtaz, S.; et al. Sodium nitroprusside application improves morphological and physiological attributes of soybean (Glycine max L.) under salinity stress. PLoS ONE 2021, 16, e0248207. [Google Scholar] [CrossRef]
- Meddich, A.; El Mokhtar, M.A.; Bourzik, W.; Mitsui, T.; Baslam, M.; Hafidi, M. Optimizing growth and tolerance of date palm (Phoenix dactylifera L.) to drought, salinity, and vascular fusarium-induced wilt (Fusarium oxysporum) by application of arbuscular mycorrhizal fungi (AMF). In Root Biology; Springer: Berlin/Heidelberg, Germany, 2018; pp. 239–258. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| * Rt | † HRESIMS | [M + H/Na]+ | Tentative Identification |
|---|---|---|---|
| 5.85 | 739.4262 | C39H63O13 | Nuatigenin-O-[α-L-Rhamnopyranosyl-(1→2)-β-D-glucopyranoside] (terpenoid) |
| 5.87 | 885.4833 | C45H73O17 | Nuatigenin − 3-O-[α-L-Rhamnopyranosyl-(1→2)-[α-L-rhamnopyranosyl-(1→4)]-β-D-glucopyranoside] (terpenoid) |
| 7.58 | 415.3215 | C27H43O3 | Diosgenin (terpenoid) |
| 7.60 | 397.3109 | C27H41O2 | δ-tocotrienol (terpenoid) |
| 8.40 | 280.2648 | C18H34NO | Linoleamide (fatty acid derivative) |
| 8.48 | 552.4000 | Unknown | |
| 8.46 | 715.5225 | C45H72O5Na | Belamcandaquinone L (Phenolic derivative) |
| 8.47 | 352.3221 | C22H42NO2 | Unknown terpenoid derivative |
| 8.47 | 279.2084 | C16H27N2O2 | Lycocernuine (alkaloid) |
| 8.48 | 758.5683 | C42H81NO8P | L-α-phosphatidylcholine (Glycerophospholipid) |
| 8.48 | 782.5677 | C44H81NO8P | Dilinoleoylphosphatydylcholine (Glycerophospholipid) |
| 8.50 | 784.5819 | C44H83NO8P | α-Oleyl-β-linoleoylphosphatydylcholine (Glycerophospholipid) |
| 8.52 | 614.4843 | C35H65N3O4Na | Unknown |
| 8.54 | 570.4582 | C35H60N3O3 | Unknown terpenoid derivative |
| 8.55 | 526.4322 | C31H57N3O2Na | Unknown |
| 8.56 | 219.1759 | C15H23O | Germacrone (terpenoid) |
| 8.61 | 429.3736 | C29H49O2 | Cholesteryl acetate (terpenoid) |
| 8.61 | 256.2649 | C16H34NO | Palmitamide (fatty acid derivative) |
| 8.64 | 282.2804 | C18H36NO | Oleamide (fatty acid derivative) |
| 8.65 | 336.3273 | C22H42NO | Pipericine (fatty acid derivative) |
| 8.67 | 742.5819 | C42H80NO9 | Asteriacerebroside G (cerebrosides) |
| 8.96 | 284.2939 | C18H38NO | Stearamide (fatty acid derivative) |
| 9.23 | 708.5113 | C37H71N3O8Na | Unknown |
| 9.26 | 663.4533 | C40H64O6Na | Scapaundulin B (terpenoid) |
| 9.31 | 338.3425 | C22H44NO | Cis-docosenamide (fatty acid derivative) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouhouch, S.; Eshelli, M.; Slama, H.B.; Bouket, A.C.; Oszako, T.; Okorski, A.; Rateb, M.E.; Belbahri, L. Morphological, Biochemical, and Metabolomic Strategies of the Date Palm (Phoenix dactylifera L., cv. Deglet Nour) Roots Response to Salt Stress. Agronomy 2021, 11, 2389. https://doi.org/10.3390/agronomy11122389
Bouhouch S, Eshelli M, Slama HB, Bouket AC, Oszako T, Okorski A, Rateb ME, Belbahri L. Morphological, Biochemical, and Metabolomic Strategies of the Date Palm (Phoenix dactylifera L., cv. Deglet Nour) Roots Response to Salt Stress. Agronomy. 2021; 11(12):2389. https://doi.org/10.3390/agronomy11122389
Chicago/Turabian StyleBouhouch, Safa, Manal Eshelli, Houda Ben Slama, Ali Chenari Bouket, Tomasz Oszako, Adam Okorski, Mostafa E. Rateb, and Lassaad Belbahri. 2021. "Morphological, Biochemical, and Metabolomic Strategies of the Date Palm (Phoenix dactylifera L., cv. Deglet Nour) Roots Response to Salt Stress" Agronomy 11, no. 12: 2389. https://doi.org/10.3390/agronomy11122389
APA StyleBouhouch, S., Eshelli, M., Slama, H. B., Bouket, A. C., Oszako, T., Okorski, A., Rateb, M. E., & Belbahri, L. (2021). Morphological, Biochemical, and Metabolomic Strategies of the Date Palm (Phoenix dactylifera L., cv. Deglet Nour) Roots Response to Salt Stress. Agronomy, 11(12), 2389. https://doi.org/10.3390/agronomy11122389